

ABSTRACT
Ebola virus (EBOV) is an RNA virus that can cause hemorrhagic fever with high fatality rates, and there are no approved vaccines or therapies. Typically, RNA viruses have high spontaneous mutation rates, which permit rapid adaptation to selection pressures and have other important biological consequences. However, it is unknown if filoviruses exhibit high mutation frequencies. Ultradeep sequencing and a recombinant EBOV that carries the gene encoding green fluorescent protein were used to determine the spontaneous mutation frequency of EBOV. The effects of the guanosine analogue ribavirin during EBOV infections were also assessed. Ultradeep sequencing revealed that the mutation frequency for EBOV was high and similar to those of other RNA viruses. Interestingly, significant genetic diversity was not observed in viable viruses, implying that changes were not well tolerated. We hypothesized that this could be exploited therapeutically. In vitro, the presence of ribavirin increased the error rate, and the 50% inhibitory concentration (IC50) was 27 μM. In a mouse model of ribavirin therapy given pre-EBOV exposure, ribavirin treatment corresponded with a significant delay in time to death and up to 75% survival. In mouse and monkey models of therapy given post-EBOV exposure, ribavirin treatment also delayed the time to death and increased survival. These results demonstrate that EBOV has a spontaneous mutation frequency similar to those of other RNA viruses. These data also suggest a potential for therapeutic use of ribavirin for human EBOV infections.
IMPORTANCE Ebola virus (EBOV) causes a severe hemorrhagic disease with high case fatality rates; there are no approved vaccines or therapies. We determined the spontaneous mutation frequency of EBOV, which is relevant to understanding the potential for the virus to adapt. The frequency was similar to those of other RNA viruses. Significant genetic diversity was not observed in viable viruses, implying that changes were not well tolerated. We hypothesized that this could be exploited therapeutically. Ribavirin is a viral mutagen approved for treatment of several virus infections; it is also cheap and readily available. In cell culture, we showed that ribavirin was effective at reducing production of infectious EBOV. In mouse and monkey models of therapy given post-EBOV exposure, ribavirin treatment delayed the time to death and increased survival. These data provide a better understanding of EBOV spontaneous mutation and suggest that ribavirin may have great value in the context of human disease.
INTRODUCTION
Ebola virus (EBOV), a member of the Filoviridae family, is an enveloped, single-stranded, negative-sense RNA virus that can cause Ebola virus disease (EVD) with variable fatality rates of up to 90% (1, 2). There are no approved vaccines or therapies for EBOV infections, and the West African outbreak in 2014 that became the largest outbreak on record highlights the urgency for advancements in this area (1–3).
Typically, RNA viruses have high spontaneous mutation rates due to error-prone RNA-dependent RNA polymerases. However, it is unknown if filoviruses exhibit a high spontaneous mutation rate (3, 4). It had been thought that EBOV did not evolve in the context of a single outbreak (4), but recent data from the West African outbreak suggest that there is evolution (5–8). Filoviruses have a wide host range and show great potential for emergence: to new geographic regions, to new hosts, and by new modes of transmission (9–15). Information about mutation frequencies in filoviruses could help to lead to a better understanding of these topics. Furthermore, from the current outbreak, we are learning the importance of how the virus changes and how this could affect future diagnostics, treatments, and preventative measures (6–8).
We found that EBOV was unable to tolerate excessive changes in the genome, and thus it was reasoned that chemically increasing the mutation rate may decrease the number of viable virions released from a cell. Thus, an increase in mutation frequency could be an effective therapeutic mechanism, as it is for other RNA viruses (16–18). However, the effect on filoviruses is unclear (19–21).
Ribavirin, a guanosine analogue, shows broad-spectrum antiviral activity and is commonly used to treat infections caused by several RNA viruses, including hepatitis C virus and Lassa virus (17, 18, 22). Ribavirin likely acts through a variety of mechanisms but has been shown to be a potent mutagen in vitro (18), and there is some evidence to suggest that in vivo therapeutic effects may also be related to lethal mutagenesis (23, 24). If EBOV is susceptible to mutagenesis, then error catastrophe could occur, thus giving ribavirin an antiviral effect against EBOV. Even a moderate efficacy of ribavirin in the treatment of EVD may provide a significant clinical benefit: ribavirin is inexpensive, does not require a cold chain, can be delivered orally, and is already available in many areas of Africa (25). Furthermore, ribavirin has already undergone rigorous safety testing and received FDA approval for the treatment of several other viral infections. As a result, it could be implemented immediately as a therapy for EVD rather than being delayed in early-stage clinical trials. This study was designed to determine if EBOV exhibits a high mutation frequency and to evaluate the potential of ribavirin for use as a therapy in EBOV infections.
MATERIALS AND METHODS
Ethics statement.
Animal research was conducted under Institutional Animal Care and Use Committee (IACUC)-approved protocols in compliance with the Animal Welfare Act and other federal statutes and regulations relating to animals and experiments involving animals. The Texas Biomedical Research Institute (TBRI) is accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International and adheres to the principles stated in the 8th edition of the Guide for the Care and Use of Laboratory Animals (26).
Cells and virus.
BEI-sourced Vero E6 cells were grown in minimum essential medium (MEM; Gibco) containing 2 mM l-glutamine and 1 mM sodium pyruvate (henceforth referred to as normal growth medium) with 10% heat-inactivated fetal calf serum (FCS; Gibco) at 37°C with 5% CO2. To determine the mutation frequency, a recombinant Zaire ebolavirus containing green fluorescent protein (eGFP-ZEBOV; kindly provided by H. Feldmann) was used. Mouse-adapted EBOV was obtained from the U.S. Army Medical Research Institute of Infectious Diseases (USAMRIID) and then propagated in Vero E6 cells at a multiplicity of infection (MOI) of 0.01, with the viral supernatant harvested at 7 days postinfection (dpi) (27). The virus received from USAMRIID was originally obtained by Bray et al. through passage in suckling BALB/c mice (28). Ebola virus Kikwit (passage number 3, Vero E6 cells [GenBank accession number KR063672]; passage number 2, Ebola virus Homo sapiens-tc/COD/1995/Kikwit-9510621 [species Zaire ebolavirus]; acquired from T. Ksiazek, University of Texas Medical Branch) was used for the nonhuman primate (NHP) experiment.
In vitro eGFP-ZEBOV and ribavirin infections.
eGFP-ZEBOV was grown at a low MOI (MOI of 0.001) in Vero E6 cells with normal growth medium containing 2% FCS and a range of concentrations of ribavirin (Spectrum Laboratories). The concentrations used were 100 μM, 200 μM, 400 μM, 500 μM, 700 μM, and 1 mM. Viral supernatants were harvested after 5 days and titrated, and the RNAs were prepared for deep sequencing as described below.
Deep sequencing and sample preparation.
