

Abstract
Ocean acidification and warming (OAW) are occurring globally. Additionally, at a more local scale the spreading of hypoxic conditions is promoted by eutrophication and warming. In the semi-enclosed brackish Baltic Sea, occasional upwelling in late summer and autumn may expose even shallow-water communities including the macroalga Fucus vesiculosus to particularly acidified, nutrient-rich and oxygen-poor water bodies. During summer 2014 (July–September) sibling groups of early life-stage F. vesiculosus were exposed to OAW in the presence and absence of enhanced nutrient levels and, subsequently to a single upwelling event in a near-natural scenario which included all environmental fluctuations in the Kiel Fjord, southwestern Baltic Sea, Germany (54°27 ´N, 10°11 ´W). We strove to elucidate the single and combined impacts of these potential stressors, and how stress sensitivity varies among genetically different sibling groups. Enhanced by a circumstantial natural heat wave, warming and acidification increased mortalities and reduced growth in F. vesiculosus germlings. This impact, however, was mitigated by enhanced nutrient conditions. Survival under OAW conditions strongly varied among sibling groups hinting at a substantial adaptive potential of the natural Fucus populations in the Western Baltic. A three-day experimental upwelling caused severe mortality of Fucus germlings, which was substantially more severe in those sibling groups which previously had been exposed to OAW. Our results show that global (OAW), regional (nutrient enrichment) and local pressures (upwelling), both alone and co-occurring may have synergistic and antagonistic effects on survival and/or growth of Fucus germlings. This result emphasizes the need to consider combined stress effects.
Introduction
Global climate change will expose marine populations to increased eutrophication and upwelling events at a regional scale and to ocean acidification and warming (OAW) at a more global scale [1]. For the Baltic Sea, current models predict an increase in pCO2 from currently almost 400 μatm to 1000 μatm, an increase in sea surface temperature by up to 5°C and considerable enhancements of eutrophication and hypoxia during the next 100 years [2]. The combined effects of these global change factors will likely affect Baltic ecosystems [3]. Anthropogenic activities, such as agriculture, lead to eutrophic conditions in the largest part of the Baltic Sea, including the southwestern part with Kiel Bight [4]. Eutrophication may further intensify due to increased precipitation and river runoff [5, 6]. Hypoxic conditions (< 2 mg O2 L-1) are predicted to increase in the Baltic Sea during this century as modelled by Meier and Andersson [7]. In Kiel Bay, seasonal oxygen depletion has been known for decades [8], but is likely to intensify further.
Macrophytes play a key role in ecosystem services by the retention of excessive nutrients [9, 10], acting as ‘nutrient filters’ [11] and providing the benthic ecosystem with oxygen [12]. A combination of global, regional and local stressors endangers macroalgae worldwide [13, 14]. Among these, the bladder wrack Fucus vesiculosus is a dominant perennial macroalga in the intertidal and shallow subtidal of the Western, Central and Eastern Baltic Sea [15]. As a foundation species, F. vesiculosus provides habitat and food for a large variety of invertebrate assemblages [16, 17]. A drastic decline in F. vesiculosus during the last five decades was observed, e.g. by almost 95% in Kiel Bay, Western Baltic Sea [18, 19]. The shoaling of F. vesiculosus was mainly assigned to the severe indirect effects of eutrophication [18, 20–22]. These are i.a. the increased turbidity [20], competition with ephemeral algae [23] and increased palatability [24]. Also, enhanced sedimentation reduces the attachment and survival of F. vesiculosus zygotes [25, 26]. Regarding direct effects, elevated nutrient concentrations positively affect photosynthesis and growth in adult F. vesiculosus [27].
As a consequence of eutrophication in the Baltic Sea, sedimentation and bacterial re-mineralization are enhanced, leading to seasonal hypoxia and hypercapnia in sub-surface waters [28]. Further spread of (periodic) hypoxic areas in the Baltic Sea is predicted until the end of the 21st century due to the interplay between eutrophication and lower oxygen solubility under warming [7]. Hypoxia induced mortality of benthic organisms was reviewed by Gray et al. [29] and recognized at a Baltic [30] and a global scale [31]. Also, higher frequencies of local upwelling in the last decades have been documented in the Baltic Sea [32].
In addition to hypoxia, warming was observed to impact growth, survival and photosynthetic efficiency in F. vesiculosus, while acidification showed weaker effects in the early [33] and adult life-stage [34]. As a consequence of warming, poleward range shifts of seaweed populations have been observed worldwide [14, 35, 36]. Also in the Baltic Sea, northward range shifts of F. vesiculosus populations have been observed and are predicted to continue during the 21st and 22nd century [37]. In contrast, ocean acidification was reported to increase growth in non-calcifying macroalgae [27, 38]. Assumed reasons are enhanced availability of CO2 to saturate the carbon demand during photosynthesis or the saved energy when carbon concentrating mechanisms are downregulated [39]. However, physiological responses of the macroalga Macrocystis pyrifera to acidified conditions showed that increased CO2 conditions did not affect growth or photosynthesis [40]. Early life-stage macroalgae may be particularly threatened by global change [41]: Warming lowers the germination success of F. vesiculosus at 25°C [42] and reduces survival in F. serratus germlings more severely than in adults [43]. Enhanced sedimentation caused by eutrophication is more detrimental for young [23, 44] than for adult Fucus [15, 45]. Despite recognized differences in stress sensitivity among life-stages of a species, past research has mainly focussed on adult forms [46, 47].
Adaptation of marine populations to global change is favoured by genetic diversity [48, 49]. Conversely, low intraspecific genetic diversity and high phenotypic plasticity and gene flow restrict adaptation to environmental stress [50–53]. The increased tolerance of the eelgrass Zostera marina with higher genetic diversity to a summer heat wave [54] showed that genetic diversity may buffer warming stress at the population level [55].
Baltic F. vesiculosus populations show reduced genetic variation compared to Atlantic populations probably due to isolation and bottlenecks as well as the eroded genetic variation due to selection [56]. It has been argued that environmental stress, e.g. osmotic stress in the brackish Baltic, lower genetic diversity and limited dispersal capacity of F. vesiculosus gametes [57] may favour local extinctions [58]. However, this study is the first one to test genetic variation in Baltic F. vesiculosus with regard to the sensitivity towards OAW, nutrient enrichment and subsequent hypoxia.
The aim of the present study was to investigate (1) how OAW, (2) nutrient enrichment and (3) upwelling events affect the survival and growth of F. vesiculosus germlings, (4) how OAW interacts with simultaneous exposure to high nutrient concentrations, (5) how these treatments modify hypoxia sensitivity and (6) whether sibling groups vary in their tolerance towards these environmental parameters. Our experimental concept, hence, consisted in the exposure of genetically different sibling groups of F. vesiculosus germlings to increased temperature, pCO2 and nutrient conditions and to a final upwelling event while maintaining the natural variations.
Material and Methods
Collection, gamete acquisition, experimental design
A total of 64 fertile F. vesiculosus were collected in a wave exposed area with mixed hard substrate and sand bottom in the southwestern Baltic Sea (Bülk, Germany, 54°27.327 ´N, 10°11.977 ´W) in mid-June 2014. To avoid the collection of siblings and ensuring for genetic variability, individuals sampled were distanced by at least 2 meters, which is the estimated maximum dispersal distance of most F. vesiculosus gametes [57]. After collection, algae were transported to the lab in cooler boxes. Fertile receptacles were cut from these dioecious algae and gender was determined (46 females, 18 males) under the microscope at 100 x magnification (Olympus BH-2). Receptacles were rinsed with tap water, blotted dry and stored in the dark for 6 days at 14°C. Before gamete release, all receptacles from one female and one male individual (i.e. one parental pair) were put in a small dish. Gamete release followed by egg fertilisation was induced by immerging receptacles into sand-filtered seawater (15–16 psu) and exposing them to light (110 μmol photons m-2 s-1) for 3 hours. In this way, gametes were obtained from16 parental pairs. No specific permits were required for this study, the location is not privately-owned or protected and the study did not involve an endangered or protected species.
