

Abstract
Global marine biodiversity peaks within the Coral Triangle, and understanding how such high diversity is maintained is a central question in marine ecology. We investigated broad-scale patterns in the diversity of clownfishes and their host sea anemones by conducting 981 belt-transects at 20 locations throughout the Indo-Pacific. Of the 1508 clownfishes encountered, 377 fish occurred in interspecific cohabiting groups and cohabitation was almost entirely restricted to the Coral Triangle. Neither the diversity nor density of host anemone or clownfish species alone influenced rates of interspecific cohabitation. Rather cohabitation occurred in areas where the number of clownfish species exceeds the number of host anemone species. In the Coral Triangle, cohabiting individuals were observed to finely partition their host anemone, with the subordinate species inhabiting the periphery. Furthermore, aggression did not increase in interspecific cohabiting groups, instead dominant species were accepting of subordinate species. Various combinations of clownfish species were observed cohabiting (independent of body size, phylogenetic relatedness, evolutionary age, dentition, level of specialization) in a range of anemone species, thereby ensuring that each clownfish species had dominant reproductive individuals in some cohabiting groups. Clownfishes are obligate commensals, thus cohabitation is an important process in maintaining biodiversity in high diversity systems because it supports the persistence of many species when host availability is limiting. Cohabitation is a likely explanation for high species richness in other obligate commensals within the Coral Triangle, and highlights the importance of protecting these habitats in order to conserve unique marine biodiversity.
Keywords: amphiprion, anemonefish, biodiversity, coral reefs, mutualism, symbiosis
1. Introduction
Determining how multiple species manage to coexist is a central theme in ecology [1]. One explanation is that species differ in niche occupancy [2]. Through mechanisms such as niche differentiation [3], including social and behavioural specializations [4] and resource partitioning (e.g. space and habitat use), coexistence of different species can occur [5]. Despite extensive studies into the variation in physiological, morphological and demographic traits across coexisting species [6,7], a range of hypotheses exist regarding the relative importance of these different niche components in maintaining diversity [8].
Coral reefs are the most biodiverse marine ecosystem [9]. Within reef systems, there are numerous examples of species occupying different niches to avoid competition [10,11]. It has also been proposed that some coral-reef species coexist using the same resources through lottery competition [12,13] or through cohabitation [14]. Under cohabitation, multiple species appear to occupy the same niche in the same habitat, but although some species may be competitively superior, others may have different advantageous traits (e.g. higher colonization rates) that enable cohabitation to persist. It may also be too difficult to exclude the competitively inferior species [14].
Reef fishes comprise the most diverse vertebrate assemblages in the world and the greatest diversity of reef fish occurs in the East Indian region (Eastern Indonesia, New Guinea and the Philippines [15]) of the Indo-Pacific's Coral Triangle [16,17]. The Coral Triangle only covers a surface area of 3% of the tropical Indo-West and Central Pacific, but contains 52% of its total reef fish species [17]. Cohabitation has been observed within [18,19] and between coral reef fish families [20] and if cohabitation facilitates high species richness we would hypothesize that the proportion of cohabiting fishes would increase with proximity to their centre of biodiversity (i.e. the Coral Triangle).
In this study, we investigate the diversity and abundance of clownfishes (family Pomacentridae) and their host sea anemones (order Actiniaria) at 20 locations across the Indo-Pacific to determine patterns of interspecific cohabitation and to explore what factors facilitate cohabitation. Clownfishes are small bodied (maximum size 8–16 cm) and predominantly feed on plankton in the water column and algae around their host anemones [21]. They form small groups with a strong hierarchical social structure based on body size. They are protandrous hermaphrodites with the largest fish being a dominant female, second largest an adult male and the rest of the group are immature [21]. Clownfishes are an ideal model group because their taxonomy and phylogeny are well resolved and, like most coral reef organisms, their highest diversity is located in the Coral Triangle [14]. They are habitat specialists that cannot persist in nature without host anemones and demonstrate strict host specificity, limiting which anemone species are available for occupancy [22]. There are 28 recognized species of clownfish [14], but only 10 species of host anemones [14,21] and this can lead to intra- and interspecific competition [22–25]. Thus, in areas where there is a high abundance and/or diversity of clownfishes there may not be enough host anemone species. Rather than competing for limited available resources, cohabitation may facilitate high clownfish diversity.
Different drivers could affect clownfish interspecific cohabitation and we therefore set out to systematically test a hierarchy of hypotheses that could explain clownfish cohabitation patterns. We hypothesize that: (i) the proportion of clownfishes that exhibit interspecific cohabitation increases with decreasing abundance and/or diversity of anemone hosts; (ii) interspecific cohabitation is highest in regions of high clownfish density and/or diversity regardless of host anemone availability; (iii) differences in the abundance of host anemones across reef zones and selective settlement patterns of clownfishes provide spatially explicit abundance patterns that influence the proportion of cohabitating clownfishes; (iv) larger host anemones support a greater proportion of cohabiting clownfishes; and (v) the total body length of resident clownfish is greater in intraspecific social groups. We also examine which traits (e.g. trophic guild, size, morphology and evolutionary age) are linked to cohabitation in clownfishes. Finally, we determine whether cohabitation is facilitated by: (i) reduced aggression and/or (ii) fine-scale spatial separation of the host anemone. We assemble the most comprehensive dataset to-date of clownfish habitat use and document the highest incidence of clownfish interspecific cohabitation, which appears to be driven by clownfish species richness in high biodiversity areas. Thus, we contextualize the role of cohabitation in supporting species diversity within the Coral Triangle.
