

ABSTRACT
Enteropathogenic Escherichia coli (EPEC) remains a significant cause of infant diarrheal illness and associated morbidity and mortality in developing countries. EPEC strains are characterized by their ability to colonize the small intestines of their hosts by a multistep program involving initial loose attachment to intestinal epithelial cells followed by an intimate adhesion phase. The initial loose interaction of typical EPEC with host intestinal cells is mediated by bundle-forming pili (BFP). BFP are type 4b pili (T4bP) based on structural and functional properties shared with T4bP expressed by other bacteria. The major structural subunit of BFP is called bundlin, a T4b pilin expressed from the bfpA gene in the BFP operon, which contains three additional genes that encode the pilin-like proteins BfpI, BfpJ, and BfpK. In this study, we show that, in the absence of the BFP retraction ATPase (BfpF), BfpI, BfpJ, and BfpK are dispensable for BFP biogenesis. We also demonstrate that these three minor pilins are incorporated along with bundlin into the BFP filament and contribute to its structural integrity and host cell adhesive properties. The results confirm that previous findings in T4aP systems can be extended to a model T4bP such as BFP.
IMPORTANCE Bundle-forming pili contribute to the host colonization strategy of enteropathogenic Escherichia coli. The studies described here investigate the role for three minor pilin subunits in the structure and function of BFP in EPEC. The studies also suggest that these subunits could be antigens for vaccine development.
INTRODUCTION
The main virulence feature of the enteropathogenic Escherichia coli (EPEC) group of enterovirulent E. coli is their ability to produce attaching-and-effacing (A/E) lesions on the epithelial lining of the human small intestine (1). These lesions result from the rearrangement of the actin cytoskeleton, leading to the loss of microvilli and, subsequently, intimate attachment of the organisms to host intestinal epithelial cells (2). Typical EPEC strains differ from the atypical strains by their expression of bundle-forming pili (BFP), which are classified as type 4 pili (T4P). T4P are thin (50 to 80 Å in diameter), flexible filaments that have been classified into the groups T4aP and T4bP depending on the structure of their major pilin subunits. The BPF expressed by EPEC are members of the T4bP group, and the main pilin subunit is called bundlin (3). BFP assist the A/E process by promoting the formation of microcolonies and the early, nonintimate attachment of the organisms to host cells. This process is referred to as localized adherence (LA) (4, 5).
One functional feature of the T4aP group is their ability to extend from, and retract back into, the cell, a process energized by ATP hydrolysis (6, 7). In addition to participating in the early nonintimate phase of EPEC adherence to host intestinal cells, T4aP have been implicated in such processes as immune evasion, twitching motility, biofilm development, autoaggregation, and phage attachment (8–14). T4aP and T4bP pilins are synthesized as prepilin proteins, which are processed by a specific prepilin peptidase in the cytoplasmic membrane. All T4aP and T4bP pilins have a hydrophobic N-terminal segment (6 to 7 and 10 to 30 residues in length, respectively), and many also possess a methylated N-terminal amino acid residue (15). Despite the sequence variation found in the C-terminal regions of T4aP and T4bP pilins, many possess a pair of conserved cysteine residues (16) that are oxidized to form a loop structure which contributes to pilus stability and function (17).
In EPEC, the BFP-encoding genes reside in an operon on the EPEC adherence factor (EAF) plasmid. In addition to bfpA, the gene that encodes bundlin (18), there are three additional genes (bfpI, bfpJ, and bfpK) which encode pilin-like proteins that share many structural features with bundlin, including the prepilin peptidase cleavage site (19), the hydrophobic N-terminal sequence, and the conserved cysteines in their C-terminal domains. The ATPase enzymes responsible for the BFP extension and retraction processes are encoded by bfpD and bfpF, respectively. Moreover, although bfpD is absolutely required for BFP expression, the bfpF gene is not, and retraction-defective bfpF deletion mutants display a hyperadherent phenotype relative to that of the wild-type (WT) parental EPEC strain (20).
The role of BfpI, BfpJ, and BfpK in BFP structure and function still is poorly defined, because single-deletion mutants lacking any of the bfpI, bfpJ, and bfpK genes do not express BFP (19). As a result, these mutants do not exhibit a phenotype that can be attributed to any one of the pilin-like subunits (19). However, in the present study we determined that EPEC pilin-like deletion mutants, like those produced in Pseudomonas aeruginosa (21), Neisseria gonorrhoeae (22), and Neisseria meningitidis (23), do express BFP if they are created in a bfpF retraction-ATPase-deficient background strain. This study used this finding to describe the role of BfpI, BfpJ, and BfpK in BFP structure and function, and the results are presented here.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions.
The bacterial strains, plasmids, and PCR primers used in these investigations are described in Tables 1 and 2. Bacteria were routinely grown overnight in Luria broth (LB) or tryptic soy broth (TSB) without dextrose at 37°C from single colonies picked from an overnight LB agar plate. Ampicillin (100 μg/ml), kanamycin (Km; 50 μg/ml), tetracycline (12.5 μg/ml), or chloramphenicol (25 μg/ml) was added to the media when required for mutant selection purposes. The complemented plasmids for EPEC mutants were induced using 1 mM isopropyl-β-d-thiogalactopyranoside (IPTG). Bacteria also were grown in Dulbecco's modified Eagle's medium (DMEM) (Invitrogen) supplemented with 44 mM NaHCO3 and 25 mM glucose at 37°C or on tryptic soy agar (TSA) agar plates supplemented with 5% (vol/vol) defibrinated sheep blood (blood agar plates [BAP]) to induce BFP expression (3). All DNA procedures were performed using standard genetic and molecular techniques (29). Plasmid DNA and RNA were purified using Qiagen kits (Qiagen Inc.). Restriction and other DNA-modifying enzymes were obtained from New England BioLabs Inc. and were used according to the supplier's instructions.
TABLE 1.
Strains and plasmids used in this study
| Strain or plasmid | Description, genotype, or sequence | Reference or source |
|---|---|---|
| Strains | ||
| E2348/69 | Wild-type EPEC strain, serotype O127:H6 | 24 |
| UMD946 | E2348/69 ΔbfpF | 25 |
| UMD951 | E2348/69 ΔbfpI | This study |
| UMD957 | E2348/69 ΔbfpK | This study |
| CMP-J | E2348/69 ΔbfpJ | This study |
| UMD963 | E2348/69 bfpF ΔbfpI | This study |
| JPN15 | E2348/69 cured of EAF plasmid | 26 |
| BL21(DE3) pLysS | F− dcm ompT hsdS(rB− mB−) | Novagen |
| CMP-1 | E2348/69 ΔbfpF ΔbfpI | This study |
| CMP-2 | E2348/69 ΔbfpF ΔbfpJ | This study |
| CMP-3 | E2348/69 ΔbfpF ΔbfpK | This study |
| CMP-4 | E2348/69 ΔbfpIJK | This study |
| CMP-5 | E2348/69 ΔbfpF ΔbfpIJK | This study |
| Plasmids | ||
| pET48b+ | T7 promoter, His tag, N-terminus Trx tag | Novagen |
| pQE31 | Vector for expressing N-terminally 6× His-tagged proteins | Qiagen |
| pMPM-T3 | Low-copy-number cloning vector; p15A derivative; Tcr | 27 |
| pKD46 | λ Red recombinase system plasmid | 28 |
| pKD4 | Km cassette template plasmid | 28 |
| pETBfpI | bfpI cloned in pET48b+ | This study |
| pETBfpJ | bfpJ cloned in pET48b+ | This study |
| pQEBfpK | bfpK cloned in pQE31 | This study |
| pT3-BfpI | pMPM-T3 derivative carrying bfpI | This study |
| pT3-BfpJ | pMPM-T3 derivative carrying bfpJ | This study |
| pT3-BfpK | pMPM-T3 derivative carrying bfpK | This study |
TABLE 2.
