

Abstract
This article reviews the role of protein intake on metabolic programming early in life. The observations that breastfeeding in infancy reduces the risk of being overweight and obese later in life and the differences in the protein content between formula milk and human milk have generated the early protein hypothesis. The present review focuses on a mechanistic approach to programmed adiposity and the growth and development of other organs by protein intake in infancy, which may be mediated by branched-chain amino acids, insulin, and insulin-like growth factor 1 via the mammalian target of rapamycin. Observational studies and clinical trials have shown that lowering the protein content in infant and follow-on formulas may reduce the risk of becoming obese later in life. The recent body of evidence is currently being translated into new policies. Therefore, the evolution of European regulatory laws and recommendations by expert panels on the protein content of infant and follow-on formulas are also reviewed. Research gaps, such as the critical window for programming adiposity by protein intake, testing formulas with modified amino acids, and the long-term consequences of differences in protein intake on organ functionality among well-nourished infants, have been identified.
Keywords: programming, infant nutrition, protein, adiposity
Introduction
The increased prevalence rates of noncommunicable diseases, such as obesity, have been a matter of public health and of economic burden in developed countries in the recent decades.1,2
The difficulty and cost associated with treating obesity and related disorders have increased the interest in early prevention. In this context, the study of early metabolic programming has gained importance. The concept of early metabolic programming, ie, a “nutritional stimulus or insult during critical periods of development may affect health later in life,”3 has been widely demonstrated in animal models and human studies.
Rapid Early Growth Is Associated With Later Obesity Risk
One of the most important factors that predict being overweight in childhood and adulthood is early growth. A systematic review and meta-analyses published in 2013 by Weng et al concluded that early rapid weight gain is an early identifiable risk factor that increases the likelihood of being overweight and obese in childhood.4 The UK Millennium Cohort Study found that rapid weight gain during the first year of life resulted in a fourfold increased risk of being overweight or obese at three years (defined as weight for age z-score >0.67 standard deviations [SD]).5 Furthermore, a meta-analysis conducted on 10 cohort studies in 20126 concluded that each weight gain z-score from birth to one year increased the risk of being overweight or obese in adulthood. In other words, modulating weight gain during the first year of life may be a tool to prevent obesity.
Basis for the Metabolic Programming of Adiposity by Protein Intake in Infancy
In the 1900s, King stated that nothing is comparable to breast milk, the birthright of the newborn.7 However, in the 1930s, breast milk was considered to be simply food and started to be frequently replaced by infant formula.8 Since then, pediatricians have required eight decades to realize that milk and infant formula cannot be handled as simply food; rather, they are a mammalian programming system of great importance for long-term human health. Currently, breastfeeding is known as a lactation genome-controlled physiological method of infant feeding of mammals.9–11
Based on systematic reviews and meta-analyses, breastfeeding is well known to reduce obesity risk. A meta-analyses performed by Arenz et al12 showed that breastfeeding resulted in a 22% reduction in childhood obesity. More recently, in 2012, a meta-analysis published by Weng et al4 showed that having been breast-fed (independent of duration) resulted in a mean reduction in the obesity risk of 15% compared with never having been breast-fed. In 1999, an observational study had already shown a dose-dependent association between breastfeeding and the prevention of being overweight and obesity; children who had been breast-fed for <2 months had an odds ratio (OR) of being overweight at the school age of 0.85 compared with children who had never been breast-fed, whereas infants breast-fed for >12 months had an OR of <0.5.13
Several factors may influence the protective effect of breastfeeding, such as the presence of hormones and adipokines in human milk, which may contribute to the self-regulation of an infant’s appetite,14 the parental pressure to feed the infant with formula, and the lower protein supply in human milk compared with formula milk.15 Protein intake has been associated with increase in the body mass index (BMI) in children based on observational studies conducted in different countries and at different ages. The DONALD study found that children who had a high-protein intake at 18–24 months had an increased risk of being overweight and overfat at the age of seven years.16 Similarly, Scaglioni et al17 reported a significant association between protein intake at one year of age and the risk of being overweight at five years. Among others, these observations have led to the development of the so-called early protein hypothesis.
Early Protein Hypothesis
The early protein hypothesis suggests that the protective effect of breastfeeding might be due (at least in part) to the lower protein content in human milk compared with cow’s milk-based formulas. A high-protein intake in early childhood may increase the levels of insulin-releasing amino acids (branched-chain amino acids), which may in turn stimulate insulin and insulinlike growth factor 1 (IGF-1) secretion, which are all upstream of the mammalian target of rapamycin (mTOR) growth signaling network. Then, these factors may stimulate growth and adipogenic activity (Fig. 1).18 The rationale to link increased early protein intake with not only higher weight gain but also obesity risk later in life relies on the fact that faster weight gain in infancy is associated with increased adipo genesis and later obesity risk (as mentioned in the Rapid Early Growth Is Associated With Later Obesity Risk section). Therefore, the stimulation of weight gain by protein intake early in life would translate into an increased later obesity risk.
Figure 1.

The early protein hypothesis (adapted from Koletzko et al).18 High-protein intake in early childhood may increase the levels of insulin-releasing amino acids, which may consequently stimulate insulin and IGF-1 secretion and stimulate growth and adipogenic activity to increase the long-term risk of obesity and associated disorders.