2. Material and methods
(a). Clownfish cohabitation frequency
To establish patterns of clownfish interspecific cohabitation across the Indo-Pacific, host anemone and clownfish populations were investigated at 20 locations between 2005 and 2014. A further eight surveyed locations (Djibouti, Norfolk Island and six locations in the Red Sea) were excluded from the analyses, because only one clownfish species occurs at these locations and thus cohabitation is impossible. To assess the diversity, abundance and cohabitation of clownfishes across the Indo-Pacific, underwater visual surveys were conducted at the 20 locations, covering a wide range of reef zones (e.g. reef flat, crest and slope) and depths (see electronic supplementary material, table S1). The number and species of clownfish occupying a single-host anemone were defined as a social group and were assessed using standardized sampling effort, either through fixed reef area sampling 250 m2 (50 × 5 m) or restricted effort dives with multiple replicates and sites per location (see electronic supplementary material, table S1 for detailed survey methodology). Within our analysis, we also included published data from Riccardi et al. [26] and Elliott & Mariscal [27] to achieve 22 study locations (electronic supplementary material, table S1).
(i). Host anemone size
On Hoga Island, where the highest frequency of interspecific cohabitation was recorded, we tested whether cohabitation increased with host anemone size. Hoga Island is a UNESCO Biosphere Reserve and is situated in the Wakatobi Marine National Park (WMNP), southeast Sulawesi, Indonesia, in the central area of the Coral Triangle [28]. From June to August 2014, the western reefs of Hoga Island were surveyed. The Hoga reef system is a typical fringing reef with a shallow reef flat (1–4 m), reef crest (4–6 m) and a reef slope extending down to 50 m depth. Sixty-five belt-transects (50 × 5 m) were randomly placed on the reef flat to upper reef slope at depths of 1–5 m (total area surveyed: 16 250 m2).
The surface area of the oral disc of all recorded anemones was determined by using the advanced geometry technique typically applied to corals [29]. In this method, the anemone is visually divided into sections in situ, with each segment assigned a geometric form or shape. The majority of anemones had an oval shape, so the longest axial length and shortest axial length (measured to the nearest centimetre) were multiplied by π/4 [30]. Field surveys were completed between 10.00 and 15.00 h to minimize any diel variation in changing tentacle size [31]. For the three most common anemones on Hoga reefs (Entacmaea quadricolor, Heteractis crispa and Heteractis magnifica), the surface areas of three individuals per species were measured repeatedly over 3 days to determine natural variation in size. A general linear model demonstrated that size did not differ across the repeated measures.
(ii). Clownfish total length
To determine whether cohabitation of clownfishes was influenced by the total length of all clownfishes in a host anemone, fish observed in the Hoga surveys were categorized as either: new recruit (less than 2 cm), juvenile (2–5 cm) or adult (more than 5 cm) [21]. Fish were observed from a distance of 2 m. No detectable effect on fish behaviour was observed at this distance when compared with observations carried out a greater distance. Assessment of fish size followed specific training on underwater visual size estimation.
(b). Characteristics of cohabiting clownfish
To elucidate whether certain traits were linked to clownfish cohabitation, and to quantify the composition of different social groups, detailed surveys were conducted at Hoga. All social groups were surveyed by a 2-min observation from 2 m away. The area of host anemone available per fish in both inter- and intraspecific social groups was determined.
(c). Mechanisms that support clownfish cohabitation
(i). Clownfish aggression
To determine whether interspecific cohabitation is supported by reduced clownfish aggression, randomly selected inter- and intraspecific social groups encountered in the belt-transects at Hoga were assessed. Aggression was defined as rapid swimming towards other fish, head-dips, jaw clicking and excessive dorsal/ventral leaning [32]. For interspecific social groups, two factors were recorded to determine dominant and subordinate clownfishes. The first factor was which species had the largest individual fish and the second factor was which species showed the most aggression [26,32]. A size-based dominance hierarchy has been well documented for intraspecific social groups of clownfishes [33]. The percentage time of aggression was used to quantify the extent of aggression between cohabiting species. Observations were conducted on the dominant fish within each social group. Behavioural observations (n = 45) were conducted for 10-min, as preliminary observations (n = 12) demonstrated this to be most effective time for sampling the range of behavioural traits demonstrated by an individual.
(ii). Spatial separation of host anemone
To test whether fine-scale spatial separation facilitates interspecific cohabitation, we recorded the anemone host usage of the dominant (largest) clownfish of each social group observed at Hoga. Five-minute high-definition videos (Olympus E-PL5 in Olympus PT-EP10 underwater housing) were taken per social group with cameras set at least 2-m away from the focal anemone. Each video was analysed by superimposing a 6 × 6 grid (n = 36) over the anemone and documenting the amount of time the dominant fish spent in each square. Grids were always 6 × 6, but were customized and scaled for each anemone to ensure the grid cells covered the entire anemone (range in cell area 4.0–6.25 cm2). Times when fish moved out of the anemone or were hidden within the anemone were removed from the analysis. The proportion of time when clownfish were out of sight did not correspond to any particular part of the anemone.
(d). Data analysis
(i). Clownfish cohabitation
Cohabitation frequency was determined for all locations as the proportion of clownfish social groups that contained more than one species of clownfish. Within this study, Hoga was centrally located within the Coral Triangle [28] and to assess how both cohabitation and clownfish diversity changed with distance from Hoga a Pearson's Correlation was conducted. The distance of each site from Hoga was determined using their GPS points entered into Google Earth as waypoints.
A Pearson's Correlation was also used to test if cohabitation was positively associated with the density of anemones, anemone diversity (Shannon–Wiener Diversity Index (SW-Index)), the density of clownfish and/or clownfish diversity (SW-Index). Note: n = 20 as data from Riccardi et al. [26] and Elliott & Mariscal [27] were not included as detailed diversity data were unavailable. ANOVA was used to test whether the reef zone influenced the proportion of interspecific cohabitation across sites. Next, the composition of host anemone species and its influence on cohabitation at each site were examined via ordinations based on principal component analysis (PCA). Pearson's Correlation coefficient was used to examine whether extracted ordination axes were correlated with the proportion of cohabitation. Finally, an ANOVA with post hoc Tukey Kramer test was used to determine whether the ratio of clownfish species to host species influenced cohabitation.