Primers used in this study
| Primer | Sequence |
|---|---|
| bfpI F cloning in pET48b+ | 5′ ATTGGATCCGGTTATTGTGCTTGTTCAG 3′ |
| bfpI R cloning in pET48b+ | 5′ ATTGAATTCCGCTTTCTTTTCTTATCA 3′ |
| bfpJFBamHI cloning in pET48b+ | 5′ TCTCTTTCTGTGGATCCCTATTACAAC 3′ |
| bfpJEcoRI cloning in pET48b+ | 5′ TCTCTTTCTGTGGATCCCTATTACAAC 3′ |
| FbfpK BamHI cloning in pQE31 | 5′ TTGTCTGTGGATCCTATTTCTGGCG 3′ |
| bfpKRHIII cloning in pQE31 | 5′ ACACCAGAAGAAAAGCTTAACTAGTACCGT 3′ |
| bfpJpKD4F λ red recombination | 5′ TCTTTTCTTTTACAATAATTTCCGGGTGGATGTTTTATGATAAGAGTGTAG GCTGGAGCTGCTTC 3′ |
| bfpJpKD4R λ red recombination | 5′ ATCGCTATTTCAATAAGCGACAGTCCCTTTTGCTGACGTCCTTCAAAAAT CATATGAATATCCTCCTTAG 3′ |
| bfpKpKD4 λ red recombination | 5′ ATTCAGACATCAGGGCAAAATTGTGACAGAGGTGGATATGAT TTTTGAAGGACGTCAGGTGTAGGCTGGAGCTGCTTC 3′ |
| bfpKpKD4R λ red recombination | 5′ AAAATGGATACAATTAATGAAAAAAAACCACACCAGAAGAATAA CATATGAATATCCTCCTTAG 3′ |
| bfpIpK λ red recombination | 5′ AATATGTAAGTAAAAATTATGGTTCGTCTATGTTTATATTG AGAGTGTAGGCTGGAGCTGCTTC 3′ |
| bfpIFcl qPCR | 5′ TTAAGTATGCTGTTCATGAAAATGC 3′ |
| bfpIRcl qPCR | 5′ ACATCCACCCGGAAATTATTGTAAA 3′ |
| bfpJF qPCR | 5′ GGAGCGCAGAACAACATTATATG 3′ |
| bfpJR qPCR | 5′ CACAATTTTGCCCTGATGTCTGAA 3′ |
| bfpKinF qPCR | 5′ TTGTAGCAACAGATATGAATTC 3′ |
| bfpKinR qPCR | 5′ CACTTAATAACTCTAACGGGAG 3′ |
| rpoAF qPCR | 5′ GGCTTGACGATTTCGACATC 3′ |
| rpoAR qPCR | 5′ GGTGAGAGTTCAGGGCAAAG 3′ |
| bfpAF qPCR | 5′ TGATTGAATCTGCAATGGTG 3′ |
| bfpAR qPCR | 5′ AGCATTCTGCGACTTATTGG 3′ |
| BfpIHinDIIIF clone in pMPMT3 | 5′ TACAGCCAAGCTTTACATCAAAGGAATATGT 3′ |
| bfpIBamHIR clone in pMPMT3 | 5′ AAGGATCCTTTCTTTTCTTATCATAAAACA 3′ |
| bfpJSmaIF clone in pMPMT3 | 5′ CTTTTACAATAATTCCCGGGTGG 3′ |
| bfpJSacIR clone in pMPMT3 | 5′ GTCCCTTTTGCTGAGCTCCTTCAAA 3′ |
| bfpKSmaIF clone in pMPMT3 | 5′ TTCAGACACCCGGGCAAAATTGTGA 3′ |
| bfpKXbaIR clone in pMPMT3 | 5′ CCACTCTAGAAGAATAACATAACTA 3′ |
Construction of EPEC mutants and complemented strains.
The EPEC mutants UMD963, UMD951, UMD957, CMP-J, CMP-1, CMP-2, CMP-3, CMP-4, and CMP-5 were constructed using the lambda red recombinase protocol described by Datsenko and Wanner (28). Briefly, the substitution of the gene of interest, bfpI (deleted from position 15 to 527), bfpJ (deleted from position 9 to 529), or bfpK (deleted from position 21 to 488), was accomplished using the pKD4 vector as a template by PCR amplifying the aph gene, encoding kanamycin resistance, using primers complementary to part of the Km cassette sequence and to part of the upstream and downstream regions of the target genes. The resulting PCR products containing the aph gene with flanking regions of the target genes then were transformed into either the EPEC wild-type strain E2348/69 or the EPEC bfpF mutant strain (UMD946) containing pKD46, encoding the lambda red proteins under the control of an arabinose-inducible promoter. The transformed colonies were selected by growing them at 37°C on LB plates supplemented with Km, and the desired mutations were verified by PCR and sequencing. The bfpI-, bfpJ-, and bfpK-complemented strains were created by cloning the amplified regions of each of these genes into pMPM-T3 using BamHI/HindIII for bfpI, SmaI/SacI for bfpJ, and SmaI/XbaI for bfpK restriction sites.
RNA extraction and quantitative reverse transcription-PCR (RT-PCR).
Total DNA-free RNA was prepared from the wild-type EPEC strain E2348/69 and EPEC mutant strains grown in DMEM to an optical density at 600 nm (OD600) of 0.5. RNA subsequently was extracted using the Qiagen RNA extraction kit combined with DNase treatment to remove residual DNA from the preparations. One microgram of RNA was used to synthesize cDNA using qScript cDNA SuperMix (Quanta). The cDNA next was diluted 10-fold and used to perform real-time quantitative PCR with PerfeCTa SYBR green FastMix for iQ (Quanta) and specific primers for the bfpA, bfpI, bfpJ, and bfpK genes. The rpoA gene was used as a calibration standard to normalize the resulting data. The relative expression values, determined by R = 2−(ΔCT) (where CT is threshold cycle), were calculated using the formula ΔCT = CT (bfp gene) − CT (rpoA gene).
eLA assay.
The early localized adherence (eLA) assay was performed as described previously (30, 31), with slight modifications. Briefly, subconfluent HEp-2 cell monolayers were grown for 24 h on 8-well slides (Lab-Tek chamber glass slide system) at 37°C in a humidified atmosphere of 5% CO2 in minimal essential medium (MEM) supplemented with 10% fetal bovine serum (FBS). Prior to each experiment, 40 μl of TSB (without dextrose)-grown bacteria was inoculated into 4 ml of DMEM supplemented with 0.5% glucose in a CO2 incubator for 60 min to induce BFP expression. These cultures then were applied onto subconfluent HEp-2 cell monolayers at a multiplicity of 100:1 (bacteria to HEp-2 cells), and the slides were placed in the 37°C CO2 incubator for 45 min. The wells were washed five times with Na+/K+ phosphate-buffered (pH 7.2) physiological saline (PBS) to remove unbound bacteria, fixed with methanol for 10 min, and stained with Giemsa stain for 20 min. EPEC eLA was recorded using a phase contrast light microscope. A total of 10 fields and approximately 150 to 200 individual HEp-2 cells were examined, and those having attached microcolonies consisting of five or more bacteria were recorded as positive for eLA EPEC. The results are expressed as a percentage of cells positive for eLA relative to the total number of cells examined. Each experiment was performed in triplicate and repeated at least three independent times.