The EU Childhood Obesity Project (EU CHOP) is a randomized clinical trial designed to demonstrate the early protein hypothesis. Between 2002 and 2004, >1670 infants were recruited in five European countries (Germany, Belgium, Italy, Poland, and Spain) during the first two months of life. Formula-fed infants were randomized to be fed either a lower (1.77 g and 2.2 g protein/100 kcal) or a higher (2.9 g and 4.4 g protein/100 kcal, respectively) protein infant and follow-on formula, respectively (the lowest and highest ranges of the recommendations at the time of the study)19 during the first year of life.20 Breastfed infants served as a gold standard reference of growth and development. At two years, infants fed the lower protein content formula had a significantly lower BMI and weight per length, whereas the body length did not differ from that of infants fed the higher protein content formula.20
These results were partially supported by a shorter, recently published clinical trial, in which feeding infants formula with a reduced protein content starting at three months of age decelerated weight gain from 4 to 12 months compared with feeding those with a formula containing higher levels of protein.21
In addition, the EU CHOP was able to demonstrate the different steps of the mechanism proposed by the early protein hypothesis. Infants fed the higher protein formula exhibited increased blood levels of branched-chain amino acids, IGF-1, and faster weight gain at six months of life, and weight gain was associated with increased fat mass but not increased lean mass at six months.22 Children from the EU CHOP were followed up until the age of six years, when BMI and obesity risk were reduced among children who had been fed the lower protein content formula compared with the higher protein group (5% vs. 10%, respectively, obesity prevalence rate).23
Imbalanced mTORC1 Signaling Due to High-protein Formula Feeding
The target of rapamycin (TOR) is a highly conserved protein kinase24 that was first found to confer resistance to the growth inhibitory properties of rapamycin in the yeast Saccharomyces cerevisiae.25 This inhibition was later confirmed in all eukaryote organisms, including mammals.26 The mTOR is the central coordinator (checkpoint) of two signaling complexes, mTORC1 and mTORC2.27 Upstream and downstream mTORC1 signaling inputs determine the mass (size) of the cells and respond to nutrients, such as essential branched-chain amino acids (predominantly leucine, valine, and isoleucine), glutamine, and growth factors, such as insulin and IGF-1. mTORC2 determines the shape of the cell26 and is also activated by insulin and IGF-1, but not by amino acids.27 mTOR has been shown to regulate cell growth and proliferation in different tissues, such as the hypothalamus, liver, pancreas, muscle, and adipose tissue.27,28
Mammalian milk has been shown to be a species-specific endocrine signaling system that promotes growth upstream of the mTORC1 signaling system.11,29 The essential amino acids present in mammalian milk exert an insulinotropic effect, stimulating insulin synthesis and secretion.30
Insulin synthesis and secretion have been shown to be stimulated in infants fed formula containing high levels of protein, as indicated by increased urinary C-peptide levels compared with infants fed a low-protein formula31,32 and with breastfed infants.31 Furthermore, in infants fed formula containing higher levels of protein, the IGF-1 axis has been shown to be stimulated by higher serum concentrations of total and free IGF-1 and IGF-1-binding protein 3 as well as lower levels of IGF-1-binding protein 2, which were again compared with infants fed low-protein formula and with breastfed infants.31,33 These findings were independent of genetic variants.34 Amino acids, insulin, and IGF-1 converge at the same pathway to activate mTORC1, which promotes adipogenesis in adipose tissue by activating PPAR-γ.28
In 2012, Melnik presented a missing mechanistic link between the early protein hypothesis and mTORC1-mediated early childhood obesity. In this review, the author proposed that leucine in cow’s milk protein plays a key role in inducing early childhood obesity. Leucine is a precursor of fatty acid and cholesterol synthesis35 and is the main amino acid that activates mTORC1.36 In addition, Melnik reported the results that linked dietary leucine intake and serum levels to different mammalian milks. Thus, the leucine contents in an infant’s diet and an infant’s leucine serum levels are lower when they are fed human milk and higher when fed cow protein-based formulas and cow’s milk, and these changes are proportional to the total protein content. Accordingly, in the CHOP study, infants who were breast-fed had significantly lower blood levels of leucine and other branched-chain amino acids than infants who were fed with the lower and higher cow’s protein formulas.31 In this scenario, Melnik proposed that adipogenesis due to mTORC1 signaling was excessive in infants fed leucine-rich formula milk compared with those fed human milk (Fig. 2).
Figure 2.

Metabolic mechanism of human milk-mediated mTORC1 signaling and increased adipogenic activity by feeding leucine-rich infant formula (Adapted from Melnik).42
In summary, cow’s milk-based high-protein formula may stimulate adipogenesis in newborns via the joint action of insulin and IGF-1 (as growth factors), leucine (as essential substrate similar to building blocks), and energy, and these effects are all orchestrated by mTOR.
In parallel to the peripheral mechanism linking mTORC1 signaling in adipose tissue and obesity, hypothalamic mTOR has been associated with the regulation of food intake and energy balance. mTORC1 has been proposed to play an important role in the regulation of food intake in the hypothalamus by reducing the expression of orexigenic peptides (NPY and AgRP),28 and mTORC2 signaling may be associated with the regulation of energy balance.37 Thus, obesity may be driven by the stimulation of mTORC, both peripherally and in the central nervous system.
Critical Periods for Programming Adiposity by Protein Intakes
Prepregnancy, gestation, and early infancy have been defined as critical windows of programming,38 but evidence indicating how long an infant can be programmed is lacking. Because the growth and development of tissues is fastest during gestation and the first year of life, this period may be the critical window for programming. Alternatively, protein intake during the second year of life may impact the development of obesity later in life.
The EU CHOP is the only published clinical trial to demonstrate the early protein hypothesis, and it only covered the first year of life. However, other observational studies have shown similar associations between protein intake and obesity risk in older children. In 2000, an abovementioned work published by Scaglioni et al suggested that the risk of being overweight at age five was increased by higher protein intake at age two.17 One of the other longitudinal observational studies that had reported an association between protein intake and later obesity risk was the DONALD study. Interestingly, this study reported that infants who had higher protein intake at 18 and 24 months (but not earlier) were at a greater risk of being overweight (OR = 2.4) and overfatness (OR = 2.3) at age seven.39
These observations have not yet been tested in a well-designed clinical trial, but they lay a foundation for the study of preventive nutritional interventions.