(ii). Host anemone size and clownfish total length
To determine whether there was an association between the surface area of the oral disc of the anemones and the number of clownfish, a Pearson's Correlation was conducted for each of the common anemone species recorded in surveys at Hoga Island (Heteractis aurora, H. magnifica and Stichodactyla mertensii were excluded because n < 4). In the absence of site-specific knowledge of the relationship between fish length and biomass, the same analysis was conducted to compare the association between aggregate fish length per social group (calculated as the sum of the total lengths of all fish in the group) and anemone size (oral disc surface area) for both inter- and intraspecific social groups.
(iii). Characteristics of cohabiting clownfish
For the Hoga data, a Linear Mixed Effects (LME) model was used to determine whether cohabiting species exhibited similar traits. The model included mean maximum body size [21], evolutionary age (determined from [34]), host specialization (e.g. number of hosts the fish will use [21]), feeding preference [35], teeth structure [21,35,36], whether the individual was dominant or subordinate at Hoga, and the variance-to-mean-ratio (VMR) to represent host usage. Parameter estimation in LME models was based on restricted maximum likelihood (REML). A Pearson's Correlation was conducted to determine how the degree of phylogenetic relatedness (determined as the cumulative number of divergent branches of each clownfish species from their common ancestor; phylogeny from [34]) affected the proportion of cohabitation between clownfish species.
(iv). Clownfish aggression
To test whether reduced clownfish aggression supported interspecific cohabitation of clownfish, the amount of time spent on aggression (inter- and intraspecific) was compared between inter- and intraspecific social groups using a two-way ANOVA. Tests focused on the two most common cohabiting (66% of cohabitation) clownfishes at Hoga, Amphiprion clarkii and Amphiprion perideraion.
(v). Spatial separation of host anemone
To elucidate whether clownfishes in interspecific social groups used fine-scale spatial separation of the host anemone, we calculated the Index of Dispersion (VMR) for the dominant and subordinate fish of each social group at Hoga. The VMR was calculated as
| 2.8 |
where σ2 is the variance and μ is the mean percent time each clownfish spent in each of the 36 sections of the host anemone. VMR: less than 1 = uniform distribution, 1 = random distribution and more than 1 = clustered or aggregated distribution. For the two most common interspecific social groups (A. clarkii with A. perideraion, and Premnas biaculeatus with Amphiprion melanopus), a two-way ANOVA with post hoc Tukey Kramer test was used to compare the Index of Dispersion between inter- and intraspecific social groups for each social group member. Statistical analyses were conducted in R studio [37].
3. Results
(a). Clownfish cohabitation frequency
In total, 53 525 m2 of coral reef habitat were surveyed in 981 belt-transects at 20 locations across the Indo-Pacific, and combined with published survey data from reefs in Manado and Madang [26,27]. A total of 1508 clownfish individuals were recorded from 12 species and 5039 anemones across eight host species (electronic supplementary material, table S2). Clownfish diversity was negatively correlated with distance from the centre of the Coral Triangle (figure 1 and electronic supplementary material, figure S1; r = 0.54, n = 20, p < 0.05).
Figure 1.
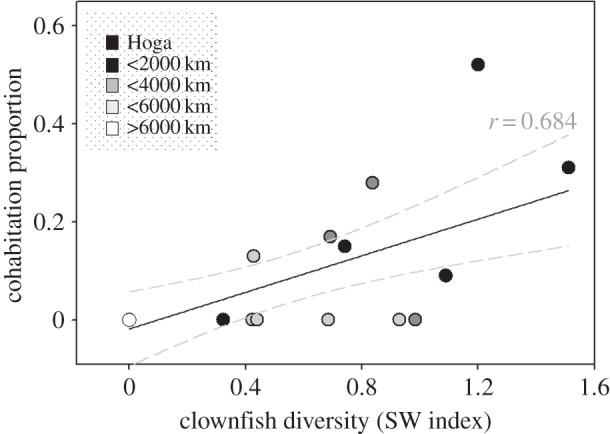
The relationship (r = 0.68, n = 20, p < 0.01) between the proportion of anemones that contained interspecific cohabiting clownfishes and clownfish diversity (S–W Index). Surveys were conducted between 2005 and 2014. The distance of each site from Hoga is illustrated by the shading of each point (see Legend). 95% confidence interval is shown (grey dashed lines).
Of the 1508 clownfish individuals that were recorded, 377 were found to be cohabitating in interspecific social groups (25%). Clownfish cohabitation was concentrated within the Coral Triangle (figure 2). The proportion of interspecific cohabitation decreased with distance from the geographic centre of the Coral Triangle (figure 1 and electronic supplementary material, figure S1; r = 0.60, n = 22, p < 0.01). Thus, interspecific cohabitation increased with clownfish diversity (figure 1; r = 0.68, n = 22, p < 0.01). The diversity of host anemones did not influence the proportion of interspecific cohabitation (electronic supplementary material, figure S2). However, interspecific cohabitation was significantly higher when the number of clownfish species exceeded the number of host anemone species (table 1; ANOVA with post hoc Tukey, p < 0.005). Clownfish and host anemone densities were not correlated with interspecific cohabitation (electronic supplementary material, figure S2) and reef zone had no influence on levels of cohabitation. Similarly, the composition of host anemones across the different reef zones did not influence interspecific cohabitation, with no correlation between the PCA extracted ordination axes and the proportion of cohabitation (electronic supplementary material, figure S3).
Figure 2.

The proportion of cohabitation in clownfishes throughout the Indo-Pacific. The proportion of cohabitating clownfish at a location was determined by dividing the number of anemones per location hosting more than one species of clownfish by the total number of anemones hosting clownfish at the location. Surveys were conducted between 2005 and 2014 and published data from Elliott & Mariscal [27] and Riccardi et al. [26] were included. Grey shading indicates the proportion of anemones that were cohabited by different species of clownfish. The black line delineates the Coral Triangle as designated by Allen & Werner [16]. The dashed circle indicates Hoga Island.
Table 1.