One mL of homogeneously suspended fertilised eggs was pipetted onto the upper surface of each of 2 x 2 cm sandstone cubes. Each cube with its settled germlings represented one experimental population. 16 different populations, each composed of germlings stemming from one parental pair, were thus produced. Culture and monitoring of germlings took place in a room with windows approximating natural light conditions during 3 weeks with weekly water exchange (15–16 psu) at 15°C until introducing them to the Kiel Outdoor Benthocosms (KOB). This facility maintains the natural in situ fluctuations and simultaneously allows manipulating environmental variables (e.g. temperature, pCO2) on top of these fluctuations, i.e. “delta treatments”. Target temperatures were obtained and maintained by computer controlled heaters and coolers, while acidification was achieved by increasing the atmospheric pCO2 within the enclosed head space above the tanks by injecting pre-mixed gas to maintain an atmospheric pCO2 of 1100 μatm. Details of the experimental set-up of the KOB and the logged conditions in the tanks are given in Wahl et al. [59].
OAW x nutrient and upwelling treatment
The OAW x nutrient experiment took place during 2 summer months from mid-July until mid-September 2014. One PVC box (70 cm x 40 cm x 12 cm) was placed within each of the 12 Benthocosm main tanks, each of them containing all 16 experimental populations of germlings. Since the upper rim of the lid-less boxes was a few centimetres above the water surface, the water body within the PVC boxes was separated from the water body of the main tank but open to the atmosphere. Thus, the boxes experienced the same treatments as the main tank regarding OAW but were insulated from gene flow from the adult Fucus population in the main tank. Twice a week the water of the PVC box was exchanged by water from the main tank which had been filtered through a 50 μm mesh to prevent the introduction of F. vesiculosus eggs (100 μm diameter). As the single-factor and combined effects of temperature and pCO2 on F. vesiculosus germlings have been investigated previously [33], we combined warming and acidification into a single factor (OAW) in the present run. The two fully crossed factors OAW and nutrients were applied at two levels each (ambient and future). Ambient and predicted future levels of OAW and nutrients were simulated by adding the expected shift to the natural fluctuations of the ambient fjord conditions as delta treatments [59].
The “ambient” condition represents the natural fjord conditions transported into the main tanks of the KOB by a continuous flow-through (1 tank-volume per day, i.e. 1500 L/ 24 h) of Kiel Fjord water pumped from 1 m depth. “Future” conditions were simulated by adding 5°C to the actual temperature of the Kiel Fjord and by increasing the pCO2 concentration in the hooded headspace of the tanks to 1100 μatm according to the predictions for the year 2110 for the Baltic Sea [2]. The bi-weekly nutrient enrichment (2x ambient) was achieved by doubling the “ambient” concentration which was taken as the seven years (2006–2013) mean for each specific date of nutrient addition (Table 1, S1 Fig). NaNO2 (Merck, Germany), NaNO3 (Carl Roth, Germany) and H2NaO4P.H2O (ACROS organics, Germany) were dissolved in fjord water 10 minutes before addition to the nutrient treatments. The ratio P: N of the Kiel Fjord is approximately 1: 1.5 and does not match the Redfield Ratio, probably due to the nutrient input of the nearby located river Schwentine. Additionally, high organism activity during summer months decreases the overall nutrient availability in shallow water. This ambient P: N ratio was not altered when NO3, NO2 and PO4 concentrations (μmol L-1) were doubled under “future” conditions. Our analysis on CN ratio in Fucus tissue (see result section below) showed no differences under “future” conditions, indicating that carbon did not become a limiting factor under nutrient enrichment.
Table 1. Nutrient concentration under ambient and future conditions.
“Ambient” nutrient concentrations of PO4, NO2, NO3 (μmol L-1) for the respective summer months and “future” nutrient concentrations (μmol L-1).
| July | August | September | ||||
|---|---|---|---|---|---|---|
| Ambient | Future | Ambient | Future | Ambient | Future | |
| PO4 | 0.46 | 0.93 | 0.59 | 1.19 | 1.06 | 2.11 |
| NO3 | 0.53 | 1.05 | 0.77 | 1.54 | 1.27 | 2.54 |
| NO2 | 0.18 | 0.36 | 0.20 | 0.40 | 0.22 | 0.44 |
The four treatment combinations, each replicated three times, thus were: OAW- N- (ambient OAW & ambient nutrients), OAW- N+ (ambient OAW & high nutrients), OAW+ N- (future OAW & ambient nutrients) and OAW+ N+ (future OAW & high nutrients). These treatment combinations were regularly distributed among the 12 experimental units.
The upwelling experiment was performed immediately after the end of the OAW x nutrient treatment phase, i.e. when all the treatments in the tanks were set back to "ambient". During three days, hypoxic fjord water from 15 m depth (O2 = 2.75 ± 0.41 mg L-1, T = 16.52 ± 0.33°C, pH = 7.40, Sal = 22.8) was pumped as flow-through into the KOB and, via a bypass into the germlings boxes continuously during 3 days. This experiment was meant to assess the effect of the compound factor "upwelling" on sibling groups pre-conditioned by the foregoing treatment combinations (OAW x nutrient enrichment). Upwelled water in this region and this season usually is characterized by lower oxygen, higher salinity and higher nutrients than surface water.
Measurement of abiotic factors
pH and temperature of the main tank of the KOB were measured daily with a calibrated hand-held sensor (pH, Mettler Toledo GmbH, Giessen, Germany) while oxygen was measured with a Multi WTW Oxy 3515 (oxygen, Wissenschaftlich Technische Werkstätten, Weilheim, Germany). Salinity was measured with a portable conductivity meter (WTW Cond 3110 + Tetra Con 325, Wissenschaftlich Technische Werkstätten, Weilheim, Germany). Additionally, temperature, pH, O2 and salinity were continuously logged at 10 min time intervals (GHL Advanced Technology, Kaiserslautern, Germany). During the hypoxia experiment, O2 and temperature within the PVC boxes were logged every 10 min using the Multi WTW Oxy 3515. Samples for nutrient concentrations were taken from the KOB before each water exchange in the PVC boxes as initial concentration. For this, water samples were immediately filtrated through a 0.45 μm Minisart syringe filter (Sartorius) in 10 mL tubes, stored at -20°C and measured with a QuAAtro nutrient analyzer [60] (SEAL Analytical; S1 Fig). More details about the measurement of the abiotic variables in the KOB are described by Wahl et al. [59].
Abiotic conditions
During the OAW x nutrient experiment, the bi-weekly measured oxygen concentrations (mean = 8.91 ± 0.38 mg L-1) varied in the low OAW treatment with a minimum of 8.01 ± 0.23 mg L-1 and a maximum of 9.44 ± 0.16 mg L-1 (mean ± SD) and within the high OAW treatment (mean = 7.76 ± 0.56 mg L-1) with a minimum of 6.75 ± 0.56 mg L-1 and a maximum of 8.72 ± 0.21 mg L-1. Day-night fluctuations measured in two hours intervals showed that O2 concentrations varied between a minimum of 7.64 mg L-1 at 4:30 am and a maximum of 9.97 mg L-1 at 16:30 pm under ambient conditions.
Ambient temperatures (factor level ‘ambient’) (mean = 19.26 ± 2.38°C) varied between bi-weekly measurements with a minimum of 15.37 ± 0.19°C and a maximum of 22.7 ± 0.08°C. Elevated temperatures (factor level ‘future’) (mean = 23.64 ± 2.68°C) varied between a minimum of 19.53 ± 0.52°C and a maximum of 27.47 ± 0.26°C, which occurred during a natural summer heat wave (mean ± SD) (Fig 1). Temperatures during day and night fluctuated between a minimum of 16.1°C at 10:30 pm and a maximum of 17.7°C at 8:30 am under ambient conditions.
Fig 1. Temperature, pH and salinity during the OAW x nutrient experiment.

(A) Temperature (°C), (B) pH (NBS, National Bureau of Standards) under the two treatment combinations ambient (OAW-), warmed and acidified (OAW+) and (C) salinity (psu) at the ambient treatment (OAW-). Data are the mean ±SD of three replicates in each treatment combination.
