

Abstract
Evolutionary biologists have long considered menopause to be a fundamental puzzle in understanding human fertility behaviour, as post-menopausal women are no longer physiologically capable of direct reproduction. Menopause typically occurs between 45 and 55 years of age, but across cultures and history, women often stop reproducing many years before menopause. Unlike age at first reproduction or even birth spacing, a woman nearing the end of her reproductive cycle is able to reflect upon the offspring she already has—their numbers and phenotypic qualities, including sexes. This paper reviews demographic data on age at last birth both across and within societies, and also presents a case study of age at last birth in rural Bangladeshi women. In this Bangladeshi sample, age at last birth preceded age at menopause by an average of 11 years, with marked variation around that mean, even during a period of high fertility. Moreover, age at last birth was not strongly related to age at menopause. Our literature review and case study provide evidence that stopping behaviour needs to be more closely examined as an important part of human reproductive strategies and life-history theory. Menopause may be a final marker of permanent reproductive cessation, but it is only one piece of the evolutionary puzzle.
Keywords: age at last birth, stopping reproduction, reproductive cessation, infertility, menopause, life-history theory
1. Introduction
Both demographers and evolutionary anthropologists have a long-standing interest in human fertility and reproduction, and recognize that even within high-fertility populations, women reproduce below their apparent physiological maximum [1–4]. To explain this observation, demographers have focused primarily on proximate correlates of population-level fertility (e.g. mortality rates, female education and access to modern contraception) [5,6]. By contrast, evolutionary anthropologists have focused on the individual level and ultimate explanations to consider how fertility relates to lifetime reproductive success (e.g. through optimal birth spacing and trade-offs between quantity and quality of offspring [7]). Recent work in this area shows just how important the timing of births is to evolutionary fitness [8].
One of the many puzzles about fertility is why long-lived women stop reproducing, often decades before their deaths. A woman's reproductive lifespan can be visualized as a timeline of key events: menarche, births and interbirth intervals, and menopause (figure 1). Here, we follow the demographic literature in using fecundity to refer to the ability to reproduce, and fertility to refer to actual births. The first and last births define a woman's realized reproductive period. After a woman's last birth, she is infertile in the sense that she has stopped reproducing, but she may still be fecund (a). She may then experience a period of subfecundity (b) during perimenopause and eventual sterility (c), but only with menopause—defined as 1 year without menstrual cycles—can we infer permanent infecundity or sterility. (For more detailed definitions, see Wood [2].)
Figure 1.

Timeline of a hypothetical woman's reproductive lifespan between menarche and menopause, with key events indicated by triangles (solid for visible events, shaded for transitions or hidden events). Fertility is shown as N births (B1 through BN) that occur during a primary window of fecundity (unshaded). Although not shown here, some births could occur during periods of subfecundity (shaded). (For simplicity, periods of temporary infecundity and subfecundity following births are not shown.) Reproductive stopping occurs with the last birth, BN, at ALB (age at last birth). Following BN, the time durations between events are denoted as shown by a, b, c and d, where d is the time from menopause to death. The time from last birth to menopause is thus a + b + c, and the post-reproductive lifespan is a + b + c + d. In theory, only the time between BN and death constrains the values of a, b, c and d, although c ≥ 1 year by definition of menopause.
Although age at last birth and stopping reproduction have been examined by evolutionary anthropologists and demographers, this is often done from very different perspectives. In evolutionary anthropology, reproductive cessation has garnered the most attention in the context of hypotheses for menopause and a long post-reproductive lifespan [9–12]. Studies of the costs of reproduction have also looked at women's age at last birth in relationship to longevity (e.g. [13,14]). For demographers, age at last birth is important in estimating age-specific fertility rates, as well as in identifying natural fertility populations (defined as those where parity-dependent control of reproduction is absent) [15,16]. Declining ages of last birth are also of particular interest to demographers, as a potential indication of the onset of demographic transitions from high to low fertility [1].
Despite this work, we believe that reproductive stopping behaviour, in and of itself, has received relatively little attention, and that some common assumptions about age at last birth merit closer study. One such assumption often made in the evolutionary literature is that reproductive cessation is largely a reflection of the physiological changes preceding menopause [2,9,12,13]. In terms of figure 1, this would imply that a is short relative to b + c. At a proximate level, this seems quite reasonable, as menopause marks the end of a physiological process after which women are no longer capable of direct reproduction. Moreover, there is strong evidence of fertility decline preceding menopause and also increases in pregnancy loss [17]. That some women approaching menopause experience such factors, however, is not in itself evidence that most women stop reproducing because of such factors. Although this causal link may be true for some women, particularly in natural fertility populations, we will highlight evidence that many women stop when they are still fecund (i.e. that a is not short relative to b + c).
Another common assumption about age at last birth is that there is a species-typical 10-year gap between age at last birth and menopause, with age at last birth occurring around 40 and menopause around 50 years of age. Looking at figure 1, the conventional wisdom is that a + b + c is typically 10 years, and, moreover, that this duration is fairly invariant [2]. Together with the physiological assumption, these lead to the prediction that age at last birth and age at menopause will be strongly correlated. And while there are broad population-level data to support these central age tendencies, a focus on means, round numbers and natural fertility populations leads us to ignore quite interesting variation in age at last birth (i.e. a + b + c is quite variable across women).
Our aim with this review is to examine a woman's last birth as a significant event in her reproductive life history, distinct from menopause. We will show how demographic and evolutionary perspectives can be combined to form an improved understanding of stopping reproduction and age at last birth. We describe variation across societies in these traits and discuss the many proximate mechanisms through which people may regulate their fertility. We also analyse one population, a sample of rural Bangladeshi women, in greater detail, and place the results in the context of evolutionary explanations for stopping reproduction and age at last birth.
2. Variation in age at last birth across and within populations
We begin with an examination of variation in age at last birth across populations (table 1). Through a Web of Science literature search, we located studies with a primary focus on age at last birth and drew data from those without obvious sources of bias (e.g. focus on a particular subset of women). We also drew from several long-term anthropological studies and from published tables found in Bongaarts & Potter [5] and Wood [2]. Our table expands on theirs not only by adding more recent studies, but also by including measures of range and variance as available. We also include some populations that do not fit the natural fertility definition, whether historical (e.g. Finland [27]) or late twentieth century with access to modern contraception (e.g. Poland [36]). Although not fully comprehensive, table 1 represents a diverse sample of human populations with published data on age at last birth.
Table 1.