Protein purification and antibody production.
Purified BfpI, BfpJ, and BfpK were obtained by amplifying and cloning their genes from EPEC strain E2348/69 lacking the DNA sequence encoding the first 25 amino acids from the N-terminal portion to enhance solubility. These then were cloned into the pET48b+ vector (Novagen) using the BamHI and EcoRI restriction sites for bfpI and bfpJ and into the pQE31 vector (Qiagen) for bfpK using the BamHI/HindIII restriction site. The pET48b+ plasmids then were transformed into E. coli BL21 cells, and protein expression was induced using 1 mM IPTG. The pQE31 vector was transformed into E. coli M15 cells, and its expression was induced using 1 mM IPTG. The three proteins were purified using nickel-nitrilotriacetic acid (Ni-NTA)–agarose (Qiagen) by following the manufacturer's protocol. New Zealand White rabbits were immunized using 100 μg of purified BfpI, BfpJ, or BfpK emulsified in 0.5 ml complete Freund's adjuvant by following the University of Calgary Health Sciences Animal Welfare Committee approved protocol M09030. The animals were given 2 booster immunizations at 3- and 4-week intervals using 50 μg purified proteins emulsified in incomplete Freund's adjuvant. After immunization, the animals were euthanized by following the standard procedure recommended by the Canadian Council on Animal Care (1993) and exsanguinated by cardiac puncture. To remove interfering antibodies and reduce background labeling, the resulting antisera were extensively absorbed against E. coli BL21.
TEM and immunogold labeling.
Bacteria were grown overnight at 37°C on BAP to induce BFP expression. Single colonies then were carefully picked from the agar and gently suspended in a solution consisting of 2% formaldehyde plus 0.5% glutaraldehyde in 50 mM cacodylate buffer (pH 7.4). The fixed samples subsequently were deposited onto Formvar/carbon-coated copper electron microscope grids and stained with 1% phosphotungstic acid in double-distilled/deionized water. Images were obtained using a Hitachi H-7650 transmission electron microscope (TEM).
For immunogold labeling, bacteria were grown overnight at 37°C on BAP. Several EPEC colonies then were carefully picked from the plates and gently suspended in 100 μl of chilled (4°C) fixative solution. Twenty microliters of each preparation next was applied to Formvar/carbon-coated nickel grids (Electron Microscopy Science), which were allowed to remain undisturbed for 15 min before subsequent processing. Excess liquid was gently removed and the grids were incubated for 20 min with 20 μl of 50 mM glycine dissolved in PBS. The glycine-PBS solution was removed and 20 μl of 3% skim milk plus 0.01% Tween 20 in PBS was applied. The grids were allowed to remain undisturbed for 30 min. After 3 washes, with 5 min between each wash and 20 μl of incubation buffer (0.01% BSA-c [Aurion] in PBS), the grids were incubated for 60 min with the rabbit polyclonal anti-BfpI, anti-BfpJ, anti-BfpK, or anti-purified BFP (a kind gift from Jorge Girón, Centro de Deteccion Biomolecular, VIEP-BUAP, Puebla, Mexico) or preimmune sera diluted 1:100 in incubation buffer. The grids then were washed 6 times with incubation buffer. A goat anti-rabbit IgG gold-conjugated (10- or 15-nm particle size; Electron Microscopy Science) secondary antibody was diluted 1:25 in incubation buffer and applied to the grids for 45 min. The grids then were washed 6 times with incubation buffer and an additional 3 times with double-distilled/deionized water. Finally, the grids were stained for 10 s with 1% phosphotungstic acid in double-distilled/deionized water, and images were obtained using a Hitachi H-7650 transmission electron microscope.
BFP isolation.
EPEC strains were harvested from heavily streaked BAP and resuspended in PBS. Samples were vigorously shaken using a Vortex mixer for 1 min and centrifuged at 3,000 × g during 30 min. The resulting supernatant solutions were centrifuged an additional 30 min at 17,000 × g. BFP were precipitated from this supernatant solution by adding saturated ammonium sulfate to a final concentration of 10%. Precipitated BFP were collected by centrifuging the ammonium sulfate solution at 3,000 × g for 30 min. The precipitated BFP subsequently were resuspended in PBS and the protein concentration was quantified. Ten micrograms of total protein was added to each lane of the immunoblot using anti-purified BFP-specific antibody as the primary antibody.
Autoaggregation assay.
The autoaggregation assay was performed as described by Anantha et al. (20), with some modifications. Bacteria were grown overnight in LB, supplemented with antibiotics as appropriate, at 37°C. The overnight cultures were diluted 1:50 in 4 ml high-glucose DMEM and incubated at 37°C for 4 h at 180 rpm. OD600 readings were taken at the end of the incubation period, and the samples then were vigorously shaken using a Vortex mixer for 30 s to disrupt bacterial aggregates. The OD600 of the agitated samples was recorded again, and the autoaggregation index at each time point was determined using the formula (AV-OD600 − BV-OD600)/BV-OD600, where BV-OD600 and AV-OD600 indicate the before and after Vortex-agitated OD600 values, respectively.
Quantitative analysis of surface bundlin expression.
Bacterial strains were grown overnight in LB at 37°C. Overnight cultures then were diluted 1:50 into 5 ml DMEM–F-12 and incubated at 37°C for 3 h with shaking at 180 rpm. An aliquot of cells next was centrifuged for 30 s at 13,000 × g, the supernatant solution was discarded, and the cells were resuspended in 100 μl PBS and heat fixed for 30 min using a 65°C heating block. The cells were centrifuged again for 30 s at 13,000 × g, resuspended in 200 μl PBS containing a 1:1,000 dilution of rabbit anti-α1-bundlin antibody (32), and incubated on a rotator overnight at 4°C. The samples were centrifuged, and the cell pellets were washed 3 times with PBS and then incubated for 1 h at 25°C with 200 μl PBS containing a 1:1,000 dilution of goat anti-rabbit antibody conjugated to Alexa Fluor 647. The cells were centrifuged again, washed 3 times with PBS, suspended in 200 μl of 1 μM Syto-16, and incubated for 30 min at 25°C. These samples were centrifuged, and the resulting cell pellets were washed once with PBS, suspended in 500 μl of PBS, and analyzed using a BDLSRII flow cytometer (Becton, Dickinson). The data from at least 104 cells were collected using FACSDiva software (Becton, Dickinson).
Statistics.
Analysis of the eLA and autoaggregation assays was accomplished with GraphPad Prism using one-way analysis of variance (ANOVA) with Tukey's multiple comparison test. Analysis for the flow cytometry assays was performed using a two-tailed t test.
RESULTS
A mutation in bfpF restores piliation in bfpI, bfpJ, and bfpK EPEC mutants.
