

Abstract
The skeleton is an exquisitely sensitive and archetypal T3-target tissue that demonstrates the critical role for thyroid hormones during development, linear growth, and adult bone turnover and maintenance. Thyrotoxicosis is an established cause of secondary osteoporosis, and abnormal thyroid hormone signaling has recently been identified as a novel risk factor for osteoarthritis. Skeletal phenotypes in genetically modified mice have faithfully reproduced genetic disorders in humans, revealing the complex physiological relationship between centrally regulated thyroid status and the peripheral actions of thyroid hormones. Studies in mutant mice also established the paradigm that T3 exerts anabolic actions during growth and catabolic effects on adult bone. Thus, the skeleton represents an ideal physiological system in which to characterize thyroid hormone transport, metabolism, and action during development and adulthood and in response to injury. Future analysis of T3 action in individual skeletal cell lineages will provide new insights into cell-specific molecular mechanisms and may ultimately identify novel therapeutic targets for chronic degenerative diseases such as osteoporosis and osteoarthritis. This review provides a comprehensive analysis of the current state of the art.
Introduction
-
Thyroid Hormone Physiology
Hypothalamic-pituitary-thyroid axis
TSH action
Extrathyroidal actions of TSH
Thyroid hormone transport
Thyroid hormone metabolism
Nuclear actions of thyroid hormones
Nongenomic actions of thyroid hormones
-
Skeletal Physiology
Bone and cartilage cell lineages
Intramembranous ossification
Endochondral ossification
Linear growth and bone maturation
The bone-remodeling cycle
-
Skeletal Target Cells and Downstream Signaling Pathways
TSH actions in chondrocytes, osteoblasts and osteoclasts
T3 actions in chondrocytes, osteoblasts, and osteoclasts
-
Genetically Modified Mice
Targeting TSHR signaling
Targeting thyroid hormone transport and metabolism
Targeting TRα
Targeting TRβ
-
Skeletal Consequences of Mutations in Thyroid Signaling Genes in Humans
TSHB
TSHR
SBP2
THRB
THRA
-
Thyroid Status and Skeletal Development
Consequences of hypothyroidism
Consequences of thyrotoxicosis
-
Thyroid Status and Bone Maintenance
Consequences of variation of thyroid status within the reference range
Consequences of hypothyroidism
Consequences of subclinical hypothyroidism
Consequences of subclinical hyperthyroidism
Consequences of hyperthyroidism
-
Osteoporosis and Genetic Variation in Thyroid Signaling
Associations with BMD
-
Osteoarthritis and Genetic Variation in Thyroid Signaling
Genetics
Mechanism
Summary and Future Directions
I. Introduction
The essential requirement for thyroid hormones during linear growth and skeletal maturation is well established and has been recognized for 125 years. Indeed, the association between goiter, cretinism, developmental retardation, and short stature had been known for centuries, and the therapeutic use of burnt sponge and seaweed in the treatment of goiter dates back to 1600 BC in China. Paracelcus provided the first clinical description of endemic goiter and congenital idiocy in 1603. Between 1811 and 1813, Bernard Courtois discovered iodine, Joseph Gay-Lussac identified it as an element, and Humphrey Davy recognized it as a halogen (1). However, in 1820 Jean-Francois Coindet was the first to use iodine as a treatment for goiter, and in the 1850s, Gaspard Chatin was the first to show that iodine in plants prevented cretinism and goiter in endemic regions. Thomas Curling described cretinism in association with athyreosis in 1850, whereas William Gull provided the causal link between the lack of a thyroid gland and cretinism in 1873. William Ord extended Gull's observations and chaired the first detailed report on hypothyroidism by the Clinical Society of London in 1878, linking cretinism, myxedema, and cachexia strumipriva (decay due to lack of goiter) as a single entity. Indeed, in a lecture to the German Society of Surgery in 1883, the Swiss Nobel Laureate Theodor Kocher described cachexia strumipriva as a specific disease that included “decreased growth in height” after removal of the thyroid gland. Ultimately, these events led to the first organotherapy for hypothyroidism by George Murray in 1891, although the ancient Chinese had used animal thyroid tissue as a treatment for goiter as early as 643 AD (1–3).
Alongside the emergence of hypothyroidism as a recognized disease, Charles de Saint-Yves, Antonio Testa, and Guiseppe Flajani reported the first cases of goiter, palpitations, and exophthalmos between 1722 and 1802, although these features were not linked at that time. Caleb Parry had recognized in 1825, whereas Robert Graves independently recognized and also published in 1835, the link between hypertrophic goiter and exophthalmos (1, 3). Carl Adolf von Basedow extended Graves' description in 1840 by adding palpitations, weight loss, diarrhea, tremor, restlessness, perspiration, amenorrhea, myxedema of the lower leg, and orbital tissue hypertrophy to describe the syndrome more completely. In 1886, Paul Möbius proposed that the cause of these symptoms was increased thyroid function, and Murray supported this view in 1891 at the time of his organotherapy for hypothyroidism (1, 3). Coincidentally, also in 1891, Friedrich Von Recklinghausen reported a patient with thyrotoxicosis and multiple fractures and was the first to identify the relationship between the thyroid and the adult skeleton (4, 5). Since then, a role for thyroid hormones in bone and mineral metabolism has become well established.
During the last 25 years, the role of thyroid hormones in bone and cartilage biology has attracted considerable and growing attention, leading to important advances in understanding the consequences of thyroid disease on the developing and adult skeleton. Major progress in defining the mechanisms of thyroid hormone action in bone has followed and has led to new insights into thyroid-related skeletal disorders. As a result, the role of the hypothalamic-pituitary-thyroid (HPT) axis in skeletal pathophysiology has become a high-profile subject. It is only now that experimental tools are becoming available to allow determination of the precise cellular and molecular mechanisms that underlie thyroid hormone actions in the skeleton. This review will discuss our current understanding by considering the published literature up to December 31, 2015.
II. Thyroid Hormone Physiology
A. Hypothalamic-pituitary-thyroid axis
Synthesis and release of the prohormone 3,5,3′,5′-L-tetraiodothyronine (thyroxine [T4]) and the active thyroid hormone 3,5,3′-L-triiodothyronine (T3) are controlled by a negative feedback loop mediated by the HPT axis (Figure 1) (6). Thyrotropin-releasing hormone (TRH) is secreted by the hypothalamic paraventricular nucleus and acts on pituitary thyrotrophs to stimulate release of thyrotropin (thyroid-stimulating hormone [TSH]). TSH subsequently acts via the TSH receptor (TSHR) on thyroid follicular cells to stimulate cell proliferation and the synthesis and secretion of T4 and T3 (7). T3, derived predominantly from local metabolism of T4, acts via thyroid hormone receptors (TR) α and β (TRα, TRβ) in the hypothalamus and pituitary to inhibit synthesis and secretion of TRH and TSH (8–11). Normal euthyroid status is maintained by a negative feedback loop that establishes a physiological inverse relationship between TSH and circulating T3 and T4, thus defining the HPT axis set point (12, 13). Systemic thyroid hormone and TSH concentrations vary significantly among individuals, indicating that each person has a unique set point (12). Twin studies indicate the set point is genetically determined with heritability for free T3 (fT3), free T4 (fT4), and TSH of 65% (14), and candidate gene and genome-wide association studies (GWAS) have identified quantitative trait loci (15–17).
Figure 1.
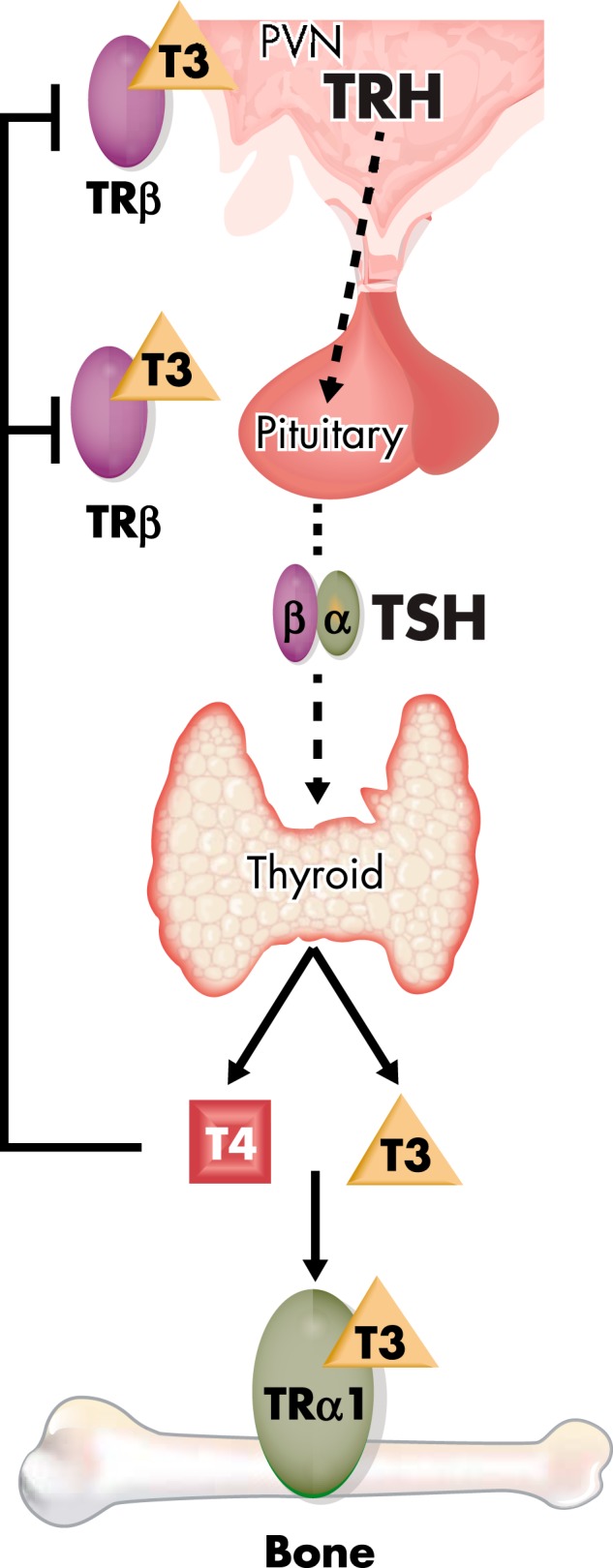
HPT axis. The thyroid gland secretes the prohormone T4 and the active hormone T3, and circulating concentrations are regulated by a classical endocrine negative feedback loop that maintains an inverse physiological relationship between TSH, and T4 and T3. PVN, paraventricular nucleus.
B. TSH action
The glycoprotein hormone TSH is composed of common α-subunits and unique β-subunits. The TSHR is a G protein-coupled receptor consisting of ligand binding, ecto- and transmembrane domains (Figure 2) (18). Although cAMP is the major second messenger following activation of the TSHR in thyroid follicular cells, alternative downstream signaling pathways have been implicated in both thyroid and extrathyroidal tissues (19–22). In the thyroid, the TSHR associates with various G proteins (23), and Gαs and Gαq are thought to compete for activation by the TSHR (20, 24).
Figure 2.

TSH action. Binding of TSH to the TSHR results in activation of G protein-coupled downstream signaling including: 1) the adenylyl cyclase (AC), cAMP, protein kinase A (PKA), and the cAMP response element binding (CREB) protein; or 2) phospholipase C (PLC), inositol triphosphate (IP3), and intracellular calcium pathway; or 3) the PLC, diacylglycerol (DAG), protein kinase C (PKC), and signal transducer and activator of transcription 3 (STAT3) pathway. PIP2, phosphatidylinositol 4,5-bisphosphate.
C. Extrathyroidal actions of TSH
The TSHR has been proposed to have diverse functions in extrathyroidal tissues, although their physiological importance has not been established. Thus, TSHR expression has been reported in anterior pituitary, brain, pars tuberalis, bone, orbital preadipocytes and fibroblasts, kidney, ovary and testis, skin and hair follicles, heart, adipose tissue, as well as hematopoietic and immune cells (19, 25–28). These data suggest direct actions of TSH, for example, in the regulation of seasonal reproduction (29–31), bone turnover (32), pathogenesis of Graves' orbitopathy (33–35), and immunomodulatory responses in the bone marrow (36–43), gut (42, 43), and skeleton (38).
D. Thyroid hormone transport
Uptake of thyroid hormones into peripheral tissues and entry into target cells is mediated by specific membrane transporter proteins (Figure 3) (44), including monocarboxylate transporter (MCT) 8 and MCT10, the organic anion transporter protein-1c1 (OATP1c1), and the nonspecific L-type amino acid transporters 1 and 2 (LAT1, LAT2) (45). The best-characterized specific transporter MCT8 is expressed widely, and its physiological importance has been demonstrated by inactivating mutations of MCT8 that cause the Allan-Herndon-Dudley X-linked psychomotor retardation syndrome (OMIM no. 300523) (46, 47).
Figure 3.

Thyroid hormone action in bone cells. A, In hypothyroidism, despite maximum DIO2 (D2) and minimum DIO3 (D3) activities, TRα1 remains unliganded and bound to corepressor, thus inhibiting T3-target gene transcription. B, In the euthyroid state, D2 and D3 activities are regulated to optimize ideal intracellular T3 availability, resulting in displacement of corepressor and physiological transcriptional activity of TRα1. C, In thyrotoxicosis, despite maximum D3 and minimum D2 activities, supraphysiological intracellular T3 concentrations result in increased TRα1 activation and enhanced T3-target gene responses.
E. Thyroid hormone metabolism
T4 is derived from thyroid gland secretion, whereas most circulating T3 is generated by deiodination of T4 in peripheral tissues. Although the circulating fT4 concentration is 4-fold greater than fT3, the TR-binding affinity for T3 is 15-fold higher than its affinity for T4 (48). Thus, T4 must be converted to T3 for mediation of genomic thyroid hormone action (Figure 3) (49). Three iodothyronine deiodinases metabolize thyroid hormones to active or inactive products (6, 50, 51). The type 1 deiodinase (DIO1) is inefficient, with an apparent Michaelis constant (Km) of 10−6 to 10−7 m, and catalyzes removal of inner or outer ring iodine atoms in equimolar proportions to generate T3, rT3, or 3,3′-diiodothyronine, depending on the substrate. Most of the circulating T3 is derived from conversion of T4 to T3 by DIO1, which is expressed mainly in the thyroid gland, liver, and kidney. Nevertheless, its physiological role remains uncertain because serum T3 concentrations are normal in Dio1−/− knockout mice (52). Activity of DIO2 in skeletal muscle may also contribute to circulating T3, although this role probably differs between species (6, 49, 53–55). DIO2 (Km, 10−9 m) is more efficient than DIO1 and catalyzes outer ring deiodination to generate T3 from T4. The physiological role of DIO2 is thus to control the intracellular T3 concentration and saturation of the nuclear TR in target tissues (56–58). Importantly, DIO2 protects tissues from the detrimental effects of hypothyroidism because its low Km permits efficient local conversion of T4 to T3. T4 treatment of cells in which MCT8 and DIO2 are coexpressed results in increased T3-target gene expression (59), indicating that thyroid hormone uptake and metabolism coordinately regulate T3 responsiveness. By contrast, DIO3 (Km, 10−9 m) irreversibly inactivates T3 or prevents T4 being activated by inner ring deiodination to generate 3,3′-diiodothyronine or rT3, respectively. The physiological role of DIO3 is thus to prevent or limit access of thyroid hormones to specific tissues at critical times during development and in tissue repair (6, 49, 51).
Consistent with this, DIO2 and DIO3 are expressed in T3-target cells, including the central nervous system, cochlea, retina, heart, and skeleton (49, 60–65), and expression of both enzymes is regulated in a temporo-spatial and tissue-specific manner (51, 66, 67). Acting together, DIO2 and DIO3 thus control cellular T3 availability (49). For example, during fetal growth, high levels of DIO3 in placenta, uterus, and fetal tissues protect developing organs from exposure to inappropriate levels of T3 and facilitate cell proliferation (68). At birth, DIO3 declines rapidly whereas expression of DIO2 increases to trigger cell differentiation and tissue maturation during postnatal development (49–51). The temporo-spatial and tissue-specific regulated expression of both DIO2 and DIO3 (66) and the TRα and TRβ nuclear receptors (69) combine to provide a complex and coordinated system for fine control of T3 availability and action in individual cell types.
F. Nuclear actions of thyroid hormones
TRα and TRβ are members of the nuclear receptor superfamily (70, 71), acting as ligand-inducible transcription factors that regulate expression of T3-target genes (Figure 3). In mammals, THRA encodes three C-terminal variants of TRα. TRα1 is a functional receptor that binds both DNA and T3, whereas TRα2 and TRα3 fail to bind T3 and act as antagonists in vitro (72). A promoter within intron 7 of mouse Thra gives rise to two truncated variants, TRΔα1 and TRΔα2, which are potent dominant-negative antagonists in vitro, although their physiological role is unclear (73). Two truncated TRα1 proteins, p28 and p43, arise from alternate start codon usage and are proposed to mediate T3 actions in mitochondria or nongenomic responses (74, 75). THRB encodes two N-terminal TRβ variants, TRβ1 and TRβ2, both of which act as functional receptors. Two further transcripts, TRβ3 and TRΔβ3, have been described, but their physiological role is uncertain (76, 77). TRα1 and TRβ1 are expressed widely, but their relative concentrations differ during development and in adulthood due to tissue-specific and temporo-spatial regulation (69), so that most T3-target tissues are either predominantly TRα1 or TRβ1 responsive or lack isoform specificity. Expression of TRβ2, however, is markedly restricted. In the hypothalamus and pituitary, it mediates inhibitory actions of thyroid hormones on TRH and TSH expression to control the HPT axis (8, 78), whereas in cochlea and retina, TRβ2 is an important regulator of sensory development (79, 80).
In the nucleus, TRs form heterodimers with retinoid X receptors (RXRs) and bind T3 response elements (TREs) in target gene promoters to regulate transcription. Unliganded TRs compete with T3-bound TRs for DNA response elements. They are potent transcriptional repressors and have critical roles during development (81–84). Unliganded TRs interact with corepressor proteins, including nuclear receptor corepressor and the silencing mediator for retinoid and TR, which recruit histone deacetylases and inhibit gene transcription (85, 86). Ligand-bound TRs interact with steroid receptor coactivator 1 and other related coactivators in a hormone-dependent fashion leading to target gene activation. The opposing chromatin-modifying effects of unliganded and liganded TRs greatly enhance the magnitude of the transcriptional response to T3 (87–89). In addition to positive stimulatory effects, T3 also mediates transcriptional repression to inhibit expression of key target genes, including TSH. Although negative regulatory effects are physiologically critical, underlying molecular mechanisms have not been fully characterized (88).
G. Nongenomic actions of thyroid hormones
Nongenomic effects of thyroid hormones include actions that do not directly influence nuclear gene expression. Nongenomic actions frequently have a short latency, are not affected by inhibitors of transcription and translation, and have agonist and antagonist affinity and kinetics divergent from classical nuclear hormone actions (90). These rapid responses are associated with second messenger pathways including: 1) the phospholipase C, inositol triphosphate, diacylglycerol, protein kinase C, and intracellular Ca2+ signaling pathway; 2) the adenylyl cyclase, protein kinase A, and cAMP-response element binding protein pathway; and 3) the Ras/Raf1 serine-threonine kinase/MAPK pathway.
Nongenomic actions of thyroid hormones have been described at the plasma membrane, in the cytoplasm, and in mitochondria (74, 88). The αVβ3 integrin has been reported to mediate cell surface responses to T4 acting, for example, via the MAPK pathway to stimulate cell proliferation and angiogenesis (91, 92). TRβ also mediates rapid responses to T3, acting via the PI3K/AKT/mTOR/p70S6K and PI3K pathways (93–99), whereas palmitoylated TRα activates the nitric oxide/protein kinase G2/Src pathway to stimulate MAPK and PI3K/AKT downstream signaling responses that mediate rapid T3 actions in osteoblastic cells (75).
III. Skeletal Physiology
Bones of the skull vault form directly from mesenchyme via intramembranous ossification, whereas long bones develop on a cartilage scaffold by endochondral ossification (Figure 4). Four key cell types are involved in these developmental programs, and they are essential for linear growth in the postnatal period and maintenance of the skeleton in later life.
Figure 4.

Intramembranous and endochondral ossification. A, Postnatal day 1 skull vault stained with alizarin red (bone) and alcian blue (cartilage) showing sutures and fontanelles. B, Schematic representation of intramembranous bone formation at a skull suture. C, Proximal tibial section at P21 growth plate stained with alcian blue (cartilage) and Van Gieson (bone matrix, red). D, Schematic representation of the growth plate.
A. Bone and cartilage cell lineages
1. Chondrocytes
Chondrocytes are the first skeletal cell type to arise during development (100). In early embryogenesis, mesenchyme precursors condense and define a template for the future skeleton. These cells differentiate into chondrocytes that proliferate and secrete a matrix containing aggrecan, elastin, and type II collagen to form a cartilage anlage or a model of the skeletal element. Cells at the center of the anlage stop proliferating and differentiate into prehypertrophic and then hypertrophic chondrocytes (101). Hypertrophic chondrocytes increase rapidly in size, synthesize a matrix rich in type X collagen, and induce formation of calcified cartilage before finally undergoing apoptosis (102). Initiation of chondrogenesis requires bone morphogenic protein (BMP) signaling and the transcription factor SOX9 acting in association with SOX5 and SOX6. Indian hedgehog (IHH) stimulates chondrocyte proliferation directly but also ensures that sufficient chondrocyte proliferation occurs by increasing PTHrP signaling, which inhibits chondrocyte hypertrophic differentiation. Canonical Wnt signaling promotes hypertrophic differentiation via inhibition of SOX9, whereas fibroblast growth factor (FGF) 18 inhibits both proliferation and differentiation of chondrocytes (102, 103).
2. Osteoblasts
Bone-forming osteoblasts comprise 5% of bone cells and derive from multipotent mesenchymal stem cells that can differentiate into chondrocytes, osteoblasts, or adipocytes. Osteoblast maturation comprises precursor cell commitment, cell proliferation, type I collagen deposition, and matrix mineralization. After bone formation, osteoblasts may differentiate into bone-lining cells or osteocytes or may undergo apoptosis (104, 105). In SOX9-expressing mesenchymal progenitors, osteoblastogenesis requires induction of two critical transcription factors, RUNX2 and Osterix (105, 106). Subsequent differentiation is regulated by the IHH, PTH, Notch, canonical Wnt, BMP, IGF-1, and FGF signaling pathways (105, 107, 108).
3. Osteocytes
Osteocytes comprise 90–95% of bone cells and derive from osteoblasts that have become embedded in bone matrix. Osteocyte dendritic processes ramify through networks of canaliculi and sense fluid shear stresses, communicating via gap junctions (109, 110). Mechanical stresses and localized microdamage stimulate osteocytes to release cytokines and chemotactic signals or induce apoptosis. In general, increased mechanical stress stimulates local osteoblastic bone formation, whereas reduced loading or microdamage results in osteoclastic bone resorption (111, 112). Osteocytes are thus mechano-sensors that control bone modeling and remodeling through their regulation of osteoclasts via the receptor activator of NF-κB (RANK) ligand (RANKL)/RANK pathway and osteoblasts via modulation of Wnt signaling (113–115).
4. Osteoclasts
Osteoclasts comprise 1–2% of bone cells. They are polarized multinucleated cells derived from fusion of mononuclear–myeloid precursors that resorb bone matrix and mineral. Attachment to bone is mediated by αVβ3 integrin that interacts with bone matrix proteins. These interactions lead to the formation of an actin ring and sealing zone with polarization of the osteoclast into ruffled border and basolateral membrane regions (116). Carbonic anhydrase II generates protons and bicarbonate within the osteoclast cytoplasm (117), and the HCO3− is exchanged for extracellular chloride at the basolateral membrane by a specific Cl−/HCO3− channel. An osteoclast-specific pump (H+-ATPase) transports protons across the ruffled border, whereas the CLCN7 channel transports chloride simultaneously. Within resorption lacuna, the acidic environment dissolves hydroxyapatite to release Ca2+ and HPO42−, whereas a secreted cysteine protease, cathepsin K, digests organic bone matrix. The degradation products are endocytosed at the ruffled border, transported across the cytoplasm in tartrate-resistant acid phosphatase-rich vesicles, and released at the basolateral membrane by exocytosis (117). Commitment of hematopoietic stem cells to the myeloid lineage is regulated by the PU.1 and micro-ophthalmia-associated transcription factors, which induce colony-stimulating factor (CSF) receptor (CSF-1R) expression. Macrophage CSF (M-CSF)/CSF-1R signaling stimulates expression of RANK, leading to osteoclast precursor commitment. RANKL/RANK signaling induces the key transcription factors, nuclear factor-κB (NF-κB) and nuclear factor of activated T cells cytoplasmic 1, leading to osteoclast differentiation and fusion (108, 118).
B. Intramembranous ossification
The flat bones of the face and skull form by intramembranous ossification, which occurs in the absence of a cartilage scaffold (Figure 4, A and B). Mesenchyme progenitors, located within vascularized connective tissue membranes, condense into nodules and differentiate to bone-forming osteoblasts. The osteoblasts secrete an osteoid matrix of type I collagen and chondroitin sulfate, which mineralizes to form an ossification center. The surrounding mesenchyme forms the periosteum, and cells at the inner surface differentiate into lining osteoblasts. Progressive bone formation results in extension of bony spicules and fusion of adjacent ossification centers (119).
C. Endochondral ossification
Endochondral ossification is the process by which long bones form on a cartilage scaffold (Figure 4, C and D) (101). Mesenchyme precursors condense and differentiate into chondrocytes, which proliferate and secrete a matrix containing type II collagen and proteoglycans that forms a cartilage template. At the primary ossification center, a coordinated program of chondrocyte proliferation, hypertrophic differentiation, and apoptosis leads to mineralization of cartilage. Subsequently, vascular invasion and migration of osteoblasts enables replacement of mineralized cartilage with trabecular bone. Concurrently, peripheral mesenchyme precursors in the perichondrium differentiate into osteoblasts and form a collar of cortical bone. Secondary ossification centers form at the ends of long bones and remain separated from the primary ossification center by the epiphyseal growth plates, where endochondral ossification continues.
D. Linear growth and bone maturation
Epiphyseal growth plates at both ends of developing bones comprise the reserve, proliferative, prehypertrophic, and hypertrophic zones, together with primary and secondary spongiosa (Figure 4, C and D) (101). The reserve zone contains uniform chondrocytes with a low proliferation index. Cells progress to the proliferative zone, become flattened, increase type II collagen synthesis, and form longitudinal columns. As chondrocytes mature, they express alkaline phosphatase, undergo terminal hypertrophic differentiation, secrete type X collagen, and increase in volume by 10-fold (120, 121). Finally, apoptosis of hypertrophic chondrocytes results in the release of angiogenic factors that stimulate vascular invasion and migration of osteoblasts and osteoclasts, leading to remodeling of calcified cartilage and formation of trabecular bone. This ordered process mediates linear growth until adulthood (101). Synchronously, the diameter of the long bone diaphysis increases by osteoblastic deposition of cortical bone beneath the periosteum, and the marrow cavity expands as a consequence of osteoclastic bone resorption at the endosteal surface.