Response variables
Growth
For growth measurements, digital images were taken of 10–15 randomly chosen individual germlings per population at 40 x magnification (SteREO Discovery. V8 –Carl Zeiss Jena GmbH) similar to Steen and Scrosati [61]. Measured individuals were chosen randomly, since germlings were too small for labelling. The projected side-view of the single germlings was measured with the image analysis software Image J 1.45s (National Institutes of Health, USA) and the mean of germlings’ area of the perpendicular projection was calculated for each population. Germlings’ area was measured at the beginning (area t0) and after 8 weeks (area t) at the end of the OAW x nutrient experiment. Relative growth rate (RGR) in % d-1 was calculated as exponential growth:
Where Δt is the time period between t0 and t in days.
Survival
Germling number was counted under a binocular at 25x magnification between the start (number t0) and after 8 weeks (number t) of the OAW x nutrient experiment. Survival of germlings was expressed as the percent of surviving germlings and calculated as:
For determining survival during the final upwelling experiment, the germling number at the end of the preceding OAW x nutrient experiment was set as t0 and number t was the germling number after the upwelling treatment.
Log-effect ratio
Log effect ratios were performed to show the direction and the strength of the sibling groups’ phenotypical responses to the different OAW and nutrients enhancements relative to the respective ambient conditions. These differences among sibling groups’ responses were determined in order to assess whether higher genetic diversity also increases the variance in responses. Sibling groups’ sensitivity to high OAW and N were calculated separately as the log effect ratios of growth or survival under future relative to actual conditions, as:
at ambient and high nutrient conditions and:
at ambient and high OAW. The same procedure was used for calculating the log effect ratios for survival. Negative growth rates observed in 3 populations were attributable to the mortality of the bigger sized individuals and a decrease in mean area t compared to t0. To avoid negative values for log effect ratio analysis, growth was measured as ratios of area t and area t0 as:
CN ratios
Germlings of five sibling groups (1, 3, 7, 12 and 14) of the two treatment combinations OAW- N- and OAW- N+ were pooled for CN analysis. For the analysis of carbon and nitrogen content, freeze-dried algal material was ground to powder and three subsamples of 2 mg from each treatment was packed and loaded into tin cartridges (6×6×12 mm). Then, the packages were combusted at 950°C and the absolute C and N contents in % dry weight (% DW) were automatically quantified in an elemental analyser (Elementar Vario EL III, Germany) using acetanilide as standard according to Verardo et al. [62].
Statistical Analysis
Growth (RGR, % d-1) and survival (%) were analysed using a split-plot ANOVA with the fixed factors ‘OAW’ (with two levels: OAW- and OAW+) and ‘Nutrients’ (with two levels: N- and N+) as well as the random factors ‘Mesocosm’ (with 12 levels) and ‘Sibling Group’ (with 16 levels). ‘Mesocosm’ was nested in the ‘OAW’ x ‘Nutrients’ interaction. This model allowed us to analyse the influence of the fixed factors. Furthermore, we were able to identify possible random-by-fixed factor interactions between ‘Sibling Group’, ‘OAW’ and ‘Nutrients’. Survival data were arcsine transformed prior to the analysis to overcome data truncation. Differences in carbon and nitrogen contents (% DW) as well as in CN ratios between five single sibling groups were identified in a further split-plot ANOVA with the fixed factor ‘Nutrients’ (two levels: see above), which was combined with OAW only, and the random factors ‘Mesocosm’ (12 levels) and ‘Sibling Group’ (16 levels), while ‘Mesocosm’ was nested in ‘Nutrients’. Split-plot ANOVAs were performed by using Satterthwaite’s method for denominator synthesis [63], which calculates appropriate error terms for the F-ratios of the respective effects. Normality of errors and homogeneity of variances were verified by using residual plots (Q-Q Plot and Standardized Residuals Plot, respectively) in STATISTICA and R. The significance level of all analyses was α = 0.05. Post-hoc tests were performed using Tukey’s HSD. All statistical analyses were conducted using the software STATISTICA v. 12 [64] and R.
Results
OAW x nutrient experiment (mid-July–mid-September 2014)
Growth and survival
Under ambient temperature, pCO2 and nutrient conditions, F. vesiculosus germlings’ relative growth rate (RGR) and survival was 2.86 ± 0.37% d-1 (Fig 2A) and 80.34 ± 3.70% (Fig 2B), respectively. Under ambient temperature and pCO2 conditions, the addition of nutrients (i.e. OAW-N+) did not change RGR and survival (Fig 2). In contrast, high OAW conditions under ambient nutrient conditions (OAW+N-) reduced RGR and survival by about 50%; when RGR was 1.37 ± 0.21% d-1 and survival was 44.48 ± 20.03% (mean, ± SD, n = 3, Table 2). The addition of nutrients almost entirely compensated the negative impact of OAW as reflected in a RGR of 2.52 ± 0.15% d-1 and survival of 73.49 ± 3.49%. The significant interaction between the factors OAW and nutrients regarding growth rate reflects this compensation effect of the nutrient treatment (Split-plot ANOVA, p < 0.05, Table 2A).
Fig 2. Growth and survival of F. vesiculosus germlings during the OAW x nutrient experiment.
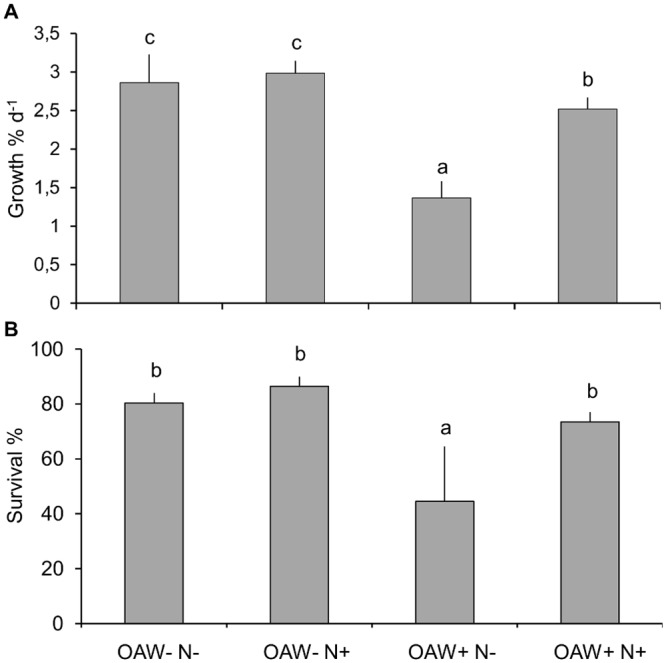
(A) Growth (% d-1) and (B) survival (%) (mean +SD, n = 3, 8–9 weeks) during summer 2014 at the four treatment levels OAW-N-, OAW-N+, OAW+N-, OAW+N+. Means were calculated from 16 sibling groups for each treatment ‘OAW’ and ‘Nutrient’ treatment combination. Different letters above the bars indicate significant differences (p-value < 0.05) between the treatments after Tukey’s HSD.
Table 2. OAW and nutrients effects on growth and survival.
Results from split-plot ANOVA with the fixed factors ‘OAW’ and ‘Nutrients’ and the random factors ‘Mesocosm’ and ‘Sibling group’. Effects are shown for (A) growth rates (% d-1) and (B) survival during the OAW x nutrient experiment and (C) survival during the upwelling experiment. Df: degrees of freedom, SS: sums of squares and MS: mean squares. ‘Den. Syn. Error df’ and ‘Den. Syn. Error MS’ describe the denominator synthesis of degrees of freedom and mean squares, respectively. (Datasets of area and growth values can be found in the PANGAEA dataset).
