

Abstract
Next-generation sequencing (NGS) is a powerful platform for identifying cancer mutations. Routine clinical adoption of NGS requires optimized quality control metrics to ensure accurate results. To assess the robustness of our clinical NGS pipeline, we analyzed the results of 304 solid tumor and hematologic malignancy specimens tested simultaneously by NGS and one or more targeted single-gene tests (EGFR, KRAS, BRAF, NPM1, FLT3, and JAK2). For samples that passed our validated tumor percentage and DNA quality and quantity thresholds, there was perfect concordance between NGS and targeted single-gene tests with the exception of two FLT3 internal tandem duplications that fell below the stringent pre-established reporting threshold but were readily detected by manual inspection. In addition, NGS identified clinically significant mutations not covered by single-gene tests. These findings confirm NGS as a reliable platform for routine clinical use when appropriate quality control metrics, such as tumor percentage and DNA quality cutoffs, are in place. Based on our findings, we suggest a simple workflow that should facilitate adoption of clinical oncologic NGS services at other institutions.
Introduction
The advance of next-generation sequencing (NGS) is a cornerstone of a recent development in molecular pathology, variably referred to as “personalized,” “precision,” or “individualized” medicine. Much of the focus of clinical NGS has been on oncology, as there are clear diagnostic, prognostic, and therapeutic implications for a multitude of genomic mutations in both solid and liquid malignancies. For example, in non-small cell lung cancers, activating mutations of the EGFR gene predict therapeutic response to tyrosine kinase inhibitors (TKI) such as erlotinib, gefitinib, and afatinib [1–3]. The KRAS protein acts downstream of EGFR, and thus mutations of the KRAS gene predict resistance to TKI [2,4–6]. In metastatic melanoma, BRAF V600 mutations predict response to dabrafenib, vemurafenib, and trametinib [7]. Mutations of the FLT3 and NPM1 genes affect the prognosis of karyotypically normal acute myeloid leukemia and aid in the decision whether or not to pursue hematopoietic stem cell transplantation [8]. In addition, activating mutations of JAK2, which encodes a tyrosine kinase essential for cytokine and growth factor signaling, are found in a large proportion of patients with myeloproliferative neoplasms [9]. Ruxolitinib is a JAK1/JAK2 inhibitor approved for the treatment of myelofibrosis [10], and additional JAK2 inhibitors are in clinical development [11].
Detection of mutations in EGFR, KRAS, BRAF, FLT3, NPM1, and JAK2 is most commonly accomplished by targeted tests that are designed to detect one or at most a small number of mutations in a single gene. However, NGS is gaining momentum as a complementary test for a number of reasons. Firstly, clinical trials for targeted cancer therapies rely on detection of mutations that are frequently not covered by existing targeted tests. In contrast to the laborious and lengthy process of validating and implementing a new molecular assay testing for one or a few mutations, NGS greatly simplifies the task of providing coverage of one of more additional mutations of interest. Secondly, targeted tests can provide misleading results. For example, the widely used FDA-approved cobas® EGFR Mutation Test only detects exon 19 deletion and L858R mutations, which together only comprise the mutations found in 85% of EGFR-mutated lung cancers [12]. In a significant proportion of cases, this test fails to identify therapeutically targetable mutations. Thirdly, targeted tests may fail to detect the very mutation they are designed to detect. Our group has recently reported a striking failure of two separate single-gene tests for the BRAF gene to detect a V600E mutation in a melanoma specimen [13]. The mutation was clearly demonstrated by concurrent NGS analysis. Finally, as has recently become apparent, tumors frequently harbor mutations that are therapeutically targetable but are not typically seen in that tumor type. Due to its massively parallel nature, NGS is very well-suited for detecting mutations in unexpected genes. One study reported a three-fold increased yield of clinically actionable mutations with NGS as compared to traditional molecular approaches targeting mutation hotspots [14].
Multiple recent studies have investigated the potential utility of NGS for detection of clinically actionable cancer mutations with encouraging results [14–25]. With the exception of one study [21], all demonstrated excellent performance of NGS on various platforms as measured by detection of point mutations and small insertions and deletions (indels). In many cases, additional potentially important variants were uncovered by NGS. All of the aforementioned studies were designed to validate clinical NGS pipelines that were not yet in clinical practice. As a consequence, they enrolled selected samples from previously examined specimens. With the exception of one commercially-sponsored study [14], in all cases a very limited number of samples was re-examined (range 13–61), often from only a single tissue type. While these important contributions confirm the potential usefulness of clinical NGS, they do not address the important question whether a well-validated NGS pipeline performs at an acceptable level in day-to-day clinical practice. Here we present a summative analysis of mutation results and quality control metrics obtained during the first year (March 2013 through March 2014) of clinical solid and liquid malignancy NGS carried out at the Center for Personalized Diagnostics at the University of Pennsylvania Health System. More than 900 specimens were submitted and processed during this time frame. We report that using our validated molecular and bioinformatics pipeline [26] with pre-determined tumor percentage and DNA quality cutoffs, we achieved excellent NGS data quality as determined by virtually perfect concordance between NGS and targeted single-gene tests for various genes in a large number of solid and liquid malignancy specimens.
Materials and Methods
Specimen Characteristics and Processing
Over the course of the study duration, 938 liquid and solid tumor specimens were submitted to the Center for Personalized Diagnostics (Table 1). Specimens were eligible for NGS if they passed the tumor percentage, DNA quality, and DNA quantity thresholds that had been determined at the time of the validation of the NGS assay, which preceded the study period. Briefly, specimens with <10% tumor were not eligible for NGS, because sequencing of samples with lower tumor percentages frequently yielded changes that were represented in fewer than five unique reads, making it difficult to distinguish true variants from sequencing artifacts. For similar reasons, DNA quality and quantity were judged to be insufficient, and the specimen was ineligible for NGS, if the DNA concentration was <1 ng/μL; the DNA concentration was <5 ng/μL with >20% DNA degraded; the DNA concentration was <50 ng/μL with >45% DNA degraded; or DNA degradation was >60%. Degraded DNA was defined as the proportion of DNA under 1000 bp in length.