Progression of endochondral ossification and linear growth is tightly regulated by a local feedback loop involving IHH and PTHrP (101, 122), and other factors including systemic hormones (thyroid hormones, GH, IGF-1, glucocorticoids, sex steroids), various cytokines, and growth factors (BMPs, FGFs, vascular endothelial growth factors) that act in a paracrine and autocrine manner (101). Linear growth continues until fusion of the growth plates during puberty, but bone mineralization and consolidation of bone mass continues until peak bone mass is achieved during the third to fourth decade (101, 123, 124).
E. The bone-remodeling cycle
Functional integrity and strength of the adult skeleton are maintained in a continuous process of repair by the “bone-remodeling cycle” (125) (Figure 5). The basic multicellular unit of bone-remodeling comprises osteoclasts and osteoblasts whose activities are orchestrated by osteocytes (113, 126, 127). Over 95% of the surface of the adult skeleton is normally quiescent because osteocytes exert resting inhibition of both osteoclastic bone resorption and osteoblastic bone formation (117).
Figure 5.

Bone remodeling compartment and “basic multicellular unit” of the bone-remodeling cycle. The bone remodeling cycle is initiated and orchestrated by osteocytes. Bone remodeling results from changes in mechanical load, structural microdamage, or exposure to systemic or paracrine factors. Monocyte/macrophage precursors differentiate to mature osteoclasts and resorb bone. Differentiation is induced by M-CSF and RANKL and inhibited by OPG. During reversal, osteoblastic progenitors are recruited to the site of resorption, synthesize osteoid, and mineralize new bone to repair the defect.
Under basal conditions, osteocytes secrete TGFβ and sclerostin, which inhibit osteoclastogenesis and Wnt-activated osteoblastic bone formation, respectively. Increased load or local microdamage results in a fall in local TGFβ levels (128), and activation of bone-lining cells leads to recruitment of osteoclast progenitors. Osteocytes and bone-lining cells express M-CSF and RANKL, the two cytokines required for osteoclastogenesis (114, 125). RANKL acts via several downstream signaling molecules, including c-fos, NF-κB, nuclear factor of activated T cells cytoplasmic 1, MAPK, and TNF receptor-associated factor-6 (129–131). RANKL also induces expression of αVβ3 integrin in osteoclast precursors, which signals via c-src to induce activation of small GTPases that are critical for formation of the actin ring sealing zone and osteoclast migration and survival. In addition to RANKL, osteoblasts and bone marrow stromal cells express osteoprotegerin (OPG). OPG is a secreted decoy receptor for RANKL and functions as the physiological inhibitor of RANK–RANKL signaling (117, 132). Thus, the RANKL:OPG ratio determines osteoclast differentiation and activity. This ratio is regulated by systemic hormones and local cytokines that control bone remodeling and include estrogen, PTH, glucocorticoids, TNF-α, IL-1, and prostaglandin E2 (133).
After this 30- to 40-day resorption phase, reversal cells remove undigested matrix fragments from the bone surface, and local paracrine signals released from degraded matrix recruit osteoblasts that initiate bone formation. Over the next 150 days, osteoblasts secrete and mineralize new bone matrix (osteoid) to fill the resorption cavity. Although commitment of mesenchyme precursors to the osteoblast lineage requires both Wnt and BMP signaling, the canonical Wnt pathway subsequently acts as the master regulator of osteogenesis (134–136). Physiological negative regulation of canonical Wnt signaling is mediated by the osteocyte, which secretes soluble factors (sclerostin, Dickkopf1-related protein 1 [DKK1], and secreted frizzled related protein 1) that interfere with the interaction between Wnt ligands and their receptor and coreceptor (113, 126). During the process of bone formation, some osteoblasts become embedded within newly formed bone and undergo terminal differentiation to osteocytes. Secretion of sclerostin and other Wnt inhibitors by these osteocytes leads to cessation of bone formation and a return to the quiescent state in which osteoblasts become bone-lining cells (113, 126).
This cycle of targeted bone modeling and remodeling enables the adult skeleton to repair old or damaged bone, react to changes in mechanical stress, and respond rapidly to the demands of mineral homeostasis.
IV. Skeletal Target Cells and Downstream Signaling Pathways
A. TSH actions in chondrocytes, osteoblasts and osteoclasts
1. Expression of TSHR and its ligands in skeletal cells
TSHR is expressed predominantly in thyroid follicular cells, but expression in chondrocytes, osteoblasts, and osteoclasts suggests that TSH exerts direct actions in cartilage and bone (32, 137). Although pituitary TSH functions as a systemic hormone, local expression of TSHR ligands in bone has also been investigated. TSHα and TSHβ subunits are not expressed in primary human or mouse osteoblasts or osteoclasts (138, 139). Nevertheless, an alternative splice variant of Tshb (Tshb-sv) has been identified in mouse bone marrow (140, 141). Expression of this variant in bone marrow-derived macrophages activated cAMP in cocultured, stably transfected TSHR-overexpressing Chinese hamster ovary cells (142). mRNA encoding the isoform was also identified at low levels in primary mouse osteoblasts but not osteoclasts (139). The alternative TSHR ligand, thyrostimulin, is also expressed in osteoblasts and osteoclasts, and studies of Gpb5−/− mice lacking thyrostimulin indicated that thyrostimulin regulates osteoblastic bone formation during early skeletal development. However, the underlying mechanisms remain unknown because thyrostimulin failed to influence osteoblast proliferation or differentiation or to activate cAMP, ERK, P38 MAPK, or AKT signaling pathways in primary osteoblasts or bone marrow stromal cells in vitro (139).
2. Chondrocytes
Only limited information has been published regarding the TSHR in cartilage. In mesenchymal stem cells, TSH stimulated self-renewal and expression of chondrogenic marker genes, suggesting that TSH may increase chondrocyte differentiation (143). Growth plate cartilage and cultured chondrocytes express TSHR, and treatment with TSH increased cAMP activity and decreased expression of SOX9 and type IIa collagen expression in primary chondrocytes (137).
Initial studies, therefore, suggest that TSHR signaling might inhibit chondrocyte differentiation (Figure 6).
Figure 6.

Actions of T3 and TSH in skeletal cells. A, T3 and TSH actions in osteocytes have not been investigated, and it is unknown whether osteocytes express thyroid hormone transporters, deiodinases, TRs, or the TSHR. B, Chondrocytes express MCT8, MCT10, and LAT1 transporters, DIO3 (D3), TRs (predominantly TRα), and TSHR. T3 inhibits proliferation and stimulates prehypertrophic and hypertrophic chondrocyte differentiation, whereas TSH might inhibit proliferation and matrix synthesis. C, Osteoblasts express MCT8 and LAT1/2 transporters, the DIO2 (D2) and D3, TRs (predominantly TRα), and TSHR. Most studies indicate that T3 stimulates osteoblast differentiation and bone formation. Contradictory data suggest that TSH may stimulate, inhibit, or have no effect on osteoblast differentiation and function. D, Osteoclasts express MCT8, D3, TRs and the TSHR. Currently, it is unclear whether T3 acts directly in osteoclasts or whether indirect effects in the osteoblast lineage mediate its actions. Most studies indicate that TSH inhibits osteoclast differentiation and function.
3. Osteoblasts
Expression of TSHR in UMR106 rat osteosarcoma cells was reported in 1998 (144), and subsequently, expression of TSHR mRNA and protein was identified in osteoblasts and osteoclasts (32, 38, 138, 139, 145). The lack of TSHα and -β expression in osteoblasts and osteoclasts (138, 139), indicates that TSH does not have local autocrine effects in these cells. Nevertheless, treatment of osteoblasts with TSH in vitro inhibited osteoblastogenesis and reduced expression of type I collagen, bone sialoprotein, and osteocalcin (32). Inhibition of low-density lipoprotein receptor-related protein 5 mRNA in these studies suggested that the effects of TSH on osteoblastogenesis and function might be mediated via Wnt signaling (32).
By contrast, Sampath et al (145) and Baliram et al (142) showed that TSH stimulates osteoblast differentiation and function. Furthermore, in embryonic stem cell (ESC) cultures, TSH stimulated osteoblastic differentiation via protein kinase C and the noncanonical Wnt pathway components Frizzled and Wnt5a (146). In human SaOS2 osteosarcoma cells, TSH also stimulated proliferation and differentiation, as measured by alkaline phosphatase, and increased IGF-1 and IGF-2 mRNA expression together with complex regulatory effects on IGF binding proteins (IGFBPs) and their proteases (147). Finally, TSH stimulated β-arrestin 1, leading to activation of ERK, P38 MAPK, and AKT signaling pathways and osteoblast differentiation in stably transfected human osteoblastic U2OS-TSHR cells that overexpress the TSHR (148).
Despite these contrasting findings, Tsai et al (149) had previously shown only low levels of TSHR expression, TSH binding, and cAMP activation in human osteoblasts and concluded that TSH was unlikely to have a physiological role. Further studies also demonstrated only low levels of TSHR protein in calvarial osteoblasts; in these studies, treatment with TSH and TSHR-stimulating antibodies failed to induce cAMP, and TSH did not affect osteoblast differentiation or function (38, 138).
Overall, findings have been interpreted to suggest that changes in TNFα, RANKL, OPG, and IL-1 signaling in response to TSH might be mediated via an alternative G protein and not by cAMP (32, 38, 150). Thus, although the TSHR is expressed in osteoblasts, in vitro data are contradictory, suggesting that TSH may inhibit, enhance, or have no effect on osteoblast differentiation and function (Figure 6). Furthermore, the physiological second messenger pathway that lies downstream of activated TSHR in osteoblasts has not yet been defined.
4. Osteoclasts
Bassett et al (138) showed that mouse osteoclasts express low levels of TSHR protein, but treatment with TSH and TSHR-stimulating antibodies did not affect osteoclast differentiation and function or elicit a cAMP response. Nevertheless, most studies have shown that TSH inhibits osteoclast formation and function. Thus, treatment of monocyte precursors with TSH inhibited osteoclastogenesis and dentine resorption (32, 38, 145), and TSH and TSHR-stimulating antibodies also inhibited osteoclastogenesis in mouse ESC cultures treated with M-CSF, RANKL, vitamin D, and dexamethasone (150). Zhang et al (151) also showed that TSH inhibits tartrate-resistant acid phosphatase, matrix metalloproteinase (MMP)-9, and cathepsin K expression and osteoclastogenesis in RAW264.7 cells.
In vitro studies using bone marrow cultures from Tshr−/− mice revealed that inhibitory effects of TSH on osteoclastogenesis were mediated by TNFα acting via disruption of activator protein 1 and NF-κB signaling (32, 38, 152), and the pathogenesis of bone loss in Tshr−/− mice was proposed to be mediated by elevated levels of TNFα (153). Nevertheless, the mechanisms of bone loss in Tshr−/− mice appear complex, and the underlying signaling pathways remain incompletely defined. Thus, TSH inhibited osteoclastogenesis in wild-type mice, and TNFα stimulated osteoclastogenesis in wild-type and Tshr−/− mice. Accordingly, TSH inhibited TNFα via activator protein 1 and RANKL-NFκB signaling pathways in osteoclasts in vitro (153), although in previous studies TSH had been shown to stimulate TNFα in ESC-derived osteoblasts (146). Together, these findings were proposed as a counter-regulatory mechanism of TNFα inhibition and stimulation in osteoclasts and osteoblasts, respectively (153).
Overall, most studies indicate that TSHR signaling inhibits osteoclastogenesis and function by complex mechanisms, primarily involving TNFα (Figure 6).
B. T3 actions in chondrocytes, osteoblasts, and osteoclasts
1. Expression of thyroid hormone transporters
The thyroid hormone transporter MCT8 is expressed and regulated by thyroid status in growth plate chondrocytes, osteoblasts, and osteoclasts (61, 154). Recent studies indicate that OATP1c1 is not expressed in the skeleton (154). MCT10 appears to be the major transporter expressed in the growth plate (155), whereas expression of the less specific LAT1 and LAT2 transporters has also been detected in bone (61, 154, 155). Nevertheless, the physiological importance and possible redundancy of thyroid hormone transporters in the skeleton has yet to be determined (Figure 6).
2. Expression of deiodinases
Thyroid hormone metabolism occurs in skeletal cells (156). Although DIO1 is not expressed in cartilage or bone (61, 156), the activating enzyme DIO2 is expressed in osteoblasts (60, 61). Dio2 mRNA has also been detected in the embryonic mouse skeleton as early as embryonic day (E) 14.5 and increases until E18.5 (157, 158). In the developing chick growth plate, DIO2 activity is restricted to the perichondrium (159), indicating that the enzyme has a role in local regulation of thyroid hormone signaling during fetal bone development. DIO2 is also expressed in primary mesenchymal stem cells, in which expression is strongly induced after treatment with BMP-7 (160). The inactivating DIO3 enzyme is present in all skeletal cell lineages, particularly during development, with the highest levels of activity in growth plate chondrocytes before weaning (61, 157). Together, these data suggest that control of tissue T3 availability by DIO2 and DIO3 is likely to be important for skeletal development, linear growth, and osteoblast function (Figure 6).
3. Expression of TRs
TRs are expressed at sites of intramembranous and endochondral bone formation. The localization of TR proteins to reserve and proliferative zone growth plate chondrocytes, but not hypertrophic cells (161, 162), suggests that progenitor cells and proliferating chondrocytes are primary T3-target cells but differentiated chondrocytes lose the ability to respond to T3. Both TRα1 and TRβ1 are expressed in bone, and quantitative RT-PCR studies reveal that levels of TRα1 are at least 10-fold greater than TRβ1 (163, 164), suggesting that TRα1 is the predominant mediator of T3 action in bone. Nevertheless, other studies also indicate that TRβ may play a role (165–167).
TRα1, TRα2, and TRβ1 are expressed in reserve and proliferative zone epiphyseal growth plate chondrocytes (161, 162, 168–173) and in immortalized osteoblastic cells from several species (170, 174–181), as well as in primary osteoblasts and osteoblastic bone marrow stromal cells (175, 182, 183). However, it is unknown whether TRs are expressed in osteocytes (184, 185). Thyroid hormones stimulate osteoclastic bone resorption (186, 187), but this effect may be indirect and mediated by T3-responsive osteoblasts (188, 189). Although immunolocalization of TR proteins and detection of TR mRNAs by in situ hybridization in osteoclasts from pathological human osteophytes and osteoclastoma tissue were reported in early studies (168, 174, 190), TR antibodies lack sufficient sensitivity to detect expression of endogenous protein, and it remains uncertain whether osteoclasts express functional TRs or respond directly to T3.
Overall, current studies indicate that reserve zone and proliferating chondrocytes, osteoblastic bone marrow stromal cells, and osteoblasts are major T3-target cells in bone and predominantly express TRα (Figure 6).
4. Chondrocytes
Hypertrophic chondrocyte differentiation and vascular invasion of cartilage are sensitive to thyroid status (191); these findings support early studies (192–194) and reinforce the critical importance of T3 for endochondral ossification and linear growth. Nevertheless, studies of T3 action in chondrocytes cultured in monolayers are conflicting due to the species, source of chondrocytes, and culture conditions studied (171, 195–199). Consequently, several three-dimensional culture systems have been devised to investigate the T3-regulated differentiation potential of chondrocytes in vitro (196, 200–202). T3 treatment of chondrogenic ATDC5 cells, mesenchymal stem cells, primary growth plate chondrocytes, and long bone organ cultures inhibits cell proliferation and concomitantly stimulates hypertrophic chondrocyte differentiation and cellular apoptosis (161, 197, 203–209). T3 promotes hypertrophic differentiation by induction of cyclin-dependent kinase inhibitors to regulate the G1-S cell cycle checkpoint (200). Subsequently, T3 stimulates BMP4 signaling, synthesis of a collagen X matrix, and expression of alkaline phosphatase and MMP13 to facilitate progression of hypertrophic differentiation and cartilage mineralization (120, 161, 197, 203–206). In addition, T3 regulation of growth plate chondrocyte proliferation and differentiation in vitro involves activation of IGF-1 and Wnt signaling (210–212).
The regulatory effects of T3 on endochondral ossification and linear growth in vivo involve interactions with key pathways that regulate growth plate maturation including IHH, PTHrP, IGF-1, Wnt, BMPs, FGFs, and leptin (213–215). IHH, PTHrP, and BMP receptor-1A participate in a negative feedback loop that promotes growth plate chondrocyte proliferation and inhibits differentiation, thereby controlling the rate of linear growth. The set point of this feedback loop is sensitive to thyroid status (162, 216) and is regulated by local thyroid hormone metabolism and T3 availability (159). Furthermore, T3 stimulates expression of genes involved in cartilage matrix synthesis, mineralization, and degradation, including matrix proteoglycans (203, 204, 217–221) and collagen degrading enzymes such as aggrecanase-2 (a disintegrin and metalloproteinase with thrombospondin type 1 motif 5 [ADAMTS5]) and MMP13 (205, 222, 223), as well as BMP4, Wnt4, and FGF receptor (FGFR) 3 (120, 210, 224–226).
In summary, thyroid hormone is essential for coordinated progression of endochondral ossification, acting to stimulate genes that control chondrocyte maturation and cartilage matrix synthesis, mineralization, and degradation.
5. Osteoblasts
Although primary osteoblasts (170, 172, 227–233) and several osteoblastic cell lines (176, 178–181, 223, 234–245) respond to T3 in vitro, the consequences of T3 stimulation vary considerably and depend on species, the anatomical origin of osteoblasts (183, 246–248), cell type, passage number, cell confluence, stage of differentiation, and the dose and duration of T3 treatment. Thus, T3 has been shown to stimulate, inhibit, or have no effect on osteoblastic cell proliferation. A general consensus, however, indicates that T3 stimulates osteoblast proliferation and differentiation and bone matrix synthesis, modification, and mineralization. T3 increases expression of osteocalcin, osteopontin, type I collagen, alkaline phosphatase, IGF-1 and its regulatory binding proteins (IGFBP-2 and -4), IL-6 and -8, MMP9, MMP13, tissue inhibitor of metalloproteinase-1, and FGFR1, leading to activation of MAPK signaling, and also regulates the Wnt pathway (165, 179, 180, 223, 227, 229, 232, 235–237, 243–245, 249–259). Furthermore, IGFBP-6 interacts directly with TRα1 to inhibit T3-stimulated increases in alkaline phosphatase activity and osteocalcin mRNA in osteoblastic cells (260). Thus, T3 stimulates osteoblast activity both directly and indirectly via complex pathways involving many growth factors and cytokines. T3 may also potentiate osteoblast responses to PTH (233) by modulating expression of the PTH/PTHrP receptor (176).
Despite the many potential T3-target genes identified in osteoblasts, little information is available regarding mechanisms by which their expression is modulated, and T3 regulation may involve other signaling pathways. For example, T3 regulates osteoblastic cell morphology, cytoskeleton, and cell-cell contacts in vitro (234, 239, 240). In addition, T3 stimulates osteocalcin via nongenomic actions mediated by suppression of Src (261). T3 also phosphorylates and activates p38 MAPK and stimulates osteocalcin expression in MC3T3 cells (250, 262), a pathway that is enhanced by AMPK activation (263) but inhibited by cAMP (264) and Rho-kinase (265). A recent study further demonstrated that nongenomic signaling in osteoblasts and osteosarcoma cells is mediated by a plasma membrane-bound N-terminal truncated isoform of TRα1 that is palmitoylated and interacts with caveolin-containing membrane domains. Acting via this isoform, T3 stimulated osteoblast proliferation and survival via increased intracellular Ca2+, nitric oxide, and cGMP leading to activation of protein kinase GII, Src, and ERK (75).
Overall, many studies in primary cultured and immortalized osteoblastic cells demonstrate the complexities of T3 action in bone and emphasize the importance of the cellular system under study. Many of these T3 actions involve interactions with bone matrix and local paracrine and autocrine factors via mechanisms that have yet to be determined.
6. Osteoclasts
Thyroid hormone excess results in increased osteoclast numbers and activity in vivo, leading to bone loss. Osteoclasts express TRα1 and TRβ1 mRNAs, but it is not clear whether functional receptors are expressed because TR antibodies lack sufficient sensitivity to detect endogenous proteins. Studies of mixed cultures containing osteoclast lineage cells and bone marrow stromal cells have been contradictory, and it is not clear whether stimulation of osteoclastic bone resorption results from direct T3 actions in osteoclasts or indirect effects mediated by primary actions in cells of the osteoblast lineage (186–188, 254, 266). Studies of fetal long bone and calvarial cultures (173, 267, 268) implicated various cytokines and growth factors including IGF-1 (269, 270), prostaglandins (186), interleukins (271), TGFβ (238, 272), and interferon-γ (186) as mediators of secondary responses in osteoclasts. Similarly, treatment of immortalized osteoblasts or primary bone marrow stromal cells resulted in increased RANKL, IL-6, IL-8, and prostaglandin E2 expression and inhibition of OPG, consistent with an indirect effect of thyroid hormones on osteoclast function (254, 258, 266). Other studies, however, suggest that the effects of T3 on osteoclastogenesis are independent of RANKL signaling (273, 274). A further complication is that whereas TR expression is well documented in osteoblastic cells, some of the effects of T3 on bone organ cultures are extremely rapid and involve mobilization of intracellular calcium stores to suggest that nongenomic TR-independent actions of T3 may be relevant (275).
Overall, it is unclear whether T3 acts directly in the osteoclast lineage or whether its stimulatory effects on osteoclastogenesis and bone resorption are secondary responses to direct actions of T3 in osteoblasts, osteocytes, stromal cells, or other bone marrow cell lineages.
V. Genetically Modified Mice (Table 1)
Table 1.