Isolation of viral RNA, sample preparation for deep sequencing, and deep sequencing were performed as previously described (29). Briefly, an aliquot of viral supernatant or serum was diluted in TRIzol LS reagent (Life Technologies). Samples were next transferred to a biosafety level 2 (BSL2) laboratory, where RNA was harvested following the manufacturer's instructions. After isolation, rRNA and mRNA were removed as described previously (29). Finally, the supernatant containing mRNA-depleted RNA was removed and prepared for deep sequencing using Illumina's TruSeq RNA sample preparation kit following the manufacturer's protocol. The final cDNA library was sequenced using sequencing by synthesis (Illumina) on an Illumina MiSeq machine. To determine the number of single nucleotide polymorphisms (SNPs) arising from errors introduced during the sequencing process (for example, from reverse transcription), we included a spiked in vitro-transcribed RNA of known sequence when samples were prepared. This RNA sequence was from a related filovirus that would recapitulate the properties of EBOV RNA during sample preparation. Data analysis began by using the Illumina pipeline to generate a FASTQ file containing all the reads, which were then mapped to the parental virus sequence by using Lasergene SeqMan NGen (DNASTAR). The abundance of each nucleotide at each position in the genome was determined and compared to the parental sequence. Mutation frequencies were then calculated as numbers of SNPs per nucleotide sequenced.
Determination of viral titers.
Viral titers were determined by plaque assay using Vero E6 cells and agarose overlay as previously described (30).
Animal studies.
Animals were randomly assigned to groups, and the studies were not blinded. For the mouse studies, a total of 80 BALB/c mice (Mus musculus; Charles River Laboratories) of 4 to 6 weeks of age and weighing 17 to 22 g were used. Mice were housed in groups of four or eight and fed mouse chow. Enrichment included shepherd shacks and nesting material. For the NHP studies, four male and four female cynomolgus macaques (Macaca fascicularis) of 2 to 4 years of age and weighing 2 to 4 kg were used. Animals were acquired from Covance and serum tested to ensure that no reactivity to filovirus antigen was present prior to purchase. Animals were shipped directly to TBRI and underwent a standard quarantine period during which all animals were allowed to acclimate to singly housed caging and the in-house diet. Health status and well-being were monitored at least twice a day, and enrichment included visual stimulation, commercial toys, and food supplements. Prior to blood collections, animals were anesthetized using Telazol (Zoetis Inc.). For all animal studies, virus challenge occurred on study day 0. Test subjects were then observed at least twice daily for up to 21 days postchallenge, at which time any survivors were euthanized. Critically ill animals were observed at least four times per day, and species-specific euthanasia criteria were followed to minimize undue pain and distress. Mice were anesthetized via intraperitoneal (i.p.) injection of a ketamine-xylazine combination and then euthanized by inhalation of carbon dioxide followed by cervical dislocation. M. fascicularis macaques were euthanized with an intravenous overdose of sodium pentobarbital.
Experimental inoculation of mice with EBOV.
For the prophylactic experiment, three groups of eight mice each were challenged i.p. with a target dose of 1,000 PFU of mouse-adapted EBOV (back titration indicated that the dose was 177 PFU) as previously described (27). Two treatment groups received i.p. injections of ribavirin 1 h prior to challenge and daily thereafter; one group received a dose of 50 mg/kg of body weight, while the other group received 80 mg/kg. For the postexposure experiment, five groups of eight mice each were challenged i.p. with a target dose of 1,000 PFU of mouse-adapted EBOV (back titration indicated that the dose was 241 PFU). One day after challenge and daily thereafter, the animals received ribavirin injections i.p. One group received saline as a vehicle control, and each of the other four groups received a different dose of ribavirin treatment (30 mg/kg, 50 mg/kg, 100 mg/kg, or 100-mg/kg loading dose with 50-mg/kg subsequent doses). To identify any detrimental effects of ribavirin treatment alone, four other groups of four animals each received the same daily ribavirin treatments but were not challenged with EBOV.
Experimental inoculation of NHPs with EBOV.
Four M. fascicularis macaques, two female and two male, were injected intramuscularly (i.m.) in the deltoid muscle with a target dose of 100 PFU of EBOV (back titration indicated that the dose was 38 PFU). One day after challenge and every 12 h thereafter, three animals received i.m. treatment injections and one animal received saline as a vehicle control. The treated animals received 30 mg/kg of ribavirin for day 1 through day 3 postchallenge. During day 4 postchallenge and throughout the rest of the study, the drug dose was decreased to 15 mg/kg. Blood samples were collected on days 0, 3, 5, and 7 relative to inoculation. During each scheduled blood collection, the rectal temperature was taken and the weight was recorded. To identify any detrimental effects of ribavirin treatment alone, four animals were not infected and were housed at animal biosafety level 2 (ABSL2). These animals received the same drug treatment regimen as the infected animals housed at ABSL4 and were not euthanized at the end of the project.
Preparation and administration of ribavirin treatment.
Ribavirin (Spectrum Laboratories) treatment doses were prepared daily to prevent loss of drug activity. The appropriate amount of powder was dispensed under sterile conditions and combined with sterile saline (Hospira). For mouse treatments, 250-μl doses were administered every 24 h throughout the duration of the study. During each morning observation and treatment administration, all animals were weighed to determine the average weight per animal for each cage; this value was used to calculate the amount of ribavirin animals in each cage would receive during the next treatment. For NHP treatments, a single concentration of drug was used, and the treatment volume was based on each animal's weight. Injections were administered i.m. every 12 h throughout the duration of the study. The body weight recorded on blood draw days was used to calculate the amount of ribavirin each animal would receive until the next weight measurement was taken.
Coagulation and blood chemistry.
Biochemical analysis was performed using a mammalian liver enzyme profile rotor on a Vet Scan analyzer (Abaxis, Inc.). Coagulation times were determined using an Idexx Coag Dx analyzer (Idexx Laboratories).
Statistics.
The log-rank Mantel-Cox test was used to analyze the survival curves, and the P value is displayed on each graph. GraphPad Prism two-way analysis of variance (ANOVA) with the Bonferroni posttest was used to analyze weight and titer data. Mouse group sizes were determined statistically (Fisher's exact test, using an α value of 0.05). The NHP study was a pilot project, and the group variances were unknown.
RESULTS
Characterization of mutation frequency of Ebola virus in vitro.
To measure the mutation frequency of EBOV, we utilized a recombinant EBOV that carries the gene encoding green fluorescent protein (eGFP-ZEBOV) (31). This permitted measurement of spontaneous mutations in the recombinant virus by quantifying errors accumulated over the GFP-encoding region—a locus less subject to selection pressures than the wild-type viral genome—which is a method that has been applied previously in studies of bacterial spontaneous mutation rates (32). Thus, using eGFP-ZEBOV allowed enumeration of errors caused by the EBOV RNA-dependent RNA polymerase over the GFP region. eGFP-ZEBOV RNA was subjected to ultradeep sequencing, and the abundance of each sequenced nucleotide at each genome position was determined and compared to a reference sequence. Mutation frequencies were then presented as numbers of SNPs per nucleotide sequenced. Over the GFP region of eGFP-ZEBOV, a total of 3.9 × 105 nucleotides were sequenced, with a mean depth of coverage of 6.7 × 102. There was a lower mean depth of coverage over the GFP region than over other regions of the viral genome (1.2 × 109 nucleotides were sequenced, with a mean depth of coverage of 6.2 × 104). This suggested that not all of the genomes sequenced had maintained the GFP gene, which suggests that negative selection took place. The difference between the highest and lowest depths of coverage for the GFP region was low (3.3-fold difference in the number of bases sequenced at each site; the range of depth of coverage per site was 3.2 × 102 to 1.0 × 103 nucleotides sequenced, with a median of 6.5 × 102 nucleotides sequenced). This implies that the whole ORF was deleterious and not well maintained in the viral genome, likely due to the viral requirement for maintaining a compact genome. This should not have affected the estimation of mutation frequency based on SNPs.
