

Abstract
The MIS pathway is a potential therapeutic target in epithelial ovarian cancer (EOC): signaling requires both type II (T2R) and type I receptors (T1R), and results in growth inhibition. MISR2 is expressed in EOC, but the prevalence and relative contributions of candidate T1R remain unknown. We sought to: a) determine expression of T1R in EOC; b) assess impact of T1R expression with clinical outcomes; c) verify MIS-dependent Smad signaling and growth inhibition in primary EOC cell cultures.
Tissue microarrays (TMA) were developed for analysis of T1Rs (ALK2/3/6) and MISR2 expression. Primary cell cultures were initiated from ascites harvested at surgery which were used to characterize response to MIS.
TMA’s from 311 primary cancers demonstrated the most common receptor combinations were: MISR2+/ALK2+3+6+ (36%); MISR2+/ALK2+3+6- (34%); MISR2-/ALK2+3+6- (18%); and MISR2-/ALK2+3+6+ (6.8%). No differences in overall survival (OS) were noted between combinations. The ALK6 receptor was least often expressed T1R and was associated with lower OS in early stage disease only (p =0.03). Most primary cell cultures expressed MISR2 (14/22 (63.6%)): 95% of these express ALK 2 and ALK3, whereas 54.5% expressed ALK6. MIS-dependent Smad phosphorylation was seen in the majority of cultures (75%). Treatment with MIS led to reduced cell viability at an average of 71% (range: 57–87%) in primary cultures. MIS signaling is dependent upon the presence of both MISR2 and specific T1R. In the majority of EOC, the T1R required for MIS-dependent signaling are present and such cells demonstrate appropriate response to MIS.
Keywords: ALK2, ALK3, ALK6, müllerian inhibiting substance, ovarian cancer
INTRODUCTION
Epithelial ovarian cancer (EOC) accounts for the majority of mortality in gynecologic cancer with 50% of female genital cancer deaths in 2010 originating from EOC [1, 2]. Obstacles to improving outcomes include: advanced stage of disease at the time of presentation, dose-limiting chemotherapy toxicity to normal tissues, and eventual disease recurrence often marked by resistance to cytotoxic drugs. These challenges have motivated research for targeted drug therapies that can block or enhance cellular pathways as a means for constraining cancer cell growth. One potential target is the Müllerian inhibiting substance (MIS) pathway. This pathway plays a key role in fetal development during müllerian duct regression in males. The receptors for this pathway are also expressed in a tissue specific manner in müllerian derived tissues. These observations have led to the concept that MIS and/or MIS signaling pathway could potentially serve as a therapeutic, tissue-specific target in the treatment of EOC.
MIS receptors are naturally occurring in human müllerian-derived cells and their activation by MIS secreted by Sertoli cells in the male fetus leads to apoptosis and regression of müllerian structures [3]. The receptors are part of the TGF-β superfamily and function as a complex of two transmembrane serine/threonine kinase proteins. MIS (ligand) binds to the Type II receptor (T2R, MISR2) and it, in turn, recruits and complexes with one of at least three candidate Type I receptors (T1R: ALK2, ALK3, or ALK6) [4, 5]. Subsequently, the relevant T1R mediates downstream signaling by phosphorylating Smad 1/5/8 proteins, which then binds the Smad4 coactivator to form one unit that translocates into the nucleus to effect downstream growth inhibition [6–8]. Behringer et al. showed that female mice chronically exposed to MIS had undetectable ovaries in adulthood due to specific activation of the MISR2 signaling pathway [6]. This work suggests that ovarian tissue is responsive to MIS and numerous investigations support that MIS signaling can also inhibit EOC cell growth [9].
Based on natural ability of MIS to inhibit growth of müllerian derived tissues, MIS is actively being studied as a potential drug to treat EOC. Fuller et al. [10] initially demonstrated that MIS inhibited colony formation in 25 out of 28 patient-derived tumor specimens in soft agar colony formation assay. Early investigations noted increased activity with increasing purification of MIS and consistent results for individual patient samples harvested at different times. Subsequent studies utilizing immunopurified rhMIS showed that MIS causes significant inhibition in a number of human cell lines in colony formation assay both in vitro and in vivo [11]. Exposure of human ovarian cancer cell lines and mouse ovarian cancer models to recombinant human MIS (rhMIS) results in significant growth inhibition both in vitro and in vivo [9]. Requirement of MIS-RII receptors for MIS mediated suppression was confirmed by transgenic expression of MISRII in mouse ovarian carcinoma (MOVCAR) cell lines [9]. MIS significantly suppressed growth of MISRII expressing MOVCAR cell line both in vitro and in vivo. These observations were reinforced by Pieretti-Vanmarcke et al. using mouse lines of EOC. Additionally, rhMIS when used in combination with subclinical concentrations of traditional cytotoxic drugs in vitro and in vivo enhanced response and efficacy of therapy [12]. Interestingly, in some cancer lines and combinations competitive effects between rhMIS and drug therapy were observed. These latter observations suggest a complex relationship possibly related to the presence or absence of MIS signaling components which yield different results depending on expression combinations or cell background. Importantly, all of these studies were limited by lack of detailed characterization of MIS receptor (type I or II) expression patterns to correlate with response.
Finally, additional relevance for MIS therapy comes from recent studies from the Donahoe’s laboratory demonstrating that MIS may preferentially inhibit stem/progenitor cells [13] as well as decrease invasion and migration in human ovarian cancer cell lines [14]. This potential increased efficacy of a stem-like cell population in EOC could have significant implications for the therapeutic value of rhMIS. Together, these data indicate that: most ovarian cancer respond to MIS; MIS can inhibit growth of ovarian cancer cells in vitro and in vivo; the response to MIS is currently not reliably predictable in combination with cytotoxic drugs.