The proportion of anemones that contained cohabiting clownfishes for each of the 22 locations across the Indo-Pacific. Symbols indicate sites where the number of host anemone species observed in surveys was higher (+), equal to (=) or lower (−) than the number of clownfish species.
| study location | cohabitation proportion | host anemone species to fish species |
|---|---|---|
| Guam | 0 | n.a |
| Maldives | 0 | + |
| Marshall Islands | 0 | + |
| Japan | 0 | + |
| Socotra | 0 | = |
| Lord Howe Island | 0 | = |
| Elizabeth Reef | 0 | = |
| Pohnpei | 0 | = |
| Cocos Islands | 0 | = |
| Abrolhos | 0 | = |
| Ashmore Reef | 0 | = |
| Lombok | 0 | + |
| Christmas Island | 0.02 | = |
| Bali | 0.07 | − |
| Komodo | 0.08 | − |
| Sunshine Coast | 0.09 | − |
| Kosrae | 0.13 | + |
| Ningaloo | 0.17 | − |
| GBR Keppels | 0.28 | − |
| Manado | 0.31 | − |
| Madang | 0.32 | − |
| Hoga | 0.52 | − |
(i). Host anemone size and clownfish total length
Across host anemones, the aggregate size of all fish (i.e. summed total lengths) in interspecific social groups was greater than that for the intraspecific groups (t112 = 3.92, p < 0.001). There was no relationship between number of clownfish and host anemone size for both inter- and intraspecific social groups. Entacmaea quadricolor was the exception, with larger anemones hosting more clownfish (number of fish: r = 0.62, n = 13, p < 0.05; total length of fish: r = 0.71, n = 13, p < 0.01). Clownfish density was twice as high in interspecific groups (2.1 anemonefish ± 0.23 per 100 cm2 of anemone) compared with intraspecific groups (1.1 clownfish ± 0.13 per 100 cm2 of clownfish) (t = 2.31112, p < 0.05).
(b). Characteristics of cohabiting clownfish
Hoga Island had the highest level of cohabitation. Anemone cover in the Hoga survey area was 0.05% and included 437 clownfish across six species, residing in 114 host anemones across five species (electronic supplementary material, table S2). Overall, the clownfish comprised 155 adults, 194 juveniles and 88 new recruits and A. clarkii was the most abundant species (57%). The mean (±s.e.) number of clownfish per anemone was 4.1 (±0.19) and ranged from one to 12 individuals.
At Hoga, all anemones were inhabited and each clownfish species occupied between one and five species of host anemones (electronic supplementary material, table S3). 52% of anemones contained two different species of clownfish (figure 2). Cohabitation varied across species (A. perideraion = 76%, A. clarkii = 69% and A. melanopus = 67%, electronic supplementary material, table S3) with different combinations of interspecific social groups ([38], electronic supplementary material, table S4).
Cohabiting species of clownfish were not phylogenetically or morphologically similar, with maximum body length, phylogenetic relatedness and evolutionary age not associated with cohabitation levels (electronic supplementary material, table S2 and figure S4). Similarly, ecological specialization was not linked to cohabitation. Cohabitation occurred independent of clownfish dentition or feeding preference (table 2). No species was observed always to be the subordinate species within interspecific social groups and thus this factor was also non-significant in the LME model.
Table 2.
Model parameters to estimate whether clownfishes cohabit based on certain traits. Clownfish species was modelled as a random effect.
| model term | estimate | s.e. | t-value | p-value |
|---|---|---|---|---|
| intercept (cohabit) | 6.67 | 10.47 | 0.64 | n.s. |
| host specificity | −0.08 | 0.17 | 0.50 | n.s. |
| maximum body length | −0.01 | 0.02 | 0.40 | n.s. |
| teeth structure | −0.30 | 1.56 | 0.19 | n.s. |
| phylogenetic relatedness | −0.37 | 0.59 | 0.63 | n.s. |
| evolutionary age | −1.21 | 1.76 | 0.69 | n.s. |
| host anemone spatial usage (VMR score) | −0.11 | 0.07 | 0.16 | n.s. |
| subordinate or dominant | 0.50 | 0.29 | 1.74 | n.s. |
| adult or juvenile | −0.40 | 0.80 | 0.50 | n.s. |
| feeding preference | −0.01 | 0.02 | 0.06 | n.s. |
(c). Mechanisms that support clownfish cohabitation
(i). Clownfish aggression
For the frequently observed cohabiting species A. clarkii and A. perideraion (electronic supplementary material, table S4), aggression decreased when they were found in interspecific rather than intraspecific social groups (A. clarkii: F1,53 = 7.55, p < 0.01; A. perideraion F1,53 = 19.65, p < 0.001). Clear species-specific responses existed, with A. clarkii significantly more aggressive than A. perideraion in all social groups (intraspecific: F1,53 = 236.48, p < 0.01; interspecific: F1,53 = 24.71, p < 0.001). Amphiprion clarkii displayed aggression on average 14% of time in intraspecific social groups and 7% of time in interspecific social groups, making it the most aggressive clownfish observed at Hoga (figure 3). Amphiprion perideraion showed very little aggression, averaging less than 2% of time displaying antagonistic behaviour.
Figure 3.