Protein Sources and the Programming of Body Composition
The effect of a protein source on the activation of the IGF-1 axis and, consequently, adiposity is another important research question.
Studies conducted in Denmark have shown that animal protein, but not vegetable protein, is associated with the stimulation of the IGF-1 axis.40 Specifically, milk protein but not meat protein increased the concentrations of IGF-1 and IGFBP-3 in 2.5-year-old children. They quantified that an increase in milk intake from 200 to 600 mL/d was associated with a 30% increase in IGF-1. Similar associations were found in eight-year-old children.41 Furthermore, animal protein intake in 12-month-old children was associated with a higher proportion of body fat later in childhood (at seven years of age), and only dairy products (but not meat or cereals) were associated with body fat.16 According to these data and the rationale by Melnik,42 milk and dairy products may be the primary protein sources that can stimulate IGF-1 secretion and jointly (amino acids and IGF-1) stimulate mTORC1 to activate adipogenesis. The level to which different milk sources and their products (cow’s milk and goat’s milk) are able to stimulate adipogenesis requires further study.
Can Protein Intake in Infancy Affect Other Tissues?
In critical periods of growth and development, organs are being formed and grow rapidly. Therefore, in addition to adipose tissue, other tissues and systems in the body are affected by protein supply variations.
The critical windows during which specific tissues might be affected depend on the target tissue or organ. For example, in the human heart and brain, new cells are formed not only during gestation but also postnatally.43,44 However, this process differs in the kidney. Although animal models show that nephron endowment occurs pre- and postnatally, the formation of new cells in humans only occurs in utero until the third trimester of gestation. Thus, human kidney cells can only grow in size postnatally (hypertrophy), but not in number (hyperplasia).45
Brain
Studies of undernourished populations have consistently shown that protein supplementation early in life improves brain functioning, leading to children and adults with better mental performance.46 Similarly, studies of preterm infants, a population vulnerable to nutritional adverse environments, have been conducted. A clinical trial of 360 preterm infants demonstrated that infants who were fed a protein- and energy-supplemented formula had improved intelligence quotients in childhood (7.5–8 years of age)47 compared with children who had been fed a standard formula.
The EU CHOP studied the effect of variations in protein supply within the normal ranges on the brain development of healthy infants during the first year of life. Children (eight years of age) were subjected to a comprehensive battery of neuropsychological tests, including several measures of memory, attention, and executive functions,48 but the results of these tests did not differ between children fed high- and low-protein formulas.
Renal system
Animal models have shown that protein restriction during critical periods, such as pregnancy, produces offspring with smaller kidneys that feature fewer nephrons and reduced renal function later in life.49,50 In infants, being formula fed (compared to breastfed)51 or being fed a higher protein content formula (compared with the lower protein content formula)52 resulted in larger kidneys and increases in function markers. The reversibility of this protein-induced kidney growth and changes in kidney function early in life is unknown. Results from an observational study51 showed an increase in kidney size among formula-fed infants at three months compared with breastfed infants. At 18 months, these differences in kidney size due to feeding early in infancy had disappeared, suggesting that protein-induced kidney growth is reversible. Similarly, the results from the Generation R cohort study recently showed that protein intake early in life is not associated with later kidney size and function at age six.53 However, the protein intake of children was assessed at age one, and this level does not necessarily reflect the protein intake during the first six months of life, when differences by food type (breast vs. formula milk) may be larger. Protein intake may increase kidney size via three physiological mechanisms: first, kidneys increase in volume to support an increased need to excrete nitrogen metabolites. Kidney growth due to this increased workload would be reversible. Second, higher protein intake stimulates IGF-1 secretion, which stimulates kidney growth and body weight gain. Third, total body weight gain may indirectly promote the growth of organs to support a higher metabolic work load.54 The authors from the EU CHOP study hypothesized that the programming mechanism, if any, might be mediated by IGF-1 and its systemic effect on the entire body.54
Heart and blood pressure
Some evidence has suggested that protein intake early in life may change cardiac function and structure. For example, fetal exposure to a maternal low-protein diet has been associated with altered left ventricular function.55 In infancy and childhood, protein–energy malnutrition has been associated with reduced left ventricular mass and functioning.56–58 On the contrary, animal studies have shown that overnutrition early in life is associated with ventricular hypertrophy.59 However, data supporting that modulating protein supply affects heart size and function in well-nourished populations have not been published.
From a mechanistic point of view and in accordance with the early protein hypothesis, the protein-induced growth of the brain and heart is likely mediated by IGF-1 (as is the case in the kidney) because IGF-1 has been shown to mediate cardiac hypertrophy in acromegaly60 and brain growth in Laron syndrome.61
Furthermore, the activation of mTORC1 signaling has been associated with the sympathetic nervous system and cardiovascular function. Thus, mTORC1 signaling in the hypothalamus due to leucine increases blood flow and arterial pressure62 and may also drive cardiac hypertrophy.
Immune system
Notably, accelerated growth in infancy has been linked to allergy development. The BMI in childhood has been associated with the development of asthma,63 but the infantile growth velocity64 and gain in BMI over time during infancy seem to be more important predictors of asthma than excess weight at any specific age.63
Asthma and obesity have been linked because their development shares a common mechanism: both pathological conditions have been associated with enhanced mTORC1 signaling in infancy, encompassing accelerated postnatal growth.65 Enhanced mTORC1 signaling suppresses the development and maturation of FoxP3+ regulatory T cells in the thymus,66 which may increase the risk of allergic asthma development.