Important obstacles in assessing the MIS pathway as a target in EOC include: i) lack of research on the role of the specific T1R necessary for response to MIS; ii) lack of data reporting the prevalence of T1R expression in EOC or their impact on tumor biology; iii) limitations in the large scale production of MIS. The scientific and MIS research community is actively addressing the latter point. Our lab has previously shown that the majority of primary EOC express MISR2 [15], however there is little data concerning T1R expression in gynecologic cancers. Studies from other TGF-β receptor family members demonstrate that signaling and response are dependent upon the specific T1R and lend important insights into the need for such characterization. Weiser et al., showed impaired TGF-β-dependent signaling with mutated T1R proteins [16]. Nohno et al. reported that different receptor combinations have different signaling potentials [17]. Sedes et al. have recently identified novel target genes using an innovative siRNA strategy to decipher relative contributions from specific candidate type I receptors in a granulosa cell background [18]. Additional studies are needed in EOC to select cancers most likely to respond in the anticipated manner. Lack of such studies in EOC hinders our understanding of MIS signaling and limits our ability to manipulate this system for therapeutic gain. To begin to address these limitations in MIS research, our initial objectives were to: a) determine expression patterns of candidate T1R (specifically, ALK2, 3 and 6) in primary EOC; b) evaluate clinical outcomes based upon expression of MISR2/T1R combinations; c) test the ability of MIS to induce Smad-dependent signaling and cell inhibition in primary cell cultures expressing MISR2 and T1R.
MATERIALS AND METHODS
Patient Tumor Selection and Clinical Data
Approval was obtained from the Mayo Clinic Institutional Review Board prior to data abstraction and TMA construction. Patients undergoing surgical management of epithelial ovarian cancer from 1999 to 2007 at Mayo Clinic were prospectively consented for collection of tumor and outcomes data (age, FIGO stage, grade, residual disease, chemotherapy, recurrence and death). Inclusion criteria included primary surgery for EOC (eligible subtypes: serous, endometrioid, clear cell, mucinous, and mixed epithelial). Histologic designation followed the classification of the World Health Organization and all cases were reviewed by a single gynecologic pathologist. Residual disease was classified as either no gross residual, gross disease with < 1cm, or > 1cm residual disease. For the current study we defined platinum resistant cancers as those with recurrent disease within 12 months of completion of chemotherapy; otherwise, tumors were deemed to be platinum sensitive. Patients who died prior to completing chemotherapy and patients who were NED but lacked 1 year of follow-up could not be categorized into a platinum response group. Disease stage was dichotomized into early (I, II, IIIA, and IIIB) or advanced (IIIC and IV).
Patient Ascites Collection
Mayo Clinic IRB approval was obtained for the use of human samples. A total of 22 ascites cases were obtained from the Ovarian Cancer Tumor Bank of Mayo Clinic. Ascitic fluid specimens (200–900ml) were collected aseptically in a vacuum flask at the time of initial surgical exploration for ovarian cancer. Samples were collected prior to cytoreductive surgery to minimize contamination with blood, and heparin was added at 1000 U/L to the ascitic fluid. Patients did not receive any chemotherapy prior to the surgery.
Primary Cell Cultures from Ovarian Cancer Ascites
Cancer cells were isolated within 4 h of collection of ascites from primary surgical patients. Ascitic fluid was centrifuged at 800 x g for 10 min to pellet the cellular component and lysed with ACK lysis buffer if RBC was present. Finally, the pellet was suspended in DMEM-F12 50/50 medium supplemented with insulin (0.25 U/ml); antibiotics with 20% MIS-free female FCS. Short-term cultures were grown for 5–7 days and media was changed every second day. Adherent cells were harvested and tested to confirm epithelial cell origin by staining with monoclonal anti-pan cytokeratin and/or anti-CD46 antibodies. Adherent cells were harvested for collection of mRNA and protein lysates for expression analysis, or expanded for subsequent studies and frozen at low passage number.
Ovarian Cancer Cell Lines
Established human ovarian carcinoma cell lines SKOV3 and Z3 (SKOV3 cell line stably expressing MISR2) as described previously by our lab were used [15]. All cells were grown in 10% RPMI medium supplemented with female fetal calf serum (Aries Biologicals, Richardson, TX) penicillin, streptomycin and L.-glutamine in 5% CO2 at 37°C. Selection antibiotic G418 (400 μg/ml) was added to the Z3 cells for the maintenance of MIS type II receptor plasmid.
Antibodies & rhMIS Ligand
The monoclonal anti-MISR2 mouse antibody (12G4mab) was kindly provided by Isabelle Navarro-Teulon (Cancer Research Institute, Montpellier Cedex 5, France). ALK2 receptor (Activin r, R&D systems), ALK3 receptor (BMPR-IA, Santa Cruz Biotech, CA), and ALK6 receptor (BMPR-IB, Santa Cruz Biotech, CA). Anti-Gapdh and anti-pSmad1/5/8 antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz Biotech, CA) and Cell Signaling Technology (Danvers, MA), respectively. Recombinant human MIS (rhMIS) ligand was kindly provided by Donahoe & MacLaughlin Laboratory (Harvard-MIT Division of Health Sciences and Technology Simches Research Center, Boston, MA). Briefly, rhMIS was produced in transfected Chinese hamster ovary cells and purified as described previously, and its bioactivity was assessed in a MIS-specific organ culture for its ability to cause Müllerian duct regression [19, 20].
Construction of Tissue Microarrays (TMA)
TMAs were created from formalin-fixed, paraffin-embedded tumors. All participants provided written informed consent for an IRB-approved protocol. We used an automated Beecher Instruments ATA-27 arrayer following gynecologic pathologist review indicating tumor location. Three 0.6-mm cores were removed from each case paraffin block and placed in a recipient paraffin block according to a randomized electronic TMA map. Recipient blocks were sliced into 5-μm sections and mounted on charged slides.