Investigations into the role of minor pilin-like proteins in the structure and function of different T4aP systems have revealed that although mutant strains possessing single mutations of these genes do not express pili, strains possessing minor pilin-like protein gene mutations in addition to a mutation in the retraction ATPase (PilT) do (22, 33). The results presented in a previous report (19), confirmed in this study, also demonstrated that mutant EPEC strains lacking the ability to express BfpI, BfpJ, or BfpK do not produce BFP. Accordingly, we replaced the bfpI, bfpJ, and bfpK genes with a kanamycin resistance gene in an EPEC bfpF deletion mutant to determine if these mutants expressed BFP. The real-time quantitative RT-PCR data presented in Fig. 1 reveal the relative expression of bfpA, bfpI, bfpJ, and bfpK mRNA transcripts in the wild-type and mutant strains. These results demonstrate that inserting the Km cassette into each target gene did not affect the mRNA expression from downstream genes, and there was no statistical difference between the expression of the pilin-like genes in the different background strains. As expected from the presence of a Rho-independent transcription terminator sequence downstream of bfpA, pilin-like genes are expressed at significantly lower levels relative to that of bfpA in wild-type EPEC (34). Unfortunately, attempts to confirm the quantitative RT-PCR results at the translational level using minor pilin-specific antisera were unsuccessful, likely because the concentration of the minor pilins in the whole-cell or semicrude BFP preparations employed in these experiments was below the detection limit of the Western immunoblots.
FIG 1.
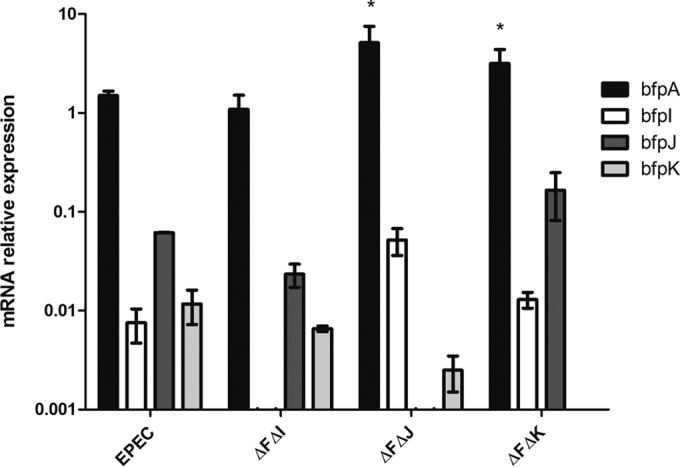
Expression for bfpA, bfpI, bfpJ, and bfpK relative to that of the housekeeping gene rpoA in wild-type and bfpF bfpI, bfpF bfpJ, and bfpF bfpK EPEC double mutant strains. Relative values are 2−(ΔCT). The assay was performed in triplicate three independent times. Error bars represent the standard errors of the means. *, P < 0.05 by two-way ANOVA comparing expression to that of WT EPEC.
TEM analysis (Fig. 2) of the double mutants revealed that the bfpF mutant strains lacking the ability to express BfpI, BfpJ, or BfpK were capable of expressing BFP. Remarkably, the BFP expressed by these mutants revealed phenotypic differences from the BFP produced by the single bfpF mutant (Fig. 2E) or by the wild-type EPEC strain (Fig. 2A). For example, the pili produced by the bfpF bfpI mutant appear to be more tightly entangled than those expressed by the bfpF mutant and are longer than the BFP expressed by the wild-type strain (Fig. 2A, B, and E). In contrast, the bfpF bfpJ mutant expresses highly fragmented pili, most of which were not attached to bacterial cells and which were detected only by immunogold staining using an anti-purified BFP antibody (Fig. 2C). Finally, the bfpF bfpK mutant strain expresses BFP that more closely resembled those produced by the wild-type EPEC strain rather than the bfpF mutant (Fig. 2A, D, and E). To determine if the absence of all three pilin-like proteins could affect the expression of BFP, we deleted all three genes from the wild-type and the bfpF background strains, thereby creating triple (bfpIJK) and quadruple (bfpFIJK) mutant strains. TEM analysis revealed that only the quadruple mutant strain produced BFP, but these pili appeared phenotypically similar to the BFP expressed by the bfpF bfpJ mutant, appearing shorter and less tangled than those expressed by the bfpF mutant (Fig. 3).
FIG 2.

Transmission electron microscope images (TEM) of wild-type EPEC and the double mutant strains grown on BAP overnight. Wild-type EPEC (A) and bfpF bfpI (B), bfpF bfpJ (C), bfpf bfpK (D), and bfpF (E) strains are shown. Panel C shows an immunogold-labeled image using the anti-purified BFP antibody. Scale bars represent 500 nm (A) and 100 nm (B, C, D, and E).
FIG 3.

TEM images of WT EPEC, bfpIJK, and bfpFIJK mutant strains. (D, E, and F) Immunogold labeling (15-nm gold particles) using an anti-BFP polyclonal antibody. The bfpIJK triple mutant (B and E) does not produce BFP. Filled arrowheads indicate flagella, open arrowheads indicate BFP, and the circle-tailed arrow indicates different pilus structures (possibly type 1 or common pili) that were not labeled with anti-BFP. Bar size, 100 nm.
We also assessed BFP expression by Western immunoblot analysis of bundlin present in isolated pili prepared from all the double as well as the triple and quadruple mutant strains. The results presented in Fig. 4 demonstrate that there was less bundlin in cell-free BFP preparations from all three of the double mutant strains than from their bfpF parent strain. As predicted by the TEM images, bundlin was not detected in samples from the triple mutant strain. Although TEM analysis revealed that the quadruple mutant strain expressed BFP, the bundlin signal was barely perceptible on the Western immunoblot presented in Fig. 4. Thus, there is a poor correlation between the TEM of the bfpF and bfpF bfpI mutants (Fig. 2B and E) and the Western immunoblot results presented in Fig. 4. This apparent discrepancy could be explained by the increased physical entanglement observed in some mutants, which may have caused a few of these pili to remain in the supernatant solution during the purification process, which relied on centrifugation to separate sheared BFP from the bacterial cells. Moreover, in the absence of all three minor pilins in the quadruple mutant, pseudopilins of the type 2 secretion system may prime for BFP assembly (35). However, the type 2 secretion system pseudopilins may not perfectly substitute for the priming function of BfpI, BfpJ, or BfpK, thereby leading to the reduced bundlin signal in the quadruple mutant.
FIG 4.
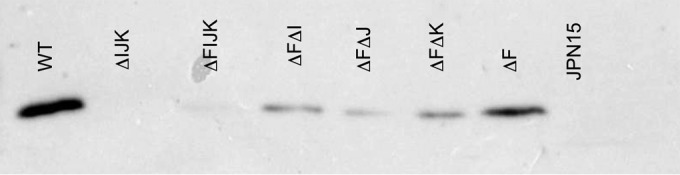
Anti-BfpA Western immunoblot analysis of isolated BFP prepared from WT EPEC mutant strains. JPN15 is an EPEC strain lacking pEAF and represents a negative control on the Western immunoblot.
Using flow cytometry, we next investigated the percentage of pilin-like mutant cells that expressed immune-reactive bundlin on their surfaces. This assay does not distinguish between unassembled bundlin and bundlin assembled into BFP. The data presented in Fig. 5 reveal that a greater percentage of the double mutant strains expressed surface bundlin than the wild-type strain. In addition, the quadruple mutant strain was capable of expressing bundlin on the surface of the bacteria (Fig. 5). These results also revealed that although we did not detect intact BFP on the single bfpJ and the triple bfpIJK mutant strains by TEM analysis, a significant percentage of these cells did express surface-exposed bundlin.
FIG 5.