| Source of variation | df | SS | MS | Den. Syn. Error df | Den. Syn. Error MS | F-value | p-value |
|---|---|---|---|---|---|---|---|
| (A) Growth rates OAW x nutrient experiment | |||||||
| OAW | 1 | 47.659 | 47.659 | 6.156 | 1.188 | 40.111 | < 0.001 |
| Nutrients | 1 | 20.468 | 20.468 | 8.469 | 1.428 | 14.335 | < 0.05 |
| OAW x Nutrients | 1 | 13.473 | 13.473 | 8 | 1.339 | 10.055 | 0.013 |
| Mesocosm (OAWxNutrients) | 8 | 10.720 | 1.340 | 135 | 0.389 | 3.443 | 0.001 |
| Sibling | 15 | 27.120 | 1.808 | 5.276 | 0.325 | 5.558 | 0.030 |
| Sibling x OAW | 15 | 3.561 | 0.237 | 135 | 0.389 | 0.610 | 0.863 |
| Sibling x Nutrient | 15 | 7.156 | 0.477 | 135 | 0.389 | 1.226 | 0.260 |
| Error | 135 | 52.542 | 0.389 | ||||
| (B) Survival OAW x nutrient experiment | |||||||
| OAW | 1 | 4.069 | 4.069 | 14.366 | 0.867 | 4.694 | 0.048 |
| Nutrients | 1 | 2.903 | 2.903 | 7.061 | 0.513 | 5.656 | 0.049 |
| OAW x Nutrients | 1 | 1.272 | 1.272 | 8 | 0.535 | 2.376 | 0.162 |
| Mesocosm (OAWxNutrients) | 8 | 4.281 | 0.535 | 135 | 0.163 | 3.284 | 0.002 |
| Sibling | 15 | 11.786 | 0.786 | 12.535 | 0.473 | 1.662 | 0.186 |
| Sibling x OAW | 15 | 7.420 | 0.495 | 135 | 0.163 | 3.036 | < 0.001 |
| Sibling x Nutrient | 15 | 2.117 | 0.141 | 135 | 0.163 | 0.866 | 0.603 |
| Error | 135 | 21.998 | 0.163 | ||||
| (C) Survival Upwelling experiment | |||||||
| OAW | 1 | 4.907 | 4.907 | 8.398 | 0.550 | 8.914 | 0.017 |
| Nutrients | 1 | 0.322 | 0.322 | 5.284 | 0.423 | 0.761 | 0.421 |
| OAW x Nutrients | 1 | 2.913 | 2.913 | 8 | 0.516 | 5.647 | 0.045 |
| Mesocosm (OAWxNutrients) | 8 | 4.127 | 0.516 | 135 | 0.164 | 3.150 | 0.003 |
| Sibling | 15 | 10.711 | 0.714 | 3.504 | 0.105 | 6.793 | 0.052 |
| Sibling x OAW | 15 | 2.976 | 0.198 | 135 | 0.164 | 1.212 | 0.270 |
| Sibling x Nutrient | 15 | 1.057 | 0.070 | 135 | 0.164 | 0.430 | 0.968 |
| Error | 135 | 22.106 | 0.164 | ||||
Sibling groups differences in growth
RGR of the 16 sibling groups differed significantly (Split-plot ANOVA, factor: sibling group, p-value < 0.05, Table 2A). Nutrient enrichment at ambient temperature and CO2 enhanced growth significantly only of the sibling groups 3, 4 and 7 (Fig 3B). In contrast, nutrient enrichment under high OAW conditions enhanced growth in most sibling groups, notably in the groups 3, 4, 5, 6, 7, 8, 9, 11, 12, 13, 14, 15 and 16. High OAW at ambient nutrient conditions decreased growth in most sibling groups, i.e. 1, 2, 3, 4, 5, 6, 7, 8, 9, 11, 12, 15 and 16. In contrast, under nutrient enrichment the negative effect of high OAW was mitigated so that growth was significantly decreased only in the 6 sibling groups 2, 4, 7, 8, 10 and 12 (Fig 3A). The factor ‘sibling group’ did not interact significantly with ‘OAW’ or with ‘nutrients’ (Split-plot ANOVA, sibling group x OAW, sibling group x nutrients, p-value > 0.05, Table 2A).
Fig 3. Log effect ratios of growth in 16 sibling groups during the OAW x nutrient experiment.

Log effect ratios (mean ± 95% confidence intervals, n = 3) of (A) OAW effects were calculated as log (growth OAW+/ growth OAW-) under ambient and high nutrient conditions (N-, N+) and (B) log effect ratios for nutrient effects calculated as log (growth N+/ growth N-) at ambient and high OAW (OAW-, OAW+).
Sibling groups differences in survival
OAW+ at ambient nutrient concentrations decreased survival significantly in 5 out of 16 sibling groups, notably 3, 7, 12, 14 and 15. The different survival responses to increased OAW of sibling groups are reflected in the significant interaction of the factors ‘OAW’ x ‘Sibling group’ (Split-plot ANOVA, p-value < 0.001, Table 2B). Nutrient enrichment tended to mitigate the negative effect of OAW and this buffering influence was significant in the 4 sibling groups 7, 12, 14 and 15 (Fig 4A). Nutrient enrichment under ambient temperatures and CO2 did not affect sibling groups’ survival. In contrast, nutrient enrichment under warming and acidification generally tended to improve survival and significantly enhance survival in the groups 3, 7, 12 and 15 (Fig 4B, Table 2B).
Fig 4. Log effect ratios of survival in 16 sibling groups during the OAW x nutrient experiment.

Log effect ratios (mean ± 95% confidence intervals, n = 3) of (A) OAW effects were calculated as log (survival OAW+/ survival OAW-) under ambient and high nutrient conditions (N-, N+); (B) log effect ratios for nutrient effects were calculated as log (survival N+/ survival N-) at ambient and high OAW (OAW-, OAW+).
CN ratio
Germling carbon and nitrogen content (% DW) as well as the CN ratio did not differ significantly among the nutrient treatments (Split-plot ANOVA, p-value > 0.05, Table 3). The CN ratio was 10.56 ± 1.13 under ambient nutrient conditions and 7.18 ± 3.06 (mean ± SD) under high nutrient conditions.
Table 3. Nutrient effect on carbon and nitrogen content (% DW) and on the CN ratio.
Split-plot ANOVA with the fixed factor ‘Nutrient’ and the random factors ‘Mesocosm’ and ‘Sibling group’. Effects of the nutrient treatment were analysed for (A) the carbon content (% DW), (B) the nitrogen content (% DW) and (C) the CN ratio. Df: degrees of freedom, SS: sums of squares and MS: mean squares. ‘Den. Syn. Error df’ and ‘Den. Syn. Error MS’ describe the denominator synthesis of degrees of freedom and mean squares, respectively.
| Source of variation | df | SS | MS | Den. Syn. Error df | Den. Syn. Error MS | F-value | p-value |
|---|---|---|---|---|---|---|---|
| (A) Carbon content | |||||||
| Nutrients | 1 | 36.563 | 36.563 | 1.998 | 17.839 | 2.05 | 0.289 |
| Mesocosm (Nutrients) | 4 | 79.878 | 19.969 | 16 | 15.482 | 1.290 | 0.315 |
| Sibling | 4 | 145.584 | 36.396 | 4 | 13.352 | 2.726 | 0.177 |
| Sibling x Nutrients | 4 | 53.407 | 13.352 | 16 | 15.482 | 0.862 | 0.507 |
| Error | 16 | 247.718 | 15.482 | ||||
| (B) Nitrogen content | |||||||
| Nutrients | 1 | 6.676 | 6.676 | 3.669 | 38.545 | 0.173 | 0.700 |
| Mesocosm (Nutrients) | 4 | 159.792 | 39.948 | 16 | 5.462 | 7.314 | 0.002 |
| Sibling | 4 | 16.647 | 4.162 | 4 | 4.059 | 1.025 | 0.491 |
| Sibling x Nutrients | 4 | 16.235 | 4.059 | 16 | 5.462 | 0.743 | 0.576 |
| Error | 16 | 87.387 | 5.462 | ||||
| (C) CN ratio | |||||||
| Nutrients | 1 | < 0.001 | < 0.001 | 4.488 | 333.810 | < 0.001 | 1 |
| Mesocosm (Nutrients) | 4 | 1226.652 | 306.663 | 16 | 42.265 | 7.256 | 0.002 |
| Sibling | 4 | 297.260 | 74.315 | 4 | 69.412 | 1.071 | 0.474 |
| Sibling x Nutrients | 4 | 277.649 | 69.412 | 16 | 42.265 | 1.642 | 0.212 |
| Error | 16 | 676.237 | 42.265 | ||||
Upwelling experiment (Mid-September 2014)
O2, salinity and temperature conditions
The mean (± SD) of O2 concentration during the experimental upwelling in the PVC boxes was 2.71 ± 0.37 mg L-1 (range 1.44 mg L-1 to 5.61 mg L-1). Mean temperature during the hypoxia experiment was 16.35 ± 0.29°C (range 15.8°C to 17.5°C). pH was 7.40 and salinity was 22.8 psu in the deep water measured once on the day before the start of the hypoxia treatment.