Immunohistochemistry Analysis and Digital Imaging
TMAs were stained with antibodies to four proteins of interest: 12G4mab, ALK2, ALK3 and ALK6. TMAs were deparaffinized with three changes of xylene and rehydrated in a series of alcohols (100%, 95%, and 70% EtOH) and rinsed well in running distilled water. Slides were then placed in a preheated 1 mM EDTA, pH 8.0 retrieval buffer for 30 min then cooled in the buffer for 5 minutes followed by a 5 minute rinse in running distilled water. After heat inactivated epitope retrieval, the antibodies were stained with their respective staining protocols. Slides stained for ALK6 and MISR2 were placed on the DAKO Autostainer and underwent processing as previously described [15]. The method for the ALK6 was similar in nature with the secondary detection changed to the Goat HRP kit (Ca. No. GHP516, Biocare, Concord, CA). For the remaining antibodies, the method for staining ALK3 and ALK2 was similar to the ALK6 with an increase in incubation time to overnight in a humidified staining chamber for the primary antibody and the secondary antibodies with Envision Dual Link (Ca. No. K4061, Dako, Carpenteria, CA).
Following their respective staining protocols, all slides were then rinsed prior to counterstaining with a Modified Schmidts' Hematoxylin for 5 minutes. Slides were then rinsed in running tap water for three minutes to blue sections, dehydrated through graded alcohols, cleared in three changes of xylene and mounted with a Leica automated coverslipper.
Digital images of each core were captured and stored using a high resolution microscope (Zeiss Axioplan, CA) and video camera interfaced with Microsoft access software to correlate each tissue core with the appropriate patient identifier. Digital images of the stained specimens were reviewed and scored independently by two reviewers, blinded to clinical information. We observed a scoring discrepancy of <3%. These inconsistencies were resolved when the two physicians, together, reviewed the images and decided on one final score. Slides were evaluated for staining intensity (0 = no staining, to 3+ = strong staining), and percentage of tumor cells stained. Scores of 0 and 1+ were categorized as “negative” for protein expression while scores of 2+ and 3+ were considered “positive” for expression. Epithelial staining considered positive had to involve at least 25% of the cells in 2 out of 3 cores to be deemed evaluable.
Statistical Analysis
Data are summarized using standard descriptive statistics: frequency and percentage for categorical variables; and mean, standard deviation (SD), median, and interquartile range (IQR) for continuous variables. Duration of follow-up was calculated from the date of surgery to the date of death or last follow-up. The outcomes of interest were death due to any cause and recurrence. Overall survival and survival-free of recurrence were estimated using Kaplan-Meier method. Associations were evaluated based on fitting Cox proportional hazards models and summarized using the hazard ratio (HR) and corresponding 95% confidence interval (CI). All calculated p-values were two-sided and p-values less than 0.05 were considered statistically significant. Statistical analyses were performed using the SAS version 9.2 software package (SAS Institute, Inc., Cary, NC).
Immunofluorescence Microscopy
Primary cell cultures were trypsinized, harvested, washed and then plated onto 8 well glass chamber slides (Thermo Fisher Scientific, Pittsburgh, PA) for processing. After 12 hour in culture media, cells were fixed for 10 min at room temperature with 4% formalin (Sigma-Aldrich, St. Louis, MO) and washed three times with PBS. The cells were blocked with 3% BSA in PBS for 30 min at 4°C and stained with the appropriate primary antibody (anti-MISR2 ab; 12G4, cytokeratin) overnight at 4°C. Unbound primary antibodies were removed by washing (4 X 5 min) followed by incubation with a fluorescent labeled secondary antibody for 1 h. Unbound secondary antibodies were removed by washing (4 X 5 min) in PBS. Coverslips were mounted using Vectorshield with DAPI for nuclear staining (Vectashield, Vector Laboratories, Burlingame, CA). Cells were imaged with an LSM 510 Confocal Microscope (Carl Zeiss Micro Imaging, Thornwood, NY).
RNA Extraction and RT-PCR
RNA was prepared using the RNeasy kit (Qiagen Inc., Valencia, CA) and was treated with DNase to remove contaminating genomic DNA, using RNase-free DNaseI set (Qiagen Inc., Valencia, CA). A total of 900 ng of RNA was reverse-transcribed with random hexamers in a 40 μL reaction using SuperScript III (Invitrogen Corp, Carlsbad, CA). Primers were synthesized for MISR2 and GapDh by SA Bioscience (Qiagen, MD), and primers for Alk2 (forward: 5′-CAGCTTGATGTGGGGAAC-3′ and reverse: 5′-GACT CGAGCGGCCGCTCAACAGTCAC-3′), Alk3 (forward: 5′-GCTCTATTTGATTACTGATTACCATG-3′ and reverse: 5′-TCCACCGATTAGACACAATTGGC-3′) and Alk6 (forward: 5′-CTCAGGGAGCGACCTGGGCA-3′ and reverse: 5′-GCGGCCCCAAATGCAGGGAT-3′) were synthesized by IDT (Coralville, Iowa). PCR was performed using 25-μl volume, containing 200 ng of cDNA, 1 unit of AmpliTaq DNA polymerase, PCR buffer, dNTPs (Applied Biosystems, Foster City, CA) and 10 pmole/μl primers, with appropriate controls. The cDNA was amplified in Bio-Rad iCycler using the following protocol: 95°C for 3 min; 35 cycles of 95°C for 30 s, 50°C for 30 s, and 72°C for 45 s; with a final step at 72°C for 7 min. PCR product of Alk2 (450bp), Alk3 (520bp), Alk6 (540bp), and MISR2 (120bp) were detected using 2% agarose gel. Gapdh was used as a housekeeping gene. Bands were visualized using G:BOX gel imager from Syngene (Frederick, MD).