Flow cytometry quantitative analysis of surface-exposed bundlin. Surface-exposed bundlin was detected by flow cytometry and expressed as the percentage of Alexa Fluor 647 fluorescence-positive cells also displaying Syto-16 fluorescence above background levels. (A) Surface expression of bundlin in WT and bfpA, bfpI, bfpJ, bfpK, and bfpIJK mutant strains. (B) Surface expression of bundlin in strains shown in panel A with a mutation in bfpF. Error bars indicate standard errors of the means from three independent experiments (**, P < 0.03 relative to the bfpA mutant; *, P < 0.03 relative to the wild-type strain; ns, not significant).
bfpF bfpI and bfpF bfpJ mutants do not adhere to HEp-2 cells.
It has been shown that only EPEC strains harboring the EAF plasmid express the LA phenotype, which was attributed to the expression of BFP. Therefore, we investigated whether the BFP produced by the pilin-like double mutants were functional in adherence to HEp-2 cells. Despite its piliated phenotype, the bfpF bfpI mutant displayed low eLA (see Materials and Methods) to HEp-2 cells (5%) relative to that for the bfpF mutant strain (80%) and the wild-type strain (52%) (Fig. 6). The bfpF bfpJ mutant also displayed significantly reduced eLA (7%). In contrast, the bfpF bfpK mutant adhered to HEp-2 cells better than either of the bfpF bfpI or bfpF bfpJ strains but less so than the bfpF deletion mutant in which it was created. Complementing the mutants with the bfpI or bfpJ gene (in trans) fully (bfpF bfpJ) or almost completely (bfpF bfpI; from 5% to 39%) restored the ability of the organisms to adhere to HEp-2 cells relative to that of the bfpF parent strain.
FIG 6.

Early localized adherence (eLA) assays performed with wild-type and T4bP mutant EPEC. WT EPEC, bfpF (ΔF), bfpI (ΔI), bfpJ (ΔJ), bfpK (ΔK), bfpF bfpI (ΔFΔI), bfpF BfpJ (ΔFΔJ), bfpF bfpK (ΔFΔK), bfpI+pT3BfpI (ΔI+I), bfpJ+pT3BfpJ (ΔJ+J), bfpK+pT3BfpK (ΔK+K), bfpF bfpI+pT3BfpI (ΔFΔI+I), bfpF bfpJ+pT3BfpJ (ΔFΔJ+J), and bfpF bfpK+pT3BfpK (ΔFΔK+K) strains are shown. The presence or absence of BFP, as revealed by TEM analysis, is indicated below the graph. The assays were performed by counting HEp-2 cells with EPEC microcolonies (at least 4 or more aggregated bacteria) in 10 fields in triplicate three separate times, and the error bars represent the standard deviations from the means. An asterisk indicates a significant difference (P < 0.001 by one-way ANOVA) between the single mutant strains and the WT, and a plus sign indicates a significant difference between the double mutant and the bfpF mutant strains.
To determine which of the three pilin-like proteins were critical to the EPEC eLA phenotype, we assessed the ability of the quadruple mutant and the corresponding complemented strains to adhere to HEp-2 cells. eLA of the bfpFIJK mutant strain was significantly diminished to 9% (Fig. 7), whereas it was 80% eLA for the bfpF mutant. Complementing the quadruple mutant strain using the pMPM-T3 plasmid containing either the bfpI, bfpJ, or bfpK gene individually revealed that bfpI significantly restored the ability of the bfpFIJK mutant to adhere to HEp-2 cells, increasing its eLA from 9% to 34% (Fig. 7). Although eLA appeared to increase when bfpJ or bfpK was reintroduced into the quadruple mutant, this increase was not significantly different relative to that displayed by the uncomplemented quadruple mutant strain (15% and 10%, respectively) but was significantly different from that of the bfpI-complemented strain.
FIG 7.
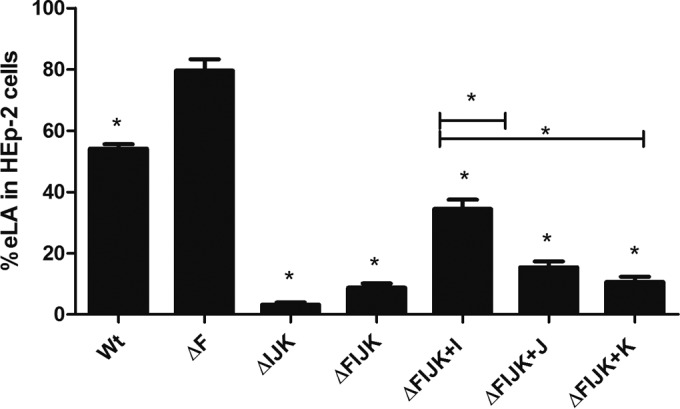
HEp-2 cell eLA of the WT, the quadruple mutant EPEC, and respective complemented strains. The assay was performed three times, each time in triplicate, allowing the strains to interact with the HEp-2 cells for 45 min. Error bars indicate standard errors of the means. *, P < 0.001 by one-way ANOVA relative to the EPEC ΔF strain and between the complemented strains.
Absence of BfpI and BfpJ affects autoaggregation in the EPEC bfpF mutant.
It was previously reported (20, 22) that T4aP and T4bP retraction ATPase mutants of both EPEC (bfpF) and N. meningitidis (pilT) display a greater autoaggregation phenotype than wild-type organisms. Consequently, we determined the autoaggregation phenotype (Fig. 8) of the double mutant in addition to the quadruple mutant strains and compared it to that of the bfpF parent mutant. The absence of BfpI from the bfpF bfpI mutant resulted in a significant (P < 0.0001 by one-way ANOVA) reduction in its autoaggregation index relative to that of the bfpF parent strain. In contrast, the bfpF bfpJ double mutant and the quadruple mutant strains displayed significantly (P < 0.001 by one-way ANOVA) less of a reduction in their autoaggregation indices than the bfpF strain (0.6 and 0.4 versus 2.9, respectively).
FIG 8.
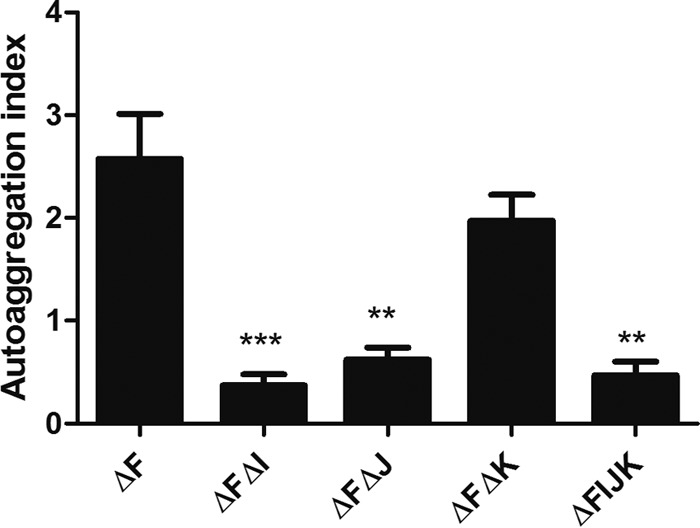
Autoaggregation index of EPEC mutants. Autoaggregation index of bfpF (ΔF), bfpF bfpI (ΔFΔI), bfpF bfpJ (ΔFΔJ), bfpF bfpK (ΔFΔK), and bfpFIJK (ΔFIJK) strains is shown. The assay was performed in triplicate. Error bars indicate standard errors of the means (P < 0.001 [**] and P < 0.0001 [***] for significant differences relative to the ΔF strain by one-way ANOVA with Tukey's multiple-comparison test).