Changes in the Recommended Protein Contents in Infant and Follow-on Formulas
Historically, formula-fed infants have received more protein than breastfed infants.15
Given the recent body of evidence, efforts are being made to decrease the protein content of infant and follow-on formulas. The European Directive from 1991 established a minimum of 1.8 g and 2.25 g protein/100 Kcal and a maximum of 3 g and 4.5 g protein/100 kcal for infant and follow-on formulas, respectively.19 In 2003, a report by the Scientific Committee on Food of the European Commission on the revision of essential requirements of infant formulae and follow-on formulae concluded that the minimum protein level in cow’s milk formulas is the same for infant and follow-on formulas (1.8 g protein/100 kcal) and that the maximum range for follow-on formulas should be reduced from 4.5 g to 3 g protein/100 kcal.67 The European Directive from 2006 partially adopted the recommended decreases in the minimum and maximum protein content of follow-on formulas of 1.8 g and 3.5 g/100 kcal, respectively.68 More recently, the European Food Safety Authority (EFSA) published a scientific opinion recommending a maximum protein level in both cow’s milk infant and follow-on formulas of 2.5 g protein/100 kcal.69 Recent recommendations and regulatory laws are provided in Figure 3.
Figure 3.
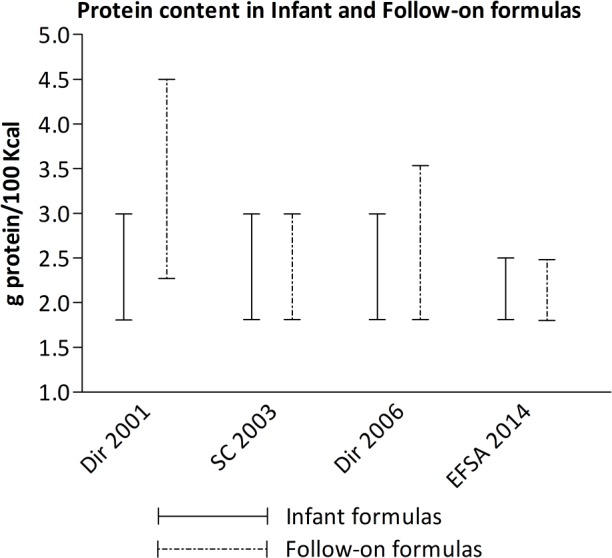
European directives and recommendations on the cow’s milk protein content of infant and follow-on formulas. Dir 2001 Commission Directive 91/321/EEC of May 14, 1991, on infant and follow-on formulae; SC 2003: Report of the Scientific Committee on Food on the revision of essential requirements of infant formulae and follow-on formulae (SCF/CS/NUT/IF/65 Final); Dir 2006 Commission Directive 2006/141/EC of December 22, 2006, on infant formulae and follow-on formulae and amending Directive 1999/21/EC; EFSA 2014 Scientific Opinion on the essential composition of infant and follow-on formulae.
The minimum levels of 1.8 g/100 kcal in infant formulas and 2.2 g/100 kcal in follow-on formulas from the EU CHOP study were shown to be safe in terms of longitudinal growth20 and long-term neurodevelopment.48 However, considering the protein supply in human milk, which is being reduced during the first year of life, these minimum ranges may still exceed nutritional needs starting at age three months.70 In a recently published study, Ziegler et al showed that an infant formula containing protein levels below the regulatory lower limit (1.61 g protein/dl) and a formula with a higher protein content provided similar longitudinal growth rates.21
Conversely, commercially available formulas for young children aged one to three (also called toddler or growing-up formula milk) are not regulated. The EFSA recently reported that these formulas cannot be considered as necessity to satisfy the nutritional requirements of infants.69 However, these formulas might be useful for reducing protein intake from dairy products in young children, and this reduction may reduce later obesity risk. To our knowledge, well-designed clinical trials have not yet demonstrated an effect of protein intake at this age on later obesity risk. Only one report from an expert panel providing recommendations for these toddler formulas proposed a range from 2.5 g to 3.75 g/100 kcal for the second year of life.71 However, consuming this level of milk protein in conjunction with family food may result in caloric excess,72,73 by providing more protein than breast milk at one year.15
In parallel to a reduction in the protein content of formula milk, the addition of bioactive protein compounds present in human milk should be tested and considered. Human milk contains many of these bioactive compounds, including α-lactalbumin, lactoferrin, osteopontin, and exosomes, and these compounds may benefit immune function, allergy prevention, and micronutrient absorption.74–76
Summary and Conclusions
Differential protein intake early in life has been shown to affect the growth and functioning of different tissues in the human body. The effects of protein intake might be driven by high leucine levels and mediated by the IGF-1 axis, which stimulates the mTOR signaling pathway.
Lowering the protein supply during the first year of life has been demonstrated to decelerate weight gain and reduce obesity risk later in childhood. This finding is remarkably important because children who are overweight or obese are likely to be overweight or obese adults.77
In parallel, differences in the protein intake during the first year of life may modulate the growth and functioning of different organs and tissues, such as brain, kidney, and possibly heart. These effects have been clearly demonstrated in vulnerable populations (such as undernourished children or preterm infants), but the permanence of these effects is not known.
Notably, recent advances in understanding the functioning of mTOR suggest that uncontrolled high-protein formula milk may increase the risk of obesity, allergy, and hypertension and these conditions are noncommunicable epidemic diseases that currently affect Western civilization.
Given these data, this review attempts to recommend and propose future scientific aims.
First, the protein content in infant formulas should be reduced; regulations guiding the industry to develop more healthy manufactured products may play a key role on obesity prevention.
On the other hand, pediatricians’ understanding that human milk is able to prevent programming of noncommunicable diseases for future generations may be crucial.
At present, formula milk that can mimic all the aspects of breast milk is not available. Therefore, policies for obesity prevention should promote and support human milk feeding.
Conversely, parents culturally provide as much protein as possible, especially after the complementary feeding introduction, and children usually prefer to eat milk-based desserts. These practices should also be addressed from an educational policy perspective. To this end, professionals should be trained to recommend specific protein and milk product portions by age to avoid overfeeding with cow’s milk.