Western Blotting
Cells were harvested and washed twice with ice-cold PBS. Whole cell extracts were prepared in the lysis buffer (20 mM Tris-HCl, pH 7.5, 150 mM NaCl, 1 mM Na2EDTA, 1 mM EGTA, 1% Triton) containing complete protease inhibitor cocktail (Roche Applied Science, Indianapolis, IN). Equal amounts of proteins (30ug) were loaded in each lane and separated by 10% SDS-PAGE under non-reducing conditions. Proteins were transferred to PVDF membranes (Bio-Rad, Hercules, CA) by electroblotting, membranes were blocked in 5% non-fat milk in TBS-T buffer. Membranes were incubated with primary antibodies overnight at 4°C, washed and incubated with secondary antibodies for 1 hour at room temperature. Proteins were visualized by enhanced chemiluminescence (ECL plus Western Blotting Detection System; Amersham Biosciences, Piscataway, NJ).
Smad Phosphorylation
Primary cell cultures or Z3 cells were plated at 3x105 cells in 6 well plates. Cells were pre-treated by incubation in serum-free DMEM medium overnight, followed by stimulation in the presence or absence of 25ng/ml BMP4 or 15 μg/ml MIS ligand for 4 hours at 37°C. Whole cell lysates were prepared in the following buffer 50 mM Tris-HCL pH 7.4, 1% Triton-X100, 0.25% DOC, 150 mM NaCl, 1 mM EDTA, 10 mM NaF and complete protease inhibitor cocktail. Membranes were created as described using 50 μg protein. Membranes were probed primarily with anti-phospho-Smad 1/5/8 antibodies (1:1000 dilution) and secondarily with anti-rabbit IgG HRP antibody (1:10,000 dilution). Proteins were visualized as described above in Immunoblotting methods.
Cell Viability Assay
The cell viability assay was measured using the 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide dye (MTT) reduction assay which was performed to determine the cell inhibition effect of the MIS ligand at various concentrations. Briefly, primary cell cultures were harvested, counted and re-plated in 96-well plates at 5000 cells/well in 200 μl media. After 24 h, cells were treated in presence or absence of MIS (30μg/ml). After 72 h of incubation, the remaining living or surviving cells were quantified as a correlative measure of proliferation by adding 20ul MTT solution (MTT One Solution Cell Proliferation Assay, Promega, Madison, WI) to the wells according to the manufacturer’s protocol. Absorbance was recorded at 550nm on a Spectra Max 190 (Molecular Devices, CA).
RESULTS
MIS Type II and Type I Expression in Primary Epithelial Ovarian Cancers
Tumor samples from 311 patients with EOC were evaluated with IHC staining of TMAs (Fig. 1). A summary of clinical variables including histology, stage, grade, debulking status, and platinum sensitivity is presented in Table 1. Forty-nine cases had excessive drop-out of tumor content across all tissue cores, and were excluded leaving 262 patients for analysis. In 10% of cases overall, IHC staining for at least one of the receptors was non-diagnostic. The frequency of MIS receptor expression is summarized in Table 2. When present, MISR2 was always complexed with at least one candidate T1R. Notably, several of the candidate T1R were expressed in nearly all cases; a single T1R was expressed in only 7 cases. MISR2 was not expressed in 27% (67/252) of cases.
Fig. (1).

Immunohistochemical staining of tissue microarray of epithelial ovarian cancer samples. Representative images depict negative and positive staining for ALK2, ALK3, ALK6 and MISR2 in a TMA core. TMAs were created from formalin-fixed, paraffin-embedded tumors and stained with antibodies to four proteins of interest:, ALK2, ALK3, ALK6 and 12G4mab. Digital images of each core were captured and stored using a high resolution microscope (Zeiss Axioplan, CA) and video camera interfaced with Microsoft access software to correlate each tissue core with the appropriate patient identifier. Digital images of the stained specimens were reviewed and scored independently by two reviewers, blinded to clinical information.
Table 1.
Demographic and clinical characteristics.
| Characteristic | Total (N=262) |
|---|---|
| Age at diagnosis (years) | |
| Mean (SD) | 61.0 (12.3) |
| Median | 61.0 |
| Range | (28 – 93) |
| FIGO Stage, n (%) | |
| I or II | 66 (25.2%) |
| IIIA or IIIB | 12 (4.6%) |
| IIIC | 140 (53.4%) |
| IV | 44 (16.8%) |
| Grade, n (%) | |
| 1 | 13 (5.2%) |
| 2 | 25 (9.9%) |
| 3 | 127 (50.4%) |
| 4 | 87 (34.5%) |
| Not documented | 10 |
| Histology, n (%) | |
| Clear cell | 15 (5.7%) |
| Endometrioid | 47 (17.9%) |
| Mucinous | 10 (3.8%) |
| Serous | 171 (65.3%) |
| Mixed Epithelial | 19 (7.3%) |
| Debulking status, n (%) | |
| No residual | 112 (43.9%) |
| Optimal (1– 10cm) | 110 (43.1%) |
| Sub-optimal (>10cm) | 33 (12.9%) |
| Unknown | 7 |
| Chemo-sensitivity, n (%) | |
| Sensitive | 185 (79.7%) |
| Resistant | 47 (20.3%) |
| Indeterminate † | 30 |
The chemo-sensitivity status was indeterminate for 30 patients who either died prior to completion of chemotherapy (n=23) or had less than one year of follow-up with no documented recurrence (n=2 death, n=5 lost to follow-up).
Table 2.
Receptor prevalence.
| Receptors | Total (N) |
|---|---|
| Presence of Type I receptors, n (%) | |
| ALK2 | 241/242 (99.6%) |
| ALK3 | 240/251 (95.6%) |
| ALK6 | 111/249 (44.6%) |
| Presence of Type II receptor, n (%) † | 185/252 (73.4%) |
| Combinations | |
| MISR2, ALK2, ALK3, ALK6 | 84/235 (36%) |
| MISR2, ALK2, and ALK3 | 81/235 (34%) |
| ALK2 and ALK3 | 43/235 (18%) |
| ALK2, ALK3, and ALK6 | 16/235 (6.8%) |
| MISR2 and ALK2 | 6 (2.5%) |
| MISR2, ALK2, and ALK6 | 4 (1.7%) |
| ALK2 present only | 1 (0.4%) |
| At least one of the 4 receptors was non-diagnostic | 27 (11.4%) |
Percentage of the patients with a diagnostic result for each specific receptor.