Table 1. Characteristics of Specimens by Tumor Site.
| Tumor site | Number of specimens submitted for NGS | Specimens with DNA quality or quantity inadequate for NGS analysis | Number of “shared” specimens (i.e., results are available from both NGS and targeted tests) |
|---|---|---|---|
| Lung | 196 | 13 (6.6%) | 101 (51.5%) |
| Brain | 174 | 9 (5.2%) | 6 (3.4%) |
| Bone Marrow | 161 | 0 | 95 (59.0%) |
| Lymph Node | 70 | 8 (11.4%) | 32 (45.7%) |
| Peripheral blood | 56 | 0 | 29 (51.8%) |
| Liver | 36 | 4 (11.1%) | 5 (13.9%) |
| Skin | 23 | 2 (8.7%) | 5 (21.7%) |
| Other | 222 | 24 (10.8%) | 31 (14.0%) |
Tumor site is not necessarily tissue of origin. Please note that in contrast to solid tumor specimens, all liquid specimens (bone marrow and peripheral blood) were adequate for NGS processing.
To determine the tumor percentage and volume of solid tumors, hematoxylin- and eosin-stained tissue specimens were evaluated by an anatomic pathologist, and the region with the highest tumor burden was marked. Genomic DNA was extracted from fresh bone marrow or peripheral blood using the Gentra Puregene Cell Kit (Qiagen, Netherlands). For formalin-fixed, paraffin-embedded (FFPE) specimens, tissues were macro-dissected from 5 μM or 10μM slides. Scrapings were dewaxed with Qiagen Deparaffinization Solution and purified with Gentra Puregene Tissue reagents following the manufacturer’s protocol (Qiagen, Netherlands). DNA quantification was performed using the Qubit Broad Range assay following manufacturer’s protocols (Life Technologies, CA). Agilent Genomic TapeScreens were used following manufacturer’s protocols to assess the degree of DNA degradation (Agilent, CA).
Targeted Molecular Testing
EGFR, KRAS, and BRAF Assays
The mutational status of EGFR exons 19 and 21 was determined using a laboratory-developed test (LDT) as previously described [27]. Briefly, genomic DNA was extracted from FFPE tissue and amplified with primers covering two regions, one that is commonly deleted in exon 19, and a part of exon 21 that encompasses codon 858. The L858R missense mutation in exon 21 creates a new Sau96I cleavage site within exon 21. The amplification products were digested with Sau96I and then separated by capillary electrophoresis.
KRAS mutations in codons 12 and 13 were assayed using a LDT as previously described [28]. Briefly, genomic DNA was extracted and amplified using primers designed to detect point mutations, hybridized to target-specific capture probes, and subjected to a bead assay (Lumina, TX).
BRAF mutations were assayed by pyrosequencing of an amplified portion of the BRAF gene including codon 600, as previously described [29].
FLT3 Assay
DNA was extracted using the QIAamp DNA Blood Mini Kit (Qiagen, Netherlands). Mutation analysis of the FLT3 gene was performed using multiplex PCR amplification with two sets of fluorescently labeled primers. For internal tandem duplication (ITD) detection, PCR was performed with the following primers: 5’-GCA ATT TAG GTA TGA AAG CCA GC-3’ (forward) and 5’-CTT TCA GCA TTT TGA CGG CAA CC-3’ (reverse); forward primers were labeled with 6-carboxyfluorescin (6-FAM), and reverse primers were labeled with VIC. An internal tandem duplication (ITD) was determined to be present if a product larger than the wild-type (329 bp) product was detected by capillary electrophoresis. Detection of the D835 mutation of FLT3 (NM_004119.2: c.2503_2505) was based on the fact that this mutation abolishes an EcoRV cleavage site. PCR was performed with the following primers: 5’-GTA AAA CGA CGG CCA GCC GCC AGG AAC GTG CTT-3’ (forward) and 5’-CAG GAA ACA GCT ATG ACG ATA TCA GCC TCA CAT TGC CCC-3’ (reverse); forward primers were labeled with NED at the 5’ end. After EcoRV digestion, PCR products were analyzed by capillary electrophoresis using a 3500xL Genetic Analyzer (Life Technologies, NY). A D835 point mutation was indicated by the presence of a 129 bp fragment.
NPM1 Assay
Total RNA was extracted using the QIAamp RNA Blood Mini Kit (Qiagen, Netherlands), reverse transcribed, and amplified in a multiplex PCR reaction using primers designed to detect common mutations in NPM1 (NM_002520.4) using the Signature NPM1 Mutations Assay (Asuragen, TX). Labeled PCR products were hybridized to target-specific capture probes covalently bound to fluorescent microspheres in a liquid bead array followed by analysis with a Luminex 100 (Luminex, TX). Interpretation was based on the mean fluorescence intensity (MFI) obtained from a minimum of 50 microspheres.
JAK2 Assay
Genomic DNA was isolated from leukocytes using the QIAamp DNA Blood Mini Kit (Qiagen, Netherlands) and amplified using real-time PCR with primers flanking JAK2 codon 617. Allelic discrimination between the normal sequence and the JAK2 V617F (NM_004972 c.1849G>T) mutation was subsequently accomplished by simultaneous differential hybridization of two sequence-specific probes, each labeled with a different fluorescent marker (MutaScreen Assay, Qiagen, Netherlands).