The GFP region accumulated 4.1 × 10−4 SNP per nucleotide sequenced, which is similar to the mutation frequencies of other RNA viruses (16, 33, 34). The number of insertions or deletions (indels) per base sequenced was 2.2 × 10−2. The number of SNPs per nucleotide sequenced across wild-type regions of the genome was determined to be below the limit of detection, as it was the same as the number calculated for the control (9.1 × 10−4 SNP per nucleotide sequenced).
Effect of guanosine analogue on Ebola virus mutation frequency in vitro.
To determine the effect of the guanosine analogue ribavirin on the EBOV mutation frequency in vitro, eGFP-ZEBOV was grown at a low MOI in Vero E6 cells with medium containing 500 μM ribavirin. The viral supernatant was removed, and viral RNA was isolated and deep sequenced. Over the GFP region of eGFP-ZEBOV, a total of 6.1 × 107 nucleotides were sequenced, and the mean depth of coverage was 1.0 × 105. eGFP-ZEBOV grown in the presence of ribavirin accumulated 27-fold more SNPs per nucleotide sequenced (1.1 × 10−2) and 17-fold more indels (3.8 × 10−1) in the GFP region than eGFP-ZEBOV grown without ribavirin. Importantly, when the virus was grown in the presence of ribavirin, the number of SNPs per nucleotide sequenced (8.1 × 106 nucleotides total, with a mean depth of coverage of 4.4 × 102) across the wild-type viral genome increased to a detectable level (1.3 × 10−2) that was similar to the number found in the GFP region. Ribavirin treatment also correlated with a high ratio of transversions to transitions (3.9:1 ratio in the wild-type viral genome and 4.7:1 ratio in the GFP region).
Effect of guanosine analogue on production of infectious Ebola virus in vitro.
To determine the effect of the guanosine analogue ribavirin on production of infectious EBOV in vitro, eGFP-ZEBOV was grown at a low MOI in Vero E6 cells with media containing a range of concentrations of ribavirin. Consistent with other published reports, ribavirin-induced cytotoxicity was not observed at the concentrations used (18, 35, 36). Viral titers of the supernatants revealed that increasing concentrations of ribavirin resulted in decreasing quantities of infectious virus (Fig. 1). eGFP-ZEBOV-infected cells treated with 1 mM ribavirin showed a titer that was >1,000-fold lower than that of untreated virus. The half-maximal inhibitory concentration (IC50) was determined to be 27 μM.
FIG 1.
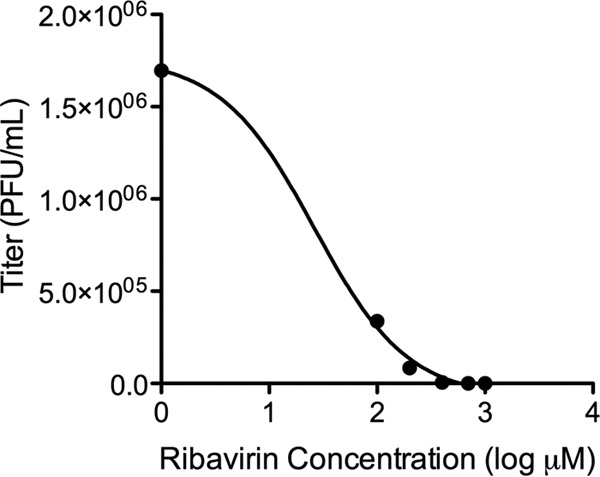
Effects of ribavirin on EBOV infectivity in vitro. EBOV was amplified in the presence of increasing concentrations of ribavirin, and plaque assays were used to determine the levels of infectious virus (measured as PFU per milliliter). Values shown represent the means for three biological replicates. Increasing concentrations of ribavirin resulted in decreased production of infectious virus. The IC50 was determined to be 27 μM. Ribavirin-induced cytotoxicity was not observed.
Effect of prophylactic ribavirin on Ebola virus infection in mouse model of EVD.
To determine the efficacy of ribavirin in a mouse model of EBOV infection, three groups of eight mice each were challenged with 1,000 PFU of mouse-adapted EBOV via the i.p. route as previously described (27). Two treatment groups received i.p. injections of either 50 or 80 mg/kg ribavirin 1 h prior to challenge and once daily thereafter; the third group received saline only as a vehicle control. All groups exhibited a decline in weight over the first week (Fig. 2). However, many of the treated mice gradually regained weight as the study progressed. Ribavirin treatment correlated with a statistically significant delay to death and increased survival compared to those of the untreated group (Fig. 3). The median day to death for the untreated group was 6 days, while the median day to death for mice treated with 80 mg/kg ribavirin was 12 days. The median day to death for the 50-mg/kg group was undefined because of the high survival rate. Treatment with 50 mg/kg ribavirin resulted in 75% survival, and treatment with 80 mg/kg resulted in 38% survival. While the 50-mg/kg group appeared to have a higher survival rate and less weight loss than the 80-mg/kg group, the difference was not statistically significant (P = 0.185; log-rank Mantel-Cox test).
FIG 2.