Skeletal Phenotype of Genetically Modified Mice
| Mouse Model | Genotype | Systemic Thyroid Status | Developing Skeleton | Adult Skeleton | T3 Action in Bone | Refs. |
|---|---|---|---|---|---|---|
| Congenital hypothyroid | ||||||
| Pax8−/− | Maximal Tshr signaling Apo TRα and TRβ |
No Thyroid T4 undetectable T3 undetectable TSH 1900× |
Impaired linear growth, delayed endochondral ossification, reduced cortical bone, reduced bone mineralization | Majority die by weaning; coarse plate-like trabeculae with impaired trabecular remodeling; reduced cortical thickness; reduced mineralization | Reduced T3 action | 138, 282, 314 |
| TSHR mutants | ||||||
| Tshr−/− | No Tshr Apo TRα and TRβ |
Thyroid hypoplasia T4 undetectable T3 undetectable TSH > 500× |
Severe growth retardation; impaired linear growth; reduced mineral density; increased bone resorption; increased bone formation or decreased bone formation | Die by weaning unless treated with thyroid extract Low bone mass, high bone turnover not reversed by thyroid extract supplementation Enhanced bone loss in supplemented animals rendered thyrotoxic |
NR | 32, 140, 153, 276 |
| TshrP556L/P556L(hyt/hyt) | Absent Tshr signaling | Thyroid hypoplasia | Impaired linear growth, delayed endochondral ossification, reduced cortical bone, reduced bone mineralization | NR | Reduced T3 action | 138, 529 |
| Apo TRα and TRβ | T4 0.06× | |||||
| T3 0.06× | ||||||
| TSH 2300× | ||||||
| Gpb5−/− | No thyrostimulin (Alternative high affinity Tshr ligand) |
Juveniles: T4 0.7×, (males only) | Normal linear growth; endochondral and intramembranous ossification; increased bone volume and mineralization due to increased osteoblastic bone formation | Skeletal phenotype resolved by early adulthood | NR | 139 |
| T3 normal | ||||||
| TSH 3 × (males only) | ||||||
| Adults: T4,T3 and TSH normal | ||||||
| Compound mutants | ||||||
| Tshr−/−Tnfα−/− | No Tshr or TNFα | Treated with thyroid extract from weaning: T4, T3, and TSH not reported | NR | Amelioration of low bone mass, high bone turnover phenotype in Tshr−/− mice | NR | 153 |
| TR mutants | ||||||
| TRα mutants | ||||||
| TRα−/− | No TRα1 or TRα2; TRΔα1 and TRΔα2 preserved |
Hypothyroid: T4 0.1× T3 0.4× TSH 2× (GH normal) |
Severe growth delay; delayed endochondral ossification; impaired chondrocyte differentiation; reduced mineralization | Die by weaning unless T3 treated | NR | 73, 310, 311 |
| TRα1−/− | No TRα1 or TRΔα1; TRα2 and TRΔα2 preserved |
Mild hypothyroidism T4 0.7 × (males only) T3 normal TSH 0.8× |
No growth retardation | NR | NR | 304 |
| TRα2−/− | No TRα2 or TRΔα2 | Mild hypothyroidism | No growth retardation | Reduced BMD; reduced cortical bone | NR | 253 |
| TRα1 and TRΔα1 over-expression | T4 0.8× | Normal endochondral ossification | ||||
| T3 0.7× | ||||||
| TSH normal | ||||||
| (GH normal, IGF-1 low) | ||||||
| TRα1GFP/GFP | No TRα2 2× increase in TRα1GFP |
Euthyroid, T4, T3 and TSH normal | Normal postnatal growth | NR | NR | 308 |
| TRα0/0 | No TRα | Euthyroid | Transient growth delay, delayed endochondral ossification, impaired chondrocyte differentiation, reduced mineral deposition | Osteosclerosis, increased trabecular bone volume, reduced osteoclastic bone resorption | Reduced T3 action | 285, 313 |
| Normal TRβ | T4 normal | |||||
| T3 normal | ||||||
| TSH normal | ||||||
| (GH normal) | ||||||
| TRα1PV/+ | Heterozygous dominant-negative TRα receptor |
Euthyroid T4 normal T3 1.2× TSH 1.5–2× (GH normal) |
Severe persistent growth retardation; delayed intramembranous and endochondral ossification; impaired chondrocyte differentiation; reduced mineralization | Grossly dysmorphic bones; impaired ossification and bone modelling; Osteosclerosis, increased trabecular and cortical bone volume; reduced osteoclastic bone resorption (resistant to T4 treatment) |
Reduced T3 action | 278, 317, 319 |
| TRα1R384C/+ | Heterozygous dominant-negative TRα receptor (10 × lower affinity for T3) |
Euthyroid adults Mild hypothyroidism (P10–35) T4 0.8× T3 0.7× TSH 0.7× (GH reduced in juveniles) |
Transient growth delay; delayed intramembranous and endochondral ossification; impaired chondrocyte differentiation | Impaired bone modelling; osteosclerosis, increased trabecular bone volume; increased mineralization; reduced osteoclastic bone resorption (ameliorated by T3 treatment) | Reduced T3 action | 312, 530 |
| TRα1R398H/+ | Heterozygous dominant-negative TRα receptor |
Euthyroid juvenilesT4 and T3 normal TSH 3.4× |
NR | NR | NR | 531 |
| TRα1L400R/+ (TRAMI/+xSycp1-Cre) | Global expression dominant-negative receptor TRα1L400R |
Euthyroid T4 and T3 normal TSH normal (GH low) |
Severe persistent growth retardation; delayed endochondral ossification | NR | NR | 320 |
| TRα1L400R/+/C1 (TRAMI/+x Col1a1-Cre) | Osteoblast expression dominant-negative receptor TRα1L400R | Euthyroid, T4 and T3 and TSH normal | No skeletal abnormalities reported after limited analysis | No skeletal abnormalities reported after limited analysis | NR | 321 |
| TRα1L400R/+/C2 (TRAMI/+x Col2a1-Cre) | Chondrocyte and osteoblast expression dominant-negative receptor TRα1L400R | Euthyroid, T4 and T3 and TSH normal | Persistent growth retardation Delay in endochondral ossification Skull abnormalities Reduced cortical and trabecular bone Decreased mineralization |
Short stature | NR | 321 |
| TRβ mutants | ||||||
| TRβ−/− | No TRβ Normal TRα |
RTH and goiter T4 3–4× T3 3–4× TSH 2.6–8× |
Persistent short stature, advanced endochondral and intramembranous ossification, increased mineral deposition | Osteoporosis, reduced mineralization, increased osteoclastic bone resorption | Increased T3 action | 9, 80, 285, 312, 313 |
| TRβ2−/− | No TRβ2 TRβ1 preserved |
Mild RTH T4 1–3× T3 1.3–1.5× TSH 1.2–2.5× |
No growth abnormality | NR | NR | 78, 309 |
| TRβPV/PV | Homozygous dominant-negative TRβ receptor |
Severe RTH and goiter T4 15× T3 9× TSH 400× |
Accelerated prenatal growth; persistent postnatal growth retardation; advanced intramembranous and endochondral ossification; increased mineralization | NR | Increased T3 action | 164, 278, 323 |
| TRβΔ337T/Δ337T | Homozygous dominant-negative TRβ receptor |
Severe RTH and goiter T4 15× T3 10× TSH 50× |
No growth phenotype | NR | NR | 82 |
| TRβ transgenics | ||||||
Tshb TRβG345R TRβG345R
|
Pituitary expression of dominant-negative TRβG345R |
RTH T4 1.2× TSH normal |
Normal growth | NR | NR | 532 |
| CgaTRβΔ337T
|
Pituitary expression of dominant-negative TRβΔ337T | T4 normal TSH 3× (Reduced bioavailability?) |
NR | NR | NR | 533 |
| ActbTRβPV
|
Ubiquitous expression of dominant-negative TRβPV |
RTH T4 1.5× TSH normal |
Impaired weight gain | NR | NR | 534 |
| CgaTRβPV
|
Pituitary expression of dominant-negative TRβPV |
Euthyroid T4, T3 and TSH normal |
Impaired weight gain | NR | NR | 535 |
| Compound mutants | ||||||
| TRα−/−TRβ−/− | No TRα1, TRα2 or TRβ TRΔα1 and TRΔα2 preserved |
RTH and small goiter T4 10× T3 10× TSH > 100× |
Growth delay similar to TRα−/−; delayed endochondral ossification; impaired chondrocyte differentiation; reduced mineralization | Die at or near weaning | NR | 311 |
| TRα1−/−TRβ−/− | No TRα1, TRΔα1 or TRβ TRα2 and TRΔα2 preserved |
RTH and large goiter T4 60× T3 60× TSH > 160× (GH/IGF-1 low) |
Persistent growth retardation; delayed endochondral ossification; reduced mineralization | Reduced trabecular and cortical BMD GH treatment corrects growth but not ossification defect |
NR | 305–307 |
| TRα2−/−TRβ−/− | No TRα2, TRΔα2 or TRβ | Mild hypothyroidism | Transient growth delay | NR | NR | 309 |
| TRα1 and TRΔα1 overexpression | T4 0.7× | |||||
| T3 0.8× | ||||||
| TSH normal | ||||||
| TRα0/0TRβ−/− | No TRα or TRβ |
RTH and goiter T4 14× T3 13× TSH > 200× (GH/IGF-1 low) |
More severe phenotype than TRα0/0; growth delay; delayed endochondral ossification; impaired chondrocyte differentiation; reduced mineralization | NR | NR | 219, 313, 314 |
| Pax8−/−TRα1−/− | No TRα1 or TRΔα1 | No Thyroid | Growth retardation similar to Pax8−/− | Die by weaning | NR | 315 |
| TRα2/TRΔα2 preserved | T4 undetectable | |||||
| Apo TRβ | T3 undetectable | |||||
| Maximal Tshr signaling | TSH NR | |||||
| Pax8−/−TRα0/0 | No TRα | No Thyroid | Growth retardation less than Pax8−/− and similar to TRα0/0β−/−; delayed endochondral ossification; mice survive to adulthood | NR | NR | 314 |
| Apo TRβ | T4 undetectable | |||||
| Maximal Tshr signaling | T3 undetectable TSH > 400× |
|||||
| Pax8−/−TRβ−/− | No TRβ | No Thyroid | Growth retardation similar to Pax8−/−; severely delayed endochondral ossification | Die by weaning | NR | 314 |
| Apo TRα | T4 undetectable | |||||
| Maximal Tshr signaling | T3 undetectable TSH > 400× |
|||||
| Deiodinase mutants | ||||||
| C3H/HeJ | 80% reduction in D1 activity | T4 1.6× T3 normal rT3 3× |
Not reported | NR | NR | 294, 296 |
| Dio1−/− | No Dio1 | T4 1.4× T3 normal TSH normal rT3 4× |
Normal growth | NR | NR | 52, 288 |
| Dio2−/− | No Dio2 | T4 1–1.3× | Normal intramembranous and endochondral ossification | Reduced bone formation | Reduced T3 action in osteoblasts | 60, 288, 536 |
| T3 normal | Increased mineralization | |||||
| TSH 3–15× | Brittle bones | |||||
| rT3 normal | ||||||
| Dio3−/− | No Dio3 |
Perinatal thyrotoxicosis Adult central hypothyroidism |
Reduced body length | NR | NR | 299, 300 |
| Compound mutants | ||||||
| Dio1−/− Dio2−/− | No Dio1 or Dio2 | T4 1.7× T3 normal TSH 15× rT3 6.5× |
Normal growth | NR | NR | 288 |
| Transporter mutants | ||||||
| Mct8−/− | No Mct8 | T4 0.7–0.3× T3 1–1.4× TSH 1–3× rT3 0.2× |
NR | NR | NR | 288, 289, 291, 292, 537 |
| Mct10Y88*/Y88* | No Mct10 |
P21: T4 normal; T3 0.8× Adult: T4 and T3 normal |
NR | NR | NR | 290, 291 |
| Oatp1c1−/− | No Oatp1c1 | T4, T3 and TSH normal | NR | NR | NR | 292, 293 |
| Compound mutants | NR | NR | ||||
| Mct8−/− Mct10Y88*/Y88* | No Mct8 or Mct10 | P21, T4 0.6×; T3 1.6×. Adult, T4 normal; T3 3× |
NR | NR | NR | 291 |
| Mct8−/− Oatp1c1−/− | No Mct10 or Oatp1c1 | T4 0.3× T3 2× TSH 5× |
Growth retarded after P16 | NR | NR | 292 |
| Mct8−/− Dio1−/− | No Mct8 or Dio1 | T4 1.4× T3 normal TSH 1.4× rT3 normal |
Mild growth retardation | NR | NR | 288 |
| Mct8−/− Dio2−/− | No Mct8 or Dio2 | T4 0.4× T3 1.7× TSH 50× rT3 0.2× |
Mild growth retardation | NR | NR | 288, 538 |
| Mct8−/− Dio1−/− Dio2−/− | No Mct8, Dio1 or Dio2 | T4 2.3× T3 1.4× TSH 100× rT3 2× |
Growth retarded | NR | NR | 288 |
Abbreviation: NR, not reported.
A. Targeting TSHR signaling
1. Skeletal development and growth
TSHR knockout (Tshr−/−) mice have congenital hypothyroidism with undetectable thyroid hormones and 500-fold elevation of TSH. Tshr−/− mice are growth retarded and usually die by 4 weeks of age (276). Nevertheless, animals supplemented with thyroid extract from weaning regain normal weight by 7 weeks. Heterozygous Tshr+/− mice are euthyroid with normal linear growth. Untreated Tshr−/− mice had a 30% reduction in bone mineral density (BMD) with evidence of increased bone formation and resorption when analyzed during growth at 6 weeks of age. Tshr−/− mice treated with thyroid extract displayed a 20% reduction in BMD and reduced calvarial thickness, although histomorphometry responses were not reported (32). Heterozygotes had a 6% reduction in total BMD, affecting only some skeletal elements, no change in calvarial thickness, and no difference in parameters of bone resorption or formation. These studies were interpreted to indicate that TSH suppresses bone remodeling, and TSH was proposed as an inhibitor of bone formation and resorption (32). Normally, T4 and T3 levels rise rapidly to a physiological peak at 2 weeks of age in mice, and growth velocity is maximal at this time (277, 278). Because Tshr−/− mice are only supplemented with thyroid extract from weaning at around 3 weeks of age (32, 276), they remain grossly hypothyroid at this critical stage of skeletal development. Thus, the phenotype reported in Tshr−/− mice also reflects the effects of severe hypothyroidism followed by incomplete “catch-up” growth and accelerated bone maturation in response to delayed thyroid hormone replacement (138, 279–281). Furthermore, treatment with supraphysiological doses of T4 for 21 days resulted in increased bone resorption and a greater loss of bone in Tshr−/− mice compared to wild-type controls, suggesting that Tshr deficiency exacerbates bone loss in thyrotoxicosis (140).
To investigate the relative importance of T3 and TSH in bone development, two contrasting mouse models of congenital hypothyroidism were compared, in which the reciprocal relationship between thyroid hormones and TSH was either intact or disrupted (138). Pax8−/− mice lack a transcription factor required for thyroid follicular cell development (282) and hyt/hyt mice harbor a loss-of-function mutation in Tshr (283). Pax8−/− mice have a 2000-fold elevation of TSH (277, 284) and a normal TSHR, whereas hyt/hyt mice have a 2000-fold elevation of TSH but a nonfunctional TSHR. Thus, if TSHR has the predominant role, these mice should display opposing skeletal phenotypes. However, Pax8−/− and hyt/hyt mice each displayed impaired linear growth, delayed endochondral ossification, reduced cortical bone mass, defective trabecular bone remodeling, and reduced bone mineralization (138). Indeed, both Pax8−/− and hyt/hyt mice have impaired chondrocyte, osteoblast, and osteoclast activities that are typical of thyroid hormone deficiency (161, 187, 189, 200, 230, 231, 246) and characteristic of juvenile hypothyroidism (278–281, 285). Nevertheless, the actions of thyroid hormone and TSH are not mutually exclusive, and the skeletal consequences of grossly abnormal thyroid hormone levels in Pax8−/− and hyt/hyt mice may mask effects of TSH on the skeleton.
To investigate further the role of the TSHR in bone, Gpb5−/− mice lacking the high-affinity ligand thyrostimulin were characterized. Juvenile Gpb5−/− mice had increased bone volume and mineralization due to increased osteoblastic bone formation, whereas no effects on linear growth or osteoclast function were identified. Resolution of these abnormalities by adulthood was consistent with transient postnatal expression of thyrostimulin in bone (139). Despite this, treatment of osteoblasts with thyrostimulin in vitro had no effect on cell proliferation, differentiation, and signaling, suggesting that thyrostimulin acts via unknown cellular and molecular mechanisms to inhibit bone formation indirectly during skeletal development.
2. Adult bone maintenance
Two animal studies have investigated the therapeutic potential of TSH to inhibit bone turnover. Ovariectomized rats were treated with TSH at doses insufficient to alter circulating T3, T4, or TSH levels (145). Intermittent TSH treatment resulted in reduced bone resorption markers, but increased formation markers, together with a dose-related increase in BMD. Bone volume, trabecular architecture, and strength parameters were either preserved or improved, although no dose relationship was evident. This osteoblastic response to TSH (145) contrasts with findings in Tshr−/− mice, which also displayed increased osteoblastic bone formation despite the absence of TSHR signaling (32). In further studies, intermittent treatment of ovariectomized rats or mice with similar concentrations of TSH was investigated by bone densitometry and micro-computed tomography. In these studies, TSH prevented bone loss and increased bone mass after ovariectomy (286). Furthermore, treatment of thyroidectomized and parathyroidectomized rats with intermittent TSH injections also suppressed bone resorption and stimulated bone formation, resulting in increased bone volume and strength (287).
B. Targeting thyroid hormone transport and metabolism
1. Thyroid hormone transporters
Mct8−/y knockout mice have elevated T3 but decreased T4 levels and recapitulate the systemic thyroid abnormalities observed in Allan-Herndon-Dudley syndrome. Mct8−/y mice, however, do not display the neurological abnormalities, and they exhibit only minor growth delay before postnatal day (P) 35, suggesting that other transporters such as OATP1c1 may compensate for a lack of MCT8 in mice (288, 289). Mct10 mutant mice harbor an ENU loss-of-function mutation and exhibit normal weight gain during growth (290, 291). Furthermore, mice lacking both MCT8 and MCT10 also showed no evidence of growth retardation (291), suggesting that both transporters are dispensable in the skeleton. Similarly, Oatp1c1−/− knockout mice exhibited normal weight gain during growth (292) and had no evidence of growth retardation (293). However, double mutants lacking both Mct8 and Oatp1c1 had growth retardation from P16 (292), confirming redundancy among thyroid hormone transporters in the regulation of skeletal growth. Finally, mice lacking Mct8 and Dio1 or Dio2 display mild growth retardation, whereas triple mutants lacking Mct8, Dio1, and Dio2 exhibit more severe growth delay (288), indicating cooperation between thyroid hormone transport and metabolism in vivo during linear growth.
2. Deiodinases
A minor and transient impairment of weight gain was reported in male Dio2−/− mice, whereas weight gain and growth were normal in Dio1−/− and DIO1-deficient C3H/HeJ mice and in C3H/HeJ/Dio2−/− mutants with DIO1 and DIO2 deficiency (294–296).
The role of DIO2 in bone was investigated in Dio2−/− mice (60), which have mild pituitary resistance to T4 characterized by a 3-fold increase in TSH, a 27% increase in T4, and normal T3 levels (297, 298). Bone formation and linear growth were normal in Dio2−/− mice, indicating that DIO2 does not have a major role during postnatal skeletal development. This is unexpected, given studies in the chick embryonic growth plate indicating that DIO2 regulates the pace of chondrocyte proliferation and differentiation during early development (159). Although skeletal development is normal, adult Dio2−/− mice have reduced bone formation resulting in a generalized increase in bone mineralization and brittle bones. Target gene analysis demonstrated the phenotype results from reduced T3 production in osteoblasts (60).
Dio3−/− mice have severe growth retardation and increased perinatal mortality. At weaning, they have a 35% reduction in body weight, which persists into adulthood (299). Interpretation of the phenotype, however, is complicated by the systemic effects of disrupted HPT axis maturation and altered thyroid status (300).
In summary, DIO1 has no role in the skeleton; DIO2 is essential for osteoblast function and the maintenance of adult bone structure and strength, whereas the role of DIO3 in bone remains to be determined.
C. Targeting TRα (Figure 7)
Figure 7.

Skeletal phenotype of TRα and TRβ mutant mice. A, Proximal tibias stained with alcian blue (cartilage) and van Gieson (bone, red) showing delayed formation of the secondary ossification center in TRα-deficient mice (TRα0/0) and grossly delayed formation in mice with dominant-negative TRα mutations (TRα1R384C/+ and TRα1PV/+). Mice with mutation or deletion of TRβ have advanced ossification with premature growth plate narrowing. B, Skull vaults stained with alizarin red (bone) and alcian blue (cartilage) showing skull sutures and fontanelles. Arrows indicate delayed intramembranous ossification in mice with dominant-negative TRα mutations (TRα1R384C/+ and TRα1PV/+) and advanced ossification in mice with mutation or deletion of TRβ. C, Trabecular bone microarchitecture in adult TR mutant mice. Backscattered electron scanning electron microscopy images show increased trabecular bone in TRα0/0 mice and severe osteosclerosis in TRα1R384C/+ and TRα1PV/+ mice. By contrast, TRβ mutant mice have reduced trabecular bone volume and osteoporosis. D, Trabecular bone micromineralization in adult TR mutant mice. Pseudo-colored quantitative backscattered electron scanning electron microscopy images showing mineralization densities in which high mineralization density is gray and low density is red. Mice with deletion or mutation of TRα have retention of highly mineralized calcified cartilage (arrows) demonstrating a persistent remodeling defect. By contrast, mice with deletion or mutation of TRβ have reduced bone mineralization (arrow) secondary to increased bone turnover.
Analysis of TR-null, Pax8-null, and TR-knock-in mice has provided further insight into the complexity of T3 actions and the relative roles of TR isoforms. Importantly, TRα1−/− and TRα2−/− mice have selective deletion of TRα1 or TRα2; TRα−/− mice represent an incomplete deletion because the TRΔα1 and TRΔα2 isoforms are still expressed, whereas TRα0/0 mice represent a complete knockout lacking all Thra transcripts (73, 185, 253, 301–303).
TRα1−/− mice retain normal TRα2 mRNA expression (304) and have 30% lower T4 but normal T3 levels and 20% reduced TSH, indicating mild central hypothyroidism. TRα1−/− mice had normal weight gain and linear growth (304). TRα1−/−TRβ−/− double-null mice have 60-fold increases in T4 and T3 with 160-fold higher TSH and decreased GH and IGF-1. Juveniles had growth retardation, delayed endochondral ossification, and decreased bone mineralization, and adults had reduced trabecular and cortical BMD (305–307).
Gene targeting to prevent TRα2 expression resulted in 3- to 5-fold and 6- to 10-fold overexpression of TRα1 mRNA in TRα2+/− and TRα2−/− mice, respectively (253). TRα2−/− mice had 25% lower levels of T4, 20% lower T3, but inappropriately normal TSH, indicating mild thyroid dysfunction. Juvenile TRα2−/− mice had normal linear growth, but adults had reduced trabecular BMD and cortical bone mass (253). Fusion of green fluorescent protein (GFP) to exon 9 of Thra in TRα1-GFP mice unexpectedly resulted in loss of TRα2 expression in homozygotes, with only a 2.5-fold increase in TRα1 mRNA (308). Homozygous TRα1-GFP mice were euthyroid with no abnormalities of postnatal development or growth, suggesting that the phenotype in TRα2−/− mice may result from abnormal overexpression of TRα1. TRα2−/−TRβ−/− double mutants have mild hypothyroidism with a 30% reduction in T4, 20% decrease in T3, and normal TSH, resulting in transiently delayed weight gain (309).
TRα−/− mice are markedly hypothyroid, have severely delayed bone development, and die around weaning unless treated with T3 (73, 310, 311). The skeletal abnormalities include delayed endochondral ossification, disorganized growth plate architecture, impaired chondrocyte differentiation, and reduced bone mineralization. TRα−/−TRβ−/− double-null mice have 10-fold increases in T4 and T3 with 100-fold elevation of TSH and similarly display delayed endochondral ossification and growth retardation (310, 311).
TRα0/0 mice are euthyroid and have less severe skeletal abnormalities than TRα−/− mutants. Juveniles display growth retardation, delayed endochondral ossification, and reduced bone mineral deposition. Although delayed ossification and reduced mineral deposition were observed in juvenile TRα0/0 mice, adults had markedly increased bone mass resulting from a bone-remodeling defect (312). Deletion of both TRs in TRα0/0TRβ−/− double-null mice resulted in a more severe phenotype of delayed bone maturation that may be due to reduced GH/IGF-1 levels or reflect partial TRβ compensation in TRα0/0 single mutants (313).
The role of unliganded TRs in bone was studied in Pax8−/− mice. The severely delayed endochondral ossification in Pax8−/− mice compared to TRα0/0TRβ−/− double mutants indicates that the presence of unliganded receptors is more detrimental to skeletal development than TR deficiency. Importantly, amelioration of the Pax8−/− skeletal phenotype in Pax8−/−TRα0/0 knockout mice, but not Pax8−/−TRβ−/− mutants (314), suggested that unliganded TRα is largely responsible for the severity of the Pax8−/− skeletal phenotype. Nevertheless, this interpretation was not supported by analysis of Pax8−/−TRα1−/− mice in which growth retardation in Pax8−/− mice was not ameliorated by additional deletion of TRα1 (315).
The essential role for TRα in bone has been confirmed by studies of mice with dominant-negative mutations of Thra1. A patient with severe resistance to thyroid hormone (RTH) was found to have a frameshift mutation in the THRB gene, termed “PV.” The mutation results in expression of a mutant TR that cannot bind T3 and acts as a potent dominant-negative antagonist of wild-type TR function (316). The homologous mutation was introduced into the mouse Thra gene to generate TRα1PV mice. The TRα1PV/PV homozygous mutation is lethal, whereas TRα1PV/+ heterozygotes display mild thyroid failure with small increases in TSH and T3, but no change in T4. TRα1PV/+ mice have a severe skeletal phenotype with growth retardation, delayed intramembranous and endochondral ossification, impaired chondrocyte differentiation, and reduced mineral deposition during growth (278, 317, 318). Adult mice had grossly abnormal skeletal morphology with increased bone mass and retention of calcified cartilage, indicating defective bone remodeling (319). Thus, TRα1PV/+ mice have markedly impaired bone development and maintenance despite systemic euthyroidism, indicating that wild-type TRα signaling in bone is impaired by the dominant-negative PV mutation in heterozygous mice. Furthermore, the presence of a dominant-negative TRα leads to a more severe phenotype than receptor deficiency alone.
TRα1R384C/+ mice, with a less potent dominant-negative mutation of TRα, are euthyroid. They display transient growth retardation with delayed intramembranous and endochondral ossification. Trabecular bone mass increased progressively with age, and adults had osteosclerosis due to a remodeling defect (312). Remarkably, brief T3 supplementation during growth, at a dose sufficient to overcome transcriptional repression by TRα1R384C, ameliorated the adult phenotype (312). Thus, even transient relief from transcriptional repression mediated by unliganded TRα1 during development has long-term consequences for adult bone structure and mineralization.
TRαAMI mice harbor a floxed Thra allele (AF2 mutation inducible [AMI]) and express a dominant-negative mutant TRα1L400R only after Cre-mediated recombination. Mice with global expression of TRα1L400R had a similar skeletal phenotype to TRα1PV/+ and TRα1R384C/+ mice (320). Furthermore, restricted expression of TRα1L400R in chondrocytes resulted in delayed endochondral ossification, impaired linear growth, and a skull base defect (321). Microarray studies using chondrocyte RNA revealed changes in expression of known T3-regulated genes, but also identified new target genes associated with cytoskeleton regulation, the primary cilium, and cell adhesion. Desjardin et al (321) also reported that restricted expression of TRα1L400R in osteoblasts unexpectedly resulted in no skeletal abnormalities.
In summary, the presence of an unliganded or mutant TRα is more detrimental to the skeleton than the absence of the receptor. Overall, these studies: 1) demonstrate the importance of thyroid hormone signaling for normal skeletal development and adult bone maintenance; and 2) identify a critical role for TRα in bone.
D. Targeting TRβ (Figure 7)
Two TRβ−/− strains have been generated, and both show similar skeletal phenotypes (9, 285, 311–313). Mice lacking TRβ recapitulate RTH with increased T4, T3, and TSH concentrations (9, 285). Juvenile TRβ−/− mice have accelerated endochondral and intramembranous ossification, advanced bone age, increased mineral deposition, and persistent short stature due to premature growth plate closure. Increased T3-target gene expression demonstrated enhanced T3 action in skeletal cells resulting from the effects of elevated thyroid hormones mediated by TRα in bone. Accordingly, the phenotype is typical of the skeletal consequences of thyrotoxicosis (13, 322). In keeping with the consequences of thyrotoxicosis, adult TRβ−/− mice have progressive osteoporosis with reduced trabecular and cortical bone, reduced mineralization, and increased osteoclast numbers and activity (285, 312).
TRβPV/PV mice with the potent dominant-negative PV mutation have a severe RTH phenotype with a 15-fold increase in T4, a 9-fold increase in T3, and a 400-fold increase in TSH (164, 278, 317, 323). Consequently, TRβPV/PV mice have more severe abnormalities than TRβ−/− mice with accelerated intrauterine growth, advanced endochondral and intramembranous ossification, craniosynostosis, and increased mineral deposition, again resulting in short stature.
Overall, skeletal abnormalities in TRβ mutant mice are also consistent with a predominant physiological role for TRα in bone.
1. Cellular and molecular mechanisms
The opposing skeletal phenotypes in mutant mice provide compelling evidence of distinct roles for TRα and TRβ in the skeleton and HPT axis. The contrasting phenotypes of TRα and TRβ mutant mice can be explained by the predominant expression of TRα in bone (163, 164) and TRβ in the hypothalamus and pituitary (8, 9). Thus, in TRα mutants, delayed ossification with impaired bone remodeling in juveniles and increased bone mass in adults is a consequence of disrupted T3 action in skeletal T3-target cells, whereas accelerated skeletal development and adult osteoporosis in TRβ mutants result from supraphysiological stimulation of skeletal TRα due to disruption of the HPT axis (318).
This paradigm is supported by analysis of T3-target gene expression in skeletal cells, which demonstrated reduced skeletal T3 action in TRα mutant animals (255, 278, 285, 312) and increased T3 action in TRβ mutant mice (164, 224, 285, 312). The studies further demonstrate that T3 exerts anabolic actions during skeletal development but catabolic actions in adult bone. The data also indicate that effects of disrupted or increased T3 action in bone predominate over skeletal responses to TSH. For example, TRα0/0, TRα1PV/+, and TRα1R384C/+ mice have grossly increased trabecular bone mass even when TSH concentrations are normal, whereas TRβ−/− and TRβPV/PV mice are osteoporotic despite markedly increased levels of TSH. Consistent with this conclusion, TRα1−/−β−/− double knockout mice, generated in a different genetic background, also display reduced bone mineralization despite grossly elevated TSH (305).
Although TRα is the major TR isoform expressed in bone, it is apparent that TRα0/0TRβ−/− mice have a more severe skeletal phenotype than TRα0/0 mice, whereas TRα0/0 mice also remain sensitive to T4 treatment, suggesting a compensatory role for TRβ in skeletal cells. This analysis is supported by studies of thyroid-manipulated adult TRα0/0 and TRβ−/− mice, in which a discrete role for TRβ in mediating remodeling responses to T3 treatment was proposed (167).