Next-Generation Sequencing and Bioinformatic Analysis
For solid tumors, target enrichment was performed with the TruSeq Amplicon Cancer Panel (Illumina, CA), a cancer gene panel consisting of 212 target amplicons covering mutation hotspots of 47 cancer genes. For hematologic malignancies, an in-house developed gene panel was utilized, which is composed of 382 amplicons covering 33 genes. For all successful sequencing runs, read depth was 250x at any given position, with 1000x mean coverage across the entire targeted sequence, and a Q30 at greater than 75% of reads. An in-house bioinformatics pipeline [26] was used to map reads, detect variants, and annotate them. Reads were de-multiplexed, mapped to the hg19 version of the human reference genome, filtered to remove off-target and poor-quality reads (Fig 1). Using custom scripts, four types of variants were extracted: single-nucleotide variants (SNVs), small indels, copy number variants, and large indels. Variants were then compared to an in-house developed knowledge base, which draws from publicly available sources such as PubMed, dbSNP database [30], COSMIC database [31], 1000 Genomes [32], and the Exome Variant Server (http://evs.gs.washington.edu). Using this knowledge base, variants were classified into 1 of 5 categories: disease associated mutation (DAM), likely pathogenic mutation (LPM), variants of uncertain significance (VUS), likely benign (LB), or benign (B). DAMs include mutations previously reported and associated with disease, including gain-of-function mutations in oncogenes (e.g., the canonical KRAS G12D mutation) and truncating mutations in known tumor suppressor genes. LPMs were classified as variants that had some evidence of disease association, such as case reports, but are not well described otherwise. Variants were classified as VUS if they had not been previously reported either as a disease-associated mutation or as a normal variant on the Exome Variant Server, but whose pathogenicity could not be established with certainty. Variants were classified as LB if there was no report of association with disease and if they occurred in regions of the gene not predicted to have pathologic consequences. Variants noted on the clinical report, which are the ones included in this analysis, included the DAM, LPM and VUS (not LB or B). All reported variants were manually reviewed using the Integrative Genomics Viewer (IGV) [33] by at least two pathologists.
Fig 1. Next-generation sequencing data analysis pipeline.

Data analysis occurs in three sequential stages, pre-processing of NGS reads, variant calling, and variant annotation. Of note, large indels are detected by an examination of reads that failed to map to target regions of the reference genome and are recovered from a pool of rejected reads (“Trash”). SNVs, single nucleotide variants. CNVs, copy number variation.
Ethics Statement
Patient data were analyzed anonymously in accordance with institutional practice guidelines. The institutional review board of the University of Pennsylvania determined this study to be exempt.
Results
Study Design and Specimen Characteristics
The University of Pennsylvania Health System began routine clinical NGS of solid tumors and hematologic malignancies in February of 2013. During validation of our NGS pipeline, we established thresholds for acceptable tumor percentage (10%) and DNA quantity and quality (see Methods for details).
For a large number of specimens, clinicians ordered targeted single-gene tests and NGS analysis on the same specimen (Table 1): this occurred frequently with pulmonary and hematological specimens, but relatively rarely with specimens derived from other sites such as brain. We reasoned that comparing results from targeted and NGS testing of all “shared” specimens (i.e. specimens that had been analyzed by both NGS and single-gene tests) would allow us to probe the robustness of our NGS pipeline. We therefore set out to compare NGS and targeted test results in specimens that underwent NGS analysis during the first year of operation of the NGS pipeline (March 1, 2013 through March 1, 2014). During this time, 938 specimens (717 solid and 221 hematologic) were submitted for NGS analysis. While the majority of solid tumor specimens were composed of >50% tumor cells (Fig 2), a fraction of solid tumors was not tested due to low tumor percentage. The most common tumor sites were lung, brain, bone marrow, lymph nodes, and peripheral blood (Table 1). A small fraction of solid tumor specimens could not be analyzed by NGS due to inadequate DNA quantity or quality (n = 60, 8.3%), however this was not an issue with peripheral blood and bone marrow specimens.
Fig 2. Tumor Percentage of Solid Tumor Specimens.
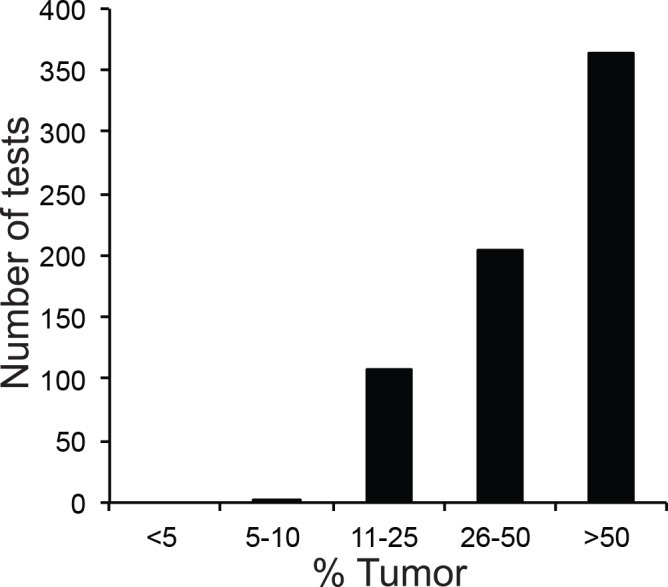
Specimens were analyzed by next-generation sequencing (NGS) within the study period of March 1, 2013 and March 1, 2014.