BfpI, BfpJ, and BfpK are incorporated into the BFP filament.
Since BfpI, BfpJ, and BfpK share structural similarities with bundlin (1) (see Fig. S1 and S2 in the supplemental material), we used the TEM immunogold labeling technique to investigate whether these subunits are coassembled along with bundlin into the BFP filaments. As demonstrated by the images presented in Fig. 9, we detected immunogold labeling of BfpI, BfpJ, and BfpK subunits in the wild-type BFP but not in their respective bfpF bfpI, bfpF bfpJ, or bfpF bfpK double mutant BFP filaments. Moreover, TEM analysis (Fig. 9) revealed that although BfpI and BfpK pilin-like proteins were detected in BFP expressed in the bfpF bfpJ mutant strain, we could not detect BfpJ in either the bfpF bfpI or bfpF bfpK mutant strain.
FIG 9.

Immunogold (arrows) labeling of BfpI, BfpJ, and BfpK in BFP from wild-type EPEC and minor pilins mutants. (A) Anti-BfpI label; (B) anti-BfpJ label; (C) anti-BfpK label. Bars, 100 nm; immunogold particle size, 15 nm.
DISCUSSION
The structural and functional roles of minor pilins have been studied mostly in T4aP systems and just recently in the T4bP expressed by enterotoxigenic E. coli (22, 36–38). The BFP of enteropathogenic Escherichia coli represent a model of T4bP, and here we report the findings of the role of the minor pilin-like proteins in these pili. In the present study, we confirmed that whereas single mutants in bfpI, bfpJ, and bfpK pilin-like genes do not produce BFP, Western immunoblot and TEM analyses revealed that mutants created in an EPEC bfpF background strain do. Moreover, immunogold labeling revealed that all three of the pilin-like subunits (BfpI, BfpJ, and BfpK) are incorporated along with bundlin (BfpA) into the wild-type BFP filament and are important for BFP assembly and function (Fig. 4 and 9). It would seem appropriate, therefore, to refer to these subunits as minor pilins as reported in the T4aP expressed by N. meningitidis and P. aeruginosa (21, 39).
TEM analysis of immunogold-labeled preparations also revealed that the BfpI and BfpK minor pilins were incorporated into BFP produced in the bfpF bfpJ mutant strain, but we could not detect BfpJ in BFP expressed by either the bfpF bfpI or bfpF bfpK mutant strains (Fig. 9). Given that BfpJ was readily detected by immunogold labeling in BFP expressed by wild-type EPEC (Fig. 9), the greatly reduced or absent BfpJ immunogold labeling in the BFP expressed by the bfpF bfpI and bfpF bfpK mutant EPEC strains (Fig. 9) is most consistent with the conclusion that, in the absence of either BfpI or BfpK, BfpJ is not incorporated into BFP. It is also consistent with the observation (Fig. 7) that the presence of BfpJ alone had little positive influence on the host cell adhesion properties of BFP.
TEM analysis also revealed that, relative to BFP expressed by the wild-type or bfpF mutant strains, the bfpF bfpJ double mutant strain produced what appeared to be fragmented BFP, which were difficult to detect on the EM grids unless we employed the immunogold labeling technique. Moreover, many of these BFP fragments did not appear to be attached to the bacterial cells. Despite this, the Western immunoblot results presented in Fig. 4 demonstrated that the bfpF bfpJ double mutant expressed what appeared to be the same amount of BFP as the bfpF bfpI and bfpF bfpK double mutant strains. These observations suggest that BFP lacking BfpJ subunits is more prone to breakage than BFP expressed by the bfpF or wild-type EPEC strains. Therefore, BfpJ, perhaps in association with BfpI or BfpK, may contribute to the structural integrity of wild-type BFP, making them less prone to breakage. Nonetheless, despite its tendency to produce fragmented BFP, the bfpF bfpJ double mutant strain demonstrated low, but still detectable, levels of eLA to HEp-2 cells in addition to its autoaggregation phenotype (Fig. 6 and 8), suggesting that BFP lacking BfpJ that remained attached to the bacterial cell surface partially maintained their ability to promote adhesion.
Previously, we reported that bundlin displayed lectin-like activity, demonstrating specificity for N-acetyl-d-lactosamine glycan receptors on host cells (30, 40). Although TEM analysis revealed that the bfpF bfpI double mutant appeared to express BFP similar to that of the bfpF parental mutant, this strain displayed a significantly reduced ability to autoaggregate (Fig. 8) or adhere to HEp-2 cells (Fig. 6). These observations are reminiscent of those reported for the minor pilin PilX. The N. meningitidis pilX mutant expresses T4aP but is completely unable to adhere to or to form microcolonies on host cells (39, 41). Our results suggest that the absence of BfpI from BFP filaments either compromises the ability of bundlin to ligate its receptors on HEp-2 cells or that BfpI also serves a direct role, perhaps as a coadhesin, in the host cell recognition process. However, we cannot exclude the possibility that BfpJ also contributes to the BFP host cell adhesive function, since BfpJ also is lacking in the BFP expressed by the bfpF bfpI mutant strain (Fig. 9). More detailed structural analysis of BfpI, BfpJ, and the BFP produced by the bfpF bfpI double mutant is required to complete our understanding of how the conformation or assembly of BFP is affected when BfpI and BfpJ are absent.
With respect to BfpK, it would appear that, in the absence of retraction, this subunit is dispensable without compromising either the BFP structure, at least at the level of resolution of TEM, or eLA function (Fig. 6). However, the TEM observations revealed that, unlike the bfpF parent strain, the bfpF bfpK mutant appears to express BFP that more closely resemble in number those produced by the wild-type organisms. This suggests that BfpK, possibly in association with BfpJ (Fig. 9), is involved in regulating the BFP extension/retraction processes. In addition, TEM analysis revealed that the quadruple bfpFIJK mutant strain expressed BFP which appeared to be similar to those expressed by the bfpF bfpJ double mutant. It has been reported that pseudopilins of the type 2 secretion system may substitute for the minor pilins in priming T4P assembly in Pseudomonas aeruginosa (36). Further studies will be required to determine whether, in the absence of all three (BfpI, BfpJ, and BfpK) minor pilins, pseudopilins play a similar role in EPEC BFP expression (35, 42).
As anticipated, flow cytometry analysis, using a bundlin-specific polyclonal antibody, revealed a low percentage of bundlin-positive cells in the bfpA mutant population, representing background levels (Fig. 5A). Analysis of the bfpI and bfpK single mutant strains produced an equivalent result. In contrast, the percentage of bundlin-positive cells was significantly greater in the bfpJ and bfpIJK mutant strains than in the bfpA mutant. Despite this, TEM analysis did not reveal BFP in either of these mutants. These observations imply that although the bfpJ single and bfpIJK triple mutant strains appear not to express BFP that could be detected by TEM, a significant percentage of these cells have surface bundlin. We cannot exclude the presence of very short truncated pilus structures, perhaps associated with the BFP outer membrane assembly apparatus, or the possibility of unassembled bundlin on the outer membranes of the organisms. This suggests that BfpJ serves a role in regulating BFP assembly in addition to stabilizing the pilus filaments, as suggested by the image presented in Fig. 2C.