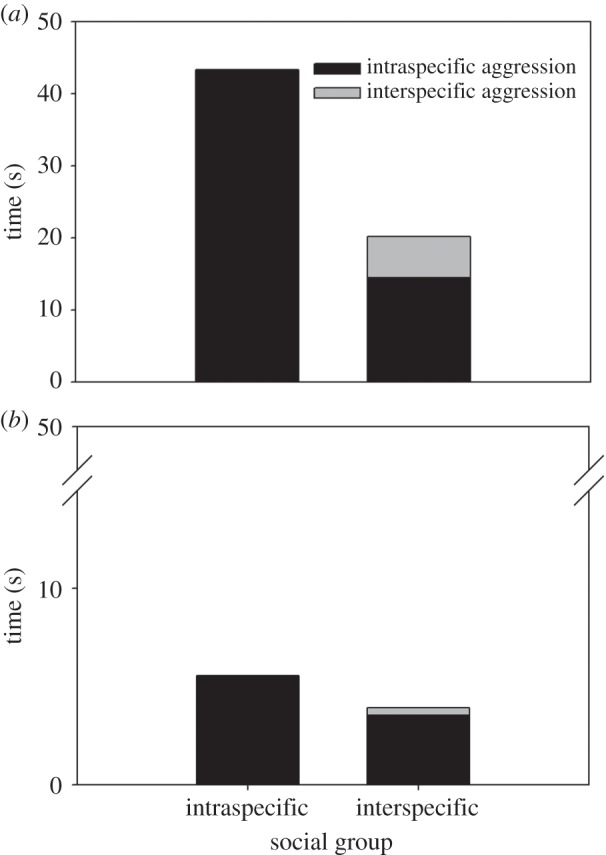
Aggression of (a) Amphiprion clarkii and (b) A. perideraion in inter- and intraspecific social groups on reefs around Hoga Island, South East Sulawesi, Indonesia. Aggression was determined by observing the dominant (largest) clownfish of each social group for a total of 10 min. In interspecific social groups, aggression was defined as intraspecific or interspecific, i.e. within or between species.
(ii). Spatial separation of host anemone
Interspecific cohabitation altered the spatial use of host anemones by the subordinate species while dominant individuals maintained similar spatial habitat usage (figure 4). Analysis on the subordinates (A. perideraion and P. biaculeatus) of the two main social groups (A. clarkii–A. perideraion and A. melanopus–P. biaculeatus) demonstrated significant clustering on the outer edge of the anemone (A. perideraion: F3,100 = 696.76, p < 0.001, post hoc Tukey, p < 0.01; P. biaculeatus: F3,16 = 1438.42, p < 0.01, post hoc Tukey, p < 0.01), a trend generally observed for all species when they were subordinate (figure 4, A. clarkii VMR > 3.1, A. perideraion VMR > 1.2 and P. biaculeatus VMR > 15.1). Dominant individuals of A. clarkii and A. melanopus showed a uniform distribution (VMR < 0.3), spending most time outside of the host anemone (figure 4, A. clarkii ca. 40% and A. melanopus ca. 50% of time). By contrast, dominant individuals of Amphiprion sandaracinos and A. perideraion occupied the centre of the anemone (VMR > 1.2, > 1.3, respectively). Dominant cohabiting A. clarkii spent 17.0 ± 1.3% more time outside of the anemone compared with when it was the sole occupant of an anemone.
Figure 4.

Spatial patterns in usage of anemones by clownfish in intra- and interspecific social groups for Hoga Island, South East Sulawesi, Indonesia. The percentage of time clownfish spent in each quadrant was determined and colour coded accordingly. Territory analysis was determined for intra- and interspecific social groups, and for the dominant (largest) and subordinate (smallest) individuals.
4. Discussion
This study demonstrates that clownfishes are rarely found cohabiting anemones outside of the Coral Triangle (figure 2). Across the Indo-Pacific, total anemone cover is often very low [38] and as the majority of clownfish species exhibit host specificity the availability of appropriate habitat is even lower [39]. Anemone density, diversity and clownfish diversity did not influence the proportion of cohabitating clownfishes. Reef zone did not influence the proportion of cohabitation, and the composition of host anemones did not vary across reef zones. By contrast, Elliott & Mariscal [27] found niche differentiation of clownfish across different reef zones and that this influenced cohabitation. However, Elliott & Mariscal [27] classified reef zones at a far larger spatial scale (including near shore, mid-lagoon, outer-barrier and off-shore). Reefs within the 20 study locations reported here did not cover this spatial extent and could only be classified within the reef as either: reef crest, reef slope, patch reef or reef flat.
The pattern of clownfish cohabitation observed within this study was best explained by the ratio of host anemone species to clownfish species; when the number of clownfish species exceeded the number of host anemone species, cohabitation was almost always documented (table 1). In the Coral Triangle, for example, the number of clownfish species (13 species) exceeded the number of species of host anemones (10 species). All available hosts were occupied, and host limitation combined with high clownfish diversity appears to drive cohabitation. Survival of clownfish outside of a host anemone is inhibited by predation [22]. Consequently, cohabitation reduces the risk of mortality and enables greater species richness in environments where microhabitats are limited.
To date, the scientific consensus is that clownfish competitively exclude other species from the host anemone [27,40]. However, in the Hoga reefs, clownfishes coexist using a size-structured hierarchy previously documented only in intraspecific groups [41]. Importantly, these cohabiting species were often phylogenetically unrelated and morphologically different, further highlighting the distinctness of the cohabiting relationships. Both generalist and specialist clownfish species were seen to cohabit [21].
Fine-scale spatial separation of the host anemone is a mechanism that allows both cohabiting species to share a common habitat, with outcomes of competitive interactions dictating which part of the anemone is inhabited by each species. Subordinate individuals tend to occupy the peripheral areas of the anemone [42], as occurs in intraspecific groups where competition forces subordinates to occupy suboptimal and less favourable environments [38–40]. Dominant species from interspecific social groups generally did not alter their spatial distribution within the host anemone, although the common dominant species A. clarkii increased the time it spent outside of the host anemone. The increased time outside of the host anemone created greater spatial separation from the subordinate species, potentially alleviating competition for space inside the host anemone. Within the host anemone, space use per fish decreased by ca. 50% in interspecific social groups compared to intraspecific social groups. Despite the greater risk of predation for A. clarkii while outside of the host anemone, it appears that the reduced competition created by being outside may outweigh any additional associated risks.