Comparison of EGFR and KRAS Gene Mutations by NGS and Targeted Testing
We were particularly interested in the performance of EGFR testing in our NGS assay, because mutations in this gene may be therapeutically targetable, and because it is often challenging to analyze, as DNA degradation and low tumor percentage are frequently encountered in lung cancer specimens. Since KRAS mutations were frequently evaluated alongside EGFR in cases of lung cancers at our institution, we also wanted to compare the performance of both methods on this gene. The tumor percentages for all specimens for which targeted EGFR mutation analysis was reported during the study period (n = 283) are shown in Fig 3. Approximately 10% of the cases evaluated by targeted testing had a tumor percentage below the cutoff for NGS.
Fig 3. Tumor Percentage of Specimens Analyzed by Targeted EGFR Tests.
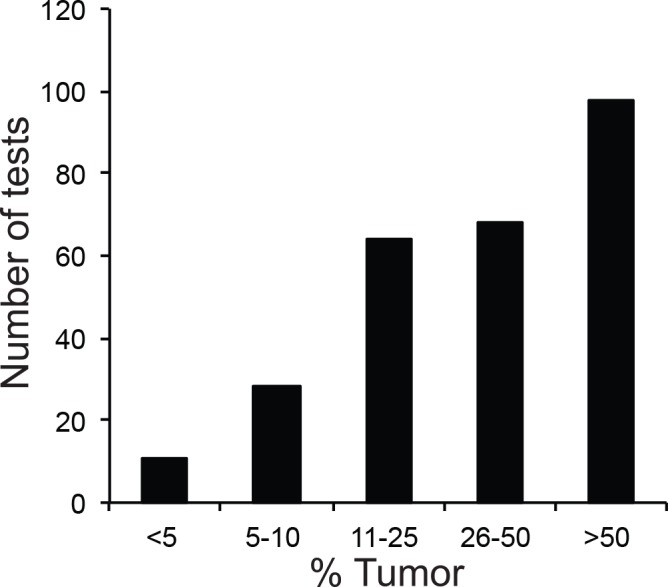
Cases were placed into 5 bins (<5%, 5–10%, 11–25%, 26–50%, >50% tumor cells in specimen).
We compared NGS and targeted testing in 139 specimens that had been tested for EGFR mutations (Fig 4A) and 138 that had been tested for KRAS mutations by both methods (Fig 4B). Among the shared specimens, all generated a result with the targeted single-gene methods. However, 15/139 (11%) of shared EGFR-tested and 13/138 (9%) of shared KRAS-tested specimens were excluded from NGS analysis due to insufficient DNA quality or quantity.
Fig 4. Concordance Analysis of Solid Tumor Specimens.

The specimens shown were submitted for both NGS and targeted tests for EGFR (A), KRAS (B), and BRAF (C) mutations. Note that all mutations seen by targeted testing were also found by NGS when specimens with inadequate DNA quantity and/or quality are excluded.
In the remaining specimens, all mutations detected by the targeted assays were also detected by NGS. Conversely, NGS identified a number of mutations that the targeted tests were not designed to detect. For example, our targeted EGFR mutation test only covers deletions in exon 19 and the L858R mutation in exon 21. Similarly, the targeted KRAS test only detects mutations in codons 12 and 13. In 10 EGFR shared cases (6%), additional pathogenic mutations were detected by NGS. In two of these cases, the T790M mutation was found, which predicts resistance to TKI therapy [4]. Additionally, NGS detected EGFR amplification in five cases; the predictive value of this copy number alteration in the context of TKI therapy is currently unclear. In four KRAS shared cases (3%), NGS detected mutations in codon 61, which predict resistance to TKI therapy [34].
Comparison of BRAF Gene Mutations by NGS and Targeted Testing
The BRAF single-gene test was performed less frequently in parallel with solid tumor NGS than the EGFR or KRAS tests, because thyroid fine-needle aspiration samples, for which BRAF testing was frequently ordered, were not originally validated for the NGS assay. Of 224 specimens that were tested with targeted BRAF tests, 38 were also analyzed by NGS (Fig 4C). Among the shared specimens, all generated a result with the targeted BRAF test. In contrast, 8/38 shared cases (21%) could not be analyzed by NGS due to poor DNA quality or inadequate DNA quantity. All remaining shared specimens showed perfect concordance between NGS and targeted testing.
Comparison FLT3, NPM1, and JAK2 Mutations by NGS and Targeted Testing
Of 221 hematologic specimens tested by NGS during the study period, 118 were also tested with the FLT3, and 98 with the NPM1 targeted tests. In two cases, the targeted FLT3 test detected an internal tandem duplication (ITD) that was not automatically called by the NGS analysis pipeline (Fig 5A). However, manual review of the sequencing data demonstrated ITD mutations at allele frequencies of 1.3% and 1.6% (Fig 6A and 6C, respectively). In one of these two cases, the hematologic malignancy NGS panel additionally detected a pathogenic FLT3 D839G (c. 2516A>G) mutation (Fig 6B); this mutation was not detected by the FLT3 single-gene test that is designed to detect only ITDs and D835 mutations.
Fig 5. Concordance Analysis of Hematologic Malignancy Specimens.

The specimens shown were submitted for both NGS and targeted tests for FLT3 (A), NPM1 (B), and JAK2 (C) mutations. Note that all samples were adequate for testing by both single-gene assays and NGS.
Fig 6. Two Specimens with Low-Allele Frequency FLT3 Internal Tandem Duplications.

In one specimen (A and B), a 24 bp internal tandem duplication (ITD) was seen in 7 out of 529 reads for an allele frequency of 1.3%. (A) Four of the reads containing insertions (purple bars) are shown using the Integrative Genomics Viewer. This specimen additionally harbored a FLT3 D839G mutation in 45% of reads (B). A second specimen (C) harbored a 33 bp FLT3 ITD in 12 out of 739 reads, for an allele frequency of 1.6%. Nine of the reads carrying an ITD are pictured.