The flow cytometry technique also revealed that a significantly higher percentage of cells from all three double mutant strains and from the quadruple mutant expressed unassembled surface bundlin relative to percentages for the wild-type strain. Despite these results, TEM analysis did not reveal any apparent increase but rather the opposite in BFP expression by either the bfpF bfpJ, bfpF bfpK, or bfpFIJK mutant strain. TEM, however, would not have the same capability as the flow cytometry technique to detect immune-reactive short truncated BFP or unassembled bundlin on the surface of the bacteria. These observations do suggest, though, that apparent phenotypic differences in the pili expressed by the double mutant strains are due not to defects in the processing and transport of bundlin to the outer membrane BFP assembly apparatus but rather to pilus polymerization and stability. Furthermore, these results reveal a fundamental difference between the bfpJ and the bfpI and bfpK single mutants, since we could detect bundlin on the surface of the bfpJ single mutant strain but not in the bfpI or bfpK single mutants, while TEM analysis did not reveal assembled BFP in any of these single mutant strains. The ability of double retraction-deficient, minor pilin mutant strains to make T4P while single minor pilin strains cannot has been interpreted as evidence that minor pilins facilitate pilus extension, impede retraction, or both. Whatever the function with regard to the relative efficiency of pilus extension and retraction, it appears that BfpJ plays less of a role than the other pilin proteins, since bundlin monomers still can reach the surface in the bfpJ mutant but not in the bfpI or bfpK mutants. Moreover, analysis of the triple bfpIJK mutant indicates that the net effect of losing all three minor pilins still permits some bundlin to reach the bacterial cell surface.
Although it is tempting to use our immunogold labeling results to estimate the number of BfpI, BfpJ, and BfpK subunits incorporated into the BFP filaments relative to bundlin, this would require using antibodies with avidities similar to those of their respective antigens. However, enzyme-linked immunosorbent assay (ELISA) results (data not shown) indicate that the BfpI-, BfpJ-, and BfpK-specific antisera used in the immunogold labeling experiments do not fulfill this requirement. Moreover, because they form dense rope-like structures, it is also extremely challenging to determine the relative frequency or localization of BfpI, BfpJ, or BfpK incorporation into individual BFP filaments.
In sum, here we present evidence that the minor pilins BfpI, BfpJ, and BfpK are incorporated into BFP expressed by EPEC. BfpI might have a direct role as a coadhesin or indirectly participate in maintaining the BFP host cell recognition properties. The results also suggest that BfpJ is required to maintain BFP flexibility or tensile strength, and that BfpK regulates either the extension or the retraction phase of BFP expression.
Supplementary Material
ACKNOWLEDGMENTS
We thank Jorge Girón, Centro de Deteccion Biomolecular, VIEP-BUAP, Puebla, Mexico, for providing the BFP-specific rabbit antiserum.
C.F.M.D.L.P. was the recipient of CONACYT scholarship 309086 from the Government of Mexico. L.D.M., S.N., and M.S.D. were supported by Public Health Service award R01 AI37606 from the National Institutes of Health, and L.D.M. also received support from Public Health Service award T32 DK067872 from the National Institutes of Health. Funding to G.D.A. was from the Alberta Glycomics Centre. J.T. was the recipient of an NSERC Summer Studentship.
Footnotes
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JB.00818-15.
REFERENCES
- 1.Nataro JP, Kaper JB. 1998. Diarrheagenic Escherichia coli. Clin Microbiol Rev 11:142–201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Moon HW, Whipp SC, Argenzio RA, Levine MM, Giannella RA. 1983. Attaching and effacing activities of rabbit and human enteropathogenic Escherichia coli in pig and rabbit intestines. Infect Immun 41:1340–1351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Giron JA, Ho AS, Schoolnik GK. 1991. An inducible bundle-forming pilus of enteropathogenic Escherichia coli. Science 254:710–713. doi: 10.1126/science.1683004. [DOI] [PubMed] [Google Scholar]
- 4.Scaletsky IC, Silva ML, Trabulsi LR. 1984. Distinctive patterns of adherence of enteropathogenic Escherichia coli to HeLa cells. Infect Immun 45:534–536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nougayrede JP, Fernandes PJ, Donnenberg MS. 2003. Adhesion of enteropathogenic Escherichia coli to host cells. Cell Microbiol 5:359–372. doi: 10.1046/j.1462-5822.2003.00281.x. [DOI] [PubMed] [Google Scholar]
- 6.Maier B, Potter L, So M, Long CD, Seifert HS, Sheetz MP. 2002. Single pilus motor forces exceed 100 pN. Proc Natl Acad Sci USA 99:16012–16017. doi: 10.1073/pnas.242523299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Merz AJ, So M, Sheetz MP. 2000. Pilus retraction powers bacterial twitching motility. Nature 407:98–102. doi: 10.1038/35024105. [DOI] [PubMed] [Google Scholar]
- 8.Bradley DE. 1980. A function of Pseudomonas aeruginosa PAO polar pili: twitching motility. Can J Microbiol 26:146–154. doi: 10.1139/m80-022. [DOI] [PubMed] [Google Scholar]
- 9.Kaiser D. 2000. Bacterial motility: how do pili pull? Curr Biol 10:R777–R780. doi: 10.1016/S0960-9822(00)00764-8. [DOI] [PubMed] [Google Scholar]
- 10.Aas FE, Wolfgang M, Frye S, Dunham S, Lovold C, Koomey M. 2002. Competence for natural transformation in Neisseria gonorrhoeae: components of DNA binding and uptake linked to type IV pilus expression. Mol Microbiol 46:749–760. doi: 10.1046/j.1365-2958.2002.03193.x. [DOI] [PubMed] [Google Scholar]
- 11.Aas FE, Lovold C, Koomey M. 2002. An inhibitor of DNA binding and uptake events dictates the proficiency of genetic transformation in Neisseria gonorrhoeae: mechanism of action and links to type IV pilus expression. Mol Microbiol 46:1441–1450. doi: 10.1046/j.1365-2958.2002.03265.x. [DOI] [PubMed] [Google Scholar]
- 12.Buchanan TM, Pearce WA. 1976. Pili as a mediator of the attachment of gonococci to human erythrocytes. Infect Immun 13:1483–1489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bieber D. 1998. Type IV pili, transient bacterial aggregates, and virulence of enteropathogenic Escherichia coli. Science 280:2114–2118. doi: 10.1126/science.280.5372.2114. [DOI] [PubMed] [Google Scholar]
- 14.Craig L, Pique ME, Tainer JA. 2004. Type IV pilus structure and bacterial pathogenicity. Nat Rev Microbiol 2:363–378. doi: 10.1038/nrmicro885. [DOI] [PubMed] [Google Scholar]
- 15.Pasloske BL, Paranchych W. 1988. The expression of mutant pilins in Pseudomonas aeruginosa: fifth position glutamate affects pilin methylation. Mol Microbiol 2:489–495. doi: 10.1111/j.1365-2958.1988.tb00055.x. [DOI] [PubMed] [Google Scholar]
- 16.Strom MS, Lory S. 1993. Structure-function and biogenesis of the type IV pili. Annu Rev Microbiol 47:565–596. doi: 10.1146/annurev.mi.47.100193.003025. [DOI] [PubMed] [Google Scholar]