Among the 262 patients, 145 were dead, with a median time to death of 26.8 months (Interquartile range (IQR): 12.9 – 41.4 months) following the surgery. Of the remaining 117 who were alive at last contact, the median duration of follow-up was 49.2 months (IQR, 33.1 – 75.4 months). The overall survival was 86.5%, 74.3%, and 43.7% at 12, 24, and 60 months following surgery. Among 195 patients who experienced a remission, 87 developed a recurrence with a median time of 20.3 months (IQR, 14.4 – 29.1 months). The survival free of recurrence was 98.4%, 68.5%, and 46.4% at 12, 24, and 60 months.
Among the patients who had adequate tissue cores for evaluation of all four receptors (n=235) the most frequently observed receptor combinations were: MISR2 with ALK2/ALK3/ALK6 (84/235, 36%); MISR2 with ALK2/ALK3 (81/235, 34%); ALK2/ALK3 (43/235, 18%); and ALK2/ALK3/ALK6 (16/235, 7%). There was no difference in the frequency of the four major receptor combinations between early and advanced stage disease. There was no correlation between receptor combination and histology (p=1.0), grade (p=0.62), or age (p=1.0). All patients received platinum-based chemotherapy and 80% were classified as platinum-sensitive: platinum sensitivity was not correlated with receptor expression (p = 0.3). There was no statistically significant difference in overall or disease free survival between the four receptor combinations (p = 0.91 and p = 0.89, respectively).
Overall, 73.4% of cases expressed MISR2. Presence of MISR2 was associated with favorable surgical-pathologic findings. MISR2 negative cases were more likely to be advanced stage relative to MISR2 positive cases (67.6% vs 80.6%, p = 0.04) and more likely to have visible disease at the completion of primary debulking (52% vs 69.7%, p = 0.013). Despite these findings, MISR2 status was not significantly associated with time to recurrence (p = 0.84); further, the overall survival was not different for MISR2 expressing cancers (p = 0.47). Survival relationships were unchanged when the cohort was restricted to advanced stage disease and stratified by debulking status.
Since ALK6 was rarely expressed, we assessed its impact on survival. We observed a significant overall survival benefit in ALK6 non-expressing cancers for early stage disease (p = 0.03) but not in advanced stage cases (p=0.42) (Fig. 2). Patients with tumors expressing ALK6 were 3.2 times more likely to die than patients without ALK6 expression (95% CI 1.1–9.6).
Fig. (2).

ALK6 expression is associated with decreased survival in early stage EOCs. (A), Kaplan-Meier overall survival curves for ALK6 positive and negative early stage EOCs. Among patients with early stage disease, presence of the ALK 6 receptor was associated with a significantly increased risk of death (p = 0.03). Patients with tumors expressing ALK6 (n = 35) were 3.2 times more likely to die than patients without ALK6 (n = 38) expression (95% CI 1.1–9.6). (B) Among patients with advanced stage disease, expression of ALK 6 receptor did not affect overall survival. Associations were evaluated based on fitting Cox proportional hazards models and summarized using the hazard ratio (HR) and corresponding 95% confidence interval (CI).
Expression Pattern of T1R & MISR2 at mRNA and Protein Levels in Primary Cell Cultures
Cell surface expression of MISR2 was determined by immunoflourescence staining using 12G4 antibody in Z3 (positive control), SKOV3 (negative control) cell lines and primary cultures derived from 22 ovarian cancer cases (Fig. 3). Demographics for the 22 cases from which primary cell cultures were obtained including histology, stage, grade and debulking status is presented in Table 3. We confirmed expression of MISR2 and T1R mRNA and protein (representative data shown in Fig. 4A, B). All 22 cases expressed ALK2 and ALK3 mRNA whereas only 15 samples (68.2%) were positive for ALK6. Similar to IHC analysis, 14 out of 22 (63.6%) expressed MISR2. MISR2 protein expression was observed in 64% (14/22) and ALK2 and ALK3 protein was expressed in 95% (21/22) with ALK6 protein observed in 59% of cases. These results are consistent with the IHC analysis of primary cancers and demonstrate that the majority of ovarian cancers express MISR2 and the candidate T1R: additionally expression appears to be retained in primary cultures.
Fig. (3).

MISR2 is expressed in primary EOC cultures. Ovarian cancer cells were examined by immunofluorescence by staining with 12G4 primary antibody and Texas Red secondary antibody (A/B: left panels), or FITC-cytokeratin (A/B: right panels), as described in methods. Representative primary ovarian cancer cells are shown in A, and MISR2 overexpressing Z3 cells (positive controls) are shown in panel B. Stained cells were imaged with an LSM 510 Confocal Microscope (Carl Zeiss Micro Imaging, Thornwood, NY).
Table 3.
Clinical characteristics and receptor prevalence.
| Characteristics and Presence of Receptors | Total (N=22) |
|---|---|
| FIGO Stage, n (%) | |
| I | 3 (13.6%) |
| III | 17 (77.3%) |
| IV | 2 (9.09%) |
| Grade, n (%) | |
| 2 | 2 (9.09%) |
| 3 | 10 (45.45%) |
| 4 | 7 (31.81%) |
| Not documented | 3 |
| Histology, n (%) | |
| Endometrioid | 3 (13.6%) |
| Mucinous | 1 (4.5%) |
| Serous | 18 (81.81%) |
| Debulking status, n (%) | |
| Optimal (1- 10cm) | 20 (90.90%) |
| Sub-optimal (>10cm) | 2 (9.09%) |
| mRNA | |
| ALK2 and ALK3 | 22 (100%) |
| ALK6 | 12 (54.5%) |
| MISR2 | 14 (63.6%) |
| Protein | |
| ALK2 and ALK3 | 21 (95%) |
| ALK6 | 11 (50%) |
| MISR2 | 14 (63.6%) |
Fig. (4).