2. Downstream signaling responses in skeletal cells
GH receptor and IGF-1 receptor mRNA was reduced in growth plate chondrocytes in TRα0/0 and TRα1PV/+ mice, and phosphorylation of their secondary messengers signal transducer and activator of transcription 5 (STAT5) and protein kinase B (AKT) was also impaired (278, 312). By contrast, GH receptor and IGF-1 receptor expression and downstream signaling were increased in TRβ−/− and TRβPV/PV mice (278, 285). Furthermore, studies in osteoblasts from knockout mice revealed that T3/TRα1 bound to a response element in intron 1 of the Igf1 gene to stimulate transcription (259). These studies demonstrate that GH/IGF-1 signaling is a downstream mediator of T3 action in the skeleton in vivo.
Activation of FGFR1 stimulates osteoblast proliferation and differentiation (324), and FGFR3 regulates growth plate chondrocyte maturation and linear growth (325). Fgfr3 and Fgfr1 expression was reduced in growth plates of TRα0/0, TRα1R384C/+, and TRα1PV/+ mice, and Fgfr1 expression was reduced in osteoblasts from TRα0/0 and Pax8−/− mice (138, 224, 255, 278, 285, 312). By contrast, Fgfr3 and Fgfr1 expression was increased in growth plates of TRβ−/− and TRβPV/PV mice, and Fgfr1 expression was increased in osteoblasts from TRβPV/PV mice (164, 224, 285, 312). Thus, FGF/FGFR signaling is a downstream mediator of T3 action in chondrocytes and osteoblasts in vivo.
T3 is essential for cartilage matrix synthesis, and heparan sulfate proteoglycans (HSPGs) are key matrix components essential for FGF and IHH signaling. Studies in thyroid-manipulated rats and TRα0/0TRβ−/− and Pax8−/− mice demonstrated reduced HSPG expression in thyrotoxic animals, increased expression in TRα0/0TRβ−/− mice, and more markedly increased expression in hypothyroid rats and congenitally hypothyroid Pax8−/− mice (219). Thus, T3 coordinately regulates FGF/FGFR and IHH/PTHrP signaling within the growth plate via regulation of HSPG synthesis.
In neonatal TRβPV/PV mice, Runx2 expression was increased in perichondrial cells surrounding the developing growth plate, and in 2-week-old mice Rankl expression was decreased in osteoblasts (252). Although the findings are consistent with increased canonical Wnt signaling pathway during postnatal growth, expression of Wnt4 was decreased. Unliganded TRβ physically interacts with and stabilizes β-catenin to increase Wnt signaling, but this interaction is disrupted by T3 binding. However, although TRβPV similarly interacts with β-catenin, its interaction is not disrupted by T3 (326), and this leads to persistent activation of Wnt signaling in TRβPV/PV mice despite reduced expression of Wnt4 (252). Tsourdi et al (327) investigated Wnt signaling in hypothyroid and thyrotoxic adult mice. In hyperthyroid animals, bone turnover was increased and, consistent with this, the serum concentration of the Wnt inhibitor DKK1 was decreased. In hypothyroid mice, bone turnover was reduced, and the DKK1 concentration increased. Surprisingly, however, concentrations of another Wnt inhibitor, sclerostin, were increased in both hyperthyroid and hypothyroid mice (327). These preliminary studies suggest that increased Wnt signaling may also lie downstream of T3 action in bone.
VI. Skeletal Consequences of Mutations in Thyroid Signaling Genes in Humans (Table 2)
Table 2.
Skeletal Phenotype in Individuals With TSHB, TSHR, SECISBP2, THRA, and THRB Mutations
| Mutation | Genotype | Systemic Thyroid Status | Skeletal Phenotype | Refs. |
|---|---|---|---|---|
| TSHB loss-of-function (non-goitrous congenital hypothyroidism type 4; OMIM no. 275100) | ||||
| TSHBQ49X/Q49X | Premature stop | Low T4, T3, and TSH | T4 replacement from birth | 328 |
| TSH deficiency | BMD normal in two children at 7 y and 10 y | |||
| TSHR loss-of-function (non-goitrous congenital hypothyroidism type 1; OMIM no. 275 200) | ||||
| TSHRP52A/I167N | 1) Impaired function | Normal T4 | Normal growth and bone age (14 y) | 334 |
| 2) Severely impaired function | Elevated TSH 20× | (Without T4 treatment) | ||
| TSHRR109Q/W546X | 1) Impaired function | Normal T4 | Normal growth and bone maturation (3 y) | 333 |
| 2) Frameshift/premature stop | Elevated TSH 50× | (Without T4 treatment) | ||
| TSHRA553T/A553T | Reduced cell surface expression | Hypoplastic thyroid | Normal birth length and head circumference | 331 |
| Low T4 | Normal growth (3 y) | |||
| Elevated TSH 250× | (T4 treatment) | |||
| TSHRC390W/ fs419X | 1) Impaired function | Hypoplastic thyroid | Normal linear growth and bone age (5 y) | 332 |
| 2) Frameshift/premature stop | Low T4 | (T4 treatment) | ||
| Elevated TSH 40× | ||||
| TSHRIVS6+3 G>C/fs656X | 1) Skipping of exon 6 | Severe hypoplasia | Delayed bone age/enlarged fontanelles at birth | 336 |
| 2) Frameshift/premature stop | Very low T4 | Normal growth (2 y) | ||
| Elevated TSH 200× | (T4 treatment) | |||
| TSHRIVS5-1G>A/IVS5-1G>A | Skipping of exon 6 | Severe hypoplasia | Enlarged fontanelles at birth | 335 |
| Very low T4 and T3 | Normal growth (12 y) | |||
| Elevated TSH 250× | (T4 treatment) | |||
| TSHR gain-of-function (non-autoimmune hyperthyroidism; OMIM no. 609152) | ||||
| TSHRM453T/+ | Constitutive activation of cAMP | Goiter | Advanced bone age | 539 |
| High T4 and T3 | Normalized after treatment (12 y) | |||
| Suppressed TSH | ||||
| TSHRM453T/+ | Constitutive activation of cAMP | Goiter | Advanced bone age (newborn) | 339 |
| High T4 and T3 | ||||
| Suppressed TSH | ||||
| TSHRS505N/+ | Constitutive activation of cAMP | High T4 and T3 | Bone age advanced by 4 y at 6 mo old | 341 |
| Suppressed TSH | Craniosynostosis | |||
| Treatment improved growth and bone age (4 y) | ||||
| TSHRT632I/+ | Constitutive activation of cAMP | High T4 and T3 | Craniosynostosis | 344 |
| Suppressed TSH | ||||
| TSHRF629L/+ | Constitutive activation | Goiter | Advanced bone age | 338 |
| High T4 and T3 | Craniosynostosis | |||
| Suppressed TSH | Development normal after treatment (10 y) | |||
| TSHRR528H/S281N | 1) Polymorphism | High T4 and T3 | Birth length 90th centile | 343 |
| 2) Constitutive activation of cAMP | Suppressed TSH | Craniosynostosis requiring surgery | ||
| TSHRV597L/+ | Constitutive activation of cAMP | High T4 and T3 | Advanced bone age (9 mo) | 340 |
| Suppressed TSH | ||||
| TSHRM453T/+ | Constitutive activation of cAMP | Goiter | Bone age advanced by 5 y | 342 |
| High T4 and T3 | Craniosynostosis and mid-face hypoplasia | |||
| Suppressed TSH | Shortened 5th metacarpals and phalanges | |||
| TSHRT32I/+ | Constitutive activation | High T4 and T3 | Bone age advanced by 4 y | 345 |
| Suppressed TSH | Craniosynostosis requiring surgery | |||
| Treatment stabilized bone age | ||||
| (Height: 90th centile at 6 mo but 18th centile at 4 y) | ||||
| SECISBP2 loss-of-function (Abnormal thyroid hormone metabolism OMIM: 609698) | ||||
| SECISBP2R540Q/R540Q | Hypomorphic allele with abnormal SECIS binding | High T4 and rT3, low T3 | Delayed growth and bone age | 346 |
| Slightly elevated TSH | Normal final height | |||
| SECISBP2K438X/IVS8DS+29G-A | 1) LoF truncation | High T4 and rT3, low T3 | Transient growth retardation | 346 |
| 2) Splicing defect | Slightly elevated TSH | |||
| SECISBP2R128X/R128X | Truncated protein lacking N-terminal | High T4 and rT3, low T3 Normal TSH |
Growth retardation and delayed bone age Responsive to T3 treatment |
348 |
| SECISBP2R120X/R770X | 1) LoF truncation | High T4 and rT3, low T3 | Intrauterine growth retardation | 347 |
| 2) Impaired SECIS binding | Slightly elevated TSH | Craniofacial dysmorphism | ||
| Bilateral clinodactyly | ||||
| Delayed growth and bone age | ||||
| SECISBP2fs255X/ fs | 1) Frameshift/premature stop | High T4 and rT3 | Genu valgus | 349 |
| 2) Splicing defect | Normal T3 | External rotation of the hip | ||
| Normal TSH | Normal final height | |||
| SECISBP2C691R/fs | 1) Increased proteasomal degradation 2) Splicing defect |
High T4 and rT3 Low T3 Normal TSH |
Delayed growth T3 treatment improved growth |
349 |
| SECISBP2M515fs563X/Q79X | 1) Frameshift/premature stop | High T4 | Delayed growth and bone age | 350 |
| 2) Premature stop | Slightly low T3 | GH improved height but not bone age | ||
| Slightly elevated TSH | GH+T3 normalized height and bone age | |||
| RTHβ: THRB dominant-negative mutations (non-goitrous congenital hypothyroidism type 6; OMIM no. 614450) | ||||
| TRβ?/+ | Unknown | Goiter | Delayed bone maturation | 351 |
| High T4, T3 | Stippled epiphyses | |||
| Nonsuppressed TSH | ||||
| TRβ?/+ | Unknown | Goiter | Normal height and body proportions | 360 |
| High T4, T3 | Normal head circumference | |||
| Nonsuppressed TSH | Normal bone age and epiphyses (8 y) | |||
| TRβ?/+ | Unknown | High T4, T3 and TSH | Short stature and delayed bone age | 540 |
| TRβ?/+ | Unknown | Goiter | Short stature | 377 |
| High T4, T3 | Bone age 3 y at 2.9 y old | |||
| Nonsuppressed TSH | Craniosynostosis (9 y) | |||
| Shortened 4th metacarpals and metatarsals | ||||
| TRβG345R/+ | No T3 binding | Goiter | Normalizing T4 slowed growth | 352 |
| High T4, T3 | ||||
| Nonsuppressed TSH | ||||
| TRβΔT337/ΔT337 | Dominant-negative TRβ | High T4, T3 | Growth retardation | 368, 370 |
| Homozygous single amino acid deletion | Markedly elevated TSH | Delayed bone age | ||
| No T3 binding | ||||
| TRβ−/− | Deletion of TRβ coding region | High T4, T3 | Stippled epiphyses and growth delay | 541 |
| Nonsuppressed TSH | ||||
| TRβR438H/+ | Dominant-negative TRβ | Goiter | Delayed bone age | 542 |
| Missense mutation | High T4 and TSH | |||
| TRβC434X/+ | Dominant-negative TRβ | High T4, T3 | Advanced bone age | 376 |
| C-terminal premature stop | Nonsuppressed TSH | |||
| No T3 binding | ||||
| TRβR338L/+ | Dominant-negative TRβ | High T4, T3 | Reduced BMD | 375 |
| Missense mutation | Nonsuppressed TSH | |||
| TRβM310L/+ | Dominant-negative TRβ | Goiter | Intrauterine growth retardation | 366 |
| Missense mutation | High T4, T3 | Delayed bone age | ||
| Reduced T3 affinity | Nonsuppressed TSH | Persistent short stature (17 y) | ||
| TRβF438fs422X/+ | Potent dominant-negative TRβ | Small goiter | Growth retardation and delayed bone age | 367 |
| Frameshift | Very high T4, T3 | Short stature | ||
| C-terminal premature stop | High TSH | Frontal bossing | ||
| No T3 binding | ||||
| TRβK443fs458X/+ | Potent dominant-negative TRβ | High T4, T3 and TSH | Short stature (7 y) | 316 |
| Frameshift | ||||
| C-terminal premature stop | ||||
| No T3 binding | ||||
| TRβR429W/+ | Normal T3-binding affinity and transactivation | Goiter High T4, T3 Nonsuppressed TSH |
Increased osteocalcin and decreased BMD (19 y) | 378 |
| TRβG344A/+ | Dominant-negative TRβ | High T4 | Mild intrauterine growth retardation | 364, 543 |
| Missense mutation | Nonsuppressed TSH | Persistent short stature | ||
| No T3 binding | Delayed bone age | |||
| TRβG347A/+ | Missense mutation | Goiter | Persistent short stature (21 y) | 379 |
| High T4, T3 | Increased bone turnover markers | |||
| Nonsuppressed TSH | ||||
| TRβA317T/+; TRβR438H/+; TRβP453T/+; TRβP453fs463X/+; TRβN331H/+ | Missense mutations Frameshift C-terminal premature stop |
High T4, T3 Nonsuppressed or high TSH |
Reduced whole body and lumbar spine BMD in adults but not in children | 374 |
| RTHα: THRA dominant-negative mutations (nongoitrous congenital hypothyroidism type 6; OMIM no. 614450) | ||||
| TRα1E403X/+ | Dominant-negative TRα1 | Low fT4, normal fT3 | Disproportionate short stature (6 y) | 382 |
| C-terminal premature stop | Low fT4:fT3 ratio Low rT3 Normal TSH |
Epiphyseal dysgenesis and grossly delayed bone age Macrocephaly and patent skull sutures Delayed tooth eruption, defective bone mineralization |
||
| TRα1F397fs406X/+ | Dominant-negative TRα1 | Low fT4 and high fT3 | Delayed bone age and persistent short stature (3 y) | 384, 385 |
| Frameshift | Low fT4:fT3 ratio | Macrocephaly/flattened nasal bridge/patent skull sutures | ||
| C-terminal premature stop | Low rT3 | Delayed tooth eruption and congenital hip dislocation | ||
| Normal TSH | Brief increased growth after T4 treatment | |||
| Persistent short stature and otosclerosis (47 y) | ||||
| TRα1A382Pfs389X/+ | Dominant-negative TRα1 | Normal fT4 and fT3 | Disproportionate persistent short stature (45 y) | 383 |
| Frameshift | Low fT4:fT3 ratio | Macrocephaly and cortical bone thickening | ||
| C-terminal premature stop | Low rT3 | T4 treatment (10–15 y) increased growth rate | ||
| Borderline elevated TSH | ||||
| TRα1C392X/+ | C-terminal premature stop | Low fT4 High fT3 Low fT4:fT3 ratio Normal TSH |
Delayed growth short stature with shortened limbs Macrocephaly/flattened nasal bridge/hypertelorism Micrognathia/elongated thorax/lumbar kyphosis (18 y) T4 treatment did not improve syndrome |
388 |
| TRα1E403X/+ | Dominant-negative TRα1 | Low normal fT4 | Delayed growth short stature with shortened limbs | 388 |
| C-terminal premature stop | High normal fT3 Low fT4:fT3 ratio Normal TSH |
Macrocephaly/flattened nasal bridge/hypertelorism Elongated thorax/lumbar kyphosis (14 y) T4 treatment did not improve syndrome |
||
| TRα1E403K/+ | Missense mutation | Low normal fT4 High normal fT3 Low fT4:fT3 ratio Normal TSH |
Short stature with shortened limbs Macrocephaly/flattened nasal bridge/hypertelorism Elongated thorax (12 y) T4 treatment did not improve syndrome |
388 |
| TRα1P398R/+ | Missense mutation | Low normal fT4 | Delayed growth with shortened limbs | 388 |
| High normal fT3 | Flattened nasal bridge/hypertelorism | |||
| Low fT4:fT3 ratio | Elongated thorax (8 y) | |||
| Low normal TSH | T4 treatment did not improve syndrome | |||
| TRαA263V/+ | Weak dominant-negative | Low normal fT4 | Growth retardation | 389 |
| Missense mutation | High normal fT3 | Macrocephaly/broad face/flattened nasal bridge | ||
| Transactivation activity restored by supraphysiological T3 | Low fT4:fT3 ratio | Thickened calvarium | ||
| No effect on TRα2 action | Low rT3 | T4 treatment improved growth rate | ||
| Normal TSH | Macrocephaly and increased BMD in adult | |||
| TRαN359Y/+ | Dominant-negative TRα1 Missense mutation with decreased T3 binding and transactivation activity Weak reduction in TRα2 action |
Normal fT4 High normal fT3 Low fT4:fT3 ratio Low normal rT3 Low TSH (initially normal) |
Growth retardation with shortened limbs Macrocephaly/hypertelorism/micrognathia/broad nose Elongated thorax with clavicular and 12th rib agenesis with scoliosis, ovoid vertebrae, and congenital hip dislocation Humeroradial synostosis and syndactyly |
390 |
Abbreviation: LoF, loss of function.
A. TSHB
1. Loss-of-function mutations
Several individuals have been described with mutations of TSHB leading to biologically inactive TSH and congenital nongoitrous hypothyroidism (OMIM no. 275100), but skeletal consequences have only been documented in two cases. Two brothers aged 10 and 7 years were described with isolated TSHβ deficiency and normal BMD after thyroid hormone replacement from birth (328). These cases demonstrate that the absence of TSH throughout normal skeletal development and growth does not affect bone mineral accumulation by 10 years of age.
B. TSHR
1. Loss-of-function mutations
Loss-of-function mutations in TSHR have been described at more than 30 different amino acid positions and result in varying degrees of TSH resistance and congenital nongoitrous hypothyroidism (OMIM no. 275200) (329, 330). However, skeletal consequences have only been reported in a few children. Individuals with mild and compensated hypothyroidism have normal growth and bone age (331–334), whereas subjects with severe thyroid hypoplasia and very low T4 and T3 levels have delayed intramembranous ossification and bone age at birth but display normal growth and postnatal skeletal development after T4 replacement (335, 336). These cases demonstrate that impaired TSHR signaling does not impair linear growth and bone maturation when thyroid hormones are adequately replaced.
2. Gain-of-function mutations
Nonautoimmune autosomal dominant hyperthyroidism (OMIM no. 609152) is rare, and patients are frequently treated at an early age with thyroid surgery and radioiodine ablation followed by physiological T4 replacement (337). Skeletal manifestations at presentation include classical features of juvenile hyperthyroidism, with advanced bone age (338–342), craniosynostosis (338, 342–345), shortening of the fifth metacarpal bones and middle phalanges of the fifth fingers (342) all described. The skeletal consequences in treated adults have not been reported, but in several younger patients, amelioration of the phenotype was reported after treatment and normalization of circulating thyroid hormone levels (338, 341, 344, 345). These cases indicate that early normalization of thyroid status improves skeletal abnormalities in patients with nonautoimmune autosomal dominant hyperthyroidism despite continued constitutive activation of the TSHR.
C. SBP2
1. Loss-of-function mutations
The deiodinases are selenoproteins that require incorporation of the rare amino acid selenocysteine (Sec) for enzyme activity. Individuals with loss-of-function mutations of SBP2, which encodes the Sec incorporation sequence binding protein 2 (SBP2) essential for incorporation of Sec, have abnormal thyroid hormone metabolism resulting in low circulating total and fT3 levels despite elevated total and fT4 and rT3 levels (OMIM no. 609698). Affected patients have evidence of skeletal thyroid hormone deficiency during prepubertal growth with transient growth retardation, short stature, and delayed bone age (346–349) that cannot be compensated by treatment with GH (350).
D. THRB
1. Dominant-negative mutations
RTH, an autosomal dominant condition caused by heterozygous dominant-negative mutations of the THRB gene encoding TRβ (OMIM no. 188570; RTHβ), was described in 1967 (351), with the first mutations identified in 1989 (352, 353). The incidence of RTHβ is 1 in 40 000; over 3000 cases have been documented, with 80% harboring THRB mutations, of which 27% are de novo (354). The mutant TRβ disrupts negative feedback of TSH secretion, resulting in the characteristic elevation of T4 and T3 concentrations and inappropriately normal or increased TSH. The syndrome results in a complex mixed phenotype of hyperthyroidism and hypothyroidism, depending on the specific mutation and target tissue. Thus, an individual patient can have symptoms and signs of both thyroid hormone deficiency and excess. Tissue responses to T3 are dependent upon: the severity of the TRβ mutation; genetic background; the relative concentrations of TRα, wild-type TRβ, and dominant-negative mutant TRβ proteins in individual cells; and whether the patient has received prior treatment with antithyroid drugs or surgery. Consequently, interpretation of the skeletal phenotype in RTHβ is complex, and a broad range of abnormalities has been described.
The skeletal consequences of RTHβ have been reported in a limited number of patients with a spectrum of mutations. Abnormalities include major or minor somatic defects, such as scaphocephaly, craniosynostosis, bird-like facies, vertebral anomalies, pigeon breast, winged scapulae, prominent pectoralis, and short fourth metacarpals (355). The findings are inconsistent, and factors other than mutations of TRβ may be responsible (356). In kindreds with the same THRB mutation, individuals may have different phenotypes, especially with respect to growth velocity or the degree of abnormality in thyroid status. Thus, variable skeletal abnormalities have been reported in three kindreds (357), in 14 affected individuals within a four-generation kindred (358), and in four unrelated families with affected individuals aged 14 months to 29 years (359), whereas a normal skeletal phenotype was reported in an affected 8 year old (360).
Some subjects have been reported with skeletal abnormalities that are similar to the consequences of hypothyroidism. Delayed growth below the fifth percentile has been estimated in 20% of subjects (361), whereas delayed bone age of greater than 2 SD was reported to occur in 29–47% (362). A National Institutes of Health study of 104 patients from 42 kindreds and 114 unaffected relatives found short stature in 18%, low weight for height in 32% of children, and variable tissue resistance from kindred to kindred. RTHβ patients were shorter than unaffected family members and had no catch-up growth later in life. However, delayed bone age was documented in only a minority (363). In a study of 36 family members with RTHβ in four generations, delayed bone maturation with short stature was observed in affected adults and children. Of note, in affected individuals born to an affected mother, growth retardation was less marked. Seven of eight children with RTHβ, but only one of six controls, had bone age 2 SD below the mean (364). Delayed bone maturation was also found in a series of eight children (365). In individual cases, stippled epiphyses were reported in a 6 year old (351), intrauterine and postnatal growth retardation with delayed bone age in a 26-month-old girl (366), and delayed bone age >3 SD below the mean for chronological age in a 22 month old (367). Four patients with homozygous THRB mutations have been identified, and markedly delayed linear growth and skeletal maturation were reported (368–370).
Other individuals with RTHβ, however, have been reported with skeletal abnormalities that are similar to the consequences of hyperthyroidism. Short metacarpals, metatarsals, and advanced bone age have all been described (371), as well as craniofacial abnormalities, craniosynostosis, advanced bone age, short stature, osteoporosis, and fracture, together with increased bone turnover and changes in calcium, phosphorous, and magnesium metabolism (364, 372, 373). In 14 patients, including eight adults and six children, the affected children had short stature below the third centile, no significant delay in bone age, increased calcium, decreased phosphate, and elevated FGF23. Adults had low lumbar spine and whole body BMD (374). Five adults from two unrelated kindreds with the same THRB mutation were followed for 3–11 years and were found to have reduced BMD (375). In individual cases, a 15-year-old girl with short stature and advanced bone age equivalent to age 17 years (376) was reported; craniosynostosis with frontal bossing and minor skeletal abnormalities including short metacarpals were seen in a 9 year old (377); a 19-year-old man with increased osteocalcin and mildly reduced BMD was reported (378); and a 21-year-old female with short stature, normal alkaline phosphatase, but elevated urinary deoxypyridinoline was described (379).
Overall, these findings should be considered in the context of data from mouse models of RTHβ. In mutant mice, the skeletal phenotype is consistent with increased thyroid hormone action in bone manifest by accelerated skeletal development in juveniles and increased bone turnover with osteoporosis in adults (318). Importantly, the clear and consistent findings in mutant mice result from studies of single mutations in genetically homogeneous backgrounds that are not confounded by therapeutic intervention. Reports in human RTHβ result from patients with a broad range of mutations of varying severity studied in diverse genetic backgrounds. Phenotype diversity is also likely to be due to additional confounding factors, including variable phenotype analyses and nomenclature among studies, retrospective and cross-sectional study design, differing surgical and pharmacological interventions, and analysis of individuals at differing ages. Well-controlled, long-term prospective analysis of large families with specific mutations will be the only way to determine the skeletal consequences of individual mutations in human RTHβ.
E. THRA
1. Dominant-negative mutations
Individuals with mutations of THRA encoding TRα were recently identified (OMIM no. 614450; RTHα), and this has resulted in reclassification of the inheritable syndromes of impaired sensitivity to thyroid hormone (380). By analogy to the phenotype variability seen in RTHβ, it was considered likely that individuals with a spectrum of THRA mutations would be identified (381). Heterozygous mutations of THRA were first reported in three families in 2012 and 2013 (382–385). Affected individuals all had grossly delayed skeletal development but variable motor and cognitive abnormalities. All had normal serum TSH with low/normal T4 and high/normal T3 concentrations, and a characteristically reduced fT4:fT3 ratio (369, 386, 387).
2. Mutations affecting TRα1
A 6-year-old girl had skeletal dysplasia, growth retardation with grossly delayed bone age and tooth eruption, patent skull sutures with wormian bones, macrocephaly, flattened nasal bridge, disproportionate short stature (decreased subischial leg length with a normal sitting height), epiphyseal dysgenesis, and defective bone mineralization (382). A 3-year-old girl displayed a similar phenotype with short stature, macrocephaly, delayed closure of the skull sutures, delayed tooth eruption, delayed bone age with absent hip secondary ossification centers and congenital hip dislocation. Treatment with T4 resulted in a brief initial catch-up of growth, whereas GH treatment had no effect. Her subsequent height, however, remained 2 SD below normal. Her 47-year-old father was short (−3.77 SD), but with normal BMD, and he had hearing loss due to otosclerosis (384, 385). A 45-year-old female was subsequently described with disproportionate short stature and macrocephaly (head circumference +9 SD), together with skull vault and long-bone cortical thickening. Between the ages of 10 and 15 years, T4 treatment increased her rate of growth, although final adult height was 2.34 SD below predicted (383). Most recently, a series of four individuals with truncating and missense mutations affecting TRα1 was described and included data from 18 years of follow-up (388). A consistent skeletal phenotype included disproportionate growth retardation with relatively short limbs, hands, and feet and a long thorax, and a mild skeletal dysplasia comprising a flat nasal bridge with round, puffy, and coarse flattened facial features, upturned nose, hypertelorism, and macrocephaly. Radiological features of hypothyroidism included: ovoid immature vertebral bodies, ossification defects of lower thoracic and upper lumbar bodies, and hypoplasia of the acetabular and supra-acetabular portions of the ilia, and coxa vara. Hand x-rays showed changes in the tubular bones, which were short, wide, and morphologically abnormal. A phenotype-genotype correlation was noted, with missense mutations associated with a milder phenotype than the more severe phenotype seen in individuals with THRA truncation mutations. T4 treatment had no effect in any patient (388).