- 17.Zhang HZ, Donnenberg MS. 1996. DsbA is required for stability of the type IV pilin of enteropathogenic Escherichia coli. Mol Microbiol 21:787–797. doi: 10.1046/j.1365-2958.1996.431403.x. [DOI] [PubMed] [Google Scholar]
- 18.Giron JA, Ho AS, Schoolnik GK. 1993. Characterization of fimbriae produced by enteropathogenic Escherichia coli. J Bacteriol 175:7391–7403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ramer SW, Schoolnik GK, Wu CY, Hwang J, Schmidt SA, Bieber D. 2002. The type IV pilus assembly complex: biogenic interactions among the bundle-forming pilus proteins of enteropathogenic Escherichia coli. J Bacteriol 184:3457–3465. doi: 10.1128/JB.184.13.3457-3465.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Anantha RP, Stone KD, Donnenberg MS. 1998. Role of BfpF, a member of the PilT family of putative nucleotide-binding proteins, in type IV pilus biogenesis and in interactions between enteropathogenic Escherichia coli and host cells. Infect Immun 66:122–131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Giltner CL, Habash M, Burrows LL. 2010. Pseudomonas aeruginosa minor pilins are incorporated into type IV pili. J Mol Biol 398:444–461. doi: 10.1016/j.jmb.2010.03.028. [DOI] [PubMed] [Google Scholar]
- 22.Winther-Larsen HC, Wolfgang M, Dunham S, Van Putten JPM, Dorward D, Løvold C, Aas FE, Koomey M. 2005. A conserved set of pilin-like molecules controls type IV pilus dynamics and organelle-associated functions in Neisseria gonorrhoeae. Mol Microbiol 56:903–917. doi: 10.1111/j.1365-2958.2005.04591.x. [DOI] [PubMed] [Google Scholar]
- 23.Carbonnelle E, Hélaine S, Prouvensier L, Nassif X, Pelicic V. 2005. Type IV pilus biogenesis in Neisseria meningitidis: PilW is involved in a step occurring after pilus assembly, essential for fibre stability and function. Mol Microbiol 55:54–64. [DOI] [PubMed] [Google Scholar]
- 24.Levine MM, Bergquist EJ, Nalin DR, Waterman DH, Hornick RB, Young CR, Sotman S. 1978. Escherichia coli strains that cause diarrhoea but do not produce heat-labile or heat-stable enterotoxins and are non-invasive. Lancet i:1119–1122. [DOI] [PubMed] [Google Scholar]
- 25.Lieberman JA, Frost NA, Hoppert M, Fernandes PJ, Vogt SL, Raivio TL, Blanpied TA, Donnenberg MS. 2012. Outer membrane targeting, ultrastructure, and single molecule localization of the enteropathogenic Escherichia coli type IV pilus secretin BfpB. J Bacteriol 194:1646–1658. doi: 10.1128/JB.06330-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Jerse AE, Yu J, Tall BD, Kaper JB. 1990. A genetic locus of enteropathogenic Escherichia coli necessary for the production of attaching and effacing lesions on tissue culture cells. Proc Natl Acad Sci U S A 87:7839–7843. doi: 10.1073/pnas.87.20.7839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mayer MP. 1995. A new set of useful cloning and expression vectors derived from pBlueScript. Gene 163:41–46. doi: 10.1016/0378-1119(95)00389-N. [DOI] [PubMed] [Google Scholar]
- 28.Datsenko KA, Wanner BL. 2000. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc Natl Acad Sci USA 97:6640–6645. doi: 10.1073/pnas.120163297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sambrook J, Russell D. 2001. Molecular cloning: a laboratory manual, 3rd ed Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. [Google Scholar]
- 30.Hyland RM, Sun J, Griener TP, Mulvey GL, Klassen JS, Donnenberg MS, Armstrong GD. 2008. The bundlin pilin protein of enteropathogenic Escherichia coli is an N-acetyllactosamine-specific lectin. Cell Microbiol 10:177–187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Vanmaele RP, Heerze LD, Armstrong GD. 1999. Role of lactosyl glycan sequences in inhibiting enteropathogenic Escherichia coli attachment. Infect Immun 67:3302–3307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fernandes PJ, Guo Q, Donnenberg MS. 2007. Functional consequences of sequence variation in bundlin, the enteropathogenic Escherichia coli type IV pilin protein. Infect Immun 75:4687–4696. doi: 10.1128/IAI.00009-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Carbonnelle E, Hélaine S, Nassif X, Pelicic V. 2006. A systematic genetic analysis in Neisseria meningitidis defines the Pil proteins required for assembly, functionality, stabilization and export of type IV pili. Mol Microbiol 61:1510–1522. doi: 10.1111/j.1365-2958.2006.05341.x. [DOI] [PubMed] [Google Scholar]
- 34.Donnenberg MS, Giron JA, Nataro JP, Kaper JB. 1992. A plasmid-encoded type IV fimbrial gene of enteropathogenic Escherichia coli associated with localized adherence. Mol Microbiol 6:3427–3437. doi: 10.1111/j.1365-2958.1992.tb02210.x. [DOI] [PubMed] [Google Scholar]
- 35.Cisneros DA, Pehau-Arnaudet G, Francetic O. 2012. Heterologous assembly of type IV pili by a type II secretion system reveals the role of minor pilins in assembly initiation. Mol Microbiol 86:805–818. doi: 10.1111/mmi.12033. [DOI] [PubMed] [Google Scholar]
- 36.Nguyen Y, Sugiman-Marangos S, Harvey H, Bell SD, Charlton CL, Junop MS, Burrows LL. 2015. Pseudomonas aeruginosa minor pilins prime type IVa pilus assembly and promote surface display of the PilY1 adhesin. J Biol Chem 290:601–611. doi: 10.1074/jbc.M114.616904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kuchma SL, Griffin EF, O'Toole GA. 2012. Minor pilins of the type IV pilus system participate in the negative regulation of swarming motility. J Bacteriol 194:5388–5403. doi: 10.1128/JB.00899-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kawahara K, Oki H, Fukakusa S, Maruno T, Kobayashi Y, Motooka D, Taniguchi T, Honda T, Iida T, Nakamura S, Ohkubo T. 2015. Cloning, expression, purification, crystallization and X-ray crystallographic analysis of CofB, the minor pilin subunit of CFA/III from human enterotoxigenic Escherichia coli. Acta Crystallogr F Struct Biol Commun 71:663–667. doi: 10.1107/S2053230X15005890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Helaine S, Dyer DH, Nassif X, Pelicic V, Forest KT. 2007. 3D structure/function analysis of PilX reveals how minor pilins can modulate the virulence properties of type IV pili. Proc Natl Acad Sci USA 104:15888–15893. doi: 10.1073/pnas.0707581104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Humphries RM, Griener TP, Vogt SL, Mulvey GL, Raivio T, Donnenberg MS, Kitov PI, Surette M, Armstrong GD. 2010. N-acetyllactosamine-induced retraction of bundle-forming pili regulates virulence-associated gene expression in enteropathogenic Escherichia coli. Mol Microbiol 76:1111–1126. doi: 10.1111/j.1365-2958.2010.07192.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Helaine S, Carbonnelle E, Prouvensier L, Beretti JL, Nassif X, Pelicic V. 2005. PilX, a pilus-associated protein essential for bacterial aggregation, is a key to pilus-facilitated attachment of Neisseria meningitidis to human cells. Mol Microbiol 55:65–77. [DOI] [PubMed] [Google Scholar]
- 42.Cisneros DA, Bond PJ, Pugsley AP, Campos M, Francetic O. 2012. Minor pseudopilin self-assembly primes type II secretion pseudopilus elongation. EMBO J 31:1041–1053. doi: 10.1038/emboj.2011.454. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.