Variation in age at last birth (ALB) across populations.
| continent | century | population | mean ALB | range | s.d. | N | source |
|---|---|---|---|---|---|---|---|
| Africa | 19th–early 20th | Botswana (Herero) | 29.7 | 239 | Pennington & Harpending [18] | ||
| mid-20th | Kenya (Kipsigis) | 39.7 | 25.6–48.2 | 33 | Borgerhoff Mulder [19] | ||
| late 20th | Morocco (Amizmiz) | 37 | 7.3 | 842 | Varea [20] | ||
| mid-to-late 20th | Egypt | 35.5a | 870 | Horne & Suchindran [21] | |||
| mid-to-late 20th | Gambia | 39 | 26–51 | 5.2 | 110 | R. Sear et al. ([22] and personal communication) | |
| mid-20th | Kalahari Desert (!Kung) | 34.4 | 19–47 | 8.2 | Howell [23] | ||
| Asia | mid-to-late 20th | Bangladesh | 36.2 | 15–54 | 1037 | Towner et al. (this paper) | |
| mid-20th | Sri Lanka | 35.7 | 6810 | Gajanayake [24] | |||
| late 20th | Indonesia | 31.9 | 16–51 | 6.2 | 1801 | K. Snopkowski et al. ([25] and personal communication) | |
| Europe | 16th–19th | Finland | 37.7 | 16.0–52.3 | 6.3 | 5336 | Lahdenpera et al. [26] |
| 17th–18th | France (Tourouvre au Perche) | 40.3 | after Bongaarts & Potter [5] | ||||
| 17th–18th | France (Crulai) | 40.0 | after Bongaarts & Potter [5] | ||||
| 17th–19th | Finland (Utsjoki) | 41.6 | 4.6 | 61 | Helle et al. [13] | ||
| 17th–19th | Finland (Inari) | 41.7 | 3.6 | 144 | Helle et al. [13] | ||
| 17th–19th | Finland (Enontekio) | 42.1 | 3.8 | 101 | Helle et al. [13] | ||
| 18th | Finland | 30.4 | 6.6 | 75 | Korpelainen [27] | ||
| 19th | Finland | 31.7 | 5.6 | 91 | Korpelainen [27] | ||
| 18th–19th | Germany (four Waldeck Villages) | 40.3 | 1589 | Knodel [28] | |||
| 18th–19th | Germany (Grafenhausen) | 38.2 | 1149 | Knodel [28] | |||
| 18th–19th | Sweden (Scania) | 39.4 | 4.5 | 11 031 | Dribe [29] | ||
| 18th–19th | France | 39.5 | 4.4 | 994 | Desjardins et al. [30] | ||
| early 19th | The Netherlands (Utrecht) | 40.1 | 4.2 | 481 | Van Bavel & Kok [31] | ||
| late 19th | The Netherlands (Utrecht) | 36.9 | 5.5 | Van Bavel & Kok [31] | |||
| 19th | Belgium (Sart) | 40.3 | 386 | Alter et al. [32] | |||
| early 20th | Mykonos | 39 | Hionidou [33] | ||||
| early-to-mid-20th | Portugal | 32.3 | 6.6 | 320 | Gama [34] | ||
| mid-to-late 20th | Norway | 29.5 | 5.1 | 744 784 | Grundy & Kravdal [35] | ||
| mid-to-late 20th | Poland | 31.3 | 1738 | Colleran & Mace [36] | |||
| North America (European) | 17th | Quebec | 41.5 | 3.3 | 2226 | Desjardins et al. [30] | |
| 17th | Canada | 40.1 | after Charbonneau [37] | ||||
| 17th–18th | Massachusetts (Hingham) | 39.1 | after Charbonneau [37] | ||||
| 18th | Canada | 41 | after Charbonneau [37] | ||||
| 18th | New England (Quakers) | 39.6 | after Charbonneau [37] | ||||
| 18th–20th | USA (Amish) | 38.6 | 5.2 | 937 | McArdle et al. [38] | ||
| 19th | Utah (Mormons) | 40 | 18.7–54.9 | 4.6 | 13 897 | Smith et al. [39] | |
| early-to-mid-20th | Bois-Vert | 40.7 | after Charbonneau [37] | ||||
| early-to-mid-20th | Cris | 38.7 | after Charbonneau [37] | ||||
| mid-to-late 20th | St Jean-de-Cherbourg | 38.6 | after Charbonneau [37] | ||||
| 20th | USA (Hutterite) | 35.1 | 22.5–47.1 | 5.3 | 353 | Kosova et al. [40] | |
| North America (Indigenous) | early 20th | USA (Red Lake) | 37.8 | Shoemaker [41] | |||
| early 20th | USA (Cherokee) | 37.1 | Shoemaker [41] | ||||
| early 20th | USA (Navajo) | 34.6 | Shoemaker [41] | ||||
| early 20th | USA (Seneca) | 33.2 | Shoemaker [41] | ||||
| early 20th | USA (Yakama) | 32.6 | Shoemaker [41] | ||||
| mid-20th | Mexico (Xculoc Mayan) | 38.8a | ≈40 | Kramer [42] | |||
| South America | early-to-mid-20th | Paraguay (Ache: forest period) | 42.1 | 34–48 | 20 | Hill & Hurtado [43] | |
| mid-20th | Yanomamo | 37.9 | after Hill & Hurtado [43] | ||||
| late 20th | Paraguay (Ache: reservation period) | 38.5 | 6.5 | Hill & Hurtado [43] | |||
| mid-to-late 20th | Tierra del Fuego (Porvenir) | 32.1 | 6.3 | 155 | Pascual et al. [44] |
aMedian.
Taken as a whole, table 1 reveals a central tendency, but also shows considerable variation in age at last birth—in other words, the potential for women to stop reproduction at many different ages, both long before menopause and very close to menopause. Even including several controlled fertility populations, the median value of the ALB reported in table 1 is 38.6, which is consistent with the 39–41 year range noted by demographers [2,5]. Where reported, however, the age ranges and standard deviations always indicate considerable variation (e.g. in nineteenth century Utah, mean = 40, range 18.7–54.9 years of age, s.d. 4.6 [45]). Assuming a normal distribution with a mean of 40 and s.d. of 4.5, approximately 25% of women would be expected to have their last birth by 37 years of age and about 10% by age 34. This speaks nothing to cause, but it does call into question the notion that an age at last birth of around 40 is an invariant aspect of human female reproductive biology, even among natural fertility populations.
Furthermore, a number of populations reported in table 1 show a mean age at last birth closer to 35 years of age, with higher standard deviations as well. This reflects our method of study selection, which includes populations that would not meet the ‘natural fertility’ definition. From an evolutionary perspective, the realized fertility of women across a spectrum of population types is arguably more relevant to understanding human reproductive strategies than is the focus on a subset of populations practising limited fertility control. However, even with this broader focus, the variation seen across populations reveals little about the causal mechanisms or even life-history correlates of variation in age at last birth and reproductive stopping behaviour. We next consider some such correlates for the Bangladeshi sample reported in table 1.
3. Variation in stopping reproduction and age at last birth in rural Bangladesh
We know much less about variation in age at last birth within populations than across populations, particularly since so many studies report only mean values (table 1). Here, we use a sample of rural Bangladeshi women to examine age at last birth, including its relationship to other life-history events such as menopause. Our goal is not to conduct an exhaustive analysis of stopping behaviour in Bangladesh, but rather to use these data as a deeper case study to inform our review. After briefly describing the study population and sample, we present summary statistics for age of last birth as it relates to menopause, followed by a regression model of age at last birth.