3. Mutations affecting both TRα1 and TRα2
A 60-year-old woman presented in childhood with growth failure, macrocephaly, broad face, flattened nasal bridge, and a thickened calvarium (389). Her growth improved after T4 treatment during childhood following a radiological opinion suggesting that the skeletal features were similar to hypothyroidism. Her 30- and 26-year-old sons presented similarly and were also treated during childhood, resulting in a good growth response. Nevertheless, they each had a persistently abnormal facial appearance and macrocephaly, together with increased BMD between +0.8 and +1.9 SD. In vitro studies demonstrated that the mutant TRα1 was a weak dominant-negative antagonist, but its transcriptional activity could be restored by supraphysiological concentrations of T3, whereas the function of the mutant TRα2 protein did not differ from wild-type (389). Recently, a 27-year-old female with a grossly abnormal skeletal phenotype and low fT4:fT3 ratio was described in association with a N359Y mutation affecting both TRα1 and TRα2 (390). Her phenotype included intrauterine growth retardation and failure to thrive, macrocephaly, hypertelorism, micrognathia, short and broad nose, clavicular and 12th rib agenesis, elongated thorax, ovoid vertebrae, scoliosis, congenital hip dislocation, short limbs, humeroradial synostosis, and syndactyly (390). This severe and atypical phenotype extends the abnormalities described in patients with THRA mutations and raises questions regarding whether TRα2 has a functional role in skeletal development or whether the RTHα phenotype is more variable and extensive than previously reported.
Together, these reports define a new genetic disorder, RTHα, characterized by profound and consistent developmental abnormalities of the skeleton that recapitulate findings in mice with dominant-negative Thra mutations (278, 312, 319–321, 391). Initially identified THRA mutations resulted in severe phenotypes due to expression of truncated TRα1 proteins with no T3-binding capability but potent dominant-negative activity (382–385). Recognition of individuals with milder mutations and phenotypes has been predicted to be more difficult because disruption of THRA does not affect the HPT axis significantly (319).
VII. Thyroid Status and Skeletal Development
A. Consequences of hypothyroidism
Hypothyroidism is the most common congenital endocrine disorder, with an incidence of 1 in 1800. Congenital and juvenile acquired hypothyroidism result in delayed skeletal development and bone age with short stature. Abnormal endochondral ossification and epiphyseal dysgenesis are evidenced by “stippled epiphyses” on x-rays. In severe or undiagnosed cases (Figure 8), there is complete postnatal growth arrest and cessation of bone maturation with a complex skeletal dysplasia that includes broad flat nasal bridge, hypertelorism, broad face, patent fontanelles, scoliosis, vertebral immaturity and absence of ossification centers, and congenital hip dislocation (392). Thyroid hormone replacement induces rapid “catch-up” growth and accelerated skeletal maturation, demonstrating the exquisite sensitivity of the developing skeleton to thyroid hormones. Nevertheless, predicted adult height may not be attained, and in such cases, the final height deficit is related to the duration and severity of hypothyroidism before diagnosis and treatment (281), although the underlying mechanisms remain unknown. Overall, most children with congenital hypothyroidism that are treated early with T4 replacement ultimately reach their predicted adult height and achieve normal BMD after 8.5 years of follow-up (393, 394).
Figure 8.

X-rays of a 36-year-old female with severe untreated congenital hypothyroidism. (X-rays were kindly provided by Dr. Jonathan LoPresti, Keck School of Medicine, University of Southern California). A, Lateral and anteroposterior skull images showing persistently patent sutures and fontanelles and delayed tooth eruption. B, Lower limb x-ray showing severe epiphyseal dysgenesis with grossly delayed formation of secondary ossification centers (arrows). C, Hand x-ray demonstrating a 35-year delay in bone maturation (bone age, 14 months). D, Bone age advanced by 8 years after T4 replacement for a period of only 18 months, demonstrating rapid acceleration of endochondral ossification and “catch-up growth.”
B. Consequences of thyrotoxicosis
During skeletal development and growth, T3 regulates the pace of chondrocyte differentiation in the epiphyseal growth plates (120, 159, 161, 162, 224). Graves' disease is the commonest cause of thyrotoxicosis in children but remains rare. In contrast to hypothyroidism during childhood, juvenile thyrotoxicosis is characterized by accelerated skeletal development and rapid growth, although advanced bone age results in early cessation of growth and persistent short stature due to premature fusion of the growth plates. In severe cases in young children, early closure of the cranial sutures can result in craniosynostosis (13, 322, 345), whereas maternal hyperthyroidism per se may also be a risk factor for craniosynostosis (395). These observations demonstrate further the marked sensitivity of the developing skeleton to the actions of thyroid hormone.
VIII. Thyroid Status and Bone Maintenance
Numerous studies have investigated the consequences of altered thyroid function on BMD and fracture risk in adults (396–399). Interpretation is difficult because studies are frequently confounded by inclusion of subjects with a variety of thyroid diseases and comparison of cohorts that include combinations of pre- and postmenopausal women or men (400–405). Many studies lack statistical power because of small numbers, cross-sectional design, or insufficient follow-up (401, 406–411). Some studies measure TSH but not thyroid hormones, whereas others determine thyroid hormones but not TSH (402, 408). Other problems include inadequate control for confounding factors including: age; prior or family history of fracture; body mass index; physical activity; use of estrogens, glucocorticoids, bisphosphonates, or vitamin D; prior history of thyroid disease or use of T4; and smoking or alcohol intake.
Different methods have also been used for skeletal assessment, including analysis of bone geometry in some studies (411, 412). Regulation of bone turnover has been investigated by histomorphometry in early studies (413–416), measurement of proinflammatory cytokines (417), and a variety of biochemical markers of bone formation (serum alkaline phosphatase, osteocalcin, carboxy-terminal propeptide of type 1 collagen) and resorption (urinary pyridinoline and deoxypyridinoline collagen cross-links, hydroxyproline, carboxy-terminal cross-linked telopeptide of type 1 collagen, cathepsin K), which are all elevated in hyperthyroidism and generally correlate with disease severity (401, 408, 418–422). Overall, hyperthyroidism shortens the bone remodeling cycle in favor of increased resorption (Figure 5). This results in a high bone turnover state with increased bone resorption and formation rates. The frequency of initiation of bone remodeling is also increased. The duration of bone formation and mineralization is reduced to a greater extent than the duration of bone resorption. This leads to reduced bone mineralization, a net 10% loss of bone per remodeling cycle, and osteoporosis (415). BMD has also been determined by several methods including dual x-ray absorptiometry, single-photon absorptiometry, dual-photon absorptiometry, quantitative computed tomography (398), ultrasound (400), and high-resolution peripheral quantitative computed tomography (423). Although many retrospective and cross-sectional studies of the effects of thyroid dysfunction on bone turnover and mass have been reported, only a small number of studies have been prospective, whereas even fewer have investigated fracture susceptibility. Interpretation of the literature is, therefore, difficult and complex.
1. Discriminating thyroid hormone and TSH effects on the skeleton
Studies investigating osteoporosis, fracture, and thyroid hormone excess have been conflicting regarding the relative roles of thyroid hormone and TSH (424). Importantly, this issue cannot be resolved in individuals in whom the HPT axis is intact and the reciprocal relationship between thyroid hormones and TSH is maintained (13). Nevertheless, simultaneously increased thyroid hormone and TSH signaling occurs in three clinical situations: nonautoimmune autosomal dominant hyperthyroidism and RTHβ, as described above, together with Graves' disease, in which TSHR-stimulating antibodies persistently activate the TSHR and increase T4 and T3 production. Conventionally, secondary osteoporosis in Graves' disease is considered to be a consequence of elevated circulating thyroid hormone levels and increased T3 actions in bone. By contrast, TSH has been proposed as a negative regulator of bone remodeling (32), and thus suppressed TSH levels in thyrotoxicosis were suggested to be the primary cause of bone loss. Nevertheless, because Graves' disease is characterized by persistent autoantibody-mediated TSHR stimulation, such patients should be protected. Despite this, Graves' disease remains an established and important cause of secondary osteoporosis and fracture.
The effects of TSH on bone turnover have also been investigated in clinical studies. In patients receiving TSH-suppressive doses of T4 in the management of differentiated thyroid cancer, administration of recombinant human TSH (rhTSH) increases TSH levels to >100 mU/L but does not affect circulating T4 and T3 concentrations because patients have previously undergone total thyroidectomy. In this context, treatment of premenopausal women with rhTSH had no effect on serum markers of bone turnover (425–427). In postmenopausal women, two studies reported a reduction in bone resorption markers accompanied by an increase in bone formation markers after rhTSH administration (426, 427), whereas two studies reported no effect (425, 428).
A. Consequences of variation of thyroid status within the reference range
1. Effect on bone turnover markers
No studies have been reported in premenopausal women. In 3261 postmenopausal women from the Study of Health in Pomerania, higher TSH was associated with increased bone turnover and stiffness, but no association was reported with levels of sclerostin. In 2654 men from the same study, however, TSH was not related to bone turnover, stiffness, or sclerostin. Importantly, T3 and T4 levels were not determined in this study (429). In a study of 60 postmenopausal women, Zofkova and Hill (430) reported a correlation between high TSH and low concentrations of the bone resorption marker urinary deoxypyridinoline, but no relationship with the bone formation marker serum procollagen type I C propeptide, although individuals with treated hypothyroidism, subclinical hyperthyroidism, and secondary hyperparathyroidism were included.
2. Effect on BMD
One study in 2957 euthyroid healthy Taiwanese men and women of varying ages reported a weak inverse correlation between fT4 and BMD but did not identify any relationship between TSH and BMD (431).
A 6-year prospective study of 1278 healthy euthyroid postmenopausal women from five European cities (OPUS study) found that thyroid status at the upper end of the normal range was associated with lower BMD (432). This finding has been supported by a number of cross-sectional studies. Thyroid function in the upper normal reference range was associated with reduced BMD in 1426 healthy euthyroid perimenopausal women (433). A study of 756 euthyroid Korean women aged 65 and older showed that BMD was positively correlated with TSH (434). In 581 postmenopausal American women, TSH at the lower end of the reference range was associated with a 5-fold increased risk of osteoporosis compared to TSH at the upper end of the normal range (435). Kim et al (436) showed that low normal TSH levels were associated with lower BMD in 959 Korean postmenopausal women. In 648 healthy postmenopausal women, higher fT4 levels within the normal range were associated with a relationship with deterioration in trabecular bone microarchitecture (437).
In 1151 euthyroid men and women over the age of 55 years, femoral neck BMD correlated positively with TSH and inversely with fT4 (438). The association with fT4 was much stronger than the association with TSH. A population study of 993 postmenopausal women and 968 men from Tromsø showed that subjects with TSH below the 2.5th percentile had a low forearm BMD, whereas those with TSH above the 97.5th percentile had high femoral neck BMD (439). A study of 677 healthy men aged 25–45 years found that higher fT3, total T3, and total T4 levels were associated with lower BMD (440).
3. Fracture risk
Leader et al (441) investigated 13 325 healthy subjects with at least one TSH measurement during 2004 and demonstrated an increased risk of hip fracture during the subsequent 10 years in euthyroid women with TSH levels in the lower normal range, although no effect was found in men. Svare et al (442) analyzed prospective data from over 16 000 women and nearly 9000 men in the Hunt2 study and found no relationship between baseline TSH and hip or forearm fracture, but there were weak positive associations in women between hip fracture risk and both low and high TSH. In 130 postmenopausal women with normal thyroid function, Mazziotti et al (443) found that TSH levels at the low end of the normal range were associated with an increased prevalence of vertebral fractures in women previously known to have osteoporosis or osteopenia. van der Deure et al (438), however, found no relationship between fT4 or TSH and fracture in 1151 euthyroid men and women over the age of 55 years. In the OPUS study, thyroid status at the upper end of the normal range was associated with an increased risk of incident nonvertebral fracture, which was increased by 20 and 33% in women with higher fT4 or fT3, whereas higher TSH was protective and the fracture risk was reduced by 35% (432). However, in a 10-year prospective study of a cohort of only 367 women, no associations between fT3, fT4, or TSH and incident vertebral fracture were identified (444).
Overall, thyroid status at the upper end of the euthyroid reference range is associated with lower BMD and an increased risk of fracture in postmenopausal women, although studies cannot discriminate the relative contributions of thyroid hormones and TSH.
B. Consequences of hypothyroidism
The effects of hypothyroidism on bone turnover have been investigated by histomorphometry (413, 416). Manifestations include reduced osteoblast activity, impaired osteoid apposition, and a prolonged period of secondary bone mineralization. Consistent with a low bone turnover state, osteoclast activity and bone resorption are also reduced. The effect is a net increase in mineralization without a major change in bone volume, although bone mass may increase as a result of the prolonged remodeling cycle (413, 445). To identify changes in bone mass resulting from hypothyroidism would require long-term follow-up of untreated patients, and data from such studies are not available.
1. Effect on bone turnover markers
No studies have been reported in pre- or postmenopausal women or men.
2. Effect on BMD
Consistent with histomorphometry data, two studies reported normal BMD in patients newly diagnosed with hypothyroidism (446, 447). A cross-sectional cohort study of 49 patients with well-substituted hypothyroidism was compared to 49 age- and sex-matched controls; the study investigated BMD by dual x-ray absorptiometry, bone geometry by pQCT and bone strength by finite element analysis. The study showed no difference between controls and patients receiving T4 (423). A retrospective study of 400 Puerto Rican postmenopausal women also showed no relationship between hypothyroidism and BMD (448). Nevertheless, Paul et al (449) reported a 10% reduction in BMD in the femur but no effect on lumbar spine BMD after T4 replacement in 31 hypothyroid premenopausal women treated for at least 5 years, whereas Kung and Pun (450) reported reduced BMD in 26 hypothyroid premenopausal women receiving T4 for between 1 and 24 years. These studies are difficult to interpret because of the confounding effects of patient compliance to T4 replacement and the likely variability in the adequacy or sustainment of restored euthyroidism during follow-up.
3. Fracture risk
Although the effects on fracture risk of T4 replacement for hypothyroidism have not been investigated directly, retrospective data failed to identify an association (451–453). Nevertheless, large cross-sectional population studies have identified an association between hypothyroidism and fracture. Patients with a prior history of hypothyroidism or increased TSH concentration had a 2- to 3-fold increased relative risk of fracture, which persisted for up to 10 years after diagnosis (447, 454–457). One recent study of T4 treatment of 74 patients with adult-onset hypopituitarism also suggested the possibility that overtreatment of patients with T4 may increase vertebral fracture risk in some patients, particularly those with coexistent untreated GH deficiency (458).
In postmenopausal women, a database cohort study of 11 155 women over age 65 and receiving T4 for hypothyroidism revealed an increased risk of fracture in patients with a prior history of osteoporosis who were receiving more than 150 μg of T4, suggesting that overtreatment is detrimental (459). Nevertheless, González-Rodríguez et al (448) did not identify a relationship between hypothyroidism and fracture in Puerto Rican postmenopausal women.
Overall, these studies suggest that hypothyroidism per se is unlikely to be related to fracture risk, whereas long-term supraphysiological replacement with thyroid hormones may result in an increased risk of fracture.
C. Consequences of subclinical hypothyroidism
1. Effect on bone turnover markers
A randomized controlled trial in a heterogeneous group of 61 patients with subclinical hypothyroidism demonstrated that treatment with T4 to restore euthyroidism resulted in increased bone turnover at 24 and 48 weeks and reduced BMD after 48 weeks (460).
2. Effect on BMD and fracture risk
A study of subclinical hypothyroidism in 4936 US men and women aged 65 years and older followed up for 12 years showed no association with BMD or incident hip fracture (461). A prospective study in men aged 65 and older who were followed for 4.6 years also revealed no association between subclinical hypothyroidism and bone loss (462). A prospective cohort study of 3567 community-dwelling men more than 65 years of age revealed a 2.3-fold increased risk of hip fracture over 13 years of follow-up in men with subclinical hypothyroidism after adjustment for covariates, including the use of thyroid hormones (463).
3. Meta-analysis
Importantly, an individual participant meta-analysis of 70 298 individuals during 762 401 person-years of follow-up found no association between subclinical hypothyroidism and fracture risk (464).
D. Consequences of subclinical hyperthyroidism
1. Effect on bone turnover markers
Management of patients with differentiated thyroid cancer frequently involves prolonged treatment with T4 at doses that suppress TSH and may be detrimental to bone. A few studies investigated the effect of TSH suppression on bone turnover in small numbers of patients, and findings have been inconsistent (398). Some reported increased bone formation and resorption markers in patients receiving T4 (401, 465), whereas others reported no effect on either resorption or formation markers (409, 466).
2. Effect on BMD
Most studies in premenopausal women reported no effect of TSH suppression therapy on BMD at any anatomical site, although one study showed that BMD may be reduced (467). Early studies reporting the effect of TSH suppression on BMD in postmenopausal women were conflicting because they frequently evaluated TSH only without measurement of thyroid hormones and were thus unable to exclude overt hyperthyroidism. Overall, therefore, these heterogeneous studies should be interpreted with caution. For example, Franklyn et al (468) investigated 26 UK postmenopausal women treated for 8 years and found no effect of TSH suppression on BMD, whereas Kung and Yeung (469) studied 46 postmenopausal Asian women and found decreased total body, lumbar spine, and femoral neck BMD. Direct comparison between these studies is not possible, however, because TSH was fully suppressed in only 80% of patients in one (468), mean calcium intake was low in the other (469), while both were performed in small numbers of patients but from differing ethnic backgrounds. Similar conflicting data have been reported at various anatomical sites in many less well-controlled cross-sectional and longitudinal studies (398, 409, 410). Recent data have similarly been contradictory; a 12-year follow-up study of endogenous subclinical hyperthyroidism in 1317 men and women aged 65 years and older showed no association with BMD (461). Despite this, a 1-year prospective study of 93 women with differentiated thyroid cancer showed that TSH-suppressive therapy resulted in accelerated bone loss in postmenopausal women, but only in the early postoperative period after thyroidectomy (403). In men, no association between subclinical hyperthyroidism and BMD was identified in two recent studies (461, 462), which supports overall findings from previous cross-sectional studies, of which only one reported reduced BMD in men receiving TSH-suppressive doses of T4 (470).
3. Fracture risk
In a retrospective population study of 2004 individuals, subclinical hyperthyroidism was associated with a 1.25-fold increased risk of fracture, although no association with TSH concentration was identified, and the relationship between fracture and thyroid hormone levels was not reported (471). Nevertheless, a nested case-control study of 213 511 individuals from Ontario reported a 1.9-fold increased fracture risk in patients treated with T4 and demonstrated a clear dose-response relationship (472). A prospective cohort study in postmenopausal women with case-cohort sampling identified an association between suppressed TSH and increased fracture risk, with 3- to 4-fold increases in both hip and vertebral fractures in individuals with TSH suppressed below 0.01 mU/L (473). A 2.5-fold increased rate of hospital admission for fracture in subjects with TSH less than 0.05 mU/L (455) and an exponential rise in the association between fracture risk and longer duration of TSH suppression (474) have also been reported. Further analysis demonstrated an increased risk of fracture in patients with hypothyroidism that was strongly related to the cumulative duration of periods with a low TSH due to excessive thyroid hormone replacement (475). Similarly, in the Fracture Intervention Trial, Jamal et al (476) analyzed a subgroup of patients with TSH suppressed below 0.5 mU/L and found an increased risk of vertebral fracture, although individuals with untreated thyrotoxicosis were not formally excluded.
Recently, Lee et al (463) demonstrated in a 14-year prospective follow-up study that men, but not women, with endogenous subclinical hyperthyroidism had a 5-fold increased hazard ratio of hip fracture, whereas men with all causes of subclinical hyperthyroidism had a 3-fold increased hazard ratio. Nevertheless, a prospective study in men aged 65 and older revealed no association between subclinical hyperthyroidism and fracture risk during a 4.6-year follow-up, although a weak increased risk of hip fractures in subjects with lower serum TSH was identified (462). No association was seen between hip fracture risk and endogenous subclinical hyperthyroidism in 4936 US men and women aged 65 years and older followed up for 12 years (461).
In summary, large population studies have revealed increased bone turnover, reduced BMD, and an increased risk of fracture, particularly in postmenopausal women with subclinical hyperthyroidism (447, 455, 456, 473). Importantly, a prospective study of low-risk differentiated thyroid cancer patients treated with suppressive doses of T4 demonstrated an increased risk of postoperative osteoporosis without beneficial effect on tumor recurrence, and the authors suggested that future intervention in the long-term treatment of low-risk disease should focus toward avoiding therapeutic harm (477).
4. Systematic reviews and meta-analyses
Levels of bone resorption and formation markers have been reported to be either elevated or normal in subclinical hyperthyroidism. Similarly, BMD has been found to be either reduced or unaffected, and a comprehensive meta-analysis was inconclusive (478). Heemstra et al (397) analyzed 12 cross-sectional and four prospective studies of premenopausal women receiving suppressive doses of T4 but found that a formal meta-analysis could not be performed due to heterogeneity. The authors concluded that suppressive doses of T4 are unlikely to affect BMD in premenopausal women. This conclusion supported an earlier review of eight studies by Quan et al (479). A meta-analysis of 27 studies investigating the effects of TSH suppression on BMD identified no effect in premenopausal women or men but found that suppressive doses of T4 for up to 10 years in postmenopausal women led to reductions in BMD of between 5 and 7% (480). Systematic reviews of effects in postmenopausal women receiving suppressive doses of T4 for approximately 8.5 years suggested an increased rate of annual bone loss that results in a reduction in BMD of 7% at the lumbar spine and 5% at the hip. These reviews recommended monitoring BMD in postmenopausal women with subclinical hyperthyroidism (397, 398, 479). Nevertheless, although a recent systematic review and meta-analysis of seven population-based cohorts also suggested that subclinical hyperthyroidism may be associated with an increased risk of hip and nonspine fracture, a firm conclusion could not be reached due to limitations of the cohorts (481).
Importantly, the largest meta-analysis of 70 298 individuals during 762 401 person-years of follow-up demonstrated an increased risk of hip and other fractures in individuals with subclinical hyperthyroidism, particularly in those with TSH suppressed below 0.1 mU/L and those with endogenous disease (464).
E. Consequences of hyperthyroidism
1. Effect on bone turnover markers and BMD
The effects of thyrotoxicosis on bone turnover are consistent with histomorphometry data. Markers of bone formation and resorption are elevated and correlate with disease severity in pre- and postmenopausal women and men (418–421). In premenopausal women, treatment of thyrotoxicosis resulted in a 4% increase in BMD within 1 year (482).
2. Fracture risk
Severe osteoporosis due to uncontrolled thyrotoxicosis is now rare because of prompt diagnosis and treatment, although undiagnosed hyperthyroidism is an important contributor to secondary bone loss and osteoporosis in patients presenting with fracture (483). The presence of thyroid disease as a comorbidity factor has also been suggested to increase 1- and 2-year mortality rates in elderly patients with hip fracture (484). Several studies have investigated the skeleton in untreated thyrotoxicosis or have analyzed population cohorts to determine the relationship between hyperthyroidism and fracture. One cross-sectional study (456) and four population studies identified an association between fracture and a prior history of thyrotoxicosis. These studies could not determine whether reduced BMD or thyrotoxicosis was causally related to fracture risk, although one prospective study (451) demonstrated that a prior history of thyroid disease was independently associated with hip fracture after adjustment for BMD. Bauer et al (473, 485) demonstrated that low TSH was associated with a 3- to 4-fold increased risk of fracture, although a specific relationship between TSH and BMD was not identified. In agreement, Franklyn et al (486) identified an increased standardized mortality ratio due to hip fracture in a follow-up population register study of patients treated with radioiodine for hyperthyroidism. One cross-sectional study (487) identified an association between fracture and a prior history of thyrotoxicosis in postmenopausal women. A large population study reported persistence of an increased risk of fracture 5 years after initial diagnosis (456), but others failed to demonstrate a relationship between a history of thyrotoxicosis and fracture (452, 488). Recently, however, population studies have shown reduced BMD and an increased risk of fracture in postmenopausal women with thyrotoxicosis (447, 455, 456, 464, 473).
3. Systematic reviews and meta-analyses
A meta-analysis of 20 studies of patients with thyrotoxicosis calculated that BMD was reduced at the time of diagnosis and there was an increased risk of hip fracture; further investigation of the effect of antithyroid treatment demonstrated the low BMD at diagnosis returned to normal after 5 years (489).
IX. Osteoporosis and Genetic Variation in Thyroid Signaling
A. Associations with BMD
GWAS in osteoporosis cohorts have not identified associations between variation in thyroid-related genes and BMD or fracture (490–492).
Candidate gene studies have investigated relationships between polymorphisms in the TSHR, THRA, and DIO2 genes and BMD. The TSHR D727E polymorphism was associated with serum TSH concentration and with osteoporosis diagnosed by quantitative ultrasound in 150 male subjects compared to 150 controls, whereas the D36H polymorphism was not (493). By contrast, in 156 patients treated for differentiated thyroid cancer, the TSHR D727E polymorphism was associated with higher BMD at the femoral neck independent of thyroid status, but this association did not persist after adjustment for body mass index (494). A similar association between TSHR D727E and increased BMD at the femoral neck was identified in 1327 subjects from the Rotterdam study (438).
A candidate gene association study of 862 men over age 65 investigated genetic variation in THRA and BMD at the femoral neck and lumbar spine (495, 496), whereas a much larger study investigated the THRA locus in relation to BMD, fracture risk, and bone geometry in 27 326 individuals from the Genetic Factors for Osteoporosis (GEFOS) consortium and the Rotterdam Study 1 and 2 populations (497). Both studies failed to identify any relationship between variation in bone parameters and genetic variation across the THRA locus. In a further study, a cohort of 100 healthy euthyroid postmenopausal women with the highest BMD was selected from the OPUS population, and sequencing of THRA revealed no abnormalities, indicating that THRA mutations are not a common cause of high BMD (498). However, Yerges et al (495) identified an association between an intronic single nucleotide polymorphism in THRB and trabecular BMD in the Osteoporotic Fractures in Men (MrOS) study.
Variation in deiodinases has been described as a potential genetic determinant of bone pathology. Two studies investigated the relationship between deiodinase polymorphisms and skeletal parameters. The DIO2 T92A polymorphism was associated with decreased femoral neck and total hip BMD and several markers of bone turnover in 154 athyreotic patients treated for differentiated thyroid cancer (499), suggesting a role for DIO2 in the regulation of bone formation. However, in 641 young healthy men, no relationships between DIO1 or DIO2 variants and bone mass were identified (500). Sequencing of DIO2 in healthy euthyroid postmenopausal women with high BMD from the OPUS population also failed to identify any abnormalities (498).