In the remainder of FLT3 shared cases, and also in all NPM1 shared cases (Fig 5B), NGS and targeted tests were concordant. Additionally, a small number of specimens (n = 8) that had been tested for JAK2 mutations by both modalities were analyzed and showed complete concordance (Fig 5C).
Discussion
In this study, we report the properties of solid and liquid malignancy specimens processed during the first year of clinical oncologic NGS performed within the University of Pennsylvania Health System. We found that when we adhered to two predetermined quality control metrics, i.e., tumor percentage and DNA quantity and quality, we achieved excellent NGS data as determined by virtually perfect concordance between NGS and targeted, single-gene testing. While a number of recent studies have confirmed the potential utility of clinical NGS for oncology [14–25], there have been none, to our knowledge, that have evaluated the quality of data generated during day-to-day practice at a clinical oncologic sequencing facility. Additionally, in contrast to previous validation studies, we examined a larger number of specimens from a greater variety of tissues, and our data is therefore less subject to sample selection bias and may also more accurately reflect the expected annual case volume and distribution of tissue types encountered at a major academic medical center.
Solid tumors and hematologic malignancy specimens differed considerably in their performance across the established quality control measures. We found that a sizable fraction of solid but not liquid specimens yielded DNA of insufficient quality or quantity for NGS testing. With respect to specimens tested for EGFR and KRAS (predominantly lung), this was likely largely due to formalin fixation, which degrades DNA through cross-linking as well as other less well understood mechanisms [35]. One potential approach to reduce the number of samples that are currently rejected from NGS analysis might be to determine the amplifiability of extracted genomic DNA, for example by using the human genomic DNA quantitation and quality control assay by Kapa Biosystems (Wilmington, MA).
Our study highlights the complementarity of NGS and targeted tests for mutation detection. In a number of instances, NGS detected clinically important mutations that were not captured by targeted assays. For example, the EGFR test specifically interrogates potential exon 19 deletions and L858R mutations, which constitute about 85% of EGFR mutations in lung cancers [12]. In 9 shared cases, EGFR mutations were found in different regions of the gene by the NGS assay (Fig 4A). Similarly, clinically important KRAS mutations occur in codons 12, 13, and 61, but only codons 12 and 13 were evaluated by the in-house KRAS single-gene test. In 4 shared cases, KRAS mutations in codon 61 were detected by NGS only.
Targeted assays generally outperformed NGS in specimens with low tumor percentage, DNA quantity, or quality. In fact, we identified actionable mutations by targeted analysis in multiple cases that were unable to be analyzed by NGS (data not shown). In two specimens, a FLT3 ITD mutation, which was not automatically called by the NGS pipeline, was readily detected by the targeted assay. While FLT3 ITDs can be challenging to detect by NGS due to the complex structure of the mutation [36], the problem in these two cases was that the allele frequency fell below the validated threshold for automatic detection of 4%. However, manual inspection of the FLT3 exon 14 and the flanking intronic sequence using the Integrative Genomics Viewer (IGV) [33] clearly showed the presence of ITD mutations in both cases (Fig 6A and 6C). The detection of NGS for indel mutations, in contrast to single nucleotide variants, is not fundamentally limited by PCR artifacts and sequencing errors. Therefore, we validated our pipeline for the detection of FLT3 ITDs with allele frequencies of 1%, and we altered the indel allele frequency calling threshold specifically for FLT3 ITDs. Reanalysis of the two FLT3 cases with updated parameters, which was part of the revalidation study, revealed the expected results. These findings highlight the importance of manual review of the sequencing data.
Low tumor percentage was also limiting in solid tumor cases. Approximately 10% of specimens submitted for EGFR mutation testing were found to contain less than 10% tumor (Fig 3), thereby failing tumor percentage requirements for NGS testing. All of these specimens generated a result with the targeted assay. It should be noted, however, that as NGS methodologies continue to mature, the ability of NGS to detect single nucleotide variants in specimens with lower tumor percentage will improve. Rare variant detection by NGS is hampered by the high rate of amplification and sequencing errors, which can approach 1% for single nucleotide variants [37]. Various approaches to increase analytic sensitivity for rare variants by NGS have recently been described, including barcoding of each DNA fragment before amplification [38–40], barcoding of both strands of each fragment [41], and generation of multiple linked tandem copies of each DNA fragment by rolling circle amplification [42]. In particular, the latter method has been shown to improve sensitivity of rare variant detection by more than 100-fold without introducing excessive computational inefficiency.
There are a number of limitations to this study. First, only genes for which both NGS analysis and single-gene tests are performed at our institution were included in the analysis. While the genes examined in this study currently represent the essential core of cancer gene testing, it is possible that NGS might function less reliably with certain other genes or specific mutations that were not assessed. Of note, our NGS pipeline produced excellent results for the challenging genes EGFR and FLT3. We therefore expect that most other genes covered by our cancer panels generate NGS data of similarly high quality. Second, not all specimens that were submitted for NGS analysis were also tested by single-gene methods. For example, only 3.4% of brain tumor specimens submitted for NGS were at the same time examined by targeted tests, and therefore our conclusions may not necessarily extend to all tumors at this time. Additionally, the specimens tested did not contain the entire spectrum of mutations that can be evaluated by our targeted tests. Finally, our findings may not be generalizable to other platforms, especially those that utilize more complex gene panels.
Recently issued recommendations for validation and quality control of clinical NGS data [43] include monitoring of quality metrics (e.g., sequencing quality scores, depth of coverage, uniformity of coverage, mapping quality), proficiency testing, and confirmation of actionable results by independent methods. Multiple studies that appeared after these recommendations were published have established that particularly for solid tumors, tumor percentage and DNA quality are important additional metrics [14,22,23], and we found this as well in our validation studies (not shown). Proficiency testing for cancer NGS is not yet available but is currently being developed by the College of American Pathologists (CAP).