Our data come from the 1996 Matlab Health and Socio-economic Survey [46] (MHSS), which includes individual-level data from 4364 households in Matlab, Bangladesh. The Matlab population is rural, relying primarily on agriculture for subsistence and income. Bangladesh has gone through a well-documented demographic transition in the last 50 years. We analyse data from women who at the time of the survey were 50–69 years old (born 1927–1946). In our sample, the median year of last birth is 1975, at which time the total fertility rate (TFR) in Bangladesh was 6.34 [5]. (At the population level, the TFR is the expected number of births to a hypothetical woman following age-specific fertility rates over her childbearing years [2].) Thus, even though Bangladesh was well into its demographic transition by 1996, the majority of women in our analysis reproduced during a period characterized by relatively high fertility, high mortality and limited access to modern contraception. Given their age, most women in our sample also would have had little formal education [47].
Because we are interested in age at last birth among fertile women, we limit the sample to ever-married women with at least one live birth, with recorded dates for menarche and menopause in the MHSS. We exclude some women with missing or problematic dates for key events (e.g. age at last birth after age at menopause), as well as women with menopause before 40, as this is considered to be premature menopause and may indicate other underlying health problems [48]. We include widows, but exclude five separated and divorced women due to their rarity and social marginalization in Matlab [49]. Our final sample includes 1037 women aged 50–69.
Figure 2 boxplots summarize women's ages at key life-history events—menarche, first birth, last birth, and menopause. Figure 3 provides additional comparisons of ages at last birth and menopause. All women experienced their last birth by age 54 and menopause by age 59, with age at menopause showing a steeper decline than age at last birth (figure 3a). A number of women reported the same age at last birth and menopause (seen as zeros in figure 3b). This could be due to imprecise dating or, in some cases, a surgical menopause at delivery. Many could also be women who never resumed their menstrual cycle after their last birth—that is, post-partum amenorrhea may have transitioned immediately into secondary sterility (as noted among some Dobe !Kung women [23]). Even in this moderately high-fertility sample, the median age at last birth (37 years) is 11 years prior to the median age at menopause (48 years). In other words, more than half of all women last gave birth at least a decade before menopause (figure 3b).
Figure 2.
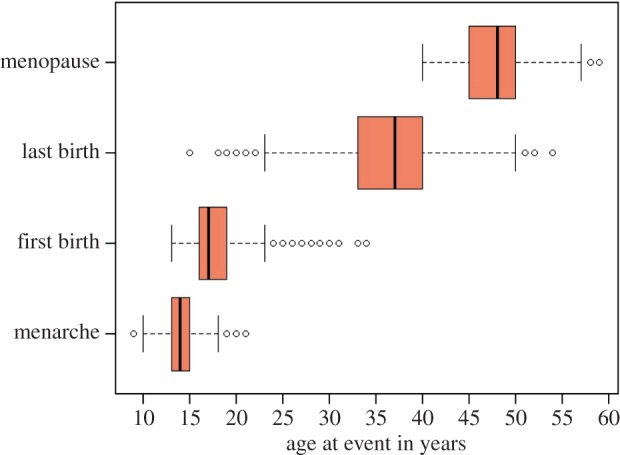
Critical life-history events for Bangladeshi women (n = 1037 aged 50–69 years old in the MHSS). Boxplots give the median event age (line within rectangle), first and third quartile age values (rectangle boundaries), and minimum and maximum age values (brackets), excluding outliers. Summary statistics: age at menarche (mean 14.0, median 14, range 9–21, s.d. 1.4), first birth (mean 17.9, median 17, range 13–34, s.d. 3.0), last birth (mean 36.2, median 37, range 15–54, s.d. 5.5) and menopause (mean 47.4, median 48, range 40–59, s.d. 3.5). (Online version in colour.)
Figure 3.
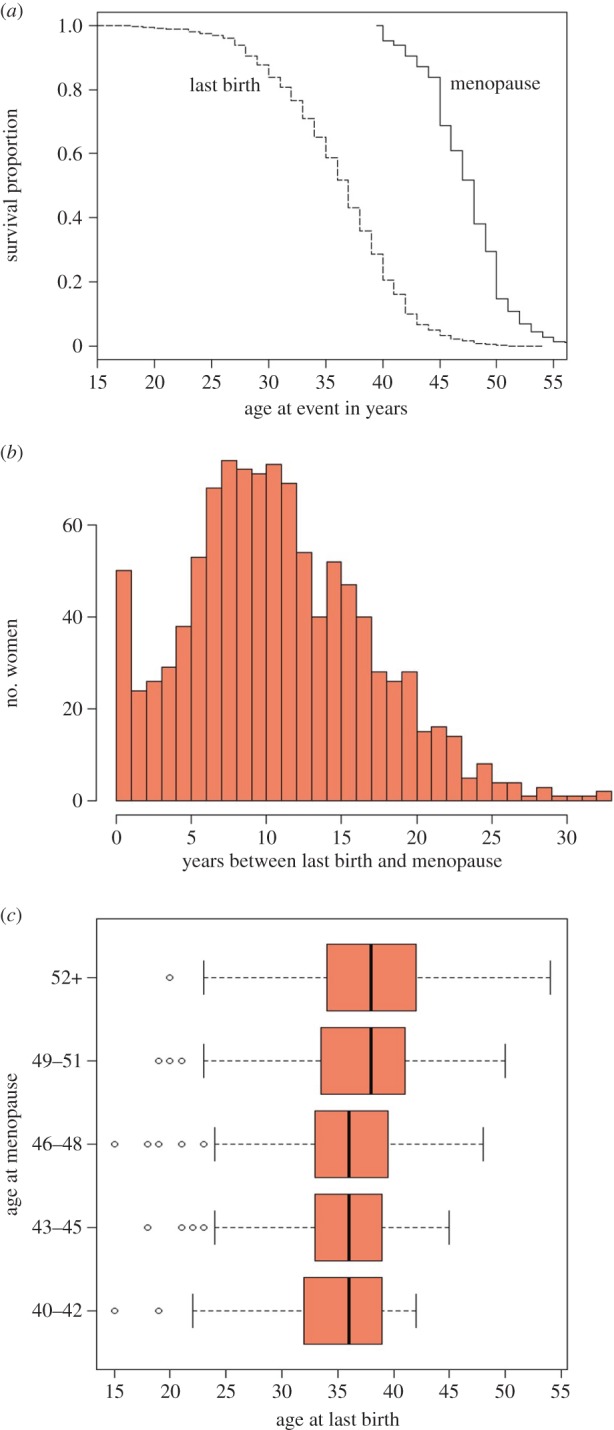
Age at last birth compared to age at menopause for Bangladeshi women (same sample as in figure 2): (a) Kaplan–Meier survival curves for age at last birth (dashed line) and age at menopause (solid line), (b) histogram of number of years between last birth and menopause (mean: 11.2, median: 11, range: 0–33, s.d.: 6.05) and (c) boxplots of age at last birth on the x-axis according to age at menopause (divided into five 3-year age groupings) on the y-axis. (See figure 2 legend for explanation of boxplot features.) (Online version in colour.)
Despite a modest positive correlation (r = 0.17), age at last birth does not appear to be strongly related to age at menopause. Figure 3c shows that neither the median nor the range in age at last birth shifts much with menopausal age, although a younger age at menopause does place an upper limit on age at last birth. This suggests that declining fecundity—e.g. through ovarian depletion—is unlikely to explain the extent of variation seen in age at last birth or the variation in elapsed time between last birth and menopause (figure 3b). To explore this further, we used ordinary least-squares regression to model age at last birth according to age at menopause, as well as other life-history variables: year of birth, age at menarche, number of live births, number of stillbirths and marital status (table 2).