Survival
In all sibling groups with different treatment histories, the three days of hypoxia reduced germling survival significantly. Survival was highest for germlings pre-treated at ambient conditions (OAW-N-) and lowest for germlings previously treated at high OAW and ambient nutrients (OAW+N-) (Fig 5). Thus, high OAW significantly doubled the sensitivity to subsequent hypoxia as compared to a non-warmed, non-acidified regime (Split-plot ANOVA, OAW, p-value < 0.05, Fig 5, Table 2C). Nutrient addition during the preceding experiment enhanced sensitivity to hypoxia in germlings stemming from a regime of ambient temperature and CO2 conditions but not for those from a OAW+ regime as reflected in the significant interaction between ‘OAW’ and ‘Nutrients’ (Split-plot ANOVA, OAW x Nutrients, p-value < 0.05, Fig 5, Table 2C).
Fig 5. Survival of F. vesiculosus germlings after 3 days of hypoxic upwelling.

Survival (%, mean +SD, n = 3) in 16 sibling groups of F. vesiculosus germlings previously treated under the four treatment combinations of the OAW x nutrient experiment: OAW-N-, OAW-N+, OAW+ N-, OAW+N+. Different letters above the bars indicate significant differences (p-value < 0.05) between the treatments after Tukey’s HSD.
Discussion
The simulated OAW as expected for 2110 strongly reduced survival and growth of F. vesiculosus germlings. The analysis of the single factors warming and acidification in a previous study at the KOB showed that warming was the main driver of mortality of F. vesiculosus germlings, while acidification played a minor role [33]. At temperatures > 27°C, reduction in growth and photosynthetic efficiency were observed in adult F. vesiculosus, finally resulting in necrosis [34]. Reduced growth of germlings under heat stress (> 25°C) was also observed in this work. This shows that the upper limits of thermotolerance of F. vesiculosus performance are similar in early and adult life-stages.
However, in our multi-factorial design, the negative effects of warming on germling survival and growth rates were strongly mitigated by high nutrient concentrations. Earlier studies on Baltic adult F. vesiculosus have shown that nutrient enrichment increases nutrient uptake [65] and enhances photosynthetic efficiency [27]. Similar responses were also observed in other algal species. Ulva rigida cultured under nutrient enrichment reacted with higher nitrogen uptake, higher nitrate reductase activity and higher growth rates. Moreover, the nitrogen reductase activity was enhanced under future (1000 μatm) compared to ambient pCO2 (400 μatm) conditions [66]. Similarly, high pCO2 enhanced nitrogen assimilation in the brown alga Hizikia fusiforme [67] and may have decreased the relative investment in the nitrogen-intensive protein biosynthesis [67]. Consequently, nitrogen may have been freed for other processes, such as growth and nitrogen storage [68]. Possibly, the F. vesiculosus germlings in our experiments also took up more nitrogen under nutrient enrichment when additional CO2 was provided under acidified (and warmed) conditions, resulting in increased growth and survival. Our experimental design did not allow for disentangling the different possible mechanisms of mitigating effects of nutrients on either warming or acidification. Photosynthesis is regarded as one of the most heat sensitive metabolic activities in the plant cell [69, 70], with at least three major heat-stress sensitive sites in the photosynthetic machinery: the photosystems (mainly photosystem II with its oxygen-evolving complex), the ATP generating and the carbon assimilation processes [71]. Moreover, respiration rates are increased under warming [72]. As our response variables growth and survival represent responses integrating over many metabolic processes, several compensatory effects caused by high nutrient levels appear possible. In conclusion, nutrient enrichment compensated to some degree the severe negative effects of future heat stress on F. vesiculosus germlings, which may be further mitigated by higher carbon availability under acidified conditions. However, such direct beneficial effects of nutrient enrichment may be overridden by indirect detrimental effects of eutrophication (such as increases in water turbidity, sedimentation, grazing and abundance of epibiotic filamentous algae) at field conditions, as reviewed by Berger et al. [45]. During the OAW x nutrient experiment, the epibiota under ambient and enriched nutrient conditions have not been determined. Regular filtration of the water content of our experimental boxes kept the fouling load relatively low under both, ambient and high nutrient conditions, assuming that epibiota had no strong effects on the F. vesiculosus germlings.
Increased nitrogen uptake is accompanied by higher CO2 uptake (even at ambient pCO2), hence a constant CN ratio is maintained [66]. Our findings show that the CN ratio in F. vesiculosus germlings was lower in the high nutrient treatment. Although this difference was not significant, it suggests higher uptake rates of nitrogen under nutrient enrichment. Since the nitrogen concentration in Baltic F. vesiculosus thalli is lowest in summer [73], nutrient enrichment effects may be most conspicuous in this season. This may have contributed to the observed stress-mitigating effect of nutrient enrichment in F. vesiculosus germlings. The buffering of stress impact by additional resources was also observed in juvenile blue mussel Mytilus edulis, when high food conditions enhanced the tolerance to ocean acidification [74].
The different sibling groups showed high variations in survival under warming and acidification, indicating the enhanced potential for adaptation in genetically diverse populations [48]. The crucial role in genetic variation for recovery from disturbances has also been reported in estuarine macrophytes [75] allowing for adaptation under global change stress [76].
The three day hypoxia experiment in the KOB simulating an upwelling event induced substantial germling mortality. During a local upwelling event, deep water with low oxygen concentration, low temperatures an increased pCO2 and high salinity is shoaling [77, 78]. In our upwelling treatment, temperature did not decrease considerably (16.41 ± 0.33°C) compared to previous values of 19.26 ± 2.38°C. Likewise, salinity (22.8 psu) did increase only slightly relative to the previous condition (Fig 1C) and the elevated CO2 (as associated with hypoxic upwelling) has minor effects on germling survival [33] (Fig 1C). This leaves the low oxygen concentration (2.75 ± 0.41 mg L-1) during the upwelling event as the most likely driver of germling mortality. Mortality during the three days of upwelling was considerably higher than during the two months of the preceding experiment, illustrating the high susceptibility of F. vesiculosus germlings to hypoxia. This susceptibility is probably due to reduced respiration rates under dark conditions, as it was also observed under hypoxic conditions in Cladophora vagabunda and Gracilaria tikvahiae [79]. Reduced respiration rates are accompanied by decreased provision of ATP and biosynthetic precursors leading to higher stress sensitivity [80] and to a reduced metabolism [79]. Susceptibility to hypoxia impacts was highest on germlings which previously experienced warmed and acidified conditions. Thus, the impact of hypoxic upwelling events in the future may be amplified by synchronous OAW. The assumed increased respiration under warming [72] may have further increased the O2 debt, which could not be balanced under hypoxic conditions. Consequently, F. vesiculosus germlings grown under high compared to ambient temperatures were less tolerant to hypoxia. Thus, germlings grown under high nutrient levels experienced higher mortality under hypoxia compared to those grown at low nutrient levels. Zou et al. [68] demonstrated that under high-nitrogen conditions, respiration was enhanced by high CO2 compared to ambient CO2 conditions in the macroalga H. fusiforme. Consequently, in algae growing under nutrient enrichment increased respiration might be necessary to support higher maintenance demands (e.g. due to increased RUBISCO contents) and greater uptake of extra nitrogen [68]. This nutrient-driven higher metabolism may have rendered these germlings more susceptible to hypoxia.
In summary, the responses to hypoxia depended on the preceding OAW x nutrient treatments we applied. This suggests that there are different protective mechanisms in F. vesiculosus germlings that vary with the type of stressor. Future expansions of hypoxic areas in the Baltic Sea [7] will have severe effects on F. vesiculosus recruitments, as observed in this experiment, as well as on the benthic community in general [81]. We demonstrated that the net impact of global change including warming, acidification, eutrophication and hypoxia may depend on the interaction among these global and regional factors. This finding underscores the importance for analysing the combined effects of multiple stressors and their interconnectivity for accurate predictions of future scenarios [3]. Moreover, the indirect effects of global change may be more significant than the direct effects [13]. Scaling up multiple stressors is crucial for predicting the fate of F. vesiculosus populations [82].