Aggression did not increase in interspecific social groups despite the reduced amount of available habitat per fish. Thus, the acceptance of a subordinate congeneric by a dominant species is a mechanism that allows multiple species to cohabit a single habitat. Subordinates probably avoid aggressive interactions with the dominant species owing to fear of injury, mortality or eviction from the anemone (e.g. [43]). Living on the periphery of the host anemone, despite the higher risk of predation [44,45], is a better option than having no host anemone. Occupying a part of the anemone also provides an opportunity for social queuing, eventually allowing the subordinate to become dominant and reproduce [43]. For the dominant species, there may be a benefit from allowing another species to cohabit (e.g. enhanced growth and protection of anemone [45,46]). Furthermore, it may be more beneficial for dominant individuals to accept a smaller congeneric rather than a conspecific because a congeneric is less likely to have niche overlap (e.g. different diets) or pose a risk to stealing a mate. Given that clownfish produce sounds during agonistic interactions that are species- and size-specific [47,48], it is possible that this form of communication is used to convey information on status and facilitate acceptance, thereby helping to stabilize the cohabiting group.
Importantly, no species was observed always to be the subordinate when cohabiting. Consequently, although clownfishes have competitive hierarchies [40], other factors (e.g. colonization ability [41,43–45]) may influence which species attains the highest rank and therefore dominance within the cohabitating social group. Demographic differences will help promote evolutionary stability of cohabitation because it allows each species to become dominant in some instances, and thus become the reproductively active individual and species within each social group. If hierarchies were fixed, the cohabiting subordinate species would struggle to persist in the region as they would face prolonged suppression of reproduction. Whether both species are able to breed while cohabiting has yet to be confirmed, but egg clutches were only observed during this study for the dominant clownfish species of each social group. Cohabitation may support stable populations even within subordinate species through social queuing whereby the subordinate species waits their turn to become the dominant individual, a process well documented within intraspecific groups [43]. The increased levels of interaction and reduced spatial separation of species during cohabitation may also explain historical hybridization events [49].
Cohabitation of species has been well documented in rainforests (e.g. scorpions [50]), where there is high species diversity and often habitat limitation analogous to coral reefs [51]. The presence of cohabitation within both coral reefs and rainforests suggests that it is an important mechanism facilitating diversity in specialists/commensal species where habitat resources are limiting. Within this study, cohabitation was not limited to one species pair in one host species, but rather we observed a range of different clownfishes cohabiting on several different host species. In other groups, shrimps that are commensal with anemones and mushroom corals (e.g. Heliofungia actiniformis) also achieve high species richness in the Coral Triangle through cohabitation [14,52]. Thus, the total diversity of commensals on microhabitats such as anemones can be very high because there is cohabitation within and between numerous taxonomic groups (e.g. fish, shrimps and crabs). The reliance of several species on a specific microhabitat within high diversity areas increases the value of the host habitat. Thus, the consequences of microhabitat loss are likely to exaggerate impacts on species richness in high biodiversity environments.
Overall, this study tested several hypotheses to determine what drives cohabitation and demonstrated that this was best explained by the ratio of host anemones to clownfish species. At sites in the highly diverse Coral Triangle, various combinations of clownfishes can co-inhabit a range of host anemones without increased aggression. Cohabitation is facilitated by niche partitioning over a fine spatial scale and through the acceptance of a subordinate species by a dominant species. Given that multiple marine species (e.g. fishes, crustaceans) cohabit anemones (and other microhabitats) within the Coral Triangle, protecting these microhabitats and preserving cohabiting relationships are important to conserving this global centre of marine biodiversity.
Supplementary Material
Supplementary Material
{kind=link}
Acknowledgments
We are extremely grateful to three anonymous reviewers for their insightful comments that helped to improve upon an earlier version of the manuscript. For the work on Hoga Island, the authors are grateful for support from Operation Wallacea, The Wallacea Biodiversity Institute, Balai Taman Nasional Wakatobi (Wakatobi National Park), The Wakatobi Government, Ministry for Research, Technology, and Higher Education, Indonesia (RISTEK-DIKTI). We also thank the following for logistical support at our other study locations: F. Benzoni, University of Milano-Bicocca MaRHE Centre, Lord Howe Island Marine Park, Parks Australia, Capricorn Star, Wet and Dry Adventures, Howea Divers, Kosrae Village, University of Guam Marine Lab, D. Hess (College of the Marshall Islands), Marshall Islands Marine Resources Authority, University of Western Australia and P. Wruck. For field assistance we thank all Operation Wallacea volunteers, as well as: M. Beger, J. DiBattista, A. Frisch, J. Gilligan, S. Gudge, I. Kerr, C. McDonald, J. Neilson, M. Priest, Z. Richards, L. Rochford, T. Sinclair-Taylor, B. Taylor and M. van der Meer.
Ethics
All research adhered to ethical and local standards of research within Indonesia and was completed under a research permit issued to Smith.
Data accessibility
Within the manuscript or electronic supplementary materials.
Authors' contributions
E.F.C., M.D.B., J.-P.A.H., A.J.D. and D.J.S. designed the study. E.F.C. and M.D.B. collected the Hoga data and J.-P.A.H. collected the data at all other sites. E.F.C., M.D.B. and J.-P.A.H. carried out the data analysis; A.J.D., E.F.C. and J.-P.A.H. contributed to the statistical analysis; E.F.C., D.J.S., A.J.D. and J.-P.A.H. led the writing of the manuscript with all authors contributing and approving the final draft.
Competing interests
We have no competing interests.
Funding
For the work on Hoga Island, the authors are grateful for financial and logistical support from Operation Wallacea and The Wallacea Biodiversity Institute.