Table 2.
Regression model of life-history traits on age at last birth. Adjusted-R2 0.49, d.f. 1029, p < 0.001.
| covariate | estimate | s.e. | t-value | p-value |
|---|---|---|---|---|
| intercept | 116.74 | 48.35 | 2.4 | 0.02 |
| year of birth | −0.06 | 0.02 | −2.3 | 0.02 |
| age at menarche | −0.05 | 0.09 | −0.6 | 0.58 |
| age at first birth | 0.71 | 0.04 | 16.4 | <0.01 |
| live births | 1.49 | 0.06 | 26.6 | <0.01 |
| stillbirths | 0.91 | 0.14 | 6.6 | <0.01 |
| age at menopause | 0.15 | 0.04 | 4.3 | <0.01 |
| widowed | −1.63 | 0.27 | −6.0 | <0.01 |
The results indicate that later ages at first birth are associated with later ages at last birth. Age at menarche does not appear to have a direct effect on age at last birth, particularly with age at first birth in the model. As seen above with the bivariate correlation, later ages at menopause are modestly correlated with later ages at last birth, but a comparison of coefficient estimates indicates that the relationship between age at first birth and age at last birth is approximately 4.5 times stronger. Widowhood is also associated with earlier ages at last birth, and age at last birth seems to be declining with time (women born earlier in the sample have an older age at last birth). At a proximate level, this could have been facilitated by an ambitious family planning programme that was implemented in 1977 in some intervention study areas in Matlab [50], which some women (about 17%) in our sample may have been a part of prior to their last birth.
Both number of live births and number of stillbirths are positively correlated with ages at last birth, with number of live births being the strongest predictor in the model. These correlations do not reveal a causal direction or imply intentional stopping behaviour, as women with later ages of last birth may simply be able to ‘fit in’ more births. Looking at simple bivariate correlation coefficients, age at last birth is highly correlated with number of live births (r = 0.56); the correlation between age at menopause and number of live births is also positive, but small by comparison (r = 0.06). This suggests that regardless of the causal direction, age at last birth conveys more information about fertility than does age at menopause.
In sum, we find both substantial variation in age at last birth and often more than a decade passing between last birth and menopause. Although some women's reproductive careers may have been cut short by premenopausal subfecundity and menopause, many more appear to have had their last births at an age when other proximate mechanisms for reproductive cessation seem more probable.
4. Proximate determinants of age at last birth
Our aim in this section is to review the wide variety of underlying proximate mechanisms that shape the timing of a woman's last birth, ranging from those over which a woman has no control to those over which she has some control, regardless of whether the latter are employed in a conscious effort to permanently stop reproducing. For the most part, these proximate mechanisms shaping age at last birth are the same as those that produce variation in fertility in general [5], not just at the end of reproduction. However, some proximate mechanisms are more salient than others when considering age at last birth. As a starting point, there is solid evidence that in natural fertility populations, marital fertility declines with age. A simple population estimate of this decline is that the fertility levels of 35–39 year olds are 70% those of 20–24 year olds, with 40–44 year olds dropping to 36% and 45–49 year olds to only 5% of 20–24 year olds [5,16].
Even assuming identical fertility rates across women, by chance women will experience different waiting times to conception and different histories of pregnancy loss and infant mortality. If there is some age after which women are unable to conceive, women will approach this endpoint at different stages in their reproductive schedules (e.g. pregnant, breastfeeding, fertile). Thus, we expect some variation in age at last birth to arise simply due to random processes [51]. Variation in age at last birth could also arise through heterogeneity across women in the timing of infecundity [52]. Teasing apart these processes has proved a challenge to demographers in distinguishing deliberate stopping behaviour from chance [51,53,54]. The physiological contributions to declining fecundity are difficult enough to study in living women, let alone historical records.
The leading candidate for physiological causes of infecundity is ovarian depletion, and due either to differences in the quantity of primary follicles or their rates of atresia, ovarian depletion may occur at different ages across women [4,12]. As the ovaries become more depleted, endocrine changes may also lead ovulatory cycles to become irregular, decreasing the likelihood of successful conceptions [55]. From this perspective, declining fecundity would seem to be a simple extension of menopause itself. This process, however, may be quite heterogeneous, with more fecund women reproducing to later ages [52,56], despite the fact that rates of pregnancy loss increase with age [48]. For example, among Utah women who continued to reproduce into their 40s, there was little evidence of declining fecundity with increased parity once age was controlled for [45].
Other physiological and anatomical causes of female infecundity may also become more pronounced with maternal age, including problems with non-reproductive endocrine functioning (e.g. thyroid hormone, androgens, and cortisol) [4]. Anatomical causes might include problems such as endometriosis, uterine fibroids, and pelvic inflammatory disease (which often results from sexually transmitted infections, but may also result from puerperal infections after giving birth) [2]. Cervical insufficiency may make it difficult to carry pregnancies to term, and uterine prolapse, in which the uterus protrudes into the vaginal canal, becomes more likely with higher parity [57]. A woman's nutritional status may also impact on her fertility. Women who are nutritionally compromised may have irregular ovarian cycles and longer waiting times to conception, higher rates of pregnancy loss and longer durations of post-partum amenorrhea [4].
The above-mentioned causes of subfecundity and sterility may be largely out of a woman's control, but many proximate determinants of age at last birth are shaped by the behaviour of both women and men. The deliberate use of contraception, for example, will contribute to variation in age at last birth, whether or not the explicit goal is stopping reproduction permanently. Women with access to modern methods such as hormonal contraception or intrauterine devices may have quite accurate control in this respect, but there is abundant historical and ethnographic evidence for women using a wide variety of birth control methods in the past, including coitus interruptus, douching, barrier devices for both men and women, and herbal remedies [58]. Worldwide, female sterilization is used by approximately 19% of married (or in a union) women aged between 15 and 49 [59]. Where it is now available, surgical sterilization may thus quite directly limit age at last birth since reversibility is costly and not highly effective or available. The rapidity and uptake of this method suggests that it is meeting a pre-existing need among women—to avoid unwanted pregnancies later in life. That so many women of reproductive age would choose surgical sterilization as a method is also evidence of an underlying preference to permanently stop once a certain age or desired family size is reached.
Declining coital frequency and abstinence could fall under contraceptive mechanisms. Here, though, we consider them in the context of marital (or other) sexual relationships that would otherwise put a woman at risk of conception. Coital frequency declines with duration of marriage, which, holding all else equal, should decrease conception rates [2,60]. More explicit taboos against sexual relations at certain times are also well documented across cultures. Most frequent are those which prohibit sex during menstruation [61] and also sex for a period of time following childbirth or when a woman is breastfeeding [62]. Taboos against sex after a certain age or event (e.g. marriage of the oldest son, birth of the first grandchild) are less well known, but have been documented, for example, among the Yoruba of Nigeria [63]. Outright taboos may be rare, but there is abundant evidence for shame and embarrassment associated with later-aged pregnancies (e.g. in Greece, Iran, India, and France) [64]. In other societies, there seems to be no such stigma (e.g. the Ache, Tsimane and Shuar of South America), and mothers and their own daughters are often pregnant at the same time [22,65]. Marriage forms may also influence age at last birth, with some evidence among polygynously married women of a ‘social menopause’ occurring when their husbands turn their mating efforts towards younger wives [19,66]. Similarly, widowhood might bring an abrupt end to reproduction in societies preventing women from remarriage [1].