Supporting Information
Nutrient concentrations within the experimental germling boxes before (A, B) and after (C, D) the bi-weekly addition of the nutrients (PO4, NO3, NO2) as well as initial NH4 conditions in μmol L-1 in July and August. Initial nutrient concentrations were measured six times per month in the main KOB tank before the water addition to the boxes. Nutrient concentrations after additions were determined by adding the sum of the initial and additional nutrient concentration.
(TIFF)
Acknowledgments
We gratefully thank Claudia Starke and Maysa Ito for their participation at the experiment and Björn Buchholz for the maintenance of the Kiel Outdoor Benthocosms. We thank all members of the BIOACID II consortium ‘Benthic Assemblages’. The research was supported by the Project BIOACID II of the German Federal Ministry of Education and Research (BMBF; FKZ 03F0655, A). We thank Trystan Sanders for the linguistic revision and three reviewers for valuable comments.
Data Availability
All data are uploaded in the repository PANGAEA (Data Publisher for Earth & Environmental Science). doi: http://doi.pangaea.de/10.1594/PANGAEA.853938.
Funding Statement
The research was funded by the Project BIOACID II of the German Federal Ministry of Education and Research (BMBF; FKZ 03F0655, A). They support this project, the experimental setup and all accompanied costs.
References
- 1.IPCC. Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA: 2013. [Google Scholar]
- 2.BACC II. Second Assessment of Climate Change for the Baltic Sea Basin. Geesthacht GmbH, Germany: International Baltic Earth Secretariat, Helmholtz-Center, 2015. 10.1007/978-3-319-16006-1 [DOI] [Google Scholar]
- 3.Jutterström S, Andersson HC, Omstedt A, Malmaeus JM. Multiple stressors threatening the future of the Baltic Sea–Kattegat marine ecosystem: Implications for policy and management actions. Mar Pollut Bull. 2014;86(1–2):468–80. 10.1016/j.marpolbul.2014.06.027 [DOI] [PubMed] [Google Scholar]
- 4.HELCOM. Eutrophication status of the Baltic Sea Balt. Sea Environ. Proc., 2007–2011. [Google Scholar]
- 5.Neumann T. Climate-change effects on the Baltic Sea ecosystem: A model study. J Marine Syst. 2010;81:213–24. 10.1016/j.jmarsys.2009.12.001 [DOI] [Google Scholar]
- 6.Ærtebjerg G, Andersen JH, Hansen OS. Nutrients and eutrophication in danish marine waters. A challenge for science and management. National Environmental Research Institute. 2003:126 pp. [Google Scholar]
- 7.Meier HEM, Andersson HC, Eilola K, Gustafsson BG, Kuznetsov I, Müller-Karulis B, et al. Hypoxia in future climates: A model ensemble study for the Baltic Sea. Geophys Res Lett. 2011;38(24):L24608 10.1029/2011GL049929 [DOI] [Google Scholar]
- 8.Weigelt M. Short-and long-term changes in the benthic community of the deeper parts of Kiel Bay (Western Baltic) due to oxygen depletion and eutrophication. Meeresforschung 1991;33:197–224. [Google Scholar]
- 9.Schmidt A, Coll M, Romanuk T, Lotze H. Ecosystem structure and services in eelgrass Zostera marina and rockweed Ascophyllum nodosum habitats. Mar Ecol-Prog Ser. 2011;437:51–68. 10.3354/meps09276 [DOI] [Google Scholar]
- 10.Lloret J, Marín A. The role of benthic macrophytes and their associated macroinvertebrate community in coastal lagoon resistance to eutrophication. Mar Pollut Bull. 2009;58(12):1827–34. 10.1016/j.marpolbul.2009.08.001 [DOI] [PubMed] [Google Scholar]
- 11.Lloret J, Marín A. The contribution of benthic macrofauna to the nutrient filter in coastal lagoons. Mar Pollut Bull. 2011;62(12):2732–40. 10.1016/j.marpolbul.2011.09.014 [DOI] [PubMed] [Google Scholar]
- 12.Häder D-P, Schäfer J. In-situ measurement of photosynthetic oxygen production in the water column. Environ Monit Assess. 1994;32(3):259–68. 10.1007/BF00546280 [DOI] [PubMed] [Google Scholar]
- 13.Wahl M, Molis M, Hobday AJ, Dudgeon S, Neumann R, Steinberg P, et al. The responses of brown macroalgae to environmental change from local to global scales: direct versus ecologically mediated effects. Perspec Phycol. 2015. 10.1127/pip/2015/0019 [DOI] [Google Scholar]
- 14.Wernberg T, Russell BD, Thomsen MS, Gurgel CFD, Bradshaw CJA, Poloczanska ES, et al. Seaweed communities in retreat from ocean warming. Curr Biol. 2011;21(21):1828–32. 10.1016/j.cub.2011.09.028 [DOI] [PubMed] [Google Scholar]
- 15.Wahl M, Jormalainen V, Eriksson BK, Coyer JA, Molis M, Schubert H, et al. Stress ecology in Fucus: abiotic, biotic and genetic interactions. Adv Mar Biol. 2011;59:37–105. Epub 2011/07/05. 10.1016/b978-0-12-385536-7.00002-9 . [DOI] [PubMed] [Google Scholar]
- 16.Lotze HK, Worm B, Sommer U. Strong bottom-up and top-down control of early life stages of macroalgae. Limnol Oceanogr. 2001;46(4):749–57. 10.4319/lo.2001.46.4.0749 [DOI] [Google Scholar]
- 17.Wikström SA, Kautsky L. Structure and diversity of invertebrate communities in the presence and absence of canopy-forming Fucus vesiculosus in the Baltic Sea. Estuar Coast Shelf S. 2007;72(1–2):168–76. 10.1016/j.ecss.2006.10.009 [DOI] [Google Scholar]
- 18.Torn K, Krause-Jensen D, Martin G. Present and past depth distribution of bladderwrack (Fucus vesiculosus) in the Baltic Sea. Aquat Bot. 2006;84(1):53–62. 10.1016/j.aquabot.2005.07.011 [DOI] [Google Scholar]
- 19.Vogt H, Schramm W. Conspicuous decline of Fucus in Kiel Bay (Western Baltic): What are the causes? Mar Ecol Prog Ser. 1991;69:189–94. 10.3354/meps069189 [DOI] [Google Scholar]
- 20.Kautsky N, Kautsky H, Kautsky U, Waern M. Decreased depth penetration of Fucus vesiculosus (L.) since the 1940s indicates eutrophication of the Baltic Sea. Mar Ecol Prog Ser. 1986;28:1–8. 10.3354/meps028001 [DOI] [Google Scholar]
- 21.Eriksson BK, Johansson G, Snoeijs P. Long-term changes in the sublittoral zonation of brown algae in the southern Bothnian Sea. Eur J Phycol. 1998;33(3):241–9. 10.1080/09670269810001736743 [DOI] [Google Scholar]
- 22.Rohde S, Hiebenthal C, Wahl M, Karez R, Bischof K. Decreased depth distribution of Fucus vesiculosus (Phaeophyceae) in the Western Baltic: effects of light deficiency and epibionts on growth and photosynthesis. Eur J Phycol. 2008;43(2):143–50. 10.1080/09670260801901018 [DOI] [Google Scholar]
- 23.Worm B, Sommer U. Rapid direct and indirect effects of a single nutrient pulse in a seaweed-epiphyte-grazer system. Mar Ecol Prog Ser. 2000;202:283–8. 10.3354/meps202283 [DOI] [Google Scholar]
- 24.Hemmi A, Jormalainen V. Nutrient enhancement increases performance of a marine herbivore via quality of its food alga. Ecology. 2002;83(4):1052–64. 10.2307/3071913 [DOI] [Google Scholar]
- 25.Eriksson BK, Johansson G. Sedimentation reduces recruitment success of Fucus vesiculosus (Phaeophyceae) in the Baltic Sea. Eur J Phycol. 2003;38(3):217–22. 10.1080/0967026031000121688 [DOI] [Google Scholar]
- 26.Rinne H, Salovius-Laurén S, Mattila J. The occurrence and depth penetration of macroalgae along environmental gradients in the northern Baltic Sea. Estuar Coast Shelf S. 2011;94(2):182–91. 10.1016/j.ecss.2011.06.010 [DOI] [Google Scholar]
- 27.Nygård CA, Dring MJ. Influence of salinity, temperature, dissolved inorganic carbon and nutrient concentration on the photosynthesis and growth of Fucus vesiculosus from the Baltic and Irish Seas. Eur J Phycol. 2008;43(3):253–62. 10.1080/09670260802172627 [DOI] [Google Scholar]
- 28.Burnett LE. The challenges of living in hypoxic and hypercapnic aquatic environments. Am Zool. 1997;37(6):633–40. 10.1093/icb/37.6.633 [DOI] [Google Scholar]
- 29.Gray JS, Wu RSS, Or YY. Effects of hypoxia and organic enrichment on the coastal marine environment. Mar Ecol Prog Ser. 2002;238:249–79. 10.3354/meps238249 [DOI] [Google Scholar]
- 30.Carstensen J, Conley D, Bonsdorff E, Gustafsson B, Hietanen S, Janas U, et al. Hypoxia in the Baltic Sea: Biogeochemical Cycles, Benthic Fauna, and Management. AMBIO. 2014;43(1):26–36. 10.1007/s13280-013-0474-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Brewer PG, Peltzer ET. Limits to marine life. Science. 2009;324(5925):347–8. 10.1126/science.1170756 [DOI] [PubMed] [Google Scholar]