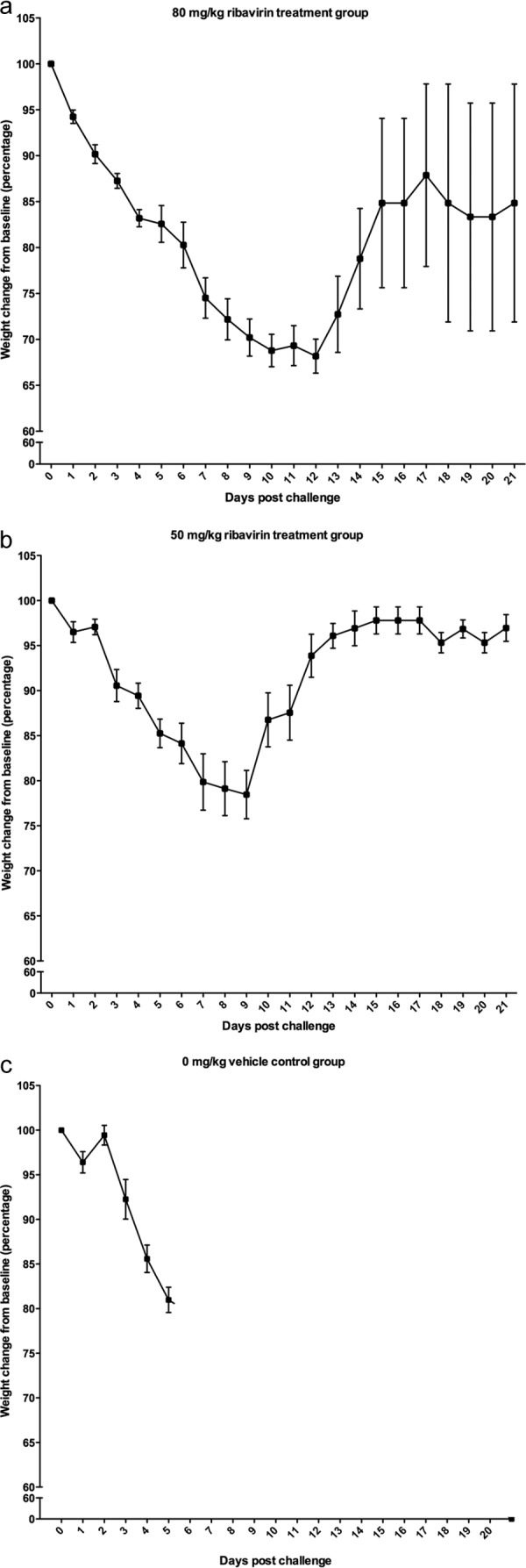
Weights of mice challenged with EBOV and prophylactically treated with ribavirin. Mice were infected with 1,000 PFU of mouse-adapted EBOV and treated with ribavirin 1 h prior to challenge and daily thereafter. Percent weight changes from baseline for each treatment group (80 mg/kg [a], 50 mg/kg, and 0 mg/kg [c]; n = 8 for each group) over the 21-day course of the study are shown. Means and standard errors of the means (SEM) are presented.
FIG 3.
Survival of mice challenged with EBOV and prophylactically treated with ribavirin. Mice were infected with 1,000 PFU of mouse-adapted EBOV and treated with ribavirin 1 h prior to challenge and daily thereafter (n = 8 for each group). Survival over the course of the study is displayed. The experiment was performed one time, and two graphs are shown in order to compare each treatment group to the one untreated group. Ribavirin treatment correlated with increased survival and a significant delay to death compared to the results for the untreated group (P = 0.0001 for the 80-mg/kg group [a] and P < 0.0001 for the 50-mg/kg group). The median day to death for the untreated group was 6 days, while the median day to death for animals treated with 80 mg/kg was 12 days. Treatment with 50 mg/kg resulted in 75% survival, and treatment with 80 mg/kg resulted in 38% survival. While the 50-mg/kg group appeared to have slightly better outcomes than those of the 80-mg/kg group, there was no statistically significant difference in survival between the two groups (P = 0.185).
Effect of postexposure ribavirin on Ebola virus infection in a mouse model.
To determine the effects of ribavirin treatment by use of a post-EBOV exposure model, five groups of eight mice each were challenged i.p. with 1,000 PFU of mouse-adapted EBOV. Four groups received various amounts of ribavirin (group 8, 30 mg/kg; group 7, 50 mg/kg; group 6, 100 mg/kg; and group 5, 100-mg/kg loading dose and 50-mg/kg subsequent doses), delivered i.p. at 24 h postchallenge and daily thereafter. Group 9 received saline as a vehicle control. To identify any detrimental effects of ribavirin treatment alone, four other groups of four mice each (groups 1, 2, 3, and 4) received an identical treatment regimen but were not infected with EBOV.
During infection, weight loss (Fig. 4) followed a pattern similar to that observed in the prophylactic study; all groups exhibited a decline in weight over the first week, but the survivors gradually regained weight as the study progressed. By day 7, all unchallenged groups exhibited significantly less weight loss than the corresponding challenged groups (Fig. 4) (P < 0.001; two-way ANOVA with the Bonferroni posttest). Postexposure ribavirin treatment correlated with increased survival and a significant delay in the median day to death (Fig. 5). The median day to death for untreated group 9 was 5 days, while the median day to death for treated mice was 7 days. The survival rate was 22%. There was no significant difference in survival or median day to death between the different treatment groups (P > 0.65; log-rank Mantel-Cox test) (Fig. 5). None of the unchallenged mice exhibited serious clinical signs, and all animals survived throughout the duration of the study. While ribavirin treatment was well tolerated in the unchallenged mice, multiple animals exhibited some weight loss (Fig. 4A) that may have been related to treatment or handling.
FIG 4.

Weights of mice challenged with EBOV and treated with ribavirin postexposure. Mice were infected with 1,000 PFU of mouse-adapted EBOV and treated with different doses of ribavirin at 1 day postchallenge and daily thereafter (n = 8 for each group). (a) Four groups (n = 4 for each group) received the same treatment regimen but were left uninfected. Percent weight changes from baseline for each treatment group (30 mg/kg [e], 50 mg/kg [d], 100 mg/kg [c], 100-mg/kg loading dose with 50-mg/kg subsequent doses, and 0 mg/kg [f]) over the 21-day course of the study are shown. All groups exhibited a decline in weight over the first week. As the study progressed, many of the treated animals gradually regained weight. Means and SEM are presented for challenged animals. Means and data for each replicate are presented for unchallenged animals.
FIG 5.

Survival of mice challenged with EBOV and treated with ribavirin postexposure. Mice were infected with 1,000 PFU of mouse-adapted EBOV and treated with different doses of ribavirin at 1 day postchallenge and daily thereafter (n = 8 for each group). Four groups (n = 4 for each group) received the same treatment regimen but were left uninfected. Survival over the course of the study is displayed for each treatment group compared to the untreated group. Postexposure ribavirin treatment correlated with increased survival and a significant delay in the median day to death (P = 0.004 for 30 mg/kg [a], P = 0.019 for 50 mg/kg, P = 0.0003 for 100 mg/kg [c], and P = 0.0003 for 100-mg/kg loading dose with 50-mg/kg subsequent doses [d]). The median day to death for the untreated group was 5 days, while the median day to death for treated animals was 7 days. The survival rate was 22%. There was no significant difference in survival or median day to death between the different treatment groups (P > 0.65).
Effect of postexposure ribavirin on Ebola virus replication in nonhuman primates.
To determine the effects of postexposure ribavirin treatment in an NHP model of EBOV infection, four cynomolgus macaques (Macaca fascicularis) were challenged i.m. with 100 PFU of passage 3 EBOV Kikwit, which has been shown to be 100% lethal at this dose (unpublished observations). One NHP received saline as a control, and each of the other three NHPs received twice-daily i.m. ribavirin treatment beginning at 24 h postinfection. During days 1 to 3, the treatment dose was 30 mg/kg, and on day 4, the dose was decreased to 15 mg/kg for the remainder of the study. Ribavirin treatment decreased the viral loads observed throughout the course of infection (Fig. 6a). Two-way ANOVA analysis revealed that on day 5 postchallenge and on the day of death for each animal, the serum titers for the treated NHPs were significantly lower than those for the untreated NHP (P < 0.001; two-way ANOVA with the Bonferroni posttest). Treatment also correlated with a delay in the time to death (Fig. 6b). The untreated NHP succumbed on day 6 postchallenge, while one treated NHP succumbed on day 7 and the remaining two treated NHPs survived until day 8. The median day to death for the untreated NHP was 6 days, while the median day to death for treated NHPs was 8 days, and the mean was 7.7 days.