Even after conception, women may terminate unwanted pregnancies with abortion—an ancient and diverse cross-cultural practice [58]. Despite the common perception that abortion is mostly practiced by young women in the USA, a study by Hill & Low [67] found a U-shaped pattern, with higher abortion rates seen among women aged 35 and older relative to women between 25 and 34 years of age. Infanticide is also found across cultures and in historical records [3]. Although infanticide by definition presumes a live birth, it has long been used as an effective method for curtailing investment in new offspring prior to menopause. Ethnographic explanations for infanticide that would correlate with maternal age include having too large a family and widowhood [68]. Caldwell & Caldwell review evidence that infanticide was widespread in agrarian Asia (India, China, and Japan) and used as way to manage problems of inheritance. In this context, family size and sex of offspring may both play an important role in determining when a woman stops reproduction [3,69,70].
5. Ultimate explanations for stopping reproduction
In addition to the proximate mechanisms detailed above, stopping reproduction calls for an evolutionary exploration in that, like menopause, it would initially seem to run counter to simple fitness-maximization models. Moreover, an evolutionary approach seems promising given that age at last birth is variable, at least partly under behavioural control, and has a close relationship to number of offspring. If stopping is part of an adaptive life-history strategy, what ultimate hypotheses could explain how it could increase fitness enough to outweigh the forgone fertility? Of direct relevance here are hypotheses for the evolution of menopause and a long post-reproductive lifespan, as well as hypotheses for optimal family size and fertility reduction seen through the demographic transition. These have been thoroughly reviewed elsewhere [9–11,71,72]. Our aim here is to highlight insights brought by considering age at last birth.
Starting with the puzzle of human menopause, initial evolutionary arguments focused on trade-offs between early and late reproduction, particularly in the face of eventual senescence [73]. For example, the costs of pregnancy and lactation might become increasingly difficult to bear as a woman's body aged, putting both her and the fetus at higher risk of mortality with each pregnancy. Consequently, menopause might allow women to channel their reproductive efforts into current offspring (and not put them in jeopardy), leading to higher lifetime fitness. Extending this explanation to age at last birth and not menopause per se, we might predict that women facing higher mortality rates in their reproductive years would tend to stop reproducing at earlier ages. Although evidence is limited, our review of age at last birth across populations suggests that it is in traditional or pre-demographic transition societies—where a woman's life expectancy is typically shorter—where we find later ages of stopping. This would seem to contradict the prediction regarding the mother's mortality risks, but other differences, for example, in infant mortality rates, make comparisons difficult.
The grandmother hypothesis for menopause and a long post-reproductive lifespan considers the inclusive fitness benefits that women might derive from shifting their energy away from direct reproduction into helping their own children to reproduce. This help could take various forms, from food provisioning, to caring for older children (for example, when a daughter has just given birth and is taking care of a newborn). Some theoretical models (e.g. [6]) also predict that it would be quite difficult to gain enough inclusive fitness benefits to favour menopause itself, which does not bode well for grandmother effects to explain stopping reproduction a decade or more before menopause. Nevertheless, if we extend the grandmother hypothesis to age at last birth, we might predict that women will be more likely to stop reproducing many years before menopause in two circumstances: (i) when their own children live close by and are themselves starting to reproduce and (ii) when their help can make a critical difference to the survival of their grandchildren [74,75].
Many studies have looked for the impact of grandmothers on grandchildren survival, with mixed results [6,9,11,76,77]. In situations where mothers are able to benefit from the help from others, we would predict that women living among investing kin would have later ages at last birth than women living neolocally, as is the case in many post-demographic transition societies. However, we might also expect that older reproducing women may find themselves with less and less support from close kin, as those relatives begin to disappear or direct their efforts towards younger women with higher reproductive value. In other words, at the same time that older women are increasing in opportunities to invest in grandchildren, they may be losing help from others. This line of thinking still focuses on opportunities for and availability of kin investment in shaping age at last birth.
By contrast, a more recent hypothesis suggests that conflict between a woman and her adult offspring could be responsible for menopause, as the reproducing generation in a family system turns over from parents to their own offspring [78,79]. As with the grandmother hypothesis, the high costs in time and energy of producing children are key, but rather than emphasizing the potential benefits of kin investment, the model focuses on competition for limited familial resources, with skewed reproduction and non-overlapping generations being the eventual outcome of this competition. Some of the strongest evidence in favour of this hypothesis comes from a recent study of pre-industrial Finnish families, where the overlapping reproduction of in-laws was associated with a sizeable drop in offspring survival [80]. In terms of reproductive senescence, this hypothesis predicts earlier menopauses in patrilocal societies, where competition is expected to be more intense. Snopkowski et al. [25], however, found no evidence for such a relationship in a study of residence patterns in Indonesia.
Anthropologists studying family systems have viewed stopping behaviour in the light of constructing families [70]. There is strong evidence from agricultural societies with heritable resources that humans deliberately shape their family sizes, and sex-composition of offspring, to meet compositional goals [5,70]. For example, historical evidence from some regions in Asia document high levels of infanticide, not just of daughters but also sons and higher parity offspring [81]. Agrarian societies are also generally where we see strong evidence of shaming and cultural taboos directed against older women giving birth (see above). In addition, in such societies finding suitable spouses and paying for the marriages of young adult children can sometimes require a considerable investment of time and resources, and parental attention may be increasingly focused on the high costs of getting the older children ‘fledged’ rather than taking care of a newborn.
These last examples bring us back to parental investment explanations for reproductive cessation. Rather than focus on menopause or the long reproductive lifespan, a parental investment perspective focuses our attention on optimal, not maximal, family sizes [9]. Although such models have recently concentrated on explaining the demographic transition, they are not limited to such. Humans invest enormous energy into producing and raising offspring that have a very long period of dependency. Under such circumstances, ‘front-loading’ reproduction and then stopping early may be the winning fitness strategy [8,9]. This may be especially true in societies with heritable wealth or high costs of education and producing competitive children [65]. Age at last birth is thus both a reflection and outcome of parental strategizing, when such strategizing is favoured, and is highly facultative in a way that menopause is not.
6. Conclusion
Stopping reproduction and the age at which a woman last gives birth have important implications, both theoretical and empirical, for a variety of academic fields. For human reproductive biology and a medical perspective, the proximate constraints on fertility and the probability of conception are of huge social relevance as women try to conceive ‘late’ in life and experience fertility problems [48]. The age at last pregnancy also has important health consequences for both the mother and developing fetus, as such pregnancies are correlated with higher risks. From an evolutionary perspective, the issues go beyond the questions of menopause and the post-reproductive lifespan, and include implications for population growth and evolutionary models of fitness and generation times [8].