A proposed requirement to confirm actionable mutations by independent molecular methods [43] has been called into question [23]. Since single-gene tests such as Sanger sequencing may not have inherently greater sensitivity than a well-scaled NGS pipeline [15,44], verification by these methods might not improve the accuracy of test results. Accordingly, very recent guidelines by the College of American Pathologists leave the decision when and how to perform confirmatory testing of NGS results to the clinical laboratory [44]. Our EGFR single-gene test had greater analytical sensitivity than NGS, detecting mutations in samples with well under 10% tumor. However, our finding that within pre-defined tumor percentage and DNA quality cutoffs NGS showed perfect concordance confirms the notion that it is unnecessary to confirm each actionable mutation detected via NGS by a single-gene test. In addition, single-gene tests frequently do not cover important disease-associated mutations and in some instances fail to detect the very mutations they target [13].
Based on these considerations, we propose a workflow that integrates NGS as an adjunct diagnostic modality for solid and liquid neoplasms (Fig 7). The typical turn-around time of the NGS assay at the time of the study was around 7–10 days from receipt in the sequencing facility to the time the final report was signed out in the electronic medical record. Turnaround is generally faster for targeted tests, and thus it is advantageous to perform targeted tests, perhaps in addition to NGS, in clinically urgent situations. Specimens with low tumor percentage or DNA quantity or quality should be subjected to targeted tests that we found to be more analytically sensitive than NGS. On the other hand, when DNA quality and quantity is adequate and a turnaround time of 7–10 days is acceptable, NGS holds clear advantages, including increased clinical sensitivity within targeted genes of interest and additional information from other covered genes on the panel. In these cases, our data suggest that targeted tests may be safely omitted.
Fig 7. Proposed Workflow for NGS and Single-Gene Assays.

Three main decision points are highlighted. Specimens requiring an urgent turnaround time are routed directly for single-gene testing (possibly followed by NGS). Additionally, single-gene testing is performed on samples with less than 10% tumor or DNA inadequate for NGS (i.e., degraded or low quantity). In samples not meeting any of the above criteria, NGS is performed instead of single-gene testing. NGS results do not require confirmation by single-gene testing.
Supporting Information
(XLSX)
Acknowledgments
We thank Daniel Desloover for comments on the manuscript.
Data Availability
Raw deep sequencing cannot be provided because these data are derived from patient samples. Because of the Minimum Necessary Requirement of the HIPAA Privacy Rule these data may not be deposited into a public repository. A Supporting Information File accompanying the submission contains the de-identified results and interpretations of all clinical NGS tests included in our analysis next to the results of single-gene testing of the same specimen.
Funding Statement
The authors received no specific funding for this work.
References
- 1.Shepherd FA, Rodrigues Pereira J, Ciuleanu T, Tan EH, Hirsh V, Thongprasert S, et al. Erlotinib in previously treated non-small-cell lung cancer. N Engl J Med. 2005;353: 123–132. 10.1056/NEJMoa050753 [DOI] [PubMed] [Google Scholar]
- 2.Jackman DM, Miller VA, Cioffredi L-A, Yeap BY, Jänne PA, Riely GJ, et al. Impact of epidermal growth factor receptor and KRAS mutations on clinical outcomes in previously untreated non–small cell lung cancer patients: results of an online tumor registry of clinical trials. Clin Cancer Res. AACR; 2009;15: 5267–5273. 10.1158/1078-0432.CCR-09-0888 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.West H, Lilenbaum R, Harpole D, Wozniak A, Sequist L. Molecular analysis-based treatment strategies for the management of non-small cell lung cancer. J Thorac Oncol. 2009;4: S1029–39. 10.1097/JTO.0b013e3181b27170 [DOI] [PubMed] [Google Scholar]
- 4.Pao W, Wang TY, Riely GJ, Miller VA, Pan Q, Ladanyi M, et al. KRAS mutations and primary resistance of lung adenocarcinomas to gefitinib or erlotinib. PLoS Med. Public Library of Science; 2005;2: e17 10.1371/journal.pmed.0020017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Eberhard DA, Johnson BE, Amler LC, Goddard AD, Heldens SL, Herbst RS, et al. Mutations in the epidermal growth factor receptor and in KRAS are predictive and prognostic indicators in patients with non-small-cell lung cancer treated with chemotherapy alone and in combination with erlotinib. J Clin Oncol. 2005;23: 5900–5909. 10.1200/JCO.2005.02.857 [DOI] [PubMed] [Google Scholar]
- 6.Massarelli E, Varella-Garcia M, Tang X, Xavier AC, Ozburn NC, Liu DD, et al. KRAS mutation is an important predictor of resistance to therapy with epidermal growth factor receptor tyrosine kinase inhibitors in non-small-cell lung cancer. Clin Cancer Res. 2007;13: 2890–2896. 10.1158/1078-0432.CCR-06-3043 [DOI] [PubMed] [Google Scholar]