FIG 6.

Effect of daily postexposure ribavirin treatment on EBOV-challenged M. fascicularis macaques. Four M. fascicularis macaques were challenged i.m. with 100 PFU of EBOV. Three NHPs received twice-daily i.m. ribavirin treatment beginning at 1 day postinfection, and one NHP received saline as a control. During days 1 to 3, the treatment dose was 30 mg/kg, and on day 4, the dose was decreased to 15 mg/kg for the remainder of the study. (a) Treated animals exhibited lower serum titers than the untreated animal throughout the course of infection. (b) Treatment correlated with a delay in the median day to death (P = 0.083). The median day to death for the untreated NHP was 6 days, while the median day to death for treated NHPs was 8 days. (c) Activated partial thromboplastin times (aPTT) for blood collected on days 0, 3, 5, and 7 and at euthanasia. (d) Alkaline phosphatase (ALP) levels in blood collected on days 0, 3, 5, and 7 and at euthanasia. (e) Gamma glutamyltransferase (GGT) levels in blood collected on days 0, 3, 5, and 7 and at euthanasia. (f) Partial thromboplastin (PT) times for blood collected on days 0, 3, 5, and 7 and at euthanasia.
Blood was collected on days 0, 3, 5, and 7 and at euthanasia for analysis of blood chemistry and coagulation times. All animals also exhibited increases in serum alkaline phosphatase (ALP) (Fig. 6d) and gamma glutamyltransferase (GGT) (Fig. 6e) near the time of death (consistent with previous observations of filovirus infection in experimentally infected NHPs [37, 38]). In contrast, the ribavirin-treated animals exhibited a delay in this increase compared to the untreated animal. Abnormal coagulation times were detected (Fig. 6c and f); all animals exhibited prolonged activated partial thromboplastin times (aPTT) near the day of death, but the treated animals exhibited a delay in the appearance of this symptom compared to the untreated animal. However, the untreated animal exhibited a very elevated partial thromboplastin (PT) time on day 5, but the treated animals never exhibited major changes in PT times.
To identify any effects of ribavirin treatment alone, four additional uninfected NHPs at ABSL2 received the same treatment regimen as the infected NHPs at ABSL4. No significant changes were seen in weight, no signs of illness were observed for the uninfected and treated animals, and all survived the duration of the study.
Mutagenic effect of guanosine analogue on Ebola virus in vivo.
Ultradeep sequencing was performed on sera collected from mice and NHPs to determine if ribavirin caused increased mutagenesis in vivo. For the preexposure mouse study, virus was harvested from sera taken immediately prior to humane euthanasia. In an untreated mouse at 5 dpi, the mutation frequency was below the limit of detection (3.0 × 108 nucleotides total, with a mean depth of coverage of 1.6 × 104). In a mouse treated with 50 mg/kg ribavirin, the mutation frequency at 7 dpi was 2.2 × 10−4 SNP per base sequenced (6.6 × 107 nucleotides total, with a mean depth of coverage of 3.8 × 103). In a mouse treated with 80 mg/kg ribavirin, the mutation frequency at 8 dpi was 4.1 × 10−3 SNP per base sequenced (2.2 × 106 nucleotides total, with a mean depth of coverage of 2.5 × 102). For the NHP study, sequencing was performed on virus harvested from sera taken at 5 dpi and immediately prior to humane euthanasia. In the untreated NHP at 5 dpi, the mutation frequency was 1.4 × 10−3 SNP per base sequenced (6.3 × 106 nucleotides total, with a mean depth of coverage of 2.7 × 103). In the treated NHPs at 5 dpi (n = 3), the average mutation frequency was 1.4 × 10−2 SNP per base sequenced (2.1 × 105 nucleotides total, with a mean depth of coverage of 3.4 × 102). This represented a 10-fold increase in mutation frequency associated with ribavirin treatment. In the untreated NHP at the time of death (6 dpi), the mutation frequency was relatively unchanged, at 1.1 × 10−3 SNP per base sequenced (2.2 × 108 nucleotides total, with a mean depth of coverage of 1.3 × 104). In the treated NHPs at the time of death (3, 7, or 8 dpi), the average mutation frequency was 2.8 × 10−3 SNP per base sequenced (8.9 × 107 nucleotides total, with a mean depth of coverage of 5.6 × 103), a 2.6-fold increase.
DISCUSSION
Ebola virus is an RNA virus that can cause hemorrhagic fever, with variable fatality rates of up to 90% (2). No approved vaccines or therapies exist for EBOV infections, despite great need. Typically, RNA viruses have high spontaneous mutation rates, but the mutation rate for EBOV is unknown. The data described herein highlight the potential for EBOV adaptation during an outbreak. Additionally, as therapeutic agents become available, the data will help us to understand the potential for the appearance of drug-resistant viruses.
We used a recombinant EBOV that carries the gene expressing GFP to investigate the mutation frequency of EBOV. This allowed quantification of errors accumulated during replication at a locus protected from selection pressures. The number of SNPs per nucleotide sequenced in the GFP region was high and similar to those for other RNA viruses (16, 33, 34). In contrast, the mutation frequency over wild-type regions of the genome was below the limit of detection for this assay. This implies that while the polymerase replicates with poor fidelity, there is a low tolerance for changes over the wild-type regions of the genome. It is likely that most mutant genomes were not packaged or the virions were not viable and therefore were not harvested for the sequencing assay.
The low tolerance for spontaneous changes suggests that EBOV may be susceptible to chemical mutagens. To this end, we investigated the effects of the guanosine analogue ribavirin on mutation frequency and virus replication. In vitro, ribavirin resulted in a substantial increase in the accumulation of SNPs across both the GFP and wild-type regions of eGFP-ZEBOV. This was associated with a >1,000-fold decrease in the production of infectious virus. The decrease in infectious virus production was also dose dependent, and the IC50 was 27 μM; this is similar to or lower than the IC50s observed for other RNA viruses that are susceptible to in vivo ribavirin treatment (39–41). Based on the mechanism of action against other RNA viruses (39), the decreased infectivity may be a result of error catastrophe from the increased mutation frequency.
Since in vitro susceptibility to ribavirin correlates with a therapeutic benefit in vivo for other RNA viruses, we next tested the effects of ribavirin in vivo in both mouse and NHP models of EBOV infection. First, the mouse model was used to test the effects of ribavirin treatment given pre-EBOV exposure. Mice were treated with ribavirin 60 min prior to receiving a lethal dose of EBOV and then received daily ribavirin treatments. Ribavirin treatment corresponded with 75% survival and a significantly delayed time to death for treated mice.
Next, mice and NHPs were challenged with a lethal dose of EBOV and then received daily ribavirin treatments starting at 24 h postchallenge. Ribavirin treatment corresponded with better survival outcomes for treated mice and NHPs. In mice, ribavirin treatment significantly delayed the time to death and increased survival. Ribavirin-treated NHPs exhibited a delayed time to death, from 6 days for the untreated control to a mean of 7.7 days for treated animals. In NHPs, ribavirin also decreased the viral load by 3.75 × 107 PFU/ml at 5 days postinfection. It is worth noting that the NHP study utilized M. fascicularis rather than Macaca mulatta macaques; M. mulatta macaques exhibit a slower disease progression (42) and thus may demonstrate greater efficacy of ribavirin treatment than what was seen for M. fascicularis.