- 32.Elken J, Lehmann A, Myrberg K. Recent change—marine circulation and stratification, p. 131–144. In Second assessment of climate change for the Baltic Sea basin. The BACC II Author Team (editors), Springer. 2015. 10.1007/978-3-319-16006-1_7 [DOI] [Google Scholar]
- 33.Al-Janabi B, Kruse I, Graiff A, Karsten U, Wahl M. Genotypic variation influences tolerance to warming and acidification of early life-stage Fucus vesiculosus L. (Phaeophyceae) in a seasonally fluctuating environment. Mar Biol. 2016;163(1):1–15. 10.1007/s00227-015-2804-8 [DOI] [Google Scholar]
- 34.Graiff A, Liesner D, Karsten U, Bartsch I. Temperature tolerance of western Baltic Sea Fucus vesiculosus–growth, photosynthesis and survival. J Exp Mar Biol Ecol. 2015;471(0):8–16. 10.1016/j.jembe.2015.05.009 [DOI] [Google Scholar]
- 35.Assis J, Serrão EA, Claro B, Perrin C, Pearson GA. Climate-driven range shifts explain the distribution of extant gene pools and predict future loss of unique lineages in a marine brown alga. Mol Ecol. 2014;23(11):2797–810. 10.1111/mec.12772 [DOI] [PubMed] [Google Scholar]
- 36.Nicastro K, Zardi G, Teixeira S, Neiva J, Serrão E, Pearson G. Shift happens: trailing edge contraction associated with recent warming trends threatens a distinct genetic lineage in the marine macroalga Fucus vesiculosus. BMC Biol. 2013;11(1):1–13. 10.1186/1741-7007-11-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Jueterbock A, Tyberghein L, Verbruggen H, Coyer JA, Olsen JL, Hoarau G. Climate change impact on seaweed meadow distribution in the North Atlantic rocky intertidal. Ecol Evol. 2013;3(5):1356–73. 10.1002/ece3.541 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Olischläger M, Bartsch I, Gutow L, Wiencke C. Effects of ocean acidification on growth and physiology of Ulva lactuca (Chlorophyta) in a rockpool-scenario. Phycol Res. 2013;61(3):180–90. 10.1111/pre.12006 [DOI] [Google Scholar]
- 39.Beardall J, Giordano M. Ecological implications of microalgal and cyanobacterial CO2 concentrating mechanisms, and their regulation. Funct Plant Biol. 2002;29(3):335–47. 10.1071/PP01195 [DOI] [PubMed] [Google Scholar]
- 40.Fernandez PA, Roleda MY, Hurd CL. Effects of ocean acidification on the photosynthetic performance, carbonic anhydrase activity and growth of the giant kelp Macrocystis pyrifera. Photosynth Res. 2015;124(3):293–304. Epub 2015/04/15. 10.1007/s11120-015-0138-5 . [DOI] [PubMed] [Google Scholar]
- 41.Coelho S, Rijstenbil J, Brown M. Impacts of anthropogenic stresses on the early development stages of seaweeds. J Aquat Ecosyst Stress Recovery. 2000;7(4):317–33. 10.1023/A:1009916129009 [DOI] [Google Scholar]
- 42.Maczassek K. Environmental drivers of fertility, fertilization and germination of Fucus vesiculosus on the German coast [Ph.D. Thesis]. Kiel: Christian-Albrechts-University; 2014.
- 43.Nielsen S, Nielsen H, Pedersen M. Juvenile life stages of the brown alga Fucus serratus L. are more sensitive to combined stress from high copper concentration and temperature than adults. Mar Biol. 2014;161(8):1895–904. 10.1007/s00227-014-2471-1 [DOI] [Google Scholar]
- 44.Bergström L, Berger R, Kautsky L. Negative direct effects of nutrient enrichment on the establishment of Fucus vesiculosus in the Baltic Sea. Eur J Phycol. 2003;38(1):41–6. 10.1080/0967026031000096236 [DOI] [Google Scholar]
- 45.Berger R, Bergström L, Granéli E, Kautsky L. How does eutrophication affect different life stages of Fucus vesiculosus in the Baltic Sea?–a conceptual model. Hydrobiologia. 2004;514(1–3):243–8. 10.1023/B:hydr.0000018222.44511.b7 [DOI] [Google Scholar]
- 46.Pineda MC, McQuaid CD, Turon X, López-Legentil S, Ordóñez V, Rius M. Tough adults, frail babies: an analysis of stress sensitivity across early life-history stages of widely introduced marine invertebrates. PLoS One. 2012;7(10):e46672 10.1371/journal.pone.0046672 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Wahl M, Saderne V, Sawall Y. How good are we at assessing the impact of ocean acidification in coastal systems? Limitations, omissions and strengths of commonly used experimental approaches with special emphasis on the neglected role of fluctuations. Mar Freshwater Res. 2015. 10.1071/MF14154 [DOI] [Google Scholar]
- 48.Pauls SU, Nowak C, Bálint M, Pfenninger M. The impact of global climate change on genetic diversity within populations and species. Mol Ecol. 2013;22(4):925–46. 10.1111/mec.12152 [DOI] [PubMed] [Google Scholar]
- 49.Frankham R, Ballou JD, Bricoe DA. Introduction to conservation genetics. 2nd edition Cambridge University Press, Cambridge, UK: 2010. 10.1017/CBO9780511809002 [DOI] [Google Scholar]
- 50.Frankham R. Genetics and conservation biology. Comptes Rendus Biologies. 2003;326, Supplement 1(0):22–9. 10.1016/S1631-0691(03)00023-4 [DOI] [PubMed] [Google Scholar]
- 51.Hoffmann AA, Sgro CM. Climate change and evolutionary adaptation. Nature. 2011;470(7335):479–85. 10.1038/nature09670 [DOI] [PubMed] [Google Scholar]
- 52.Sanford E, Kelly MW. Local adaptation in marine invertebrates. Ann Rev Mar Sci. 2011;3(1):509–35. 10.1146/annurev-marine-120709-142756 . [DOI] [PubMed] [Google Scholar]
- 53.Kawecki TJ, Ebert D. Conceptual issues in local adaptation. Ecol Lett. 2004;7(12):1225–41. 10.1111/j.1461-0248.2004.00684.x [DOI] [Google Scholar]
- 54.Reusch TBH, Ehlers A, Hämmerli A, Worm B. Ecosystem recovery after climatic extremes enhanced by genotypic diversity. Proc Natl Acad Sci USA. 2005;102(8):2826–31. 10.1073/pnas.0500008102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ehlers A, Worm B, Reusch TBH. Importance of genetic diversity in eelgrass Zostera marina for its resilience to global warming. Mar Ecol Prog Ser. 2008;355:1–7. 10.3354/meps07369 [DOI] [Google Scholar]
- 56.Johannesson K, André C. Life on the margin: genetic isolation and diversity loss in a peripheral marine ecosystem, the Baltic Sea. Mol Ecol. 2006;15(8):2013–29. Epub 2006/06/20. 10.1111/j.1365-294X.2006.02919.x . [DOI] [PubMed] [Google Scholar]
- 57.Serrão EA. Distributional success of the marine seaweed Fucus vesiculosus L. in the brackish Baltic Sea correlates with osmotic capabilities of Baltic gametes. Oecologia. 1996. 10.1007/bf00582229 [DOI] [PubMed] [Google Scholar]
- 58.Johannesson K, Smolarz K, Grahn M, André C. The future of Baltic sea populations: local extinction or evolutionary rescue? AMBIO. 2011;40(2):179–90. 10.1007/s13280-010-0129-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wahl M, Buchholz B, Winde V, Golomb D, Guy-Haim T, Müller J, et al. A mesocosm concept for the simulation of near-natural shallow underwater climates: The Kiel Outdoor Benthocosms (KOB). Limnol Oceanogr Methods. 2015. [Google Scholar]
- 60.Winde V, Böttcher ME, Escher P, Böning P, Beck M, Liebezeit G, et al. Tidal and spatial variations of DI13C and aquatic chemistry in a temperate tidal basin during winter time. Journal of Marine Systems. 2014;129:396–404. 10.1016/j.jmarsys.2013.08.005 [DOI] [Google Scholar]
- 61.Steen H, Scrosati R. Intraspecific competition in Fucus serratus and F. evanescens (Phaeophyceae: Fucales) germlings: effects of settlement density, nutrient concentration, and temperature. Mar Biol. 2004;144(1):61–70. 10.1007/s00227-003-1175-8 [DOI] [Google Scholar]
- 62.Verardo DJ, Froelich PN, McIntyre A. Determination of organic carbon and nitrogen in marine sediments using the Carlo Erba NA-1500 analyzer. Deep-Sea Res PT A. 1990;37(1):157–65. 10.1016/0198-0149(90)90034-S [DOI] [Google Scholar]
- 63.Satterthwaite FE. An approximate distribution of estimates of variance components. Biometrics. 1946;2(6):110–4. Epub 1946/12/01. 10.2307/3002019 [DOI] [PubMed] [Google Scholar]
- 64.Inc. D. Dell Statistica (data analysis software system), version 12. software.dell.com. 2015.