In terms of life-history theory, stopping reproduction also differs from menopause in a number of interesting ways. As noted above, even if it is ultimately right-censored by menopause, age at last birth is often more variable than menopause both within and across populations. Unlike menopause, stopping behaviour is also more amenable to direct control. This is not to say that age at menopause exhibits no systematic or even adaptive variation—e.g. through genetic, developmental or nutritional affects—but compared to last birth, there is no evidence that women can easily or consciously shift their age at menopause through non-medical approaches. By contrast, behavioural influences on age at last birth include a vast array of options (e.g. through mating status, coital frequency, contraception or infanticide), and may even extend beyond the woman herself to include social pressure from others and normative cultural practices. Another reason evolutionary anthropologists should be more interested in age at last birth is that, as demographers realized long ago, it is more tightly correlated with family size than is age at menopause. As evolutionary anthropologists are more interested in reproductive success (number of surviving children) rather than fecundity per se, it makes sense to focus on a measure that is closer to the outcome variable. Last, but certainly not least, stopping reproduction is something that fathers do too. Even if men are physiologically capable of reproducing later in life, the majority do not, particularly in populations with monogamous marriage systems [65]. In the context of menopause, men's influence on family size has often been ignored, but men who reproduce do have an age at which they last father a surviving baby, and many of the behaviours that impact this age are under the influence of both the mother and father [65].
Our goal has been to bring attention to age at last birth and stopping reproduction as an important aspect of human life history. To date, evolutionary anthropologists have focused their attention on menopause and the post-reproductive lifespan rather than taking a more direct look at when women actually have their last baby. Too often, age at last birth is assumed to be relatively invariant, when in fact, it is highly variable. Demographers, in contrast, have much less to say about menopause, but have paid considerable attention to age-specific fertility rates and identifying stopping behaviour in demographic data. Their desire to estimate underlying fertility rates at the population level, however, ignores the sometimes messy, lived reproductive histories of women—for example, by focusing only on marital fertility in the absence of any control. We provide evidence both across and within populations that variation in age at last birth is substantial. For example, among Bangladeshi women, age at last birth was more variable (s.d. 5.5) than age at menopause (s.d. 3.5), with over half of all women stopping a decade or more before menopause, but also a smaller number of women continuing to reproduce into their late 40s. Rather than ignore this variation, evolutionary anthropologists should consider age at last birth in the same way that they consider age at first birth—an event with both physiological and behavioural underpinnings. We should expand our view of the end of a woman's reproductive lifespan to include not just her last menstrual cycle, but more importantly from a fitness perspective, her last successful conception.
Acknowledgements
Thanks to all NESCent working group members for their feedback and encouragement, with special gratitude to Mary Shenk, David Lawson, and Rebecca Sear. We also value the contributions of Betsy Towner Levine, Barney Luttbeg, Kristin Snopkowski, Jillian Wormington, and two anonymous reviewers.
Competing interests
We declare we have no competing interests.
Funding
I.N. was supported by a fellowship from the Foundation for Polish Science, and S.W. by the NSF REU Biological Basis of Human and Animal Behavior program (NSF SMA 1358847).
References
- 1.Coale AJ. 1984. The demographic transition. Pak. Dev. Rev. 23, 531–552. [PubMed] [Google Scholar]
- 2.Wood JW. 1994. Dynamics of human reproduction: biology, biometry, demography. New York, NY: Aldine de Gruyter. [Google Scholar]
- 3.Hrdy SB. 1999. Mother nature: a history of mothers, infants, and natural selection. New York, NY: Pantheon Books. [DOI] [PubMed] [Google Scholar]
- 4.Ellison PT. 2001. On fertile ground. Cambridge, MA: Harvard University Press. [Google Scholar]
- 5.Bongaarts J, Potter RG. 1983. Fertility, biology, and behavior: an analysis of the proximate determinants. New York, NY: Academic Press. [Google Scholar]
- 6.Hill K, Hurtado AM. 1991. The evolution of premature reproductive senescence and menopause in human females: an evaluation of the ‘grandmother hypothesis’. Hum. Nat. 2, 313–350. ( 10.1007/BF02692196) [DOI] [PubMed] [Google Scholar]
- 7.Lawson DW, Mace R. 2011. Parental investment and the optimization of human family size. Phil. Trans. R. Soc. B 366, 333–343. ( 10.1098/rstb.2010.0297) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Jones JH, Bird RB. 2014. The marginal valuation of fertility. Evol. Hum. Behav. 35, 65–71. ( 10.1016/j.evolhumbehav.2013.10.002) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Peccei JS. 2001. Menopause: adaptation or epiphenomenon? Evol. Anthropol. 10, 43–57. ( 10.1002/evan.1013) [DOI] [Google Scholar]
- 10.Hawkes K, Coxworth JE. 2013. Grandmothers and the evolution of human longevity: a review of findings and future directions. Evol. Anthropol. 22, 294–302. ( 10.1002/evan.21382) [DOI] [PubMed] [Google Scholar]
- 11.Shanley DP, Sear R, Mace R, Kirkwood TBL. 2007. Testing evolutionary theories of menopause. Proc. R. Soc. B 274, 2943–2949. ( 10.1098/rspb.2007.1028) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sievert LL. 2014. Anthropology and the study of menopause: evolutionary, developmental, and comparative perspectives. Menopause 21, 1151–1159. ( 10.1097/GME.0000000000000341) [DOI] [PubMed] [Google Scholar]
- 13.Helle S, Lummaa V, Jokela J. 2005. Are reproductive and somatic senescence coupled in humans? Late, but not early, reproduction correlated with longevity in historical Sami women. Proc. R. Soc. B 272, 29–37. ( 10.1098/rspb.2004.2944) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Menken J, Duffy L, Kuhn R. 2003. Childbearing and women's survival: new evidence from rural Bangladesh. Popul. Dev. Rev. 29, 405–426. ( 10.1111/j.1728-4457.2003.00405.x) [DOI] [Google Scholar]