The evidence in the literature regarding the activity of ribavirin against filoviruses in vitro is equivocal; some studies have observed no effect (19, 20), while others have observed some antiviral activity (21). In vivo studies have shown that ribavirin delayed the mean time to death in guinea pigs, but no effect was seen in NHPs (20, 43). Different experimental conditions and study designs may explain the discrepancies found in the literature and between our results and previous reports. Our results from the in vitro and preexposure in vivo experiments indicate that EBOV is susceptible to ribavirin. The deep-sequencing data further suggest that ribavirin may cause mutagenesis in the virus in vivo. Virus in sera from ribavirin-treated mice and NHPs showed a higher mutation frequency than that in sera from untreated animals. However, in NHPs, this difference in mutation frequency was more dramatic at 5 dpi than at 7 dpi. This may explain the lack of survival seen in the NHPs. Animals received a higher dose of ribavirin during the beginning of the study, so it is possible that the drop in dose caused the decline in mutagenesis.
However, it is possible that there is another therapeutic mechanism involved as well. For example, ribavirin probably exhibits indirect antiviral activity, such as cytokine modulation, against other viruses (39). Further work is needed to understand the mode(s) of action against EBOV and how to best exploit the mechanism. We speculate that the efficacy of ribavirin in both mice and NHPs may provide utility for human infections, particularly in combination therapy. For example, coupled with another therapy (interferon, for example, as used in combination with ribavirin for hepatitis C virus [HCV] therapy) or administered while a patient awaits experimental therapies (such as Zmapp [44]), the decrease in viral load should enhance the efficacy of other treatments and lead to improved outcomes; decreased viral loads have previously been associated with survival (45, 46). Inclusion of a mutagen, such as ribavirin, may also be beneficial when administering certain therapies, such as antibody cocktails (47), that may suffer from antiviral resistance. Furthermore, it is critical to consider that these studies were not performed in the context of supportive care and rehydration, which can be instrumental in improving outcomes for human infections (48). In this context, we speculate that ribavirin may provide sufficient time for the immune system to mount a protective response. Additionally, ribavirin is cost-effective, does not require a cold chain, and is often widely available in rural areas. Finally, newly developed treatments and countermeasures face rigorous testing before they can be used in the clinic, but ribavirin is already FDA approved for other viral infections, so it could promptly be applied to EBOV.
Our results suggest that EBOV exhibits a high mutation frequency and may be susceptible to ribavirin, and thus the use of ribavirin as a potential therapeutic warrants consideration.
ACKNOWLEDGMENTS
H. Feldmann kindly provided the recombinant Ebola virus containing GFP. We give special thanks to the following veterinary staff for their assistance with the in vivo challenge studies: Laura Rumpf, Matthew Stautzenberger, George Villanueva, Tony Bowers, Robert Geiger, Juan Zapata, Alberto Torres, Dave VanDenberg, Alex Limon, and Yenny Goez-Gazi.
This work was conducted in facilities constructed with support from the Research Facilities Improvement Program (grant number C06 RR012087) from the NCRR.
K.J.A. and A.G. designed and conducted experiments and wrote the paper. R.C. and G.W. designed experiments.
We declare that we have no competing interests.
Funding Statement
Internal funds were used for this research.
REFERENCES
- 1.Baize S, Pannetier D, Oestereich L, Rieger T, Koivogui L, Magassouba N, Soropogui B, Sow MS, Keïta S, De Clerck H, Tiffany A, Dominguez G, Loua M, Traoré A, Kolié M, Malano ER, Heleze E, Bocquin A, Mély S, Raoul H, Caro V, Cadar D, Gabriel M, Pahlmann M, Tappe D, Schmidt-Chanasit J, Impouma B, Diallo AK, Formenty P, Van Herp M, Günther S. 2014. Emergence of Zaire Ebola virus disease in Guinea. N Engl J Med 371:1418–1425. doi: 10.1056/NEJMoa1404505. [DOI] [PubMed] [Google Scholar]
- 2.Feldmann H, Sanchez A, Geisbert TW. 2013. Filoviridae: Marburg and Ebola viruses, Chapter 32, p 923–924. In Knipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed Lippincott Williams & Wilkins, Philadelphia, PA. [Google Scholar]
- 3.Sala M, Wain-Hobson S. 1999. Drift and conservatism in RNA virus evolution: are they adapting or merely changing, Chapter 6 In Domingo E, Webster R, Holland J (ed), Origin and evolution of viruses. Academic Press, San Diego, CA. [Google Scholar]
- 4.Rodriguez LL, De Roo A, Guimard Y, Trappier SG, Sanchez A, Bressler D, Williams AJ, Rowe AK, Bertolli J, Khan AS, Ksiazek TG, Peters CJ, Nichol ST. 1999. Persistence and genetic stability of Ebola virus during the outbreak in Kikwit, Democratic Republic of the Congo, 1995. J Infect Dis 179(Suppl 1):S170–S176. doi: 10.1086/514291. [DOI] [PubMed] [Google Scholar]
- 5.Hoenen T, Safronetz D, Groseth A, Wollenberg KR, Koita OA, Diarra B, Fall IS, Haidara FC, Diallo F, Sanogo M, Sarro YS, Kone A, Togo ACG, Traore A, Kodio M, Dosseh A, Rosenke K, de Wit E, Feldmann F, Ebihara H, Munster VJ, Zoon KC, Feldmann H, Sow S. 2015. Virology mutation rate and genotype variation of Ebola virus from Mali case sequences. Science 348:117–119. doi: 10.1126/science.aaa5646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gire SK, Goba A, Andersen KG, Sealfon RSG, Park DJ, Kanneh L, Jalloh S, Momoh M, Fullah M, Dudas G, Wohl S, Moses LM, Yozwiak NL, Winnicki S, Matranga CB, Malboeuf CM, Qu J, Gladden AD, Schaffner SF, Yang X, Jiang P-P, Nekoui M, Colubri A, Coomber MR, Fonnie M, Moigboi A, Gbakie M, Kamara FK, Tucker V, Konuwa E, Saffa S, Sellu J, Jalloh AA, Kovoma A, Koninga J, Mustapha I, Kargbo K, Foday M, Yillah M, Kanneh F, Robert W, Massally JLB, Chapman SB, Bochicchio J, Murphy C, Nusbaum C, Young S, Birren BW, Grant DS, Scheiffelin