- 7.Sosman JA, Kim KB, Schuchter L, Gonzalez R, Pavlick AC, Weber JS, et al. Survival in BRAF V600-mutant advanced melanoma treated with vemurafenib. N Engl J Med. 2012;366: 707–714. 10.1056/NEJMoa1112302 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Schlenk RF, Döhner K, Krauter J, Fröhling S, Corbacioglu A, Bullinger L, et al. Mutations and treatment outcome in cytogenetically normal acute myeloid leukemia. N Engl J Med. 2008;358: 1909–1918. 10.1056/NEJMoa074306 [DOI] [PubMed] [Google Scholar]
- 9.Kralovics R, Passamonti F, Buser AS, Teo S-S, Tiedt R, Passweg JR, et al. A gain-of-function mutation of JAK2 in myeloproliferative disorders. N Engl J Med. 2005;352: 1779–1790. 10.1056/NEJMoa051113 [DOI] [PubMed] [Google Scholar]
- 10.Mesa RA, Yasothan U, Kirkpatrick P. Ruxolitinib. Nat Rev Drug Discov. Nature Publishing Group; 2012;11: 103–104. 10.1038/nrd3652 [DOI] [PubMed] [Google Scholar]
- 11.Gowin K, Mesa R. Emerging therapies for the treatment of chronic Philadelphia chromosome-negative myeloproliferative neoplasm-associated myelofibrosis. Expert Opin Investig Drugs. 2013;22: 1603–1611. 10.1517/13543784.2013.832199 [DOI] [PubMed] [Google Scholar]
- 12.Lindeman NI, Cagle PT, Beasley MB, Chitale DA, Dacic S, Giaccone G, et al. Molecular testing guideline for selection of lung cancer patients for EGFR and ALK tyrosine kinase inhibitors: guideline from the College of American Pathologists, International Association for the Study of Lung Cancer, and Association for Molecular Pathology. The Journal of molecular diagnostics: JMD. 2013. pp. 415–453. 10.1016/j.jmoldx.2013.03.001 [DOI] [PubMed] [Google Scholar]
- 13.Wilson MA, Morrissette JJD, McGettigan S, Roth D, Elder D, Schuchter LM, et al. What you are missing could matter: a rare, complex BRAF mutation affecting codons 599, 600, and 601 uncovered by next generation sequencing. Cancer Genet. 2014;207: 272–275. 10.1016/j.cancergen.2014.06.022 [DOI] [PubMed] [Google Scholar]
- 14.Frampton GM, Fichtenholtz A, Otto GA, Wang K, Downing SR, He J, et al. Development and validation of a clinical cancer genomic profiling test based on massively parallel DNA sequencing. Nat Biotechnol. 2013;31: 1023–1031. 10.1038/nbt.2696 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hadd AG, Houghton J, Choudhary A, Sah S, Chen L, Marko AC, et al. Targeted, high-depth, next-generation sequencing of cancer genes in formalin-fixed, paraffin-embedded and fine-needle aspiration tumor specimens. J Mol Diagn. 2013;15: 234–247. 10.1016/j.jmoldx.2012.11.006 [DOI] [PubMed] [Google Scholar]
- 16.Grossmann V, Roller A, Klein H-U, Weissmann S, Kern W, Haferlach C, et al. Robustness of amplicon deep sequencing underlines its utility in clinical applications. J Mol Diagn. 2013;15: 473–484. 10.1016/j.jmoldx.2013.03.003 [DOI] [PubMed] [Google Scholar]
- 17.Singh RR, Patel KP, Routbort MJ, Reddy NG, Barkoh BA, Handal B, et al. Clinical validation of a next-generation sequencing screen for mutational hotspots in 46 cancer-related genes. J Mol Diagn. 2013;15: 607–622. 10.1016/j.jmoldx.2013.05.003 [DOI] [PubMed] [Google Scholar]
- 18.Spencer DH, Sehn JK, Abel HJ, Watson MA, Pfeifer JD, Duncavage EJ. Comparison of clinical targeted next-generation sequence data from formalin-fixed and fresh-frozen tissue specimens. J Mol Diagn. 2013;15: 623–633. 10.1016/j.jmoldx.2013.05.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Beadling C, Neff TL, Heinrich MC, Rhodes K, Thornton M, Leamon J, et al. Combining highly multiplexed PCR with semiconductor-based sequencing for rapid cancer genotyping. J Mol Diagn. 2013;15: 171–176. 10.1016/j.jmoldx.2012.09.003 [DOI] [PubMed] [Google Scholar]
- 20.Kanagal-Shamanna R, Portier BP, Singh RR, Routbort MJ, Aldape KD, Handal BA, et al. Next-generation sequencing-based multi-gene mutation profiling of solid tumors using fine needle aspiration samples: promises and challenges for routine clinical diagnostics. Mod Pathol. 2014;27: 314–327. 10.1038/modpathol.2013.122 [DOI] [PubMed] [Google Scholar]
- 21.McCourt CM, McArt DG, Mills K, Catherwood MA, Maxwell P, Waugh DJ, et al. Validation of next generation sequencing technologies in comparison to current diagnostic gold standards for BRAF, EGFR and KRAS mutational analysis. PLoS ONE. Public Library of Science; 2013;8: e69604 10.1371/journal.pone.0069604 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Endris V, Penzel R, Warth A, Muckenhuber A, Schirmacher P, Stenzinger A, et al. Molecular diagnostic profiling of lung cancer specimens with a semiconductor-based massive parallel sequencing approach: feasibility, costs, and performance compared with conventional sequencing. J Mol Diagn. 2013;15: 765–775. 10.1016/j.jmoldx.2013.06.002 [DOI] [PubMed] [Google Scholar]
- 23.Cottrell CE, Al-Kateb H, Bredemeyer AJ, Duncavage EJ, Spencer DH, Abel HJ, et al. Validation of a next-generation sequencing assay for clinical molecular oncology. J Mol Diagn. 2014;16: 89–105. 10.1016/j.jmoldx.2013.10.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Nikiforova MN, Wald AI, Roy S, Durso MB, Nikiforov YE. Targeted next-generation sequencing panel (ThyroSeq) for detection of mutations in thyroid cancer. J Clin Endocrinol Metab. 2013;98: E1852–60. 10.1210/jc.2013-2292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pritchard CC, Salipante SJ, Koehler K, Smith C, Scroggins S, Wood B, et al. Validation and implementation of targeted capture and sequencing for the detection of actionable mutation, copy number variation, and gene rearrangement in clinical cancer specimens. J Mol Diagn. 2014;16: 56–67. 10.1016/j.jmoldx.2013.08.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Daber R, Sukhadia S, Morrissette JJD. Understanding the limitations of next generation sequencing informatics, an approach to clinical pipeline validation using artificial data sets. Cancer Genet. 2013;206: 441–448. 10.1016/j.cancergen.2013.11.005 [DOI] [PubMed] [Google Scholar]