- 15.Henry L. 1961. La fécondité naturelle. Observation - théorie - résultats. Population 16, 625–636. ( 10.2307/1526583) [DOI] [Google Scholar]
- 16.Coale AJ, Trussell TJ. 1974. Model fertility schedules: variations in the age structure of childbearing in human populations. Popul. Index 40, 185–258. ( 10.2307/2733910) [DOI] [PubMed] [Google Scholar]
- 17.O'Connor KA, Holman DJ, Wood JW. 1998. Declining fecundity and ovarian ageing in natural fertility populations. Maturitas 30, 127–136. ( 10.1016/S0378-5122(98)00068-1) [DOI] [PubMed] [Google Scholar]
- 18.Pennington R, Harpending H. 1993. The structure of an African pastoralist community: demography, history, and ecology of the Ngamiland Herero. Oxford, UK: Oxford University Press. [Google Scholar]
- 19.Borgerhoff Mulder M. 1989. Menarche, menopause and reproduction in the Kipsigis of Kenya. J. Biosoc. Sci. 21, 179–192. [DOI] [PubMed] [Google Scholar]
- 20.Varea C. 1993. Marriage, age at last birth and fertility in a traditional Moroccan population. J. Biosoc. Sci. 25, 1–15. ( 10.1017/S002193200002023X) [DOI] [PubMed] [Google Scholar]
- 21.Horne AD, Suchindran CM. 1988. Maternal age at last birth in Egypt. J. Biosoc. Sci. 20, 313–320. ( 10.1017/S0021932000006647) [DOI] [PubMed] [Google Scholar]
- 22.Sear R, Mace R, McGregor IA. 2000. Maternal grandmothers improve nutritional status and survival of children in rural Gambia. Proc. R. Soc. Lond. B 267, 1641–1647. ( 10.1098/rspb.2000.1190) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Howell N. 2000. Demography of the Dobe !Kung, 2nd edn New Brunswick, NJ: Aldine Transaction. [Google Scholar]
- 24.Gajanayake I. 1987. Cessation of childbearing in the absence of contraception in Sri-Lanka. J. Biosoc. Sci. 19, 65–71. ( 10.1017/S002193200001662X) [DOI] [PubMed] [Google Scholar]
- 25.Snopkowski K, Moya C, Sear R. 2014. A test of the intergenerational conflict model in Indonesia shows no evidence of earlier menopause in female-dispersing groups. Proc. R. Soc. B 281, 20140580. ( 10.1098/rspb.2014.0580) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lahdenperä M, Mar KU, Lummaa V. 2014. Reproductive cessation and post-reproductive lifespan in Asian elephants and pre-industrial humans. Front. Zool. 11, 54 ( 10.1186/s12983-014-0054-0) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Korpelainen H. 2000. Fitness, reproduction and longevity among European aristocratic and rural Finnish families in the 1700s and 1800s. Proc. R. Soc. Lond. B 267, 1765–1770. ( 10.1098/rspb.2000.1208) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Knodel J. 1979. From natural fertility to family limitation: the onset of fertility transition in a sample of German villages. Demography 16, 493–521. ( 10.2307/2060931) [DOI] [PubMed] [Google Scholar]
- 29.Dribe M. 2004. Long-term effects of childbearing on mortality: evidence from pre-industrial Sweden. Popul. Stud. J. Demogr. 58, 297–310. ( 10.1080/0032472042000272357) [DOI] [PubMed] [Google Scholar]
- 30.Desjardins B, Bideau A, Brunet G. 1994. Age of mother at last birth in two historical populations. J. Biosoc. Sci. 26, 509–516. ( 10.1017/S0021932000021635) [DOI] [PubMed] [Google Scholar]
- 31.Van Bavel J, Kok J. 2005. The role of religion in the Dutch fertility transition: starting, spacing, and stopping in the heart of the Netherlands, 1845–1945. Contin. Change 20, 247–263. ( 10.1017/S0268416005005473) [DOI] [Google Scholar]
- 32.Alter G, Dribe M, Van Poppel F. 2007. Widowhood, family size, and post-reproductive mortality: a comparative analysis of three populations in nineteenth-century Europe. Demography 44, 785–806. ( 10.1353/dem.2007.0037) [DOI] [PubMed] [Google Scholar]
- 33.Hionidou V. 1998. The adoption of fertility control on Mykonos, 1879–1959: stopping, spacing or both? Popul. Stud. J. Demogr. 52, 67–83. ( 10.1080/0032472031000150186) [DOI] [PubMed] [Google Scholar]
- 34.Gama A. 2011. The reproductive period and fertility in rural Portuguese women: time trends of biological and behavioural determinants. Ann. Hum. Biol. 38, 655–658. ( 10.3109/03014460.2011.572916) [DOI] [PubMed] [Google Scholar]
- 35.Grundy E, Kravdal O. 2008. Reproductive history and mortality in late middle age among Norwegian men and women. Am. J. Epidemiol. 167, 271–279. ( 10.1093/aje/kwm295) [DOI] [PubMed] [Google Scholar]
- 36.Colleran H, Mace R. 2015. Social network- and community-level influences on contraceptive use: evidence from rural Poland. Proc. R. Soc. B 282, 20150398 ( 10.1098/rspb.2015.0398) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Charbonneau H. 1979. Les régimes de fécondité naturelle en Amérique du Nord. Natural fertility. Patterns and determinants of natural fertility: proceedings of a seminar on natural fertility (eds Leridon H, Menken J), pp. 442–491. Liege, Belgium: Ordina Editions. [Google Scholar]
- 38.McArdle PF, et al. 2006. Does having children extend life span? A genealogical study of parity and longevity in the Amish. J. Gerontol. Biol. Sci. Med. Sci. 61, 190–195. ( 10.1093/gerona/61.2.190) [DOI] [PubMed] [Google Scholar]
- 39.Smith KR, Mineau GP, Bean LL. 2002. Fertility and post-reproductive longevity. Soc. Biol. 49, 185–205. ( 10.1080/19485565.2002.9989058) [DOI] [PubMed] [Google Scholar]
- 40.Kosova G, Abney M, Ober C. 2010. Colloquium papers: heritability of reproductive fitness traits in a human population. Proc. Natl Acad. Sci. USA 107(Suppl 1), 1772–1778. ( 10.1073/pnas.0906196106) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shoemaker N. 1999. American Indian population recovery in the twentieth century. Albuquerque, NM: University of New Mexico Press. [Google Scholar]
- 42.Kramer K. 2005. Maya children: helpers at the farm. Cambridge, MA: Harvard University Press. [Google Scholar]
- 43.Hill K, Hurtado AM. 1996. Ache life history: the ecology and demography of a foraging people. New York, NY: Aldine de Gruyter. [Google Scholar]
- 44.Pascual J, García-Moro CE, Hernández M. 2005. Biological and behavioral determinants of fertility in Tierra del Fuego. Am. J. Phys. Anthropol. 127, 105–113. ( 10.1002/ajpa.20065) [DOI] [PubMed] [Google Scholar]
- 45.Hawkes K, Smith KR. 2010. Do women stop early? Similarities in fertility decline in humans and chimpanzees. Ann. NY Acad. Sci. 1204, 43–53. ( 10.1111/j.1749-6632.2010.05527.x) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rahman O, Menken J, Foster A, Peterson CE, Khan MN, Kuhn R, Gertler P. 1999. The Matlab health and socioeconomic survey: overview and user's guide. In DRU-2018/1-NIA. Santa Monica, CA, USA.