In addition, research should focus on different open fields, testing amino acid-modified formulas (ie, reducing the leucine and branched-chain amino acid supply) and determining the critical time window of programming, because some evidence suggests that metabolic programming occurs not only during the first year of life but also during the second year. However, the long-term effects of protein-induced growth acceleration in early life on noncommunicable diseases in adulthood should also be investigated.
Moreover, the exploration of new treatment options has recently become possible due to a better understanding of the role of the mTOR pathway in programming. Therefore, understanding programming may provide a basis not only for prevention but also for treatment.
Footnotes
ACADEMIC EDITOR: Joseph Zhou, Editor in Chief
PEER REVIEW: Four peer reviewers contributed to the peer review report. Reviewers’ reports totaled 2470 words, excluding any confidential comments to the academic editor.
FUNDING: Authors disclose no external funding sources.
COMPETING INTERESTS: Authors disclose no potential conflicts of interest.
Paper subject to independent expert single blind peer review. All editorial decisions made by independent academic editor. Upon submission manuscript was subject to anti-plagiarism scanning. Prior to publication all authors have given signed confirmation of agreement to article publication and compliance with all applicable ethical and legal requirements, including the accuracy of author and contributor information, disclosure of competing interests and funding sources, compliance with ethical requirements relating to human and animal study participants, and compliance with any copyright requirements of third parties. This journal is a member of the Committee on Publication Ethics (COPE).
Provenance: the authors were invited to submit this paper.
Author Contributions
Wrote the first draft of the manuscript: VL. Contributed to the writing of the manuscript: RC, JE, NF. Agree with manuscript results and conclusions: VL, RC, JE, NF. Jointly developed the structure and arguments for the paper: VL. Made critical revisions and approved final version: RC, JE, NF. All authors reviewed and approved of the final manuscript.
REFERENCES
- 1.Wang Y, Lobstein T. Worldwide trends in childhood overweight and obesity. Int J Pediatr Obes. 2006;1(1):11–25. doi: 10.1080/17477160600586747. [DOI] [PubMed] [Google Scholar]
- 2.Wang G, Dietz WH. Economic burden of obesity in youths aged 6 to 17 years: 1979–1999. Pediatrics. 2002;109(5):E81–E81. doi: 10.1542/peds.109.5.e81. [DOI] [PubMed] [Google Scholar]
- 3.Lucas A. Programming by early nutrition: an experimental approach. J Nutr. 1998;128(2 suppl):401s–406s. doi: 10.1093/jn/128.2.401S. [DOI] [PubMed] [Google Scholar]
- 4.Weng SF, Redsell SA, Swift JA, Yang M, Glazebrook CP. Systematic review and meta-analyses of risk factors for childhood overweight identifiable during infancy. Arch Dis Child. 2012;97(12):1019–1026. doi: 10.1136/archdischild-2012-302263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Weng SF, Redsell SA, Nathan D, Swift JA, Yang M, Glazebrook C. Estimating overweight risk in childhood from predictors during infancy. Pediatrics. 2013;132(2):e414–e421. doi: 10.1542/peds.2012-3858. [DOI] [PubMed] [Google Scholar]
- 6.Druet C, Stettler N, Sharp S, et al. Prediction of childhood obesity by infancy weight gain: an individual-level meta-analysis. Paediatr Perinat Epidemiol. 2012;26(1):19–26. doi: 10.1111/j.1365-3016.2011.01213.x. [DOI] [PubMed] [Google Scholar]
- 7.King FT. Natural Feeding of Infants. London: Whitcombe & Tombs; 1918. p. 7. [Google Scholar]
- 8.Bryder L. From breast to bottle: a history of modern infant feeding. Endeavour. 2009;33(2):54–59. doi: 10.1016/j.endeavour.2009.04.008. [DOI] [PubMed] [Google Scholar]
- 9.Melnik BC, John SM, Schmitz G. Milk is not just food but most likely a genetic transfection system activating mTORC1 signaling for postnatal growth. Nutr J. 2013;12:103. doi: 10.1186/1475-2891-12-103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ward TL, Hosid S, Ioshikhes I, Altosaar I. Human milk metagenome: a functional capacity analysis. BMC Microbiol. 2013;13:116. doi: 10.1186/1471-2180-13-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Melnik BC. Milk—a nutrient system of mammalian evolution promoting mTORC1-dependent translation. Int J Mol Sci. 2015;16(8):17048–17087. doi: 10.3390/ijms160817048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Arenz S, Ruckerl R, Koletzko B, von Kries R. Breast-feeding and childhood obesity—a systematic review. Int J Obes Relat Metab Disord. 2004;28:1247–1256. doi: 10.1038/sj.ijo.0802758. [DOI] [PubMed] [Google Scholar]