- 27.Aisner DL, Deshpande C, Baloch Z, Watt CD, Litzky LA, Malhotra B, et al. Evaluation of EGFR mutation status in cytology specimens: an institutional experience. Diagn Cytopathol. 2013;41: 316–323. 10.1002/dc.21851 [DOI] [PubMed] [Google Scholar]
- 28.Bauml J, Mick R, Zhang Y, Watt CD, Vachani A, Aggarwal C, et al. Determinants of survival in advanced non—small-cell lung cancer in the era of targeted therapies. Clin Lung Cancer. 2013;14: 581–591. 10.1016/j.cllc.2013.05.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Frank R, Baloch ZW, Gentile C, Watt CD, LiVolsi VA. Multifocal Fibrosing Thyroiditis and Its Association with Papillary Thyroid Carcinoma Using BRAF Pyrosequencing. Endocr Pathol. 2013;25: 236–240. 10.1007/s12022-013-9289-0 [DOI] [PubMed] [Google Scholar]
- 30.Smigielski EM, Sirotkin K, Ward M, Sherry ST. dbSNP: a database of single nucleotide polymorphisms. Nucleic Acids Res. 2000;28: 352–355. 10.1007/s10549-007-9613-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Forbes S, Clements J, Dawson E, Bamford S, Webb T, Dogan A, et al. COSMIC 2005. Br J Cancer. 2006;94: 318–322. 10.1038/sj.bjc.6602928 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.1000 Genomes Project Consortium, Abecasis GR, Altshuler D, Auton A, Brooks LD, Durbin RM, et al. A map of human genome variation from population-scale sequencing. Nature. 2010;467: 1061–1073. 10.1038/nature09534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Robinson JT, Thorvaldsdóttir H, Winckler W, Guttman M, Lander ES, Getz G, et al. Integrative genomics viewer. Nat Biotechnol. 2011;29: 24–26. 10.1038/nbt.1754 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Roberts PJ, Stinchcombe TE. KRAS mutation: should we test for it, and does it matter? J Clin Oncol. 2013;31: 1112–1121. 10.1200/JCO.2012.43.0454 [DOI] [PubMed] [Google Scholar]
- 35.Srinivasan M, Sedmak D, Jewell S. Effect of fixatives and tissue processing on the content and integrity of nucleic acids. Am J Pathol. 2002;161: 1961–1971. 10.1016/S0002-9440(10)64472-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Spencer DH, Abel HJ, Lockwood CM, Payton JE, Szankasi P, Kelley TW, et al. Detection of FLT3 internal tandem duplication in targeted, short-read-length, next-generation sequencing data. J Mol Diagn. 2013;15: 81–93. 10.1016/j.jmoldx.2012.08.001 [DOI] [PubMed] [Google Scholar]
- 37.Loman NJ, Misra RV, Dallman TJ, Constantinidou C, Gharbia SE, Wain J, et al. Performance comparison of benchtop high-throughput sequencing platforms. Nat Biotechnol. 2012;30: 434–439. 10.1038/nbt.2198 [DOI] [PubMed] [Google Scholar]
- 38.Kinde I, Wu J, Papadopoulos N, Kinzler KW, Vogelstein B. Detection and quantification of rare mutations with massively parallel sequencing. Proc Natl Acad Sci USA. 2011;108: 9530–9535. 10.1073/pnas.1105422108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Casbon JA, Osborne RJ, Brenner S, Lichtenstein CP. A method for counting PCR template molecules with application to next-generation sequencing. Nucleic Acids Res. 2011;39: e81–e81. 10.1093/nar/gkr217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hiatt JB, Patwardhan RP, Turner EH, Lee C, Shendure J. Parallel, tag-directed assembly of locally derived short sequence reads. Nat Methods. 2010;7: 119–122. 10.1038/nmeth.1416 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Schmitt MW, Kennedy SR, Salk JJ, Fox EJ, Hiatt JB, Loeb LA. Detection of ultra-rare mutations by next-generation sequencing. Proc Natl Acad Sci USA. 2012;109: 14508–14513. 10.1073/pnas.1208715109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lou DI, Hussmann JA, McBee RM, Acevedo A, Andino R, Press WH, et al. High-throughput DNA sequencing errors are reduced by orders of magnitude using circle sequencing. Proc Natl Acad Sci USA. 2013;110: 19872–19877. 10.1073/pnas.1319590110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gargis AS, Kalman L, Berry MW, Bick DP, Dimmock DP, Hambuch T, et al. Assuring the quality of next-generation sequencing in clinical laboratory practice. Nat Biotechnol. 2012;30: 1033–1036. 10.1038/nbt.2403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Aziz N, Zhao Q, Bry L, Driscoll DK, Funke B, Gibson JS, et al. College of American Pathologists' laboratory standards for next-generation sequencing clinical tests. Arch Pathol Lab Med. 2015;139: 481–493. 10.5858/arpa.2014-0250-CP [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(XLSX)
Data Availability Statement
Raw deep sequencing cannot be provided because these data are derived from patient samples. Because of the Minimum Necessary Requirement of the HIPAA Privacy Rule these data may not be deposited into a public repository. A Supporting Information File accompanying the submission contains the de-identified results and interpretations of all clinical NGS tests included in our analysis next to the results of single-gene testing of the same specimen.