References
- 1.Edwards KF, Stachowicz JJ. 2011. Spatially stochastic settlement and the coexistence of benthic marine animals. Ecology 92, 1094–1103. ( 10.1890/10-1332.1) [DOI] [PubMed] [Google Scholar]
- 2.Levine JM, Hille Ris Lambers J. 2009. The importance of niches for the maintenance of species diversity. Nature 461, 254–258. ( 10.1038/nature08251) [DOI] [PubMed] [Google Scholar]
- 3.Liem KF. 1980. Adaptive significance of intra- and interspecific differences in the feeding repertoires of cichlid fishes. Am. Zool. 20, 295–314. ( 10.1093/icb/20.1.295) [DOI] [Google Scholar]
- 4.Bergmüller R, Taborsky M. 2010. Animal personality due to social niche specialisation. Trends Ecol. Evol. 1267, 1–8. ( 10.1016/j.tree.2010.06.012) [DOI] [PubMed] [Google Scholar]
- 5.Griffin JN, De La Haye KL, Hawkins SJ, Thompson RC, Jenkins SR. 2008. Predator diversity and ecosystem functioning: density modifies the effect of resource partitioning. Ecology 82, 298–305. ( 10.1890/07-1220.1) [DOI] [PubMed] [Google Scholar]
- 6.MacArthur RH. 1972. Geographical ecology. New York, NY: Harper & Row. [Google Scholar]
- 7.Chase JM, Leibold MA. 2003. Ecological niches. Chicago, IL: Chicago Press. [Google Scholar]
- 8.Hubbell SP. 2006. Neutral theory and the evolution of ecological equivalence. Ecology 87, 1387–1398. ( 10.1890/0012-9658) [DOI] [PubMed] [Google Scholar]
- 9.Costanza R. 1999. The ecological, economic, and social importance of the oceans. Ecol. Econ. 31, 199–213. ( 10.1016/S0921-8009(99)00079-8) [DOI] [Google Scholar]
- 10.Schoener TW. 1974. Resource partitioning in ecological communities. Science 185, 27–39. ( 10.1126/science.185.4145.27) [DOI] [PubMed] [Google Scholar]
- 11.Munday PL, Jones GP, Caley MJ. 1997. Habitat specialisation and the distribution and abundance of coral-dwelling gobies. Mar. Ecol. Prog. Ser. 152, 227–239. ( 10.3354/meps152227) [DOI] [Google Scholar]
- 12.Sale PF. 1978. Coexistence of coral reef fishes—a lottery for living space. Environ. Biol. Fishes 3, 85–102. ( 10.1007/BF00006310) [DOI] [Google Scholar]
- 13.Munday PL. 2004. Competitive coexistence of coral-dwelling fishes: the lottery hypothesis revisited. Ecology 85, 623–628. ( 10.1890/03-3100) [DOI] [Google Scholar]
- 14.Robertson DR, Polunin NVC. 1981. Coexistence: symbiotic sharing of feeding areas and algal food by some coral reef fishes from the Western Indian Ocean. Mar. Biol. 62, 185–195. ( 10.1007/BF00388182) [DOI] [Google Scholar]
- 15.Hoeksema BW. 2007. Delineation of the Indo-Malayan Centre of Maximum Marine Biodiversity: The Coral Triangle. In Biogeography, time and place: distributions, barriers and islands (ed. Renema W.), pp. 117–178. Dordrecht, The Netherlands: Springer. [Google Scholar]
- 16.Allen GR, Werner TB. 2002. Coral reef fish assessment in the ‘Coral Triangle’ of southeastern Asia. Environ. Biol. Fishes. 65, 209–214. ( 10.1023/A:1020093012502) [DOI] [Google Scholar]
- 17.Allen GR. 2008. Conservation hotspots of biodiversity and endemism for Indo-Pacific coral reef fishes. Aquat. Conserv. 18, 541–556. ( 10.1002/aqc.880) [DOI] [Google Scholar]
- 18.Rocha LA, Robertson DR, Roman JR, Bowen BW. 2005. Ecological speciation in tropical reef fishes. Proc. R. Soc. B 272, 573–579. ( 10.1098/2004.3005) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Robertson DR. 1984. Cohabitation of competing territorial damselfishes on a Caribbean coral reef. Ecology 65, 1121–1135. ( 10.2307/1938320) [DOI] [Google Scholar]
- 20.Bos AR, Hoeksema BW. 2015. Cryptobenthic fishes and co-inhabiting shrimps associated with the mushroom coral Heliofungia actiniformis (Fungiidae) in the Davao Gulf, Philippines. Environ. Biol. Fishes 98, 1479–1489. ( 10.1007/s10641-014-0374-0) [DOI] [Google Scholar]
- 21.Fautin DG, Allen GR. 1997. Anemonefishes and their host sea anemones. Revised edition. Perth: Western Australian Museum. [Google Scholar]
- 22.Fautin DG. 1986. Why do anemonefishes inhabit only some host actinians? Environ. Biol. Fish 15, 171–180. ( 10.1007/BF00002992) [DOI] [Google Scholar]
- 23.Dunn DF. 1981. The clownfish sea anemones: Stichodactylidae (Coelenterata: Actiniaria) and other sea anemones symbiotic with pomacentrid fishes. Trans. Amer. Philos. Soc. 71, 1–115. ( 10.2307/1006382) [DOI] [Google Scholar]
- 24.Mariscal RN. 1972. Behaviour of symbiotic fishes and sea anemones. In Behaviour of marine animals, vol. 2, 1st edn (eds Winn HE, Olla BL), pp. 327–361. New York, NY: Plenum Press. [Google Scholar]
- 25.Fautin DG. 1985. Competition by anemone fishes for host actinians. Proc. Fifth Int. Coral Reef Congress, Tahiti. 5, 373–377. [Google Scholar]
- 26.Riccardi F, Boyer M, Ollerton J. 2010. Assemblage and interaction structure of the anemonefish–anemone mutualism across the Manado region Sulawesi, Indoensia. Environ. Biol. Fish. 4, 333–347. ( 10.1007/s10641-010-9606-0) [DOI] [Google Scholar]
- 27.Elliott JK, Mariscal RN. 2001. Coexistence of nine anemonefish species: differential host and habitat utilisation, size and recruitment. Mar. Biol. 138, 23–36. ( 10.1007/s002270000441) [DOI] [Google Scholar]
- 28.Clifton J, Unsworth RFK, Smith DJ. 2010. Marine research and conservation in the Coral Triangle: the Wakatobi National Park. New York, NY: Nova Science Publishers, Inc. [Google Scholar]