- 13.von Kries R, Koletzko B, Sauerwald T, et al. Breast feeding and obesity: cross sectional study. BMJ. 1999;319:147–150. doi: 10.1136/bmj.319.7203.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.DiSantis KI, Collins BN, Fisher JO, Davey A. Do infants fed directly from the breast have improved appetite regulation and slower growth during early childhood compared with infants fed from a bottle? Int J Behav Nutr Phys Act. 2011;8:89. doi: 10.1186/1479-5868-8-89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Heinig MJ, Nommsen LA, Peerson JM, Lonnerdal B, Dewey KG. Energy and protein intakes of breast-fed and formula-fed infants during the first year of life and their association with growth velocity: the DARLING study. Am J Clin Nutr. 1993;58(2):152–161. doi: 10.1093/ajcn/58.2.152. [DOI] [PubMed] [Google Scholar]
- 16.Gunther ALB, Remer T, Kroke A, Buyken AE. Early protein intake and later obesity risk: which protein sources at which time points throughout infancy and childhood are important for body mass index and body fat percentage at 7 y of age? Am J Clin Nutr. 2007;86(6):1765–1772. doi: 10.1093/ajcn/86.5.1765. [DOI] [PubMed] [Google Scholar]
- 17.Scaglioni S, Agostoni C, Notaris RD, et al. Early macronutrient intake and overweight at five years of age. Int J Obes Relat Metab Disord. 2000;24(6):777–781. doi: 10.1038/sj.ijo.0801225. [DOI] [PubMed] [Google Scholar]
- 18.Koletzko B, von Kries R, Closa R, et al. Can infant feeding choices modulate later obesity risk? Am J Clin Nutr. 2009;89(5):1502S–1508S. doi: 10.3945/ajcn.2009.27113D. [DOI] [PubMed] [Google Scholar]
- 19.Commission E. Commission directive 91/321/EEC of 14 May 1991 on infant and follow-on formulae. Off J Eur Comm. 1991;34:35–49. [Google Scholar]
- 20.Koletzko B, von Kries R, Closa R, et al. European Childhood Obesity Trial Study Group Lower protein in infant formula is associated with lower weight up to age 2 y: a randomized clinical trial. Am J Clin Nutr. 2009;89:1836–1845. doi: 10.3945/ajcn.2008.27091. [DOI] [PubMed] [Google Scholar]
- 21.Ziegler EE, Fields DA, Chernausek SD, et al. Adequacy of infant formula with protein content of 1.6 g/100 kcal for infants between 3 and 12 months: a randomized multicenter trial. J Pediatr Gastroenterol Nutr. 2015;61(5):596–603. doi: 10.1097/MPG.0000000000000881. [DOI] [PubMed] [Google Scholar]
- 22.Escribano J, Luque V, Ferre N, et al. European Childhood Obesity Trial Study Group Effect of protein intake and weight gain velocity on body fat mass at 6 months of age: the EU childhood obesity programme. Int J Obes. 2012;36:548–553. doi: 10.1038/ijo.2011.276. [DOI] [PubMed] [Google Scholar]
- 23.Weber M, Grote V, Closa-Monasterolo R, et al. European Childhood Obesity Trial Study Group Lower protein content in infant formula reduces BMI and obesity risk at school age: follow-up of a randomized trial. Am J Clin Nutr. 2014;99(5):1041–1051. doi: 10.3945/ajcn.113.064071. [DOI] [PubMed] [Google Scholar]
- 24.Loewith R, Hall MN. Target of rapamycin (TOR) in nutrient signaling and growth control. Genetics. 2011;189(4):1177–1201. doi: 10.1534/genetics.111.133363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Heitman J, Movva NR, Hall MN. Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science. 1991;23:905–909. doi: 10.1126/science.1715094. [DOI] [PubMed] [Google Scholar]
- 26.Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism. Cell. 2006;124(3):471–484. doi: 10.1016/j.cell.2006.01.016. [DOI] [PubMed] [Google Scholar]
- 27.Polak P, Hall MN. mTOR and the control of whole body metabolism. Curr Opin Cell Biol. 2009;21(2):209–218. doi: 10.1016/j.ceb.2009.01.024. [DOI] [PubMed] [Google Scholar]
- 28.Laplante M, Sabatini DM. mTOR signaling in growth control and disease. Cell. 2012;149(2):274–293. doi: 10.1016/j.cell.2012.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Melnik BC. Milk signalling in the pathogenesis of type 2 diabetes. Med Hypotheses. 2011;76(4):553–559. doi: 10.1016/j.mehy.2010.12.017. [DOI] [PubMed] [Google Scholar]
- 30.Bartolome A, Guillén C. Role of the mammalian target of rapamycin (mTOR) complexes in pancreatic β-cell mass regulation. Vitam Horm. 2014;95:425–469. doi: 10.1016/B978-0-12-800174-5.00017-X. [DOI] [PubMed] [Google Scholar]
- 31.Socha P, Grote V, Gruszfeld D, et al. European Childhood Obesity Trial Study Group Milk protein intake, the metabolic-endocrine response, and growth in infancy: data from a randomized clinical trial. Am J Clin Nutr. 2011;94(6 suppl):1776S–1784S. doi: 10.3945/ajcn.110.000596. [DOI] [PubMed] [Google Scholar]
- 32.Axelsson IE, Ivarsson SA, Raiha NC. Protein intake in early infancy: effects on plasma amino acid concentrations, insulin metabolism, and growth. Pediatr Res. 1989;26(6):614–617. doi: 10.1203/00006450-198912000-00020. [DOI] [PubMed] [Google Scholar]
- 33.Closa-Monasterolo R, Ferre N, Luque V, et al. Childhood Obesity Project Study Group Sex differences in the endocrine system in response to protein intake early in life. Am J Clin Nutr. 2011;94(6 suppl):1920S–1927S. doi: 10.3945/ajcn.110.001123. [DOI] [PubMed] [Google Scholar]
- 34.Rzehak P, Grote V, Lattka E, et al. European Childhood Obesity Trial Study Group Associations of IGF-1 gene variants and milk protein intake with IGF-I concentrations in infants at age 6 months—results from a randomized clinical trial. Growth Horm IGF Res. 2013;23(5):149–158. doi: 10.1016/j.ghir.2013.05.002. [DOI] [PubMed] [Google Scholar]