- 47.Shenk MK, Towner MC, Kress HC, Alam N. 2013. A model comparison approach shows stronger support for economic models of fertility decline. Proc. Natl Acad. Sci. USA 110, 8045–8050. ( 10.1073/pnas.1217029110) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Eijkemans MJC, van Poppel F, Habbema DF, Smith KR, Leridon H, te Velde ER. 2014. Too old to have children? Lessons from natural fertility populations. Hum. Reprod. 29, 1304–1312. ( 10.1093/humrep/deu056) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Shenk MK, Starkweather K, Kress HC, Alam N. 2013. Does absence matter? A comparison of three types of father absence in rural Bangladesh. Hum. Nat. 24, 76–110. [DOI] [PubMed] [Google Scholar]
- 50.Koenig MA, Rob U, Khan MA, Chakraborty J, Fauveau V. 1992. Contraceptive use in Matlab, Bangladesh in 1990: levels, trends, and explanations. Stud. Family Plann. 23, 352–364. ( 10.2307/1966893) [DOI] [PubMed] [Google Scholar]
- 51.Ewbank DC. 1989. Estimating birth stopping and spacing behavior. Demography 26, 473–483. ( 10.2307/2061607) [DOI] [PubMed] [Google Scholar]
- 52.Strassmann BI, Warner JH. 1998. Predictors of fecundability and conception waits among the Dogon of Mali. Am. J. Phys. Anthropol. 105, 167–184. () [DOI] [PubMed] [Google Scholar]
- 53.Okun BS. 1995. Distinguishing stopping behavior from spacing behavior with indirect methods. Hist. Methods 28, 85–96. ( 10.1080/01615440.1995.9956357) [DOI] [Google Scholar]
- 54.Van Bavel J. 2004. Distinguer contraception d'arrêt et contraception d'espacement: revue des méthodes en démographie historique. Population 59, 119–132. ( 10.3917/popu.401.0119) [DOI] [Google Scholar]
- 55.O'Connor KA, Holman DJ, Wood JW. 2001. Menstrual cycle variability and the perimenopause. Am. J. Hum. Biol. 13, 465–478. ( 10.1002/ajhb.1078) [DOI] [PubMed] [Google Scholar]
- 56.Wood JW, Holman DJ, Yashin AI, Peterson RJ, Weinstein M, Chang MC. 1994. A multistate model of fecundability and sterility. Demography 31, 403–426. ( 10.2307/2061750) [DOI] [PubMed] [Google Scholar]
- 57.Mant J, Painter R, Vessey M. 1997. Epidemiology of genital prolapse: observations from the Oxford Family Planning Association Study. Brit. J. Obstet. Gynaecol. 104, 579–585. ( 10.1111/j.1471-0528.1997.tb11536.x) [DOI] [PubMed] [Google Scholar]
- 58.Riddle JM. 1992. Contraception and abortion from the ancient world to the Renaissance. Cambridge, MA: Harvard University Press. [Google Scholar]
- 59.United Nations DoEaSA, Population Division. 2013. World Contraceptive Patterns 2013 (ST/ESA/SER.A/326).
- 60.Ruzicka LT, Bhatia S. 1982. Coital frequency and sexual abstinence in rural Bangladesh. J. Biosoc. Sci. 14, 397–420. ( 10.1017/S0021932000014280) [DOI] [PubMed] [Google Scholar]
- 61.Strassmann BI. 1992. The function of menstrual taboos among the Dogon: defense against Cuckoldry? Hum. Nat. 3, 89–131. ( 10.1007/BF02692249) [DOI] [PubMed] [Google Scholar]
- 62.Jasienska G. 2013. The fragile wisdom: an evolutionary view on women's biology and health. Cambridge, MA: Harvard University Press. [Google Scholar]
- 63.Caldwell JC, Caldwell P. 1977. Role of marital sexual abstinence in determining fertility—study of Yoruba in Nigeria. Popul. Stud. J. Demogr. 31, 193–217. ( 10.2307/2173915) [DOI] [PubMed] [Google Scholar]
- 64.Fine A, Moulinie V, Sangoi JC. 2009. From mother to daughter: the passing of fertility. L'Homme 191, 37–76. [Google Scholar]
- 65.Kaplan H, Gurven M, Winking J, Hooper PL, Stieglitz J. 2010. Learning, menopause, and the human adaptive complex. Ann. NY Acad. Sci. 1204, 30–42. ( 10.1111/J.1749-6632.2010.05528.x) [DOI] [PubMed] [Google Scholar]
- 66.Bove R, Valeggia C. 2009. Polygyny and women's health in sub-Saharan Africa. Soc. Sci. Med. 68, 21–29. ( 10.1016/j.socscimed.2008.09.045) [DOI] [PubMed] [Google Scholar]
- 67.Hill EM, Low BS. 1992. Contemporary abortion patterns—a life-history approach. Ethol. Sociobiol. 13, 35–48. ( 10.1016/0162-3095(92)90005-O) [DOI] [PubMed] [Google Scholar]
- 68.Hausfater G, Hrdy SB. 1984. Infanticide: comparative and evolutionary perspectives. New York, NY: Aldine Pub. Co. [Google Scholar]
- 69.Hrdy SB, Judge DS. 1993. Darwin and the puzzle of primogeniture: an essay on biases in parental investment after death. Hum. Nat. Interdiscip. Biosoc. Persp. 4, 1–45. ( 10.1007/BF02734088) [DOI] [PubMed] [Google Scholar]
- 70.Skinner GW. 1997. Family systems and demographic processes. In Anthropological demography: toward a new synthesis (eds Kertzer DI, Fricke T), pp. 53–95. Chicago, IL: The University of Chicago Press. [Google Scholar]
- 71.Lawson DW, Alvergne A, Gibson MA. 2012. The life-history trade-off between fertility and child survival. Proc. R. Soc. B 279, 4755–4764. ( 10.1098/rspb.2012.1635) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sear R, Coall D. 2011. How much does family matter? Cooperative breeding and the demographic transition. Popul. Dev. Rev. 37, 81–112. ( 10.1111/j.1728-4457.2011.00379.x) [DOI] [PubMed] [Google Scholar]
- 73.Williams GC. 1957. Pleiotropy, natural-selection, and the evolution of senescence. Evolution 11, 398–411. ( 10.2307/2406060) [DOI] [Google Scholar]
- 74.Levitis DA, Burger O, Lackey LB. 2013. The human post-fertile lifespan in comparative evolutionary context. Evol. Anthropol. 22, 66–79. ( 10.1002/evan.21332) [DOI] [PubMed] [Google Scholar]
- 75.Judge DS, Carey JR. 2000. Postreproductive life predicted by primate patterns. J. Gerontol. A Biol. Sci. Med. Sci. 55, B201–B209. ( 10.1093/gerona/55.4.B201) [DOI] [PubMed] [Google Scholar]
- 76.Skjaervo GR, Roskaft E. 2013. Menopause: no support for an evolutionary explanation among historical Norwegians. Exp. Gerontol. 48, 408–413. ( 10.1016/j.exger.2013.02.001) [DOI] [PubMed] [Google Scholar]
- 77.Sear R, Mace R. 2008. Who keeps children alive? A review of the effects of kin on child survival. Evol. Hum. Behav. 29, 1–18. ( 10.1016/j.evolhumbehav.2007.10.001) [DOI] [Google Scholar]
- 78.Cant MA, Johnstone RA. 2008. Reproductive conflict and the separation of reproductive generations in humans. Proc. Natl Acad. Sci. USA 105, 5332–5336. ( 10.1073/pnas.0711911105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Mace R, Alvergne A. 2012. Female reproductive competition within families in rural Gambia. Proc. R. Soc. B 279, 2219–2227. ( 10.1098/rspb.2011.2424) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Lahdenpera M, Gillespie DOS, Lummaa V, Russell AF. 2012. Severe intergenerational reproductive conflict and the evolution of menopause. Ecol. Lett. 15, 1283–1290. ( 10.1111/j.1461-0248.2012.01851.x) [DOI] [PubMed] [Google Scholar]
- 81.Caldwell JC, Caldwell BK. 2005. Family size control by infanticide in the Great Agrarian Societies of Asia. J. Comp. Family Stud. 36, 205. [Google Scholar]