- 65.Wallentinus I. Comparisons of nutrient uptake rates for Baltic macroalgae with different thallus morphologies. Mar Biol. 1984;80(2):215–25. 10.1007/BF02180189 [DOI] [Google Scholar]
- 66.Gordillo FJL, Niell FX, Figueroa FL. Non-photosynthetic enhancement of growth by high CO2 level in the nitrophilic seaweed Ulva rigida C. Agardh (Chlorophyta). Planta. 2001;213(1):64–70. 10.1007/s004250000468 [DOI] [PubMed] [Google Scholar]
- 67.Zou D. Effects of elevated atmospheric CO2 on growth, photosynthesis and nitrogen metabolism in the economic brown seaweed, Hizikia fusiforme (Sargassaceae, Phaeophyta). Aquaculture. 2005;250(3–4):726–35. 10.1016/j.aquaculture.2005.05.014 [DOI] [Google Scholar]
- 68.Zou D, Gao K, Luo H. Short- and long- term effects of elevated CO2 on photosynthesis and respiration in the marine macroalga Hizikia fusiformis (Sargassaceae, Phaeophyta) grown at low and high N supplies. J Phycol. 2011;47(1):87–97. 10.1111/j.1529-8817.2010.00929.x [DOI] [PubMed] [Google Scholar]
- 69.Berry J, Bjorkman O. Photosynthetic response and adaptation to temperature in higher plants. Annu Rev Plant Phys. 1980;31(1):491–543. 10.1146/annurev.pp.31.060180.002423 [DOI] [Google Scholar]
- 70.Yordanov I., Dilova S., Petkova R., Pangelova T., Goltsev V., Suss K. H. Mechanisms of the temperature damage and acclimation of the photosynthetic apparatus. Photobioch Photobiop. 1986;12:147–55. [Google Scholar]
- 71.Allakhverdiev S, Kreslavski V, Klimov V, Los D, Carpentier R, Mohanty P. Heat stress: an overview of molecular responses in photosynthesis. Photosynth Res. 2008;98(1–3):541–50. 10.1007/s11120-008-9331-0 [DOI] [PubMed] [Google Scholar]
- 72.Andersen GS, Pedersen MF, Nielsen SL. Temperature acclimation and heat tolerance of photosynthesis in Norwegian Saccharina latissima (Laminariales, Phaeophyceae). J Phycol. 2013;49(4):689–700. 10.1111/jpy.12077 [DOI] [PubMed] [Google Scholar]
- 73.Lehvo A, Bäck S, Kiirikki M. Growth of Fucus vesiculosus L. (Phaeophyta) in the northern Baltic proper: energy and nitrogen storage in seasonal environment. Bot Mar. 2001;p. 345 10.1515/BOT.2001.044 [DOI] [Google Scholar]
- 74.Thomsen J, Casties I, Pansch C, Koertzinger A, Melzner F. Food availability outweighs ocean acidification effects in juvenile Mytilus edulis: laboratory and field experiments. Glob Change Biol. 2013;19(4):1017–27. 10.1111/gcb.12109 WOS:000315900800005. [DOI] [PubMed] [Google Scholar]
- 75.Reusch TH, Hughes AR. The emerging role of genetic diversity for ecosystem functioning: Estuarine macrophytes as models. Estuar Coasts. 2006;29(1):159–64. 10.1007/BF02784707 [DOI] [Google Scholar]
- 76.Reusch TBH, Wood TE. Molecular ecology of global change. Mol Ecol. 2007;16(19):3973–92. 10.1111/j.1365-294X.2007.03454.x [DOI] [PubMed] [Google Scholar]
- 77.Melzner F, Thomsen J, Koeve W, Oschlies A, Gutowska M, Bange H, et al. Future ocean acidification will be amplified by hypoxia in coastal habitats. Mar Biol. 2013;160(8):1875–88. 10.1007/s00227-012-1954-1 [DOI] [Google Scholar]
- 78.Saderne V, Fietzek P, Herman PMJ. Extreme variations of pCO2 and pH in a macrophyte meadow of the Baltic sea in summer: evidence of the effect of photosynthesis and local upwelling. PLoS One. 2013;8(4):e62689 10.1371/journal.pone.0062689 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Peckol P, Rivers JS. Physiological responses of the opportunistic macroalgae Cladophora vagabunda (L.) van den Hoek and Gracilaria tikvahiae (McLachlan) to environmental disturbances associated with eutrophication. J Exp Mar Biol Ecol. 1995;190(1):1–16. 10.1016/0022-0981(95)00026-N [DOI] [Google Scholar]
- 80.Plaxton WC, Podestá FE. The functional organization and control of plant respiration. CR Rev Plant Sci. 2006;25(2):159–98. 10.1080/07352680600563876 [DOI] [Google Scholar]
- 81.Diaz RJ, Rosenberg R. Marine benthic hypoxia: a review of its ecological effects and the behavioural responses of benthic macrofauna. Oceanograph Lit Rev. 1996;43(12):1250. [Google Scholar]
- 82.Queirós AM, Fernandes JA, Faulwetter S, Nunes J, Rastrick SPS, Mieszkowska N, et al. Scaling up experimental ocean acidification and warming research: from individuals to the ecosystem. Glob Change Biol. 2015;21(1):130–43. 10.1111/gcb.12675 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Nutrient concentrations within the experimental germling boxes before (A, B) and after (C, D) the bi-weekly addition of the nutrients (PO4, NO3, NO2) as well as initial NH4 conditions in μmol L-1 in July and August. Initial nutrient concentrations were measured six times per month in the main KOB tank before the water addition to the boxes. Nutrient concentrations after additions were determined by adding the sum of the initial and additional nutrient concentration.
(TIFF)
Data Availability Statement
All data are uploaded in the repository PANGAEA (Data Publisher for Earth & Environmental Science). doi: http://doi.pangaea.de/10.1594/PANGAEA.853938.