X. Osteoarthritis and Genetic Variation in Thyroid Signaling
A. Genetics
A role for thyroid hormones in the pathogenesis of osteoarthritis (OA) has been proposed (501). A GWAS of siblings with generalized OA found a nonsynonymous coding variant (rs225014; T92A) that identified DIO2 as a disease-susceptibility locus (502). An allele-sharing approach reinforced evidence of linkage in the region of this locus (503). Replication studies confirmed this by identifying an association between symptomatic OA and a DIO2 haplotype containing the minor allele of rs225014 and major allele of rs12885300 (504). The relationship between these DIO2 single nucleotide polymorphisms and OA, however, was not replicated in the Rotterdam Study population (505), and DIO2 was not identified in recent meta-analyses (506, 507). Nevertheless, additional data suggested that the risk allele of rs225014 may be expressed at higher levels than the protective allele in OA cartilage from heterozygous patients (508), possibly because of epigenetic modifications (509). Increased DIO2 mRNA and protein expression has also been documented by RT-qPCR and immunohistochemistry in cartilage from joints affected by end-stage OA (508, 510). Furthermore, the rs12885300 DIO2 polymorphism has been suggested to influence the association between hip shape and OA susceptibility by increasing vulnerability of articular cartilage to abnormal hip morphology (511). Moreover, studies in transgenic rats that overexpress human DIO2 in chondrocytes indicate that these animals had increased susceptibility to OA after provocation surgery (510). Finally, a meta-analysis studying genes that regulate thyroid hormone metabolism and mediate T3 effects on chondrocytes identified DIO3 as a disease-modifying locus in OA (504).
Together, these data suggest that local T3 availability in joint tissues may play a role in articular cartilage renewal and repair, particularly because the associated DIO2 and DIO3 polymorphisms do not influence systemic thyroid status (16). Overall, increased T3 availability may be detrimental to joint maintenance and articular cartilage homeostasis.
B. Mechanism
Adult Dio2−/− mice have normal articular cartilage and no other features of spontaneous joint damage, but they exhibit increased subchondral bone mineral content (512). In a forced-exercise provocation model, Dio2−/− mice were protected from articular cartilage damage (513). In studies demonstrating that functional DIO2 enzyme is restricted to bone-forming osteoblasts, we showed that Dio2 mRNA expression does not necessarily correlate with enzyme activity (61). An important reason for this discrepancy is that DIO2 protein is labile, undergoing rapid degradation after exposure to increasing concentrations of T4 in a local feedback loop (50, 159). We demonstrated Dio2 mRNA in growth plate cartilage but could not detect enzyme activity using a high sensitivity assay (61), whereas others showed Dio2 mRNA expression in rat articular cartilage (510) and increased levels of DIO2 mRNA (510) and protein (508) in human articular cartilage from joints resected for end-stage OA. In each case, however, enzyme activity was not determined. The significance of increased DIO2 mRNA in end-stage OA cartilage is therefore uncertain; it is also unknown whether increased DIO2 expression might represent a secondary response to joint destruction or whether it precedes cartilage damage and might be a causative factor in disease progression.
Transgenic rats overexpressing DIO2 in chondrocytes had increased susceptibility to OA after surgical provocation (510). Nevertheless, a causal relationship between increased DIO2 expression and OA susceptibility was not established because enzyme activity was not determined (510). This is particularly important because in a previous study, transgenic mice overexpressing Dio2 in the heart surprisingly displayed only mild thyrotoxic changes in cardiomyocytes (514), likely because the phenotype was mitigated by T4-induced degradation of the overexpressed enzyme (159). Furthermore, small interfering RNA-mediated inhibition of DIO2 in primary human chondrocytes resulted in decreased expression of liver X receptor α and increased expression of IL-1β and IL1β-induced cyclooxygenase-2 expression (515). Thus, in contrast to the previous studies, these findings suggest that suppression of Dio2 results in a proinflammatory response in cartilage. The increase in DIO2 expression observed in end-stage human OA (508, 510) may, therefore, be a consequence of disease progression resulting from activation of proinflammatory pathways such as the NFκB pathway, which is known to stimulate Dio2 expression (516, 517).
Although chondrocytes resist terminal differentiation in healthy articular cartilage, a process resembling endochondral ossification occurs during OA in which articular chondrocytes undergo hypertrophic differentiation with accelerated cartilage mineralization (518, 519). ADAMTS5 (520, 521) and MMP13 (522) degrade cartilage matrix, and these enzymes are regulated by T3 in the growth plate (205, 222, 223, 523, 524). In articular cartilage, thyroid hormones stimulate terminal chondrocyte differentiation (525) and tissue transglutaminase activity (526). In cocultures of chondrocytes derived from different articular cartilage zones, interactions between chondrocytes from differing zones were identified that modulated responses to T3 and were mediated in part by PTHrP. The authors proposed that communication between zones might regulate postnatal articular cartilage organization and mineralization (527).
Currently, the pathophysiological importance of these studies is difficult to evaluate. Initial genetic studies suggesting that reduced DIO2 activity is associated with increased susceptibility to OA (502) were based on the assumption that the T92A polymorphism results in reduced enzyme activity (528). However, studies showing that transgenic rats overexpressing DIO2 in articular cartilage had increased susceptibility to OA (510) were not consistent with this conclusion, and recent findings of allelic imbalance and increased expression of DIO2 protein in OA cartilage (508, 509) further suggest that increased DIO2 activity may be detrimental for articular cartilage maintenance. On the other hand, a large GWAS and recent meta-analyses failed to identify DIO2 as a disease susceptibility locus for OA (505–507), whereas a recent study in primary human chondrocytes suggests an anti-inflammatory role for DIO2 in cartilage (515). Thus, it remains unclear whether variation in DIO2 plays a role in OA pathogenesis, although the independent identification of DIO3 as a disease susceptibility locus (504) supports a role for control of local tissue T3 availability in the regulation of joint homeostasis.
XI. Summary and Future Directions
The skeleton is exquisitely sensitive to thyroid hormones, which have profound effects on bone development, linear growth, and adult bone maintenance.
Thyroid hormone deficiency in children results in cessation of growth and bone maturation, whereas thyrotoxicosis accelerates these processes. In adults, thyrotoxicosis is an important and established cause of secondary osteoporosis, and an increased risk of fracture has now been demonstrated in subclinical hyperthyroidism. Furthermore, even thyroid status at the upper end of the normal euthyroid reference range is associated with an increased risk of fracture in postmenopausal women.
An extensive series of studies in genetically modified mice has shown that T3 exerts anabolic actions on the developing skeleton and has catabolic effects in adulthood, and these actions are mediated predominantly by TRα1. The importance of the local regulation of T3 availability in bone has been demonstrated in studies that identified a critical role for DIO2 in osteoblasts to optimize bone mineralization and strength. The translational importance and clinical relevance of such studies is highlighted by the characterization of mice harboring deletions or dominant-negative mutations of Thra, which accurately predicted the abnormalities seen in patients recently identified with RTHα. Moreover, mice with dominant-negative mutations of Thra also represent an important disease model in which to investigate novel therapeutic approaches in these patients. In addition to studies in genetically modified mice, analysis of patients with thyroid disease or inherited disorders of T3 action is consistent with a major physiological role for T3 in the regulation of skeletal development and adult bone maintenance. Analysis of Tshr−/− mice and studies of TSH administration in rodents have also suggested that TSH acts as a negative regulator of bone turnover. Nevertheless, the physiological role of TSH in the skeleton remains uncertain because its proposed actions are not wholly consistent with findings in human diseases and mouse models in which the physiological inverse relationship between thyroid hormones and TSH is dissociated.
Precise determination of the cellular and molecular mechanisms of T3 and TSH actions in the skeleton in vivo will require cell-specific conditional gene targeting approaches in individual bone cell lineages. Combined with genome-wide gene expression analysis, these approaches will determine key target genes and downstream signaling pathways and have the potential to identify new therapeutic targets for skeletal disease.
Recent studies have also identified DIO2 and DIO3 as disease susceptibility loci for OA, a major degenerative disease of increasing prevalence in the aging population. These data establish a new field of research and further highlight the fundamental importance of understanding the mechanisms of T3 action in cartilage and bone and its role in tissue maintenance, response to injury, and pathogenesis of degenerative disease.
Acknowledgments
The authors are very grateful to Dr. Jonathan LoPresti, Keck School of Medicine, University of Southern California, for kindly sharing clinical details and providing x-rays showing the profound skeletal consequences of undiagnosed congenital hypothyroidism. We also thank and acknowledge our numerous collaborators for sharing reagents over the years and the current and past members of the Molecular Endocrinology Laboratory for their hard work and dedication.
This work was supported by grants from the Medical Research Council (G108/502, G0501486, and G800261), the Wellcome Trust (GR076584), Arthritis Research UK (18292 and 19744), the European Union Marie-Curie Fellowship (509311), the Sir Jules Thorn Trust, the Society for Endocrinology, and the British Thyroid Foundation.
Disclosure Summary: The authors have nothing to disclose.
Footnotes
- BMD
- bone mineral density
- BMP
- bone morphogenic protein
- CSF
- colony-stimulating factor
- CSF-1R
- CSF receptor
- DIO1
- type 1 deiodinase
- DKK1
- Dickkopf1-related protein 1
- E
- embryonic day
- ESC
- embryonic stem cell
- FGF
- fibroblast growth factor
- FGFR
- FGF receptor
- fT3
- free T3
- fT4
- free T4
- GFP
- green fluorescent protein
- GWAS
- genome-wide association study
- HPT
- hypothalamic-pituitary-thyroid
- HSPG
- heparan sulfate proteoglycan
- IGFBP
- IGF binding protein
- IHH
- Indian hedgehog
- LAT
- L-type amino acid transporter
- M-CSF
- macrophage-CSF
- MCT
- monocarboxylate transporter
- MMP
- matrix metalloproteinase
- NF-κB
- nuclear factor-κB
- OA
- osteoarthritis
- OATP1c1
- organic anion transporter protein-1c1
- OPG
- osteoprotegerin
- P
- postnatal day
- RANK
- receptor activator of NF-κB
- RANKL
- RANK ligand
- rhTSH
- recombinant human TSH
- RTH
- resistance to thyroid hormone
- RXR
- retinoid X receptor
- SBP2
- Selenocysteine insertion sequence (SECIS)-binding protein 2
- Sec
- selenocysteine
- TR
- thyroid hormone receptor
- TRE
- T3 response element
- TSHR
- TSH receptor.
Reference
- 1. Medvei VC. A history of endocrinology. Lancaster, UK: MTP Press; 1982. [Google Scholar]
- 2. Braverman LE, Cooper DS. Werner & Ingbar's the Thyroid: a Fundamental and Clinical Text. 10th ed Philadelphia, PA: Lippincott Williams Wilkins; 2013. [Google Scholar]
- 3. Wass JA, Stewart PM. Oxford Textbook of Endocrinology and Diabetes. 2nd ed New York: Oxford University Press; 2011. [Google Scholar]
- 4. Delling G, Kummerfeldt K. Friedrich Daniel von Recklinghausen. A reminiscence on the occasion of the centenary of his publication Osteitis fibrosa or deformans, osteomalacia and osteoplastic carcinosis in their interrelationships [in German]. Dtsch Med Wochenschr. 1991;116:1976–1979. [DOI] [PubMed] [Google Scholar]
- 5. Von Recklinghausen FD. Die Fibrose oder deformierende Ostitis, die Osteomalacie und die osteoplastische Carcinose in ihren gegenseitigen Beziehungen. In: Festschrift Rudolf Virchow zum 13. Berlin: Georg Reimer Verlag; 1891. [Google Scholar]
- 6. Bianco AC, Salvatore D, Gereben B, Berry MJ, Larsen PR. Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeiodinases. Endocr Rev. 2002;23:38–89. [DOI] [PubMed] [Google Scholar]
- 7. Kopp P. The TSH receptor and its role in thyroid disease. Cell Mol Life Sci. 2001;58:1301–1322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Abel ED, Ahima RS, Boers ME, Elmquist JK, Wondisford FE. Critical role for thyroid hormone receptor β2 in the regulation of paraventricular thyrotropin-releasing hormone neurons. J Clin Invest. 2001;107:1017–1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Forrest D, Hanebuth E, Smeyne RJ, et al. Recessive resistance to thyroid hormone in mice lacking thyroid hormone receptor β: evidence for tissue-specific modulation of receptor function. EMBO J. 1996;15:3006–3015. [PMC free article] [PubMed] [Google Scholar]
- 10. Macchia PE, Takeuchi Y, Kawai T, et al. Increased sensitivity to thyroid hormone in mice with complete deficiency of thyroid hormone receptor α. Proc Natl Acad Sci USA. 2001;98:349–354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Nikrodhanond AA, Ortiga-Carvalho TM, Shibusawa N, et al. Dominant role of thyrotropin-releasing hormone in the hypothalamic-pituitary-thyroid axis. J Biol Chem. 2006;281:5000–5007. [DOI] [PubMed] [Google Scholar]
- 12. Andersen S, Bruun NH, Pedersen KM, Laurberg P. Biologic variation is important for interpretation of thyroid function tests. Thyroid. 2003;13:1069–1078. [DOI] [PubMed] [Google Scholar]
- 13. Bassett JH, Williams GR. Critical role of the hypothalamic-pituitary-thyroid axis in bone. Bone. 2008;43:418–426. [DOI] [PubMed] [Google Scholar]
- 14. Panicker V, Wilson SG, Spector TD, et al. Heritability of serum TSH, free T4 and free T3 concentrations: a study of a large UK twin cohort. Clin Endocrinol (Oxf). 2008;68:652–659. [DOI] [PubMed] [Google Scholar]
- 15. Medici M, van der Deure WM, Verbiest M, et al. A large-scale association analysis of 68 thyroid hormone pathway genes with serum TSH and FT4 levels. Eur J Endocrinol. 2011;164:781–788. [DOI] [PubMed] [Google Scholar]
- 16. Panicker V, Wilson SG, Spector TD, et al. Genetic loci linked to pituitary-thyroid axis set points: a genome-wide scan of a large twin cohort. J Clin Endocrinol Metab. 2008;93:3519–3523. [DOI] [PubMed] [Google Scholar]
- 17. Porcu E, Medici M, Pistis G, et al. A meta-analysis of thyroid-related traits reveals novel loci and gender-specific differences in the regulation of thyroid function. PLoS Genet. 2013;9:e1003266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Davies TF, Ando T, Lin RY, Tomer Y, Latif R. Thyrotropin receptor-associated diseases: from adenomata to Graves disease. J Clin Invest. 2005;115:1972–1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Bell A, Gagnon A, Dods P, Papineau D, Tiberi M, Sorisky A. TSH signaling and cell survival in 3T3–L1 preadipocytes. Am J Physiol Cell Physiol. 2002;283:C1056–C1064. [DOI] [PubMed] [Google Scholar]
- 20. Kero J, Ahmed K, Wettschureck N, et al. Thyrocyte-specific Gq/G11 deficiency impairs thyroid function and prevents goiter development. J Clin Invest. 2007;117:2399–2407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Park ES, Kim H, Suh JM, et al. Involvement of JAK/STAT (Janus kinase/signal transducer and activator of transcription) in the thyrotropin signaling pathway. Mol Endocrinol. 2000;14:662–670. [DOI] [PubMed] [Google Scholar]
- 22. Vassart G, Dumont JE. The thyrotropin receptor and the regulation of thyrocyte function and growth. Endocr Rev. 1992;13:596–611. [DOI] [PubMed] [Google Scholar]
- 23. Laugwitz KL, Allgeier A, Offermanns S, et al. The human thyrotropin receptor: a heptahelical receptor capable of stimulating members of all four G protein families. Proc Natl Acad Sci USA. 1996;93:116–120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Cleator JH, Ravenell R, Kurtz DT, Hildebrandt JD. A dominant negative Gαs mutant that prevents thyroid-stimulating hormone receptor activation of cAMP production and inositol 1,4,5-trisphosphate turnover: competition by different G proteins for activation by a common receptor. J Biol Chem. 2004;279:36601–36607. [DOI] [PubMed] [Google Scholar]
- 25. Davies T, Marians R, Latif R. The TSH receptor reveals itself. J Clin Invest. 2002;110:161–164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Klein JR. Physiological relevance of thyroid stimulating hormone and thyroid stimulating hormone receptor in tissues other than the thyroid. Autoimmunity. 2003;36:417–421. [DOI] [PubMed] [Google Scholar]
- 27. Prummel MF, Brokken LJ, Meduri G, Misrahi M, Bakker O, Wiersinga WM. Expression of the thyroid-stimulating hormone receptor in the folliculo-stellate cells of the human anterior pituitary. J Clin Endocrinol Metab. 2000;85:4347–4353. [DOI] [PubMed] [Google Scholar]
- 28. Williams GR. Extrathyroidal expression of TSH receptor. Annales d'Endocrinologie. 2011;72:68–73. [DOI] [PubMed] [Google Scholar]
- 29. Nakao N, Ono H, Yamamura T, et al. Thyrotrophin in the pars tuberalis triggers photoperiodic response. Nature. 2008;452:317–322. [DOI] [PubMed] [Google Scholar]
- 30. Ono H, Hoshino Y, Yasuo S, et al. Involvement of thyrotropin in photoperiodic signal transduction in mice. Proc Natl Acad Sci USA. 2008;105:18238–18242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Yoshimura T, Yasuo S, Watanabe M, et al. Light-induced hormone conversion of T4 to T3 regulates photoperiodic response of gonads in birds. Nature. 2003;426:178–181. [DOI] [PubMed] [Google Scholar]
- 32. Abe E, Marians RC, Yu W, et al. TSH is a negative regulator of skeletal remodeling. Cell. 2003;115:151–162. [DOI] [PubMed] [Google Scholar]
- 33. Valyasevi RW, Erickson DZ, Harteneck DA, et al. Differentiation of human orbital preadipocyte fibroblasts induces expression of functional thyrotropin receptor. J Clin Endocrinol Metab. 1999;84:2557–2562. [DOI] [PubMed] [Google Scholar]
- 34. van Zeijl CJ, Fliers E, van Koppen CJ, et al. Thyrotropin receptor-stimulating Graves' disease immunoglobulins induce hyaluronan synthesis by differentiated orbital fibroblasts from patients with Graves' ophthalmopathy not only via cyclic adenosine monophosphate signaling pathways. Thyroid. 2011;21:169–176. [DOI] [PubMed] [Google Scholar]
- 35. Kumar S, Schiefer R, Coenen MJ, Bahn RS. A stimulatory thyrotropin receptor antibody (M22) and thyrotropin increase interleukin-6 expression and secretion in Graves' orbital preadipocyte fibroblasts. Thyroid. 2010;20:59–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Balzan S, Nicolini G, Forini F, et al. Presence of a functional TSH receptor on human erythrocytes. Biomed Pharmacother. 2007;61:463–467. [DOI] [PubMed] [Google Scholar]
- 37. Dutton CM, Joba W, Spitzweg C, Heufelder AE, Bahn RS. Thyrotropin receptor expression in adrenal, kidney, and thymus. Thyroid. 1997;7:879–884. [DOI] [PubMed] [Google Scholar]
- 38. Hase H, Ando T, Eldeiry L, et al. TNFα mediates the skeletal effects of thyroid-stimulating hormone. Proc Natl Acad Sci USA. 2006;103:12849–12854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Wang HC, Dragoo J, Zhou Q, Klein JR. An intrinsic thyrotropin-mediated pathway of TNF-α production by bone marrow cells. Blood. 2003;101:119–123. [DOI] [PubMed] [Google Scholar]
- 40. Coutelier JP, Kehrl JH, Bellur SS, Kohn LD, Notkins AL, Prabhakar BS. Binding and functional effects of thyroid stimulating hormone on human immune cells. J Clin Immunol. 1990;10:204–210. [DOI] [PubMed] [Google Scholar]
- 41. Pekonen F, Weintraub BD. Thyrotropin binding to cultured lymphocytes and thyroid cells. Endocrinology. 1978;103:1668–1677. [DOI] [PubMed] [Google Scholar]
- 42. Wang J, Klein JR. Thymus-neuroendocrine interactions in extrathymic T cell development. Science. 1994;265:1860–1862. [DOI] [PubMed] [Google Scholar]
- 43. Wang J, Whetsell M, Klein JR. Local hormone networks and intestinal T cell homeostasis. Science. 1997;275:1937–1939. [DOI] [PubMed] [Google Scholar]
- 44. Visser WE, Friesema EC, Visser TJ. Minireview: thyroid hormone transporters: the knowns and the unknowns. Mol Endocrinol. 2011;25:1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Bernal J. Thyroid hormone receptors in brain development and function. Nat Clin Pract Endocrinol Metab. 2007;3:249–259. [DOI] [PubMed] [Google Scholar]
- 46. Dumitrescu AM, Liao XH, Best TB, Brockmann K, Refetoff S. A novel syndrome combining thyroid and neurological abnormalities is associated with mutations in a monocarboxylate transporter gene. Am J Hum Genet. 2004;74:168–175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Friesema EC, Grueters A, Biebermann H, et al. Association between mutations in a thyroid hormone transporter and severe X-linked psychomotor retardation. Lancet. 2004;364:1435–1437. [DOI] [PubMed] [Google Scholar]
- 48. Lin KH, Fukuda T, Cheng SY. Hormone and DNA binding activity of a purified human thyroid hormone nuclear receptor expressed in Escherichia coli. J Biol Chem. 1990;265:5161–5165. [PubMed] [Google Scholar]
- 49. Bianco AC, Kim BW. Deiodinases: implications of the local control of thyroid hormone action. J Clin Invest. 2006;116:2571–2579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Gereben B, Zavacki AM, Ribich S, et al. Cellular and molecular basis of deiodinase-regulated thyroid hormone signaling. Endocr Rev. 2008;29:898–938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. St Germain DL, Galton VA, Hernandez A. Minireview: defining the roles of the iodothyronine deiodinases: current concepts and challenges. Endocrinology. 2009;150:1097–1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Schneider MJ, Fiering SN, Thai B, et al. Targeted disruption of the type 1 selenodeiodinase gene (Dio1) results in marked changes in thyroid hormone economy in mice. Endocrinology. 2006;147:580–589. [DOI] [PubMed] [Google Scholar]
- 53. Heemstra KA, Soeters MR, Fliers E, et al. Type 2 iodothyronine deiodinase in skeletal muscle: effects of hypothyroidism and fasting. J Clin Endocrinol Metab. 2009;94:2144–2150. [DOI] [PubMed] [Google Scholar]
- 54. Larsen PR. Type 2 iodothyronine deiodinase in human skeletal muscle: new insights into its physiological role and regulation. J Clin Endocrinol Metab. 2009;94:1893–1895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Maia AL, Kim BW, Huang SA, Harney JW, Larsen PR. Type 2 iodothyronine deiodinase is the major source of plasma T3 in euthyroid humans. J Clin Invest. 2005;115:2524–2533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Fonseca TL, Correa-Medina M, Campos MP, et al. Coordination of hypothalamic and pituitary T3 production regulates TSH expression. J Clin Invest. 2013;123:1492–1500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Luongo C, Martin C, Vella K, et al. The selective loss of the type 2 iodothyronine deiodinase in mouse thyrotrophs increases basal TSH but blunts the thyrotropin response to hypothyroidism. Endocrinology. 2015;156:745–754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Werneck de Castro JP, Fonseca TL, Ueta CB, et al. Differences in hypothalamic type 2 deiodinase ubiquitination explain localized sensitivity to thyroxine. J Clin Invest. 2015;125:769–781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Friesema EC, Kuiper GG, Jansen J, Visser TJ, Kester MH. Thyroid hormone transport by the human monocarboxylate transporter 8 and its rate-limiting role in intracellular metabolism. Mol Endocrinol. 2006;20:2761–2772. [DOI] [PubMed] [Google Scholar]
- 60. Bassett JH, Boyde A, Howell PG, et al. Optimal bone strength and mineralization requires the type 2 iodothyronine deiodinase in osteoblasts. Proc Natl Acad Sci USA. 2010;107:7604–7609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Williams AJ, Robson H, Kester MH, et al. Iodothyronine deiodinase enzyme activities in bone. Bone. 2008;43:126–134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Campos-Barros A, Amma LL, Faris JS, Shailam R, Kelley MW, Forrest D. Type 2 iodothyronine deiodinase expression in the cochlea before the onset of hearing. Proc Natl Acad Sci USA. 2000;97:1287–1292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Ng L, Hernandez A, He W, et al. A protective role for type 3 deiodinase, a thyroid hormone-inactivating enzyme, in cochlear development and auditory function. Endocrinology. 2009;150:1952–1960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Ng L, Lyubarsky A, Nikonov SS, et al. Type 3 deiodinase, a thyroid-hormone-inactivating enzyme, controls survival and maturation of cone photoreceptors. J Neurosci. 2010;30:3347–3357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Simonides WS, Mulcahey MA, Redout EM, et al. Hypoxia-inducible factor induces local thyroid hormone inactivation during hypoxic-ischemic disease in rats. J Clin Invest. 2008;118:975–983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Bates JM, St Germain DL, Galton VA. Expression profiles of the three iodothyronine deiodinases, D1, D2, and D3, in the developing rat. Endocrinology. 1999;140:844–851. [DOI] [PubMed] [Google Scholar]