- 29.Naumann MS, Niggl W, Laforsch C, Glaser C, Wild C. 2008. Coral surface area quantification—evaluation of established techniques by comparison with computer tomography. Coral Reefs 28, 109–117. ( 10.1007/s00338-008-0459-3) [DOI] [Google Scholar]
- 30.Hattori A. 2002. Small and large anemonefishes can coexist using the same patchy resources on a coral reef, before habitat destruction. J. Anim. Ecol. 71, 824–831. ( 10.1046/j.1365-2656.2002.00649.x) [DOI] [Google Scholar]
- 31.Brolund TH, Tychseno A, Nielsen LE, Arvedlund M. 2004. An assemblage of the host anemone Heteractis magnifica in the northern Red Sea, and the distribution of the resident anemonefish. J. Mar. Biol. Assoc. UK 84, 671–674. ( 10.1017/s0025315404009737h) [DOI] [Google Scholar]
- 32.Goodwin J, Fautin DG. 1992. Defence of host actinians by anemonefishes. Copeia 3, 902–908. ( 10.2307/1446171) [DOI] [Google Scholar]
- 33.Buston PM. 2003. Forcible eviction and prevention of recruitment in the clown anemonefish. Behav. Ecol. 14, 576–582. ( 10.1093/beheco/arg036) [DOI] [Google Scholar]
- 34.Litsios G, Pearman PB, Lanterbecq D, Tolou N, Salamin N. 2014. The radiation of the clownfishes has two geographical replicates. J. Biogeogr. 41, 2140–2149. ( 10.1111/jbi.12370) [DOI] [Google Scholar]
- 35.Kuo SR, Shao KT. 1991. Feeding habits of damselfishes pomacentridae from the southern part of Taiwan. J. Fish. Soc. Taiwan. 18, 165–176. [Google Scholar]
- 36.Chen KC, Mok HK. 1988. Sound production in the anemonefishes, Amphiprion clarkii and A. frenatus (Pomacentridae), in captivity. Jpn. J. Ichthyol. 35, 90–97. ( 10.1007/BF02906690) [DOI] [Google Scholar]
- 37.R Development Core Team. 2011. R: a language and environment for statistical computing. See http://www.R-project.org.
- 38.Hobbs JPA, Frisch AJ, Ford BM, Thums M, Saenz-Agudelo P, Furby KA, Berumen ML. 2013. Taxonomic, spatial and temporal patterns of bleaching in anemones inhabited by anemonefishes. PLoS ONE 8, e70966 ( 10.1371/journal.pone.0070966) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Elliot JK, Elliot JM, Mariscal RN. 1995. Host selection, location, and association behaviours of anemonefishes in field settlement experiments. Mar Biol. 122, 377–389. ( 10.1007/BF00350870) [DOI] [Google Scholar]
- 40.Hattori A. 1991. Socially controlled growth and size-dependent sex change in the anemonefish Amphiprion frenatus in Okina-wa, Japan. Jpn. J. Ichthyol. 38, 165–177. ( 10.1007/BF02905541) [DOI] [Google Scholar]
- 41.Holbrook SJ, Schmitt RJ. 2002. Competition for shelter space causes density-dependent predation mortality in damselfishes. Ecology 83, 2855–2868. ( 10.2307/3072021) [DOI] [Google Scholar]
- 42.De Brauwer M, Camp E, Jompa J, Smith DJ. 2015. High levels of heterospecific cohabitation among anemonefishes in Hoga Island, Indonesia. Mar. Biodivers. 1–2. ( 10.1007/s12526-015-0343-7) [DOI] [Google Scholar]
- 43.Wong MY, Buston PM, Munday PL, Jones GP. 2007. The threat of punishment enforces peaceful cooperation and stabilizes queues in a coral-reef fish. Proc. R. Soc. B 274, 1093–1099. ( 10.1098/rspb.2006.0284) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Schmitt RJ, Holbrook SJ. 2003. Mutualism can mediate competition and promote coexistence. Ecol. Lett. 6, 898–902. ( 10.1046/j.1461-0248.2003.00514.x) [DOI] [Google Scholar]
- 45.Holbrook SJ, Schmitt RJ. 2004. Population dynamics of a damselfish: effects of a competitor that also is an indirect mutualist. Ecology 85, 979–985. ( 10.1890/03-0406) [DOI] [Google Scholar]
- 46.Hattorri A. 1995. Coexistence of two anemonefishes, Amphiprion clarkii and A. perideraion, which utilise the same host sea anemone. Environ. Biol. Fishes. 42, 345–353. ( 10.1007/BF00001464) [DOI] [Google Scholar]
- 47.Colleye O, Parmentier E. 2012. Overview on the diversity of sounds produced by clownfishes (Pomacentridae): importance of acoustic signals in their peculiar way of life. PLoS ONE 7, e49179 ( 10.1371/journal.pone.0049179) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Colleye O, Vandewalle P, Lanterbecq D, Lecchini D, Parmentier E. 2011. Interspecific variation of calls in clownfishes: degree of similarity in closely related species. BMC Evol. Biol. 11, 365 ( 10.1186/1471-2148-11-365) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Litsios G, Salamin N. 2014. Hybridisation and diversification in the adaptive radiation of clownfishes. BMC. Evol. Biol. 14, 245 ( 10.1186/s12862-014-0245-5) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lira AFA, Souza AM, Silva Filho AAC, Albuquerque CMR. 2013. Spatio-temporal microhabitat use by two co-occurring species of scorpions in Atlantic rainforest in Brazil. J. Zool. 116, 182–185. ( 10.1016/j.zool.2013.01.002) [DOI] [PubMed] [Google Scholar]
- 51.Connel JH. 1978. Diversity in tropical rain forests and coral reefs. Science 4335, 1302–1310. ( 10.1126/science.199.4335.1302) [DOI] [PubMed] [Google Scholar]
- 52.Hoeksema BW, Fransen CHJM. 2011. Space partitioning by symbiotic shrimp species cohabitating in the mushroom coral Heliofungia actiniformis at Semporna, eastern Sabah. Coral Reefs 30, 519 ( 10.1007/s00338-011-0736-4) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Within the manuscript or electronic supplementary materials.