JS, Lander ES, Happi C, Gevao SM, Gnirke A, Rambaut A, Garry RF, Khan SH, Sabeti PC. 2014. Genomic surveillance elucidates Ebola virus origin and transmission during the 2014 outbreak. Science 345:1369–1372. doi: 10.1126/science.1259657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Castilletti C, Carletti F, Gruber CEM, Bordi L, Lalle E, Quartu S, Meschi S, Lapa D, Colavita F, Chiappini R, Mazzarelli A, Marsella P, Petrosillo N, Nicastri E, Chillemi G, Valentini A, Desideri A, Di Caro A, Ippolito G, Capobianchi MR. 2015. Molecular characterization of the first Ebola virus isolated in Italy, from a health care worker repatriated from Sierra Leone. Genome Announc 3:e00639–15. doi: 10.1128/genomeA.00639-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kugelman JR, Sanchez-Lockhart M, Andersen KG, Gire S, Park DJ, Sealfon R, Lin AE, Wohl S, Sabeti PC, Kuhn JH, Palacios GF. 2015. Evaluation of the potential impact of Ebola virus genomic drift on the efficacy of sequence-based candidate therapeutics. mBio 6:e02227–14. doi: 10.1128/mBio.02227-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Leroy EM, Kumulungui B, Pourrut X, Rouquet P, Hassanin A, Yaba P, Delicat A, Paweska JT, Gonzalez JP, Swanepoel R. 2005. Fruit bats as reservoirs of Ebola virus. Nature 438:575–576. doi: 10.1038/438575a. [DOI] [PubMed] [Google Scholar]
- 10.Marsh GA, Haining J, Robinson R, Foord A, Yamada M, Barr JA, Payne J, White J, Yu M, Bingham J, Rollin PE, Nichol ST, Wang L-F, Middleton D. 2011. Ebola Reston virus infection of pigs: clinical significance and transmission potential. J Infect Dis 204:S804–S809. doi: 10.1093/infdis/jir300. [DOI] [PubMed] [Google Scholar]
- 11.Swanepoel R, Leman PA, Burt FJ, Zachariades NA, Braack LE, Ksiazek TG, Rollin PE, Zaki SR, Peters CJ. 1996. Experimental inoculation of plants and animals with Ebola virus. Emerg Infect Dis 2:321–325. doi: 10.3201/eid0204.960407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Towner JS, Amman BR, Sealy TK, Carroll SAR, Comer JA, Kemp A, Swanepoel R, Paddock CD, Balinandi S, Khristova ML, Formenty PBH, Albarino CG, Miller DM, Reed ZD, Kayiwa JT, Mills JN, Cannon DL, Greer PW, Byaruhanga E, Farnon EC, Atimnedi P, Okware S, Katongole-Mbidde E, Downing R, Tappero JW, Zaki SR, Ksiazek TG, Nichol ST, Rollin PE. 2009. Isolation of genetically diverse Marburg viruses from Egyptian fruit bats. PLoS Pathog 5:e1000536. doi: 10.1371/journal.ppat.1000536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Martina BEE, Osterhaus ADME. 2009. “Filoviruses”: a real pandemic threat? EMBO Mol Med 1:10–18. doi: 10.1002/emmm.200900005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Miranda MEG, Miranda NLJ. 2011. Reston ebolavirus in humans and animals in the Philippines: a review. J Infect Dis 204:S757–S760. doi: 10.1093/infdis/jir296. [DOI] [PubMed] [Google Scholar]
- 15.Dowall SD, Matthews DA, Garcia-Dorival I, Taylor I, Kenny J, Hertz-Fowler C, Hall N, Corbin-Lickfett K, Empig C, Schlunegger K, Barr JN, Carroll MW, Hewson R, Hiscox JA. 2014. Elucidating variations in the nucleotide sequence of Ebola virus associated with increasing pathogenicity. Genome Biol 15:540. doi: 10.1186/s13059-014-0540-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Crotty S, Cameron CE, Andino R. 2001. RNA virus error catastrophe: direct molecular test by using ribavirin. Proc Natl Acad Sci U S A 98:6895–6900. doi: 10.1073/pnas.111085598. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Graci JD, Cameron CE. 2006. Mechanisms of action of ribavirin against distinct viruses. Rev Med Virol 16:37–48. doi: 10.1002/rmv.483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Crotty S, Maag D, Arnold JJ, Zhong WD, Lau JYN, Hong Z, Andino R, Cameron CE. 2000. The broad-spectrum antiviral ribonucleoside ribavirin is an RNA virus mutagen. Nat Med 6:1375–1379. doi: 10.1038/82191. [DOI] [PubMed] [Google Scholar]
- 19.Huggins J, Zhang ZX, Bray M. 1999. Antiviral drug therapy of filovirus infections: S-adenosylhomocysteine hydrolase inhibitors inhibit Ebola virus in vitro and in a lethal mouse model. J Infect Dis 179:S240–S247. doi: 10.1086/514316. [DOI] [PubMed] [Google Scholar]
- 20.Huggins JW. 1989. Prospects for treatment of viral hemorrhagic fevers with ribavirin, a broad-spectrum antiviral drug. Rev Infect Dis 11:S750–S761. doi: 10.1093/clinids/11.Supplement_4.S750. [DOI] [PubMed] [Google Scholar]
- 21.Ölschläger S, Neyts J, Günther S. 2011. Depletion of GTP pool is not the predominant mechanism by which ribavirin exerts its antiviral effect on Lassa virus. Antiviral Res 91:89–93. doi: 10.1016/j.antiviral.2011.05.006. [DOI] [PubMed] [Google Scholar]
- 22.Feld JJ, Hoofnagle JH. 2005. Mechanism of action of interferon and ribavirin in treatment of hepatitis C. Nature 436:967–972. doi: 10.1038/nature04082. [DOI] [PubMed] [Google Scholar]
- 23.Cuevas JM, González-Candelas F, Moya A, Sanjuán R. 2009. Effect of ribavirin on the mutation rate and spectrum of hepatitis C virus in vivo. J Virol 83:5760–5764. doi: 10.1128/JVI.00201-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Young K-C, Lindsay KL, Lee K-J, Liu W-C, He J-W, Milstein SL, Lai MMC. 2003. Identification of a ribavirin-resistant NS5B mutation of hepatitis C virus during ribavirin monotherapy. Hepatology 38:869–878. doi: 10.1002/hep.1840380413. [DOI] [PubMed] [Google Scholar]
- 25.Mardani M, Jahromi MK, Naieni KH, Zeinali M. 2003. The efficacy of oral ribavirin in the treatment of Crimean-Congo hemorrhagic fever in Iran. Clin Infect Dis 36:1613–1618. doi: 10.1086/375058. [DOI] [PubMed] [Google Scholar]
- 26.National Research Council. 2011. Guide for the care and use of laboratory animals, 8th ed National Academies Press, Washington, DC. [Google Scholar]
- 27.Sun Y, Carrion R Jr, Ye L, Wen Z, Ro Y-T, Brasky K, Ticer AE, Schwegler EE, Patterson JL, Compans RW, Yang C. 2009. Protection against lethal challenge by Ebola virus-like particles produced in insect cells. Virology 383:12–21. doi: 10.1016/j.virol.2008.09.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bray M, Davis K, Geisbert T, Schmaljohn C, Huggins J. 1998. A mouse model for evaluation of prophylaxis and therapy of Ebola hemorrhagic fever. J Infect Dis 178:651–661. doi: 10.1086/515386. [DOI] [PubMed] [Google Scholar]