- 35.Rosenthal J, Angel A, Farkas J. Metabolic fate of leucine: a significant sterol precursor in adipose tissue and muscle. Am J Physiol. 1974;226(2):411–418. doi: 10.1152/ajplegacy.1974.226.2.411. [DOI] [PubMed] [Google Scholar]
- 36.Lynch CJ. Role of leucine in the regulation of mTOR by amino acids: revelations from structure-activity studies. J Nutr. 2001;131(3):861S–865S. doi: 10.1093/jn/131.3.861S. [DOI] [PubMed] [Google Scholar]
- 37.Haissaguerre M, Saucisse N, Cota D. Influence of mTOR in energy and metabolic homeostasis. Mol Cell Endocrinol. 2014;397(1–2):67–77. doi: 10.1016/j.mce.2014.07.015. [DOI] [PubMed] [Google Scholar]
- 38.Demmelmair H, von Rosen J, Koletzko B. Long-term consequences of early nutrition. Early Hum Dev. 2006;82:567–574. doi: 10.1016/j.earlhumdev.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 39.Gunther AL, Buyken AE, Kroke A. Protein intake during the period of complementary feeding and early childhood and the association with body mass index and percentage body fat at 7 y of age. Am J Clin Nutr. 2007;85(6):1626–1633. doi: 10.1093/ajcn/85.6.1626. [DOI] [PubMed] [Google Scholar]
- 40.Hoppe C, Udam TR, Lauritzen L, Molgaard C, Juul A, Michaelsen KF. Animal protein intake, serum insulin-like growth factor I, and growth in healthy 2.5-y-old Danish children. Am J Clin Nutr. 2004;80(2):447–452. doi: 10.1093/ajcn/80.2.447. [DOI] [PubMed] [Google Scholar]
- 41.Hoppe C, Molgaard C, Juul A, Michaelsen KF. High intakes of skimmed milk, but not meat, increase serum IGF-I and IGFBP-3 in eight-year-old boys. Eur J Clin Nutr. 2004;58:1211–1216. doi: 10.1038/sj.ejcn.1601948. [DOI] [PubMed] [Google Scholar]
- 42.Melnik BC. Excessive leucine-mTORC1-signalling of cow milk-based infant formula: the missing link to understand early childhood obesity. J Obes. 2012;2012:197653. doi: 10.1155/2012/197653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Andersen SL. Trajectories of brain development: point of vulnerability or window of opportunity? Neurosci Biobehav Rev. 2003;27:3–18. doi: 10.1016/s0149-7634(03)00005-8. [DOI] [PubMed] [Google Scholar]
- 44.Mollova M, Bersell K, Walsh S, et al. Cardiomyocyte proliferation contributes to heart growth in young humans. Proc Natl Acad Sci U S A. 2013;110(4):1446–1451. doi: 10.1073/pnas.1214608110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hinchliffe SA, Lynch MR, Sargent PH, Howard CV, Van Velzen D. The effect of intrauterine growth retardation on the development of renal nephrons. Br J Obstet Gynaecol. 1992;99(4):296–301. doi: 10.1111/j.1471-0528.1992.tb13726.x. [DOI] [PubMed] [Google Scholar]
- 46.Li H, Barnhart HX, Stein AD, Martorell R. Effects of early childhood supplementation on the educational achievement of women. Pediatrics. 2003;112(5):1156–1162. doi: 10.1542/peds.112.5.1156. [DOI] [PubMed] [Google Scholar]
- 47.Lucas A, Morley R, Cole TJ. Randomised trial of early diet in preterm babies and later intelligence quotient. BMJ. 1998;317(7171):1481–1487. doi: 10.1136/bmj.317.7171.1481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Escribano J, Luque V, Canals-Sans J, et al. Mental performance in 8 years-old children fed reduced protein content formula during the first year of life: safety analysis of a randomized clinical trial. Br J Nutr. 2016;22:1–9. doi: 10.1017/S0007114515000768. Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 49.Hammond KA, Janes DN. The effects of increased protein intake on kidney size and function. J Exp Biol. 1998;201(13):2081–2090. doi: 10.1242/jeb.201.13.2081. [DOI] [PubMed] [Google Scholar]
- 50.Hoppe CC, Evans RG, Bertram JF, Moritz KM. Effects of dietary protein restriction on nephron number in the mouse. Am J Physiol Regul Integr Comp Physiol. 2007;292(5):R1768–R1774. doi: 10.1152/ajpregu.00442.2006. [DOI] [PubMed] [Google Scholar]
- 51.Schmidt I, Damgaard I, Boisen K, et al. Increased kidney growth in formula-fed versus breast-fed healthy infants. Pediatr Nephrol. 2004;19(10):1137–1144. doi: 10.1007/s00467-004-1567-0. [DOI] [PubMed] [Google Scholar]
- 52.Escribano J, Luque V, Ferre N, et al. Increased protein intake augments kidney volume and function in healthy infants. Kidney Int. 2011;79(7):783–790. doi: 10.1038/ki.2010.499. [DOI] [PubMed] [Google Scholar]
- 53.Voortman T, Bakker H, Sedaghat S, et al. Protein intake in infancy and kidney size and function at the age of 6 years: the Generation R Study. Pediatr Nephrol. 2015;30(10):1825–1833. doi: 10.1007/s00467-015-3096-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Luque V, Escribano J, Grote V, et al. European Childhood Obesity Project Does insulin-like growth factor-1 mediate protein-induced kidney growth in infants? A secondary analysis from a randomised controlled trial. Pediatr Res. 2013;74(2):223–229. doi: 10.1038/pr.2013.87. [DOI] [PubMed] [Google Scholar]
- 55.Elmes MJ, Gardner DS, Langley-Evans SC. Fetal exposure to a maternal low-protein diet is associated with altered left ventricular pressure response to ischaemia-reperfusion injury. Br J Nutr. 2007;98(1):93–100. doi: 10.1017/S000711450769182X. [DOI] [PubMed] [Google Scholar]