- 67. Berry DL, Rose CS, Remo BF, Brown DD. The expression pattern of thyroid hormone response genes in remodeling tadpole tissues defines distinct growth and resorption gene expression programs. Dev Biol. 1998;203:24–35. [DOI] [PubMed] [Google Scholar]
- 68. Wasco EC, Martinez E, Grant KS, St Germain EA, St Germain DL, Galton VA. Determinants of iodothyronine deiodinase activities in rodent uterus. Endocrinology. 2003;144:4253–4261. [DOI] [PubMed] [Google Scholar]
- 69. Forrest D, Sjöberg M, Vennström B. Contrasting developmental and tissue-specific expression of α and β thyroid hormone receptor genes. EMBO J. 1990;9:1519–1528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Sap J, Muñoz A, Damm K, et al. The c-erb-A protein is a high-affinity receptor for thyroid hormone. Nature. 1986;324:635–640. [DOI] [PubMed] [Google Scholar]
- 71. Weinberger C, Thompson CC, Ong ES, Lebo R, Gruol DJ, Evans RM. The c-erb-A gene encodes a thyroid hormone receptor. Nature. 1986;324:641–646. [DOI] [PubMed] [Google Scholar]
- 72. Harvey CB, Williams GR. Mechanism of thyroid hormone action. Thyroid. 2002;12:441–446. [DOI] [PubMed] [Google Scholar]
- 73. Chassande O, Fraichard A, Gauthier K, et al. Identification of transcripts initiated from an internal promoter in the c-erbA α locus that encode inhibitors of retinoic acid receptor-α and triiodothyronine receptor activities. Mol Endocrinol. 1997;11:1278–1290. [DOI] [PubMed] [Google Scholar]
- 74. Bassett JH, Harvey CB, Williams GR. Mechanisms of thyroid hormone receptor-specific nuclear and extra nuclear actions. Mol Cell Endocrinol. 2003;213:1–11. [DOI] [PubMed] [Google Scholar]
- 75. Kalyanaraman H, Schwappacher R, Joshua J, et al. Nongenomic thyroid hormone signaling occurs through a plasma membrane-localized receptor. Sci Signal. 2014;7:ra48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76. Harvey CB, Bassett JH, Maruvada P, Yen PM, Williams GR. The rat thyroid hormone receptor (TR) Deltabeta3 displays cell-, TR isoform-, and thyroid hormone response element-specific actions. Endocrinology. 2007;148:1764–1773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77. Williams GR. Cloning and characterization of two novel thyroid hormone receptor β isoforms. Mol Cell Biol. 2000;20:8329–8342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Abel ED, Boers ME, Pazos-Moura C, et al. Divergent roles for thyroid hormone receptor β isoforms in the endocrine axis and auditory system. J Clin Invest. 1999;104:291–300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Jones I, Ng L, Liu H, Forrest D. An intron control region differentially regulates expression of thyroid hormone receptor β2 in the cochlea, pituitary, and cone photoreceptors. Mol Endocrinol. 2007;21:1108–1119. [DOI] [PubMed] [Google Scholar]
- 80. Ng L, Hurley JB, Dierks B, et al. A thyroid hormone receptor that is required for the development of green cone photoreceptors. Nat Genet. 2001;27:94–98. [DOI] [PubMed] [Google Scholar]
- 81. Chassande O. Do unliganded thyroid hormone receptors have physiological functions? J Mol Endocrinol. 2003;31:9–20. [DOI] [PubMed] [Google Scholar]
- 82. Hashimoto K, Curty FH, Borges PP, et al. An unliganded thyroid hormone receptor causes severe neurological dysfunction. Proc Natl Acad Sci USA. 2001;98:3998–4003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Venero C, Guadaño-Ferraz A, Herrero AI, et al. Anxiety, memory impairment, and locomotor dysfunction caused by a mutant thyroid hormone receptor α1 can be ameliorated by T3 treatment. Genes Dev. 2005;19:2152–2163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Wallis K, Sjögren M, van Hogerlinden M, et al. Locomotor deficiencies and aberrant development of subtype-specific GABAergic interneurons caused by an unliganded thyroid hormone receptor α1. J Neurosci. 2008;28:1904–1915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Astapova I, Lee LJ, Morales C, Tauber S, Bilban M, Hollenberg AN. The nuclear corepressor, NCoR, regulates thyroid hormone action in vivo. Proc Natl Acad Sci USA. 2008;105:19544–19549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Astapova I, Vella KR, Ramadoss P, et al. The nuclear receptor corepressor (NCoR) controls thyroid hormone sensitivity and the set point of the hypothalamic-pituitary-thyroid axis. Mol Endocrinol. 2011;25:212–224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Brent GA. Mechanisms of thyroid hormone action. J Clin Invest. 2012;122:3035–3043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Cheng SY, Leonard JL, Davis PJ. Molecular aspects of thyroid hormone actions. Endocr Rev. 2010;31:139–170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Vella KR, Ramadoss P, Costa-E-Sousa RH, et al. Thyroid hormone signaling in vivo requires a balance between coactivators and corepressors. Mol Cell Biol. 2014;34:1564–1575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90. Lösel R, Wehling M. Nongenomic actions of steroid hormones. Nat Rev Mol Cell Biol. 2003;4:46–56. [DOI] [PubMed] [Google Scholar]
- 91. Bergh JJ, Lin HY, Lansing L, et al. Integrin αVβ3 contains a cell surface receptor site for thyroid hormone that is linked to activation of mitogen-activated protein kinase and induction of angiogenesis. Endocrinology. 2005;146:2864–2871. [DOI] [PubMed] [Google Scholar]
- 92. Davis FB, Tang HY, Shih A, et al. Acting via a cell surface receptor, thyroid hormone is a growth factor for glioma cells. Cancer Res. 2006;66:7270–7275. [DOI] [PubMed] [Google Scholar]
- 93. Cao X, Kambe F, Moeller LC, Refetoff S, Seo H. Thyroid hormone induces rapid activation of Akt/protein kinase B-mammalian target of rapamycin-p70S6K cascade through phosphatidylinositol 3-kinase in human fibroblasts. Mol Endocrinol. 2005;19:102–112. [DOI] [PubMed] [Google Scholar]
- 94. Gauthier K, Flamant F. Nongenomic, TRβ-dependent, thyroid hormone response gets genetic support. Endocrinology. 2014;155:3206–3209. [DOI] [PubMed] [Google Scholar]
- 95. Martin NP, Marron Fernandez de Velasco E, Mizuno F, et al. A rapid cytoplasmic mechanism for PI3 kinase regulation by the nuclear thyroid hormone receptor, TRβ, and genetic evidence for its role in the maturation of mouse hippocampal synapses in vivo. Endocrinology. 2014;155:3713–3724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Furuya F, Hanover JA, Cheng SY. Activation of phosphatidylinositol 3-kinase signaling by a mutant thyroid hormone β receptor. Proc Natl Acad Sci USA. 2006;103:1780–1785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Storey NM, Gentile S, Ullah H, et al. Rapid signaling at the plasma membrane by a nuclear receptor for thyroid hormone. Proc Natl Acad Sci USA. 2006;103:5197–5201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Moeller LC, Dumitrescu AM, Refetoff S. Cytosolic action of thyroid hormone leads to induction of hypoxia-inducible factor-1α and glycolytic genes. Mol Endocrinol. 2005;19:2955–2963. [DOI] [PubMed] [Google Scholar]
- 99. Moeller LC, Broecker-Preuss M. Transcriptional regulation by nonclassical action of thyroid hormone. Thyroid Res. 2011;4(suppl 1):S6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Craft AM, Ahmed N, Rockel JS, et al. Specification of chondrocytes and cartilage tissues from embryonic stem cells. Development. 2013;140:2597–2610. [DOI] [PubMed] [Google Scholar]
- 101. Kronenberg HM. Developmental regulation of the growth plate. Nature. 2003;423:332–336. [DOI] [PubMed] [Google Scholar]
- 102. Karsenty G, Kronenberg HM, Settembre C. Genetic control of bone formation. Annu Rev Cell Dev Biol. 2009;25:629–648. [DOI] [PubMed] [Google Scholar]
- 103. Zuscik MJ, Hilton MJ, Zhang X, Chen D, O'Keefe RJ. Regulation of chondrogenesis and chondrocyte differentiation by stress. J Clin Invest. 2008;118:429–438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Martin TJ, Sims NA, Ng KW. Regulatory pathways revealing new approaches to the development of anabolic drugs for osteoporosis. Osteoporos Int. 2008;19:1125–1138. [DOI] [PubMed] [Google Scholar]
- 105. Long F. Building strong bones: molecular regulation of the osteoblast lineage. Nat Rev Mol Cell Biol. 2012;13:27–38. [DOI] [PubMed] [Google Scholar]
- 106. Nakashima K, de Crombrugghe B. Transcriptional mechanisms in osteoblast differentiation and bone formation. Trends Genet. 2003;19:458–466. [DOI] [PubMed] [Google Scholar]
- 107. Hartmann C. A Wnt canon orchestrating osteoblastogenesis. Trends Cell Biol. 2006;16:151–158. [DOI] [PubMed] [Google Scholar]
- 108. Zaidi M. Skeletal remodeling in health and disease. Nat Med. 2007;13:791–801. [DOI] [PubMed] [Google Scholar]
- 109. Bilezikian JP, Matsumoto T, Bellido T, et al. Targeting bone remodeling for the treatment of osteoporosis: summary of the proceedings of an ASBMR workshop. J Bone Miner Res. 2009;24:373–385. [DOI] [PubMed] [Google Scholar]
- 110. Bonewald LF, Johnson ML. Osteocytes, mechanosensing and Wnt signaling. Bone. 2008;42:606–615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Han Y, Cowin SC, Schaffler MB, Weinbaum S. Mechanotransduction and strain amplification in osteocyte cell processes. Proc Natl Acad Sci USA. 2004;101:16689–16694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Seeman E, Delmas PD. Bone quality–the material and structural basis of bone strength and fragility. N Engl J Med. 2006;354:2250–2261. [DOI] [PubMed] [Google Scholar]
- 113. Dallas SL, Prideaux M, Bonewald LF. The osteocyte: an endocrine cell … and more. Endocr Rev. 2013;34:658–690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. Nakashima T, Hayashi M, Fukunaga T, et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat Med. 2011;17:1231–1234. [DOI] [PubMed] [Google Scholar]
- 115. Bellido T. Osteocyte-driven bone remodeling. Calcif Tissue Int. 2014;94:25–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Väänänen HK, Zhao H, Mulari M, Halleen JM. The cell biology of osteoclast function. J Cell Sci. 2000;113:377–381. [DOI] [PubMed] [Google Scholar]
- 117. Tolar J, Teitelbaum SL, Orchard PJ. Osteopetrosis. N Engl J Med. 2004;351:2839–2849. [DOI] [PubMed] [Google Scholar]
- 118. Edwards JR, Mundy GR. Advances in osteoclast biology: old findings and new insights from mouse models. Nat Rev Rheumatol. 2011;7:235–243. [DOI] [PubMed] [Google Scholar]
- 119. Rice DP, Rice R. Locate, condense, differentiate, grow and confront: developmental mechanisms controlling intramembranous bone and suture formation and function. Front Oral Biol. 2008;12:22–40. [DOI] [PubMed] [Google Scholar]
- 120. Lassová L, Niu Z, Golden EB, Cohen AJ, Adams SL. Thyroid hormone treatment of cultured chondrocytes mimics in vivo stimulation of collagen X mRNA by increasing BMP 4 expression. J Cell Physiol. 2009;219:595–605. [DOI] [PubMed] [Google Scholar]
- 121. Jacenko O, LuValle PA, Olsen BR. Spondylometaphyseal dysplasia in mice carrying a dominant negative mutation in a matrix protein specific for cartilage-to-bone transition. Nature. 1993;365:56–61. [DOI] [PubMed] [Google Scholar]
- 122. Lai LP, Mitchell J. Indian hedgehog: its roles and regulation in endochondral bone development. J Cell Biochem. 2005;96:1163–1173. [DOI] [PubMed] [Google Scholar]
- 123. Karsenty G, Wagner EF. Reaching a genetic and molecular understanding of skeletal development. Dev Cell. 2002;2:389–406. [DOI] [PubMed] [Google Scholar]
- 124. Bonjour JP, Chevalley T. Pubertal timing, bone acquisition, and risk of fracture throughout life. Endocr Rev. 2014;35:820–847. [DOI] [PubMed] [Google Scholar]
- 125. Raggatt LJ, Partridge NC. Cellular and molecular mechanisms of bone remodeling. J Biol Chem. 2010;285:25103–25108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126. Bonewald LF. The amazing osteocyte. J Bone Miner Res. 2011;26:229–238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Khosla S, Westendorf JJ, Oursler MJ. Building bone to reverse osteoporosis and repair fractures. J Clin Invest. 2008;118:421–428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Heino TJ, Hentunen TA, Väänänen HK. Osteocytes inhibit osteoclastic bone resorption through transforming growth factor-β: enhancement by estrogen. J Cell Biochem. 2002;85:185–197. [DOI] [PubMed] [Google Scholar]
- 129. Burgess TL, Qian Y, Kaufman S, et al. The ligand for osteoprotegerin (OPGL) directly activates mature osteoclasts. J Cell Biol. 1999;145:527–538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Kearns AE, Khosla S, Kostenuik PJ. Receptor activator of nuclear factor κB ligand and osteoprotegerin regulation of bone remodeling in health and disease. Endocr Rev. 2008;29:155–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Yasuda H, Shima N, Nakagawa N, et al. Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc Natl Acad Sci USA. 1998;95:3597–3602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132. Lacey DL, Timms E, Tan HL, et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell. 1998;93:165–176. [DOI] [PubMed] [Google Scholar]
- 133. Teitelbaum SL, Ross FP. Genetic regulation of osteoclast development and function. Nat Rev Genet. 2003;4:638–649. [DOI] [PubMed] [Google Scholar]
- 134. Baron R, Rawadi G. Wnt signaling and the regulation of bone mass. Curr Osteoporos Rep. 2007;5:73–80. [DOI] [PubMed] [Google Scholar]
- 135. Baron R, Rawadi G. Targeting the Wnt/β-catenin pathway to regulate bone formation in the adult skeleton. Endocrinology. 2007;148:2635–2643. [DOI] [PubMed] [Google Scholar]
- 136. Canalis E, Giustina A, Bilezikian JP. Mechanisms of anabolic therapies for osteoporosis. N Engl J Med. 2007;357:905–916. [DOI] [PubMed] [Google Scholar]
- 137. Endo T, Kobayashi T. Excess TSH causes abnormal skeletal development in young mice with hypothyroidism via suppressive effects on the growth plate. Am J Physiol Endocrinol Metab. 2013;305:E660–E666. [DOI] [PubMed] [Google Scholar]
- 138. Bassett JH, Williams AJ, Murphy E, et al. A lack of thyroid hormones rather than excess thyrotropin causes abnormal skeletal development in hypothyroidism. Mol Endocrinol. 2008;22:501–512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Bassett JH, van der Spek A, Logan JG, et al. Thyrostimulin regulates osteoblastic bone formation during early skeletal development. Endocrinology. 2015;156:3098–3113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Baliram R, Sun L, Cao J, et al. Hyperthyroid-associated osteoporosis is exacerbated by the loss of TSH signaling. J Clin Invest. 2012;122:3737–3741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Vincent BH, Montufar-Solis D, Teng BB, Amendt BA, Schaefer J, Klein JR. Bone marrow cells produce a novel TSHβ splice variant that is upregulated in the thyroid following systemic virus infection. Genes Immun. 2009;10:18–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142. Baliram R, Chow A, Huber AK, et al. Thyroid and bone: macrophage-derived TSH-β splice variant increases murine osteoblastogenesis. Endocrinology. 2013;154:4919–4926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Bagriacik EU, Yaman M, Haznedar R, Sucak G, Delibasi T. TSH-induced gene expression involves regulation of self-renewal and differentiation-related genes in human bone marrow-derived mesenchymal stem cells. J Endocrinol. 2012;212:169–178. [DOI] [PubMed] [Google Scholar]
- 144. Inoue M, Tawata M, Yokomori N, Endo T, Onaya T. Expression of thyrotropin receptor on clonal osteoblast-like rat osteosarcoma cells. Thyroid. 1998;8:1059–1064. [DOI] [PubMed] [Google Scholar]
- 145. Sampath TK, Simic P, Sendak R, et al. Thyroid-stimulating hormone restores bone volume, microarchitecture, and strength in aged ovariectomized rats. J Bone Miner Res. 2007;22:849–859. [DOI] [PubMed] [Google Scholar]
- 146. Baliram R, Latif R, Berkowitz J, et al. Thyroid-stimulating hormone induces a Wnt-dependent, feed-forward loop for osteoblastogenesis in embryonic stem cell cultures. Proc Natl Acad Sci USA. 2011;108:16277–16282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Ramajayam G, Vignesh RC, Karthikeyan S, et al. Regulation of insulin-like growth factors and their binding proteins by thyroid stimulating hormone in human osteoblast-like (SaOS2) cells. Mol Cell Biochem. 2012;368:77–88. [DOI] [PubMed] [Google Scholar]
- 148. Boutin A, Eliseeva E, Gershengorn MC, Neumann S. β-Arrestin-1 mediates thyrotropin-enhanced osteoblast differentiation. FASEB J. 2014;28:3446–3455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Tsai JA, Janson A, Bucht E, et al. Weak evidence of thyrotropin receptors in primary cultures of human osteoblast-like cells. Calcif Tissue Int. 2004;74:486–491. [DOI] [PubMed] [Google Scholar]
- 150. Ma R, Morshed S, Latif R, Zaidi M, Davies TF. The influence of thyroid-stimulating hormone and thyroid-stimulating hormone receptor antibodies on osteoclastogenesis. Thyroid. 2011;21:897–906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Zhang W, Zhang Y, Liu Y, et al. Thyroid-stimulating hormone maintains bone mass and strength by suppressing osteoclast differentiation. J Biomech. 2014;47:1307–1314. [DOI] [PubMed] [Google Scholar]
- 152. Yamoah K, Brebene A, Baliram R, et al. High-mobility group box proteins modulate tumor necrosis factor-α expression in osteoclastogenesis via a novel deoxyribonucleic acid sequence. Mol Endocrinol. 2008;22:1141–1153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Sun L, Zhu LL, Lu P, et al. Genetic confirmation for a central role for TNFα in the direct action of thyroid stimulating hormone on the skeleton. Proc Natl Acad Sci USA. 2013;110:9891–9896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Capelo LP, Beber EH, Fonseca TL, Gouveia CH. The monocarboxylate transporter 8 and L-type amino acid transporters 1 and 2 are expressed in mouse skeletons and in osteoblastic MC3T3-E1 cells. Thyroid. 2009;19:171–180. [DOI] [PubMed] [Google Scholar]
- 155. Abe S, Namba N, Abe M, et al. Monocarboxylate transporter 10 functions as a thyroid hormone transporter in chondrocytes. Endocrinology. 2012;153:4049–4058. [DOI] [PubMed] [Google Scholar]
- 156. Waung JA, Bassett JH, Williams GR. Thyroid hormone metabolism in skeletal development and adult bone maintenance. Trends Endocrinol Metab. 2012;23:155–162. [DOI] [PubMed] [Google Scholar]
- 157. Capelo LP, Beber EH, Huang SA, Zorn TM, Bianco AC, Gouveia CH. Deiodinase-mediated thyroid hormone inactivation minimizes thyroid hormone signaling in the early development of fetal skeleton. Bone. 2008;43:921–930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Gouveia CH, Christoffolete MA, Zaitune CR, et al. Type 2 iodothyronine selenodeiodinase is expressed throughout the mouse skeleton and in the MC3T3-E1 mouse osteoblastic cell line during differentiation. Endocrinology. 2005;146:195–200. [DOI] [PubMed] [Google Scholar]
- 159. Dentice M, Bandyopadhyay A, Gereben B, et al. The hedgehog-inducible ubiquitin ligase subunit WSB-1 modulates thyroid hormone activation and PTHrP secretion in the developing growth plate. Nat Cell Biol. 2005;7:698–705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160. Lavery K, Hawley S, Swain P, Rooney R, Falb D, Alaoui-Ismaili MH. New insights into BMP-7 mediated osteoblastic differentiation of primary human mesenchymal stem cells. Bone. 2009;45:27–41. [DOI] [PubMed] [Google Scholar]
- 161. Robson H, Siebler T, Stevens DA, Shalet SM, Williams GR. Thyroid hormone acts directly on growth plate chondrocytes to promote hypertrophic differentiation and inhibit clonal expansion and cell proliferation. Endocrinology. 2000;141:3887–3897. [DOI] [PubMed] [Google Scholar]
- 162. Stevens DA, Hasserjian RP, Robson H, Siebler T, Shalet SM, Williams GR. Thyroid hormones regulate hypertrophic chondrocyte differentiation and expression of parathyroid hormone-related peptide and its receptor during endochondral bone formation. J Bone Miner Res. 2000;15:2431–2442. [DOI] [PubMed] [Google Scholar]
- 163. Bookout AL, Jeong Y, Downes M, Yu RT, Evans RM, Mangelsdorf DJ. Anatomical profiling of nuclear receptor expression reveals a hierarchical transcriptional network. Cell. 2006;126:789–799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164. O'Shea PJ, Harvey CB, Suzuki H, et al. A thyrotoxic skeletal phenotype of advanced bone formation in mice with resistance to thyroid hormone. Mol Endocrinol. 2003;17:1410–1424. [DOI] [PubMed] [Google Scholar]
- 165. Beber EH, Capelo LP, Fonseca TL, et al. The thyroid hormone receptor (TR) β-selective agonist GC-1 inhibits proliferation but induces differentiation and TR β mRNA expression in mouse and rat osteoblast-like cells. Calcif Tissue Int. 2009;84:324–333. [DOI] [PubMed] [Google Scholar]
- 166. Freitas FR, Capelo LP, O'Shea PJ, et al. The thyroid hormone receptor β-specific agonist GC-1 selectively affects the bone development of hypothyroid rats. J Bone Miner Res. 2005;20:294–304. [DOI] [PubMed] [Google Scholar]
- 167. Monfoulet LE, Rabier B, Dacquin R, et al. Thyroid hormone receptor β mediates thyroid hormone effects on bone remodeling and bone mass. J Bone Miner Res. 2011;26:2036–2044. [DOI] [PubMed] [Google Scholar]
- 168. Abu EO, Bord S, Horner A, Chatterjee VK, Compston JE. The expression of thyroid hormone receptors in human bone. Bone. 1997;21:137–142. [DOI] [PubMed] [Google Scholar]
- 169. Ballock Rt, Mita BC, Zhou X, Chen DH, Mink LM. Expression of thyroid hormone receptor isoforms in rat growth plate cartilage in vivo. J Bone Miner Res. 1999;14:1550–1556. [DOI] [PubMed] [Google Scholar]
- 170. Bland R, Sammons RL, Sheppard MC, Williams GR. Thyroid hormone, vitamin D and retinoid receptor expression and signalling in primary cultures of rat osteoblastic and immortalised osteosarcoma cells. J Endocrinol. 1997;154:63–74. [DOI] [PubMed] [Google Scholar]
- 171. Carrascosa A, Ferrández MA, Audi L, Ballabriga A. Effects of triiodothyronine (T3) and identification of specific nuclear T3-binding sites in cultured human fetal epiphyseal chondrocytes. J Clin Endocrinol Metab. 1992;75:140–144. [DOI] [PubMed] [Google Scholar]
- 172. Egrise D, Martin D, Neve P, Verhas M, Schoutens A. Effects and interactions of 17 β-estradiol, T3 and 1,25(OH)2D3 on cultured osteoblasts from mature rats. Bone Miner. 1990;11:273–283. [DOI] [PubMed] [Google Scholar]
- 173. Krieger NS, Stappenbeck TS, Stern PH. Characterization of specific thyroid hormone receptors in bone. J Bone Miner Res. 1988;3:473–478. [DOI] [PubMed] [Google Scholar]
- 174. Allain TJ, Yen PM, Flanagan AM, McGregor AM. The isoform-specific expression of the tri-iodothyronine receptor in osteoblasts and osteoclasts. Eur J Clin Invest. 1996;26:418–425. [DOI] [PubMed] [Google Scholar]
- 175. Gruber R, Czerwenka K, Wolf F, Ho GM, Willheim M, Peterlik M. Expression of the vitamin D receptor, of estrogen and thyroid hormone receptor α- and β-isoforms, and of the androgen receptor in cultures of native mouse bone marrow and of stromal/osteoblastic cells. Bone. 1999;24:465–473. [DOI] [PubMed] [Google Scholar]
- 176. Gu WX, Stern PH, Madison LD, Du GG. Mutual up-regulation of thyroid hormone and parathyroid hormone receptors in rat osteoblastic osteosarcoma 17/2.8 cells. Endocrinology. 2001;142:157–164. [DOI] [PubMed] [Google Scholar]
- 177. Kasono K, Sato K, Han DC, Fujii Y, Tsushima T, Shizume K. Stimulation of alkaline phosphatase activity by thyroid hormone in mouse osteoblast-like cells (MC3T3-E1): a possible mechanism of hyperalkaline phosphatasia in hyperthyroidism. Bone Miner. 1988;4:355–363. [PubMed] [Google Scholar]
- 178. LeBron BA, Pekary AE, Mirell C, Hahn TJ, Hershman JM. Thyroid hormone 5′-deiodinase activity, nuclear binding, and effects on mitogenesis in UMR-106 osteoblastic osteosarcoma cells. J Bone Miner Res. 1989;4:173–178. [DOI] [PubMed] [Google Scholar]
- 179. Rizzoli R, Poser J, Bürgi U. Nuclear thyroid hormone receptors in cultured bone cells. Metabolism. 1986;35:71–74. [DOI] [PubMed] [Google Scholar]
- 180. Sato K, Han DC, Fujii Y, Tsushima T, Shizume K. Thyroid hormone stimulates alkaline phosphatase activity in cultured rat osteoblastic cells (ROS 17/2.8) through 3,5,3′-triiodo-L-thyronine nuclear receptors. Endocrinology. 1987;120:1873–1881. [DOI] [PubMed] [Google Scholar]
- 181. Williams GR, Bland R, Sheppard MC. Characterization of thyroid hormone (T3) receptors in three osteosarcoma cell lines of distinct osteoblast phenotype: interactions among T3, vitamin D3, and retinoid signaling. Endocrinology. 1994;135:2375–2385. [DOI] [PubMed] [Google Scholar]
- 182. Siddiqi A, Parsons MP, Lewis JL, Monson JP, Williams GR, Burrin JM. TR expression and function in human bone marrow stromal and osteoblast-like cells. J Clin Endocrinol Metab. 2002;87:906–914. [DOI] [PubMed] [Google Scholar]