Candidate T1R (ALK 2, 3, and 6) are expressed in the primary EOC. Primary cell cultures were analyzed for mRNA (A) and protein expression (B) of ALK 2, ALK 3, ALK 6, and MISR2. (A) Lane 1–4 represents differential mRNA expression of ALK 2, ALK 3, ALK 6 and MISR2 in primary cell cultures detected using RT-PCR. (B) Immunoblotting of primary EOC for MISR2 and T1R expression. Primary EOC lysates were separated on 10% SDS-PAGE under non-reducing conditions, transferred to PVDF membranes and probed with MISR2 or T1R specific Abs. Lane 1–6 demonstrates variable protein expression of T1R and MISR2. GAPDH was used as a control in both RT-PCR and western blotting.
MIS-Dependent Smad 1/5/8 Activation and Impact on Cell Growth
We investigated whether primary cell cultures expressing the relevant T1R and MISR2 could activate Smad 1/5/8 in response to MIS. Exposure to 15ug/mL MIS or 25 ng/ml of BMP4 resulted in induction of pSmad 1/5/8 in Z3 control cells expressing MISR2 (Fig. 5A). We observed variable amounts of pSmad activation (3 of 4 primary cultures, 75%) after treatment with MIS in a subset of primary cell cultures (Fig. 5B). Cell growth inhibition after 72 hours of treatment with MIS was determined by MTT-based assay (Fig. 6A, B). The treatment of Z3 control cells with MIS showed dose dependent cytotoxic effect with highest cell growth inhibition observed at 30ug/mL dose (59% cell viability compared to untreated controls) (Fig. 6A). MIS treatment had no cytotoxic effect in MISR2 non- expressing SKOV3 cells as we observed more than 90% cell viability compared to control. MIS also showed cell cytotoxicity effect in primary cells, which showed cell viability ranging from 57% to 87% with average cell viability of 71% (Fig. 6B). Four out of five primary cell cultures showed significantly decreased cell viability on treatment with MIS. Thus our data suggests that MIS treatment can cause growth inhibition of primary tumor cells in ovarian cancer.
Fig. (5).
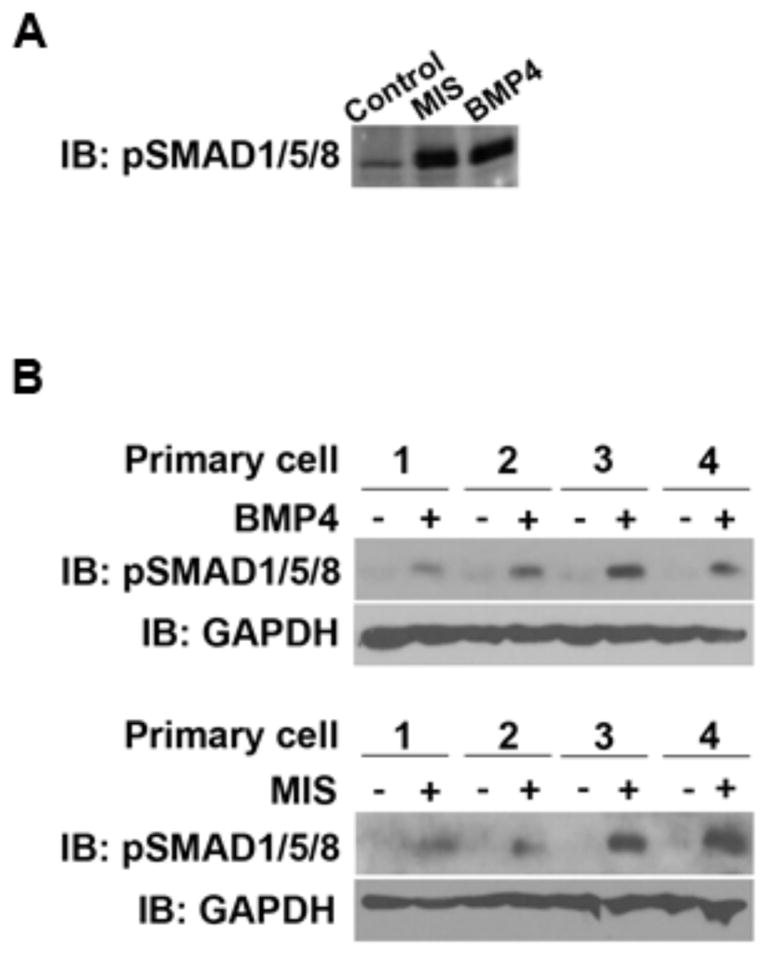
MIS ligand activates Smad phosphorylation 1/5/8 in MISR2 overexpressing Z3 and primary cell cultures. (A) Z3 cells were untreated or treated with 15ug/ml of MIS or 25ng/ml of BMP4 for 4 hours. Samples were analyzed by western blot analysis for pSmad1/5/8 protein levels in the control, MIS and BMP4-treated groups. (B) Western blot analysis for pSmad1/5/8 protein levels in primary cell cultures. Serum deprived primary cell cultures were treated with diluent or MIS 15ug/ml (upper row) and diluent or 25ng/ml of BMP4 (lower row) for 4 hours respectively and analyzed by immunoblotting as described in methods.
Fig. (6).

Effect of MIS on cell viability. (A) MIS induced a dose dependent decrease in cell viability in MISR2 overexpressing Z3 and primary cell cultures whereas no effect was observed in MISR2 negative SKOV3 cell line. Z3, SKOV3 and a representative primary cell culture cells were treated with various concentrations of MIS ranging 3.75–30ug/ml and analyzed using MTT assay for cell viability. (B) Exposure to MIS (30 ug/ml) resulted in decreased cell viability in primary cell cultures. Primary cell cultures (n=5) were treated with 30ug/ml MIS at 24 h after plating and after 72 h, cell viability was analyzed by MTT assay. Results are presented as percentage of control which was calculated using the equation: ((mean absorbance of treated cells/mean absorbance of control cells) x 100). Data are expressed as mean ± standard deviation (SD) from 3 independent experiments.