- 56.Kothari S, Patel T, Shetalwad A, Patel T. Left ventricular mass and function in children with severe protein energy malnutrition. Int J Cardiol. 1992;35(1):19–25. doi: 10.1016/0167-5273(92)90050-d. [DOI] [PubMed] [Google Scholar]
- 57.Ocal B, Unal S, Zorlu P, Tezic HT, Oguz D. Echocardiographic evaluation of cardiac functions and left ventricular mass in children with malnutrition. J Paediatr Child Health. 2001;37(1):14–17. doi: 10.1046/j.1440-1754.2001.00566.x. [DOI] [PubMed] [Google Scholar]
- 58.Faddan NH, El Sayh K, Shams H, Badrawy H. Myocardial dysfunction in malnourished children. Ann Pediatr Cardiol. 2010;3(2):113–118. doi: 10.4103/0974-2069.74036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Moreira ASB, Teixeira Teixeira M, Silveira Osso FD, et al. Left ventricular hypertrophy induced by overnutrition early in life. Nutr Metab Cardiovasc Dis. 2009;19(11):805–810. doi: 10.1016/j.numecd.2009.01.008. [DOI] [PubMed] [Google Scholar]
- 60.Maison P, Tropeano AI, Quin-Mavier I, Giustina A, Chanson P. Impact of somatostatin analogs on the heart in acromegaly: a metaanalysis. J Clin Endocrinol Metab. 2007;92(5):1743–1747. doi: 10.1210/jc.2006-2547. [DOI] [PubMed] [Google Scholar]
- 61.Laron Z, Iluz M, Kauli R. Head circumference in untreated and IGF-I treated patients with Laron syndrome: comparison with untreated and hGH-treated children with isolated growth hormone deficiency. Growth Horm IGF Res. 2012;22(2):49–52. doi: 10.1016/j.ghir.2012.02.005. [DOI] [PubMed] [Google Scholar]
- 62.Harlan SM, Guo D-F, Morgan DA, Fernandes-Santos C, Rahmouni K. Hypothalamic mTORC1 signaling controls sympathetic nerve activity and arterial pressure and mediates leptin effects. Cell Metab. 2013;17(4):599–606. doi: 10.1016/j.cmet.2013.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Brüske I, Flexeder C, Heinrich J. Body mass index and the incidence of asthma in children. Curr Opin Allergy Clin Immunol. 2014;14(2):155–160. doi: 10.1097/ACI.0000000000000035. [DOI] [PubMed] [Google Scholar]
- 64.Flexeder C, Thiering E, Brüske I, et al. GINIplus and LISAplus Study Group Growth velocity during infancy and onset of asthma in school-aged children. Allergy. 2012;67(2):257–264. doi: 10.1111/j.1398-9995.2011.02748.x. [DOI] [PubMed] [Google Scholar]
- 65.Melnik BC. The potential mechanistic link between allergy and obesity development and infant formula feeding. Allergy Asthma Clin Immunol. 2014;10(1):37. doi: 10.1186/1710-1492-10-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Haxhinasto S, Mathis D, Benoist C. The AKT-mTOR axis regulates de novo differentiation of CD4+Foxp3+ cells. J Exp Med. 2008;205(3):565–574. doi: 10.1084/jem.20071477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Food SC. Report of the Scientific Committee on Food on the Revision of Essential Requirements of Infant Formulae and Follow-On Formulae (SCF/CS/NUT/IF/65 Final) Brussels: European Commission; 2003. [Google Scholar]
- 68.Commission E. Commission directive 2006/141/EC of 22 December 2006 on infant formulae and follow-on formulae and amending directive 1999/21/EC. Off J Eur Union. 2006;49:1–33. [Google Scholar]
- 69.EFSA Panel on Dietetic Products N and A (NDA) Scientific opinion on the essential composition of infant and follow-on formulae. EFSA J. 2014;12(7):3760. [Google Scholar]
- 70.Fomon SJ. Requirements and recommended dietary intakes of protein during infancy. Pediatr Res. 1991;30(5):391–395. doi: 10.1203/00006450-199111000-00001. [DOI] [PubMed] [Google Scholar]
- 71.Lippman H, Desjeux J-F, Ding Z-Y, et al. Nutrient recommendations for growing-up milk: a report of an expert panel. Crit Rev Food Sci Nutr. 2016;56(1):141–145. doi: 10.1080/10408398.2012.702702. [DOI] [PubMed] [Google Scholar]
- 72.Michaelsen KF, Larnkjaer A, Molgaard C. Amount and quality of dietary proteins during the first two years of life in relation to NCD risk in adulthood. Nutr Metab Cardiovasc Dis. 2012;22(10):781–786. doi: 10.1016/j.numecd.2012.03.014. [DOI] [PubMed] [Google Scholar]
- 73.Michaelsen KF, Greer FR. Protein needs early in life and long-term health. Am J Clin Nutr. 2014;99(3):718S–722S. doi: 10.3945/ajcn.113.072603. [DOI] [PubMed] [Google Scholar]
- 74.Lönnerdal B. Infant formula and infant nutrition: bioactive proteins of human milk and implications for composition of infant formulas. Am J Clin Nutr. 2014;99(3):712S–717S. doi: 10.3945/ajcn.113.071993. [DOI] [PubMed] [Google Scholar]
- 75.Admyre C, Johansson SM, Qazi KR, et al. Exosomes with immune modulatory features are present in human breast milk. J Immunol. 2007;179(3):1969–1978. doi: 10.4049/jimmunol.179.3.1969. [DOI] [PubMed] [Google Scholar]
- 76.Melnik BC, John SM, Schmitz G. Milk: an exosomal microRNA transmitter promoting thymic regulatory T cell maturation preventing the development of atopy? J Transl Med. 2014;12(1):43. doi: 10.1186/1479-5876-12-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Cunningham SA, Kramer MR, Narayan KM. Incidence of childhood obesity in the United States. N Engl J Med. 2014;370(17):1660–1661. doi: 10.1056/NEJMc1402397. [DOI] [PMC free article] [PubMed] [Google Scholar]