- 29.Alfson KJ, Beadles MW, Griffiths A. 2014. A new approach to determining whole viral genomic sequences including termini using a single deep sequencing run. J Virol Methods 208:1–5. doi: 10.1016/j.jviromet.2014.07.023. [DOI] [PubMed] [Google Scholar]
- 30.Shurtleff A, Biggins J, Keeney A, Zumbrun E, Bloomfield H, Kuehne A, Audet J, Alfson K, Griffiths A, Olinger G, Bavari S. 2012. Standardization of the filovirus plaque assay for use in preclinical studies. Viruses 4:3511–3530. doi: 10.3390/v4123511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ebihara H, Theriault S, Neumann G, Alimonti JB, Geisbert JB, Hensley LE, Groseth A, Jones SM, Geisbert TW, Kawaoka Y, Feldmann H. 2007. In vitro and in vivo characterization of recombinant Ebola viruses expressing enhanced green fluorescent protein. J Infect Dis 196(Suppl 2):S313–S322. doi: 10.1086/520590. [DOI] [PubMed] [Google Scholar]
- 32.Sanjuán R, Nebot MR, Chirico N, Mansky LM. 2010. Viral mutation rates. J Virol 84:9733–9748. doi: 10.1128/JVI.00694-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Moreno H, Gallego I, Sevilla N, de la Torre JC, Domingo E, Martín V. 2011. Ribavirin can be mutagenic for arenaviruses. J Virol 85:7246–7255. doi: 10.1128/JVI.00614-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Airaksinen A, Pariente N, Menéndez-Arias L, Domingo E. 2003. Curing of foot-and-mouth disease virus from persistently infected cells by ribavirin involves enhanced mutagenesis. Virology 311:339–349. doi: 10.1016/S0042-6822(03)00144-2. [DOI] [PubMed] [Google Scholar]
- 35.Lanford RE, Chavez D, Guerra B, Lau JYN, Hong Z, Brasky KM, Beames B. 2001. Ribavirin induces error-prone replication of GB virus B in primary tamarin hepatocytes. J Virol 75:8074–8081. doi: 10.1128/JVI.75.17.8074-8081.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Barnard DL, Bischofberger N, Kim CU. 1997. Acyclic phosphonomethylether nucleoside inhibitors of respiratory viruses. Antivir Chem Chemother 8:223–233. doi: 10.1177/095632029700800306. [DOI] [Google Scholar]
- 37.Geisbert TW, Hensley LE, Larsen T, Young HA, Reed DS, Geisbert JB, Scott DP, Kagan E, Jahrling PB, Davis KJ. 2003. Pathogenesis of Ebola hemorrhagic fever in cynomolgus macaques: evidence that dendritic cells are early and sustained targets of infection. Am J Pathol 163:2347–2370. doi: 10.1016/S0002-9440(10)63591-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Carrion R, Ro Y, Hoosien K, Ticer A, Brasky K, de la Garza M, Mansfield K, Patterson JL. 2011. A small nonhuman primate model for filovirus-induced disease. Virology 420:117–124. doi: 10.1016/j.virol.2011.08.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tam RC, Lau JY, Hong Z. 2001. Mechanisms of action of ribavirin in antiviral therapies. Antivir Chem Chemother 12:261–272. doi: 10.1177/095632020101200501. [DOI] [PubMed] [Google Scholar]
- 40.Morfin F, Dupuis-Girod S, Frobert E, Mundweiler S, Carrington D, Sedlacek P, Bierings M, Cetkovsky P, Kroes ACM, van Tol MJD, Thouvenot D. 2009. Differential susceptibility of adenovirus clinical isolates to cidofovir and ribavirin is not related to species alone. Antivir Ther 14:55–61. [PubMed] [Google Scholar]
- 41.Rusnak JM. 2011. Experience with ribavirin for treatment and postexposure prophylaxis of hemorrhagic fever viruses: Crimean Congo hemorrhagic fever, Lassa fever, and hantaviruses. Appl Biosaf 16:67–87. [Google Scholar]
- 42.Bente D, Gren J, Strong JE, Feldmann H. 2009. Disease modeling for Ebola and Marburg viruses. Dis Model Mech 2:12–17. doi: 10.1242/dmm.000471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Ignat'ev GM, Strel'tsova MA, Agafonov AP, Kashentseva EA, Prozorovskiĭ NS. 1996. Experimental study of possible treatment of Marburg hemorrhagic fever with desferal, ribavirin, and homologous interferon. Vopr Virusol 41:206–209. [PubMed] [Google Scholar]
- 44.Qiu X, Wong G, Audet J, Bello A, Fernando L, Alimonti JB, Fausther-Bovendo H, Wei H, Aviles J, Hiatt E, Johnson A, Morton J, Swope K, Bohorov O, Bohorova N, Goodman C, Do Kim Pauly MH, Velasco J, Pettitt J, Olinger GG, Whaley K, Xu B, Strong JE, Zeitlin L, Kobinger GP. 2014. Reversion of advanced Ebola virus disease in nonhuman primates with ZMapp. Nature 514:47–53. doi: 10.1038/nature13777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sanchez A, Lukwiya M, Bausch D, Mahanty S, Sanchez AJ, Wagoner KD, Rollin PE. 2004. Analysis of human peripheral blood samples from fatal and nonfatal cases of Ebola (Sudan) hemorrhagic fever: cellular responses, virus load, and nitric oxide levels. J Virol 78:10370–10377. doi: 10.1128/JVI.78.19.10370-10377.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Towner JS, Rollin PE, Bausch DG, Sanchez A, Crary SM, Vincent M, Lee WF, Spiropoulou CF, Ksiazek TG, Lukwiya M, Kaducu F, Downing R, Nichol ST. 2004. Rapid diagnosis of Ebola hemorrhagic fever by reverse transcription-PCR in an outbreak setting and assessment of patient viral load as a predictor of outcome. J Virol 78:4330–4341. doi: 10.1128/JVI.78.8.4330-4341.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kugelman JR, Kugelman-Tonos J, Ladner JT, Pettit J, Keeton CM, Nagle ER, Garcia KY, Froude JW, Kuehne AI, Kuhn JH, Bavari S, Zeitlin L, Dye JM, Olinger GG, Sanchez-Lockhart M, Palacios GF. 2015. Emergence of Ebola virus escape variants in infected nonhuman primates treated with the MB-003 antibody cocktail. Cell Rep 12:2111–2120. doi: 10.1016/j.celrep.2015.08.038. [DOI] [PubMed] [Google Scholar]
- 48.Fowler RA, Fletcher T, Fischer WA, Lamontagne F, Jacob S, Brett-Major D, Lawler JV, Jacquerioz FA, Houlihan C, O'Dempsey T, Ferri M, Adachi T, Lamah M-C, Bah EI, Mayet T, Schieffelin J, McLellan SL, Senga M, Kato Y, Clement C, Mardel S, Vallenas Bejar De Villar RC, Shindo N, Bausch D. 2014. Caring for critically ill patients with Ebola virus disease. Perspectives from West Africa. Am J Respir Crit Care Med 190:733–737. doi: 10.1164/rccm.201408-1514CP. [DOI] [PubMed] [Google Scholar]