DISCUSSION
Few molecular targets have been identified for therapeutic use in the treatment of ovarian cancer. Studies to date suggest that the MIS receptor may serve as a potential candidate for targeted therapy. Important limitations to application of such therapy include relative shortage of ligand or other activating mimetic agents, lack of data on the prevalence T1R in EOC, and the impact of MISR2/T1R pairing on downstream signaling. The aims of the present study were to first characterize the expression patterns of candidate MIS T1R in primary EOC. We report that the majority of EOC express ALK2 and ALK3 along with MISR2: the combination of these receptors appears adequate for response to MIS in primary cell cultures. These results are encouraging for the widespread suitability of EOC for such studies.
Most studies on the MIS-MISR2 receptor system have specifically analyzed MISR2 and the consequences of its activation. We have previously shown that the majority of ovarian cancers express MISR2 protein [15]. These findings are confirmed in this larger cohort of EOC patients. Additionally we observed that nearly all EOC express 2 of the candidate T1R (ALK2 and ALK3, >90%). In all, 63% of EOC express MISR2 along with both ALK2 and ALK3. ALK6 in contrast is only expressed in 34% of EOC when MISR2 is expressed. We observed minimal differences in clinical outcomes associated expression of various MIS receptor combinations in primary EOC. Expression of MISR2 was associated with lower stage and a higher rate of complete cytoreduction at primary surgery. Despite these findings, in the absence of MISR2 expression, there was no impact on overall 5-year survival based on MISR2 status. This contradicts survival analysis in our prior study of a smaller dataset. These findings are not unexpected given the relative lack of serum MIS levels in post-menopausal women.
We show that T1R and MISR2 receptors are expressed at both mRNA and protein level in primary specimens and short-term cultures and their expression patterns were similar to that seen in the larger TMA cohort. Our results are in agreement with earlier studies showing that the presence of MISR2 is sufficient for Smad activation in response to MIS and that downstream effectors cause in vitro growth inhibition in [21]. Recombinant human MIS has been reported to inhibit the in vitro and in vivo growth of mouse ovarian carcinoma (MOVCAR) cells [9]. A modulated downstream response to MIS which was dependent on specific T1R was previously suggested by Visser et al. [22]. Using a mouse embryonic carcinoma cell line (P19) they determined that among the different type I candidates tested, only ALK2 was found to be important in MIS-mediated müllerian duct regression and capable of signaling using a Tlx-2 reporter gene assay. In contrast, ALK6 failed to significantly enhance the MIS-dependent reporter gene activation. Of note, ALK3 was not evaluated in this study [22]. In our study, the majority of TMA and primary OC cells expressed both ALK2 and ALK3 receptors and in the subset analyzed most cultures were capable of Smad signaling and growth inhibition in response to MIS. Collectively these data would suggest that expression of T1R is not a major limiting factor in the response to MIS by the majority of EOC. While recognizing that approximately one-third of EOC will be MISR2 non-expressing, in the era of tailored therapy, this should not pose a major clinical concern. We found activation of the MIS pathway results in up to 57–59% decrease in cell viability of primary cell culture and MISR2 over-expressing Z3 cell line. Together, these findings suggest that ALK3, in addition to ALK2, is an important T1R for MIS signaling, though there is likely redundancy in the human EOC model
An important limitation to the current study is the inability to differentiate the relative contributions of specific T1Rs in MIS-dependent growth inhibition. Additionally we were only able to test a subset of primary short-term cultures for growth inhibition to MIS. We also recognize that the use of short-term primary cultures does not rule out some contribution from associated stromal and fibroblast cells to the results. To better delineate the specific roles attributed to candidate T1Rs, future studies will utilize specific neutralizing antibodies to block one or more of these receptors. We are also developing a chimeric model, where MISR2 will be expressed with either ALK2, ALK3 or ALK6 individually linked to a unique extracellular domain to allow specificity of activation. These experiments will help to define the relative contributions of each T1R in MIS signaling. These functional studies are the next steps for gaining insight into the biology of MIS signaling in ovarian cancer and providing data that improves the purposeful manipulation of this receptor system for therapeutic gain.
In summary, our study contributes to the research on MIS as a therapeutic agent by demonstrating ALK2 and ALK3 are present in majority of EOC and in nearly all EOC expressing MISR2. While MIS induces growth inhibition in primary cultures expressing these receptors, the degree of growth inhibition is limited, suggesting a minor role for MIS alone as a therapeutic agent. Previous work suggests a role for synergy with cytotoxic drug [9]: this is a strategy which should be pursued further. Future studies should be guided by additional research into downstream MIS-dependent signaling pathways to help predict which existing therapies could synergize with MIS treatment.
Acknowledgments
Grant support was provided by the National Institute of Health Research Funding (R01CA148747 to WC).
Footnotes
Grant support was provided by the National Institute of Health Research Funding (R01CA148747 to WC).
CONFLICT OF INTEREST
The authors declare there are no financial or nonfinancial competing interests.
References
- 1.Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J Clin. 2010;60(5):277–300. doi: 10.3322/caac.20073. [DOI] [PubMed] [Google Scholar]
- 2.Josso N, Cate RL, Picard JY, et al. Anti-mullerian hormone: the Jost factor. Recent Prog Horm Res. 1993;48:1–59. doi: 10.1016/b978-0-12-571148-7.50005-1. [DOI] [PubMed] [Google Scholar]
- 3.He WW, Gustafson ML, Hirobe S, Donahoe PK. Developmental expression of four novel serine/threonine kinase receptors homologous to the activin/transforming growth factor-beta type II receptor family. Dev Dyn. 1993;196(2):133–42. doi: 10.1002/aja.1001960207. [DOI] [PubMed] [Google Scholar]
- 4.Clarke TR, Hoshiya Y, Yi SE, Liu X, Lyons KM, Donahoe PK. Mullerian inhibiting substance signaling uses a bone morphogenetic protein (BMP)-like pathway mediated by ALK2 and induces SMAD6 expression. Mol Endocrinol. 2001;15(6):946–59. doi: 10.1210/mend.15.6.0664. [DOI] [PubMed] [Google Scholar]
- 5.La Marca A, Volpe A. The Anti-Mullerian hormone and ovarian cancer. Hum Reprod Update. 2007;13(3):265–73. doi: 10.1093/humupd/dml060. [DOI] [PubMed] [Google Scholar]
- 6.Behringer RR, Cate RL, Froelick GJ, Palmiter RD, Brinster RL. Abnormal sexual development in transgenic mice chronically expressing mullerian inhibiting substance. Nature. 1990;345(6271):167–70. doi: 10.1038/345167a0. [DOI] [PubMed] [Google Scholar]
- 7.Lawrence DA. Transforming growth factor-beta: a general review. Eur Cytokine Netw. 1996;7(3):363–74. [PubMed] [Google Scholar]
- 8.Shi Y, Massague J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. 2003;113(6):685–700. doi: 10.1016/s0092-8674(03)00432-x. [DOI] [PubMed] [Google Scholar]
- 9.Pieretti-Vanmarcke R, Donahoe PK, Szotek P, et al. Recombinant human Mullerian inhibiting substance inhibits long-term growth of MIS type II receptor-directed transgenic mouse ovarian cancers in vivo. Clin Cancer Res. 2006;12(5):1593–8. doi: 10.1158/1078-0432.CCR-05-2108. [DOI] [PubMed] [Google Scholar]
- 10.Fuller AF, Jr, Krane IM, Budzik GP, Donahoe PK. Mullerian inhibiting substance reduction of colony growth of human gynecologic cancers in a stem cell assay. Gynecol Oncol. 1985;22(2):135–48. doi: 10.1016/0090-8258(85)90019-8. [DOI] [PubMed] [Google Scholar]
- 11.Chin TW, Parry RL, Donahoe PK. Human mullerian inhibiting substance inhibits tumor growth in vitro and in vivo. Cancer Res. 1991;51(8):2101–6. [PubMed] [Google Scholar]
- 12.Pieretti-Vanmarcke R, Donahoe PK, Pearsall LA, et al. Mullerian Inhibiting Substance enhances subclinical doses of chemotherapeutic agents to inhibit human and mouse ovarian cancer. Proc Natl Acad Sci USA. 2006;103(46):17426–31. doi: 10.1073/pnas.0607959103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wei X, Dombkowski D, Meirelles K, et al. Mullerian inhibiting substance preferentially inhibits stem/progenitors in human ovarian cancer cell lines compared with chemotherapeutics. Proc Natl Acad Sci USA. 2010;107(44):18874–9. doi: 10.1073/pnas.1012667107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chang HL, Pieretti-Vanmarcke R, Nicolaou F, et al. Mullerian inhibiting substance inhibits invasion and migration of epithelial cancer cell lines. Gynecol Oncol. 2011;120(1):128–34. doi: 10.1016/j.ygyno.2010.09.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bakkum-Gamez JN, Aletti G, Lewis KA, et al. Mullerian inhibiting substance type II receptor (MISIIR): a novel, tissue-specific target expressed by gynecologic cancers. Gynecol Oncol. 2008;108(1):141–8. doi: 10.1016/j.ygyno.2007.09.010. [DOI] [PubMed] [Google Scholar]
- 16.Wieser R, Wrana JL, Massague J. GS domain mutations that constitutively activate T beta R-I, the downstream signaling component in the TGF-beta receptor complex. EMBO J. 1995;14(10):2199–208. doi: 10.1002/j.1460-2075.1995.tb07214.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nohno T, Ishikawa T, Saito T, et al. Identification of a human type II receptor for bone morphogenetic protein-4 that forms differential heteromeric complexes with bone morphogenetic protein type I receptors. J Biol Chem. 1995;270(38):22522–6. doi: 10.1074/jbc.270.38.22522. [DOI] [PubMed] [Google Scholar]
- 18.Sedes L, Leclerc A, Moindjie H, et al. Anti-Mullerian hormone recruits BMPR-IA in immature granulosa cells. PLoS One. 2013;8(11):e81551. doi: 10.1371/journal.pone.0081551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Catlin EA, Tonnu VC, Ebb RG, et al. Mullerian inhibiting substance inhibits branching morphogenesis and induces apoptosis in fetal rat lung. Endocrinology. 1997;138(2):790–6. doi: 10.1210/endo.138.2.4906. [DOI] [PubMed] [Google Scholar]
- 20.Ragin RC, Donahoe PK, Kenneally MK, Ahmad MF, MacLaughlin DT. Human mullerian inhibiting substance: enhanced purification imparts biochemical stability and restores antiproliferative effects. Protein Expr Purif. 1992;3(3):236–45. doi: 10.1016/1046-5928(92)90020-w. [DOI] [PubMed] [Google Scholar]
- 21.Masiakos PT, MacLaughlin DT, Maheswaran S, et al. Human ovarian cancer, cell lines, and primary ascites cells express the human Mullerian inhibiting substance (MIS) type II receptor, bind, and are responsive to MIS. Clin Cancer Res. 1999;5(11):3488–99. [PubMed] [Google Scholar]
- 22.Visser JA, Olaso R, Verhoef-Post M, Kramer P, Themmen AP, Ingraham HA. The serine/threonine transmembrane receptor ALK2 mediates Mullerian inhibiting substance signaling. Mol Endocrinol. 2001;15(6):936–45. doi: 10.1210/mend.15.6.0645. [DOI] [PubMed] [Google Scholar]