- 183. Milne M, Kang MI, Cardona G, et al. Expression of multiple thyroid hormone receptor isoforms in rat femoral and vertebral bone and in bone marrow osteogenic cultures. J Cell Biochem. 1999;74:684–693. [DOI] [PubMed] [Google Scholar]
- 184. Bassett JH, Williams GR. The molecular actions of thyroid hormone in bone. Trends Endocrinol Metab. 2003;14:356–364. [DOI] [PubMed] [Google Scholar]
- 185. Bassett JH, Williams GR. The skeletal phenotypes of TRα and TRβ mutant mice. J Mol Endocrinol. 2009;42:269–282. [DOI] [PubMed] [Google Scholar]
- 186. Klaushofer K, Hoffmann O, Gleispach H, et al. Bone-resorbing activity of thyroid hormones is related to prostaglandin production in cultured neonatal mouse calvaria. J Bone Miner Res. 1989;4:305–312. [DOI] [PubMed] [Google Scholar]
- 187. Mundy GR, Shapiro JL, Bandelin JG, Canalis EM, Raisz LG. Direct stimulation of bone resorption by thyroid hormones. J Clin Invest. 1976;58:529–534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188. Allain TJ, Chambers TJ, Flanagan AM, McGregor AM. Tri-iodothyronine stimulates rat osteoclastic bone resorption by an indirect effect. J Endocrinol. 1992;133:327–331. [DOI] [PubMed] [Google Scholar]
- 189. Britto JM, Fenton AJ, Holloway WR, Nicholson GC. Osteoblasts mediate thyroid hormone stimulation of osteoclastic bone resorption. Endocrinology. 1994;134:169–176. [DOI] [PubMed] [Google Scholar]
- 190. Abu EO, Horner A, Teti A, Chatterjee VK, Compston JE. The localization of thyroid hormone receptor mRNAs in human bone. Thyroid. 2000;10:287–293. [DOI] [PubMed] [Google Scholar]
- 191. Urabe K, Hotokebuchi T, Oles KJ, et al. Inhibition of endochondral ossification during fracture repair in experimental hypothyroid rats. J Orthop Res. 1999;17:920–925. [DOI] [PubMed] [Google Scholar]
- 192. Ashton IK, Dekel S. Fracture repair in the Snell dwarf mouse. Br J Exp Pathol. 1983;64:479–486. [PMC free article] [PubMed] [Google Scholar]
- 193. Loftus MJ, Peterson LJ. Delayed healing of mandibular fracture in idiopathic myxedema. Oral Surg Oral Med Oral Pathol. 1979;47:233–237. [DOI] [PubMed] [Google Scholar]
- 194. Rumney TG, Wray JB. Role of the thyroid in fracture healing in the rat. Surg Forum. 1965;16:453–454. [PubMed] [Google Scholar]
- 195. Alini M, Kofsky Y, Wu W, Pidoux I, Poole AR. In serum-free culture thyroid hormones can induce full expression of chondrocyte hypertrophy leading to matrix calcification. J Bone Miner Res. 1996;11:105–113. [DOI] [PubMed] [Google Scholar]
- 196. Ballock RT, Reddi AH. Thyroxine is the serum factor that regulates morphogenesis of columnar cartilage from isolated chondrocytes in chemically defined medium. J Cell Biol. 1994;126:1311–1318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197. Ishikawa Y, Genge BR, Wuthier RE, Wu LN. Thyroid hormone inhibits growth and stimulates terminal differentiation of epiphyseal growth plate chondrocytes. J Bone Miner Res. 1998;13:1398–1411. [DOI] [PubMed] [Google Scholar]
- 198. Ohlsson C, Nilsson A, Isaksson O, Bentham J, Lindahl A. Effects of tri-iodothyronine and insulin-like growth factor-I (IGF-I) on alkaline phosphatase activity, [3H]thymidine incorporation and IGF-I receptor mRNA in cultured rat epiphyseal chondrocytes. J Endocrinol. 1992;135:115–123. [DOI] [PubMed] [Google Scholar]
- 199. Rosenthal AK, Henry LA. Thyroid hormones induce features of the hypertrophic phenotype and stimulate correlates of CPPD crystal formation in articular chondrocytes. J Rheumatol. 1999;26:395–401. [PubMed] [Google Scholar]
- 200. Ballock RT, Zhou X, Mink LM, Chen DH, Mita BC, Stewart MC. Expression of cyclin-dependent kinase inhibitors in epiphyseal chondrocytes induced to terminally differentiate with thyroid hormone. Endocrinology. 2000;141:4552–4557. [DOI] [PubMed] [Google Scholar]
- 201. Böhme K, Conscience-Egli M, Tschan T, Winterhalter KH, Bruckner P. Induction of proliferation or hypertrophy of chondrocytes in serum-free culture: the role of insulin-like growth factor-I, insulin, or thyroxine. J Cell Biol. 1992;116:1035–1042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202. Quarto R, Campanile G, Cancedda R, Dozin B. Thyroid hormone, insulin, and glucocorticoids are sufficient to support chondrocyte differentiation to hypertrophy: a serum-free analysis. J Cell Biol. 1992;119:989–995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203. Burch WM, Lebovitz HE. Triiodothyronine stimulates maturation of porcine growth-plate cartilage in vitro. J Clin Invest. 1982;70:496–504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204. Burch WM, Lebovitz HE. Triiodothyronine stimulation of in vitro growth and maturation of embryonic chick cartilage. Endocrinology. 1982;111:462–468. [DOI] [PubMed] [Google Scholar]
- 205. Himeno M, Enomoto H, Liu W, et al. Impaired vascular invasion of Cbfa1-deficient cartilage engrafted in the spleen. J Bone Miner Res. 2002;17:1297–1305. [DOI] [PubMed] [Google Scholar]
- 206. Miura M, Tanaka K, Komatsu Y, et al. Thyroid hormones promote chondrocyte differentiation in mouse ATDC5 cells and stimulate endochondral ossification in fetal mouse tibias through iodothyronine deiodinases in the growth plate. J Bone Miner Res. 2002;17:443–454. [DOI] [PubMed] [Google Scholar]
- 207. Mueller MB, Tuan RS. Functional characterization of hypertrophy in chondrogenesis of human mesenchymal stem cells. Arthritis Rheum. 2008;58:1377–1388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208. Wakita R, Izumi T, Itoman M. Thyroid hormone-induced chondrocyte terminal differentiation in rat femur organ culture. Cell Tissue Res. 1998;293:357–364. [DOI] [PubMed] [Google Scholar]
- 209. Mello MA, Tuan RS. Effects of TGF-β1 and triiodothyronine on cartilage maturation: in vitro analysis using long-term high-density micromass cultures of chick embryonic limb mesenchymal cells. J Orthop Res. 2006;24:2095–2105. [DOI] [PubMed] [Google Scholar]
- 210. Wang L, Shao YY, Ballock RT. Thyroid hormone interacts with the Wnt/β-catenin signaling pathway in the terminal differentiation of growth plate chondrocytes. J Bone Miner Res. 2007;22:1988–1995. [DOI] [PubMed] [Google Scholar]
- 211. Wang L, Shao YY, Ballock RT. Carboxypeptidase Z (CPZ) links thyroid hormone and Wnt signaling pathways in growth plate chondrocytes. J Bone Miner Res. 2009;24:265–273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212. Wang L, Shao YY, Ballock RT. Thyroid hormone-mediated growth and differentiation of growth plate chondrocytes involves IGF-1 modulation of β-catenin signaling. J Bone Miner Res. 2010;25:1138–1146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213. Minina E, Kreschel C, Naski MC, Ornitz DM, Vortkamp A. Interaction of FGF, Ihh/Pthlh, and BMP signaling integrates chondrocyte proliferation and hypertrophic differentiation. Dev Cell. 2002;3:439–449. [DOI] [PubMed] [Google Scholar]
- 214. St-Jacques B, Hammerschmidt M, McMahon AP. Indian hedgehog signaling regulates proliferation and differentiation of chondrocytes and is essential for bone formation. Genes Dev. 1999;13:2072–2086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215. Wang L, Shao YY, Ballock RT. Leptin synergizes with thyroid hormone signaling in promoting growth plate chondrocyte proliferation and terminal differentiation in vitro. Bone. 2011;48:1022–1027. [DOI] [PubMed] [Google Scholar]
- 216. Xing W, Cheng S, Wergedal J, Mohan S. Epiphyseal chondrocyte secondary ossification centers require thyroid hormone activation of Indian hedgehog and osterix signaling. J Bone Miner Res. 2014;29:2262–2275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217. Audhya TK, Gibson KD. Effects of medium composition and metabolic inhibitors on glycosaminoglycan synthesis in chick embryo cartilage and its stimulation by serum and triiodothyronine. Biochim Biophys Acta. 1976;437:364–376. [DOI] [PubMed] [Google Scholar]
- 218. Audhya TK, Segen BJ, Gibson KD. Stimulation of proteoglycan synthesis in chick embryo sternum by serum and L-3,5,3′-triiodothyronine. J Biol Chem. 1976;251:3763–3767. [PubMed] [Google Scholar]
- 219. Bassett JH, Swinhoe R, Chassande O, Samarut J, Williams GR. Thyroid hormone regulates heparan sulfate proteoglycan expression in the growth plate. Endocrinology. 2006;147:295–305. [DOI] [PubMed] [Google Scholar]
- 220. Froesch ER, Zapf J, Audhya TK, Ben-Porath E, Segen BJ, Gibson KD. Nonsuppressible insulin-like activity and thyroid hormones: major pituitary-dependent sulfation factors for chick embryo cartilage. Proc Natl Acad Sci USA. 1976;73:2904–2908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221. Gibson KD, Segen BJ, Audhya TK. The effect of β-D-xylosides on chondroitin sulphate biosynthesis in embryonic chicken cartilage in the absence of protein synthesis inhibitors. Biochem J. 1977;162:217–233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222. Makihira S, Yan W, Murakami H, et al. Thyroid hormone enhances aggrecanase-2/ADAM-TS5 expression and proteoglycan degradation in growth plate cartilage. Endocrinology. 2003;144:2480–2488. [DOI] [PubMed] [Google Scholar]
- 223. Pereira RC, Jorgetti V, Canalis E. Triiodothyronine induces collagenase-3 and gelatinase B expression in murine osteoblasts. Am J Physiol. 1999;277:E496–E504. [DOI] [PubMed] [Google Scholar]
- 224. Barnard JC, Williams AJ, Rabier B, et al. Thyroid hormones regulate fibroblast growth factor receptor signaling during chondrogenesis. Endocrinology. 2005;146:5568–5580. [DOI] [PubMed] [Google Scholar]
- 225. Karl A, Olbrich N, Pfeifer C, et al. Thyroid hormone-induced hypertrophy in mesenchymal stem cell chondrogenesis is mediated by bone morphogenetic protein-4. Tissue Eng Part A. 2014;20:178–188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226. Smith LB, Belanger JM, Oberbauer AM. Fibroblast growth factor receptor 3 effects on proliferation and telomerase activity in sheep growth plate chondrocytes. J Anim Sci Biotechnol. 2012;3:39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227. Cray JJ, Jr, Khaksarfard K, Weinberg SM, Elsalanty M, Yu JC. Effects of thyroxine exposure on osteogenesis in mouse calvarial pre-osteoblasts. PLoS One. 2013;8:e69067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228. Ernst M, Froesch ER. Triiodothyronine stimulates proliferation of osteoblast-like cells in serum-free culture. FEBS Lett. 1987;220:163–166. [DOI] [PubMed] [Google Scholar]
- 229. Huang BK, Golden LA, Tarjan G, Madison LD, Stern PH. Insulin-like growth factor I production is essential for anabolic effects of thyroid hormone in osteoblasts. J Bone Miner Res. 2000;15:188–197. [DOI] [PubMed] [Google Scholar]
- 230. Ishida H, Bellows CG, Aubin JE, Heersche JN. Tri-iodothyronine (T3) and dexamethasone interact to modulate osteoprogenitor cell differentiation in fetal rat calvaria cell cultures. Bone. 1995;16:545–549. [DOI] [PubMed] [Google Scholar]
- 231. Ohishi K, Ishida H, Nagata T, et al. Thyroid hormone suppresses the differentiation of osteoprogenitor cells to osteoblasts, but enhances functional activities of mature osteoblasts in cultured rat calvaria cells. J Cell Physiol. 1994;161:544–552. [DOI] [PubMed] [Google Scholar]
- 232. Schmid C, Schläpfer I, Futo E, et al. Triiodothyronine (T3) stimulates insulin-like growth factor (IGF)-1 and IGF binding protein (IGFBP)-2 production by rat osteoblasts in vitro. Acta Endocrinol (Copenh). 1992;126:467–473. [DOI] [PubMed] [Google Scholar]
- 233. Schmid C, Steiner T, Froesch ER. Triiodothyronine increases responsiveness of cultured rat bone cells to parathyroid hormone. Acta Endocrinol (Copenh). 1986;111:213–216. [DOI] [PubMed] [Google Scholar]
- 234. Fratzl-Zelman N, Hörandner H, Luegmayr E, et al. Effects of triiodothyronine on the morphology of cells and matrix, the localization of alkaline phosphatase, and the frequency of apoptosis in long-term cultures of MC3T3-E1 cells. Bone. 1997;20:225–236. [DOI] [PubMed] [Google Scholar]
- 235. Glantschnig H, Varga F, Klaushofer K. Thyroid hormone and retinoic acid induce the synthesis of insulin-like growth factor-binding protein-4 in mouse osteoblastic cells. Endocrinology. 1996;137:281–286. [DOI] [PubMed] [Google Scholar]
- 236. Glantschnig H, Varga F, Luegmayr E, Klaushofer K. Characterization of the mouse insulin-like growth factor binding protein 4 gene regulatory region and expression studies. DNA Cell Biol. 1998;17:51–60. [DOI] [PubMed] [Google Scholar]
- 237. Gouveia CH, Schultz JJ, Bianco AC, Brent GA. Thyroid hormone stimulation of osteocalcin gene expression in ROS 17/2.8 cells is mediated by transcriptional and post-transcriptional mechanisms. J Endocrinol. 2001;170:667–675. [DOI] [PubMed] [Google Scholar]
- 238. Klaushofer K, Varga F, Glantschnig H, et al. The regulatory role of thyroid hormones in bone cell growth and differentiation. J Nutr. 1995;125:1996S–2003S. [DOI] [PubMed] [Google Scholar]
- 239. Luegmayr E, Glantschnig H, Varga F, Klaushofer K. The organization of adherens junctions in mouse osteoblast-like cells (MC3T3-E1) and their modulation by triiodothyronine and 1,25-dihydroxyvitamin D3. Histochem Cell Biol. 2000;113:467–478. [DOI] [PubMed] [Google Scholar]
- 240. Luegmayr E, Varga F, Frank T, Roschger P, Klaushofer K. Effects of triiodothyronine on morphology, growth behavior, and the actin cytoskeleton in mouse osteoblastic cells (MC3T3–E1). Bone. 1996;18:591–599. [DOI] [PubMed] [Google Scholar]
- 241. Luegmayr E, Varga F, Glantschnig H, et al. 1,25-Dihydroxy vitamin D3 and tri-iodothyronine stimulate the expression of a protein immunologically related to osteocalcin. J Histochem Cytochem. 1998;46:477–486. [DOI] [PubMed] [Google Scholar]
- 242. Varga F, Luegmayr E, Fratzl-Zelman N, et al. Tri-iodothyronine inhibits multilayer formation of the osteoblastic cell line, MC3T3-E1, by promoting apoptosis. J Endocrinol. 1999;160:57–65. [DOI] [PubMed] [Google Scholar]
- 243. Varga F, Rumpler M, Klaushofer K. Thyroid hormones increase insulin-like growth factor mRNA levels in the clonal osteoblastic cell line MC3T3–E1. FEBS Lett. 1994;345:67–70. [DOI] [PubMed] [Google Scholar]
- 244. Varga F, Rumpler M, Luegmayr E, Fratzl-Zelman N, Glantschnig H, Klaushofer K. Triiodothyronine, a regulator of osteoblastic differentiation: depression of histone H4, attenuation of c-fos/c-jun, and induction of osteocalcin expression. Calcif Tissue Int. 1997;61:404–411. [DOI] [PubMed] [Google Scholar]
- 245. Williams GR, Bland R, Sheppard MC. Retinoids modify regulation of endogenous gene expression by vitamin D3 and thyroid hormone in three osteosarcoma cell lines. Endocrinology. 1995;136:4304–4314. [DOI] [PubMed] [Google Scholar]
- 246. Milne M, Kang MI, Quail JM, Baran DT. Thyroid hormone excess increases insulin-like growth factor I transcripts in bone marrow cell cultures: divergent effects on vertebral and femoral cell cultures. Endocrinology. 1998;139:2527–2534. [DOI] [PubMed] [Google Scholar]
- 247. Suwanwalaikorn S, Ongphiphadhanakul B, Braverman LE, Baran DT. Differential responses of femoral and vertebral bones to long-term excessive L-thyroxine administration in adult rats. Eur J Endocrinol. 1996;134:655–659. [DOI] [PubMed] [Google Scholar]
- 248. Suwanwalaikorn S, Van Auken M, Kang MI, Alex S, Braverman LE, Baran DT. Site selectivity of osteoblast gene expression response to thyroid hormone localized by in situ hybridization. Am J Physiol. 1997;272:E212–E217. [DOI] [PubMed] [Google Scholar]
- 249. Banovac K, Koren E. Triiodothyronine stimulates the release of membrane-bound alkaline phosphatase in osteoblastic cells. Calcif Tissue Int. 2000;67:460–465. [DOI] [PubMed] [Google Scholar]
- 250. Kato K, Otsuka T, Adachi S, et al. (-)-Epigallocatechin gallate inhibits thyroid hormone-stimulated osteocalcin synthesis in osteoblasts. Mol Med Rep. 2011;4:297–300. [DOI] [PubMed] [Google Scholar]
- 251. Milne M, Quail JM, Rosen CJ, Baran DT. Insulin-like growth factor binding proteins in femoral and vertebral bone marrow stromal cells: expression and regulation by thyroid hormone and dexamethasone. J Cell Biochem. 2001;81:229–240. [DOI] [PubMed] [Google Scholar]
- 252. O'Shea PJ, Kim DW, Logan JG, et al. Advanced bone formation in mice with a dominant-negative mutation in the thyroid hormone receptor β gene due to activation of Wnt/β-catenin protein signaling. J Biol Chem. 2012;287:17812–17822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 253. Saltó C, Kindblom JM, Johansson C, et al. Ablation of TRα2 and a concomitant overexpression of α1 yields a mixed hypo- and hyperthyroid phenotype in mice. Mol Endocrinol. 2001;15:2115–2128. [DOI] [PubMed] [Google Scholar]
- 254. Siddiqi A, Burrin JM, Wood DF, Monson JP. Tri-iodothyronine regulates the production of interleukin-6 and interleukin-8 in human bone marrow stromal and osteoblast-like cells. J Endocrinol. 1998;157:453–461. [DOI] [PubMed] [Google Scholar]
- 255. Stevens DA, Harvey CB, Scott AJ, et al. Thyroid hormone activates fibroblast growth factor receptor-1 in bone. Mol Endocrinol. 2003;17:1751–1766. [DOI] [PubMed] [Google Scholar]
- 256. Varga F, Rumpler M, Spitzer S, Karlic H, Klaushofer K. Osteocalcin attenuates T3- and increases vitamin D3-induced expression of MMP-13 in mouse osteoblasts. Endocr J. 2009;56:441–450. [DOI] [PubMed] [Google Scholar]
- 257. Varga F, Rumpler M, Zoehrer R, et al. T3 affects expression of collagen I and collagen cross-linking in bone cell cultures. Biochem Biophys Res Commun. 2010;402:180–185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 258. Varga F, Spitzer S, Klaushofer K. Triiodothyronine (T3) and 1,25-dihydroxyvitamin D3 (1,25D3) inversely regulate OPG gene expression in dependence of the osteoblastic phenotype. Calcif Tissue Int. 2004;74:382–387. [DOI] [PubMed] [Google Scholar]
- 259. Xing W, Govoni KE, Donahue LR, et al. Genetic evidence that thyroid hormone is indispensable for prepubertal insulin-like growth factor-I expression and bone acquisition in mice. J Bone Miner Res. 2012;27:1067–1079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 260. Qiu J, Ma XL, Wang X, Chen H, Huang BR. Insulin-like growth factor binding protein-6 interacts with the thyroid hormone receptor α1 and modulates the thyroid hormone-response in osteoblastic differentiation. Mol Cell Biochem. 2012;361:197–208. [DOI] [PubMed] [Google Scholar]
- 261. Asai S, Cao X, Yamauchi M, Funahashi K, Ishiguro N, Kambe F. Thyroid hormone non-genomically suppresses Src thereby stimulating osteocalcin expression in primary mouse calvarial osteoblasts. Biochem Biophys Res Commun. 2009;387:92–96. [DOI] [PubMed] [Google Scholar]
- 262. Ishisaki A, Tokuda H, Yoshida M, et al. Activation of p38 mitogen-activated protein kinase mediates thyroid hormone-stimulated osteocalcin synthesis in osteoblasts. Mol Cell Endocrinol. 2004;214:189–195. [DOI] [PubMed] [Google Scholar]
- 263. Kondo A, Otsuka T, Kato K, et al. AMP-activated protein kinase regulates thyroid hormone-stimulated osteocalcin synthesis in osteoblasts. Int J Mol Med. 2013;31:1457–1462. [DOI] [PubMed] [Google Scholar]
- 264. Kanno Y, Ishisaki A, Yoshida M, et al. Adenylyl cyclase-cAMP system inhibits thyroid hormone-stimulated osteocalcin synthesis in osteoblasts. Mol Cell Endocrinol. 2005;229:75–82. [DOI] [PubMed] [Google Scholar]
- 265. Kondo A, Tokuda H, Kato K, et al. Rho-kinase negatively regulates thyroid hormone-stimulated osteocalcin synthesis in osteoblasts. Biochimie. 2013;95:719–724. [DOI] [PubMed] [Google Scholar]
- 266. Miura M, Tanaka K, Komatsu Y, et al. A novel interaction between thyroid hormones and 1,25(OH)(2)D(3) in osteoclast formation. Biochem Biophys Res Commun. 2002;291:987–994. [DOI] [PubMed] [Google Scholar]
- 267. Kawaguchi H, Pilbeam CC, Raisz LG. Anabolic effects of 3,3′,5-triiodothyronine and triiodothyroacetic acid in cultured neonatal mouse parietal bones. Endocrinology. 1994;135:971–976. [DOI] [PubMed] [Google Scholar]
- 268. Kawaguchi H, Pilbeam CC, Woodiel FN, Raisz LG. Comparison of the effects of 3,5,3′-triiodothyroacetic acid and triiodothyronine on bone resorption in cultured fetal rat long bones and neonatal mouse calvariae. J Bone Miner Res. 1994;9:247–253. [DOI] [PubMed] [Google Scholar]
- 269. Lakatos P, Caplice MD, Khanna V, Stern PH. Thyroid hormones increase insulin-like growth factor I content in the medium of rat bone tissue. J Bone Miner Res. 1993;8:1475–1481. [DOI] [PubMed] [Google Scholar]
- 270. Stracke H, Rossol S, Schatz H. Alkaline phosphatase and insulin-like growth factor in fetal rat bone under the influence of thyroid hormones. Horm Metab Res. 1986;18:794. [DOI] [PubMed] [Google Scholar]
- 271. Tarjan G, Stern PH. Triiodothyronine potentiates the stimulatory effects of interleukin-1 β on bone resorption and medium interleukin-6 content in fetal rat limb bone cultures. J Bone Miner Res. 1995;10:1321–1326. [DOI] [PubMed] [Google Scholar]
- 272. Lakatos P, Stern PH. Effects of cyclosporins and transforming growth factor β 1 on thyroid hormone action in cultured fetal rat limb bones. Calcif Tissue Int. 1992;50:123–128. [DOI] [PubMed] [Google Scholar]
- 273. Kanatani M, Sugimoto T, Sowa H, Kobayashi T, Kanzawa M, Chihara K. Thyroid hormone stimulates osteoclast differentiation by a mechanism independent of RANKL-RANK interaction. J Cell Physiol. 2004;201:17–25. [DOI] [PubMed] [Google Scholar]
- 274. Saraiva PP, Teixeira SS, Padovani CR, Nogueira CR. Triiodothyronine (T3) does not induce Rankl expression in rat Ros 17/2.8 cells. Arq Bras Endocrinol Metabol. 2008;52:109–113. [DOI] [PubMed] [Google Scholar]
- 275. Lakatos P, Stern PH. Evidence for direct non-genomic effects of triiodothyronine on bone rudiments in rats: stimulation of the inositol phosphate second messenger system. Acta Endocrinol (Copenh). 1991;125:603–608. [DOI] [PubMed] [Google Scholar]
- 276. Marians RC, Ng L, Blair HC, Unger P, Graves PN, Davies TF. Defining thyrotropin-dependent and -independent steps of thyroid hormone synthesis by using thyrotropin receptor-null mice. Proc Natl Acad Sci USA. 2002;99:15776–15781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 277. Friedrichsen S, Christ S, Heuer H, et al. Regulation of iodothyronine deiodinases in the Pax8−/− mouse model of congenital hypothyroidism. Endocrinology. 2003;144:777–784. [DOI] [PubMed] [Google Scholar]
- 278. O'Shea PJ, Bassett JH, Sriskantharajah S, Ying H, Cheng SY, Williams GR. Contrasting skeletal phenotypes in mice with an identical mutation targeted to thyroid hormone receptor α1 or β. Mol Endocrinol. 2005;19:3045–3059. [DOI] [PubMed] [Google Scholar]
- 279. Boersma B, Otten BJ, Stoelinga GB, Wit JM. Catch-up growth after prolonged hypothyroidism. Eur J Pediatr. 1996;155:362–367. [DOI] [PubMed] [Google Scholar]
- 280. Marino R, Hegde A, Barnes KM, et al. Catch-up growth after hypothyroidism is caused by delayed growth plate senescence. Endocrinology. 2008;149:1820–1828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 281. Rivkees SA, Bode HH, Crawford JD. Long-term growth in juvenile acquired hypothyroidism: the failure to achieve normal adult stature. N Engl J Med. 1988;318:599–602. [DOI] [PubMed] [Google Scholar]
- 282. Mansouri A, Chowdhury K, Gruss P. Follicular cells of the thyroid gland require Pax8 gene function. Nat Genet. 1998;19:87–90. [DOI] [PubMed] [Google Scholar]
- 283. Beamer WJ, Eicher EM, Maltais LJ, Southard JL. Inherited primary hypothyroidism in mice. Science. 1981;212:61–63. [DOI] [PubMed] [Google Scholar]
- 284. Weiss RE, Chassande O, Koo EK, et al. Thyroid function and effect of aging in combined hetero/homozygous mice deficient in thyroid hormone receptors α and β genes. J Endocrinol. 2002;172:177–185. [DOI] [PubMed] [Google Scholar]
- 285. Bassett JH, O'Shea PJ, Sriskantharajah S, et al. Thyroid hormone excess rather than thyrotropin deficiency induces osteoporosis in hyperthyroidism. Mol Endocrinol. 2007;21:1095–1107. [DOI] [PubMed] [Google Scholar]
- 286. Sun L, Vukicevic S, Baliram R, et al. Intermittent recombinant TSH injections prevent ovariectomy-induced bone loss. Proc Natl Acad Sci USA. 2008;105:4289–4294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 287. Dumic-Cule I, Draca N, Luetic AT, et al. TSH prevents bone resorption and with calcitriol synergistically stimulates bone formation in rats with low levels of calciotropic hormones. Horm Metab Res. 2014;46:305–312. [DOI] [PubMed] [Google Scholar]
- 288. Liao XH, Di Cosmo C, Dumitrescu AM, et al. Distinct roles of deiodinases on the phenotype of Mct8 defect: a comparison of eight different mouse genotypes. Endocrinology. 2011;152:1180–1191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 289. Trajkovic M, Visser TJ, Mittag J, et al. Abnormal thyroid hormone metabolism in mice lacking the monocarboxylate transporter 8. J Clin Invest. 2007;117:627–635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 290. Mariotta L, Ramadan T, Singer D, et al. T-type amino acid transporter TAT1 (Slc16a10) is essential for extracellular aromatic amino acid homeostasis control. J Physiol. 2012;590:6413–6424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 291. Müller J, Mayerl S, Visser TJ, et al. Tissue-specific alterations in thyroid hormone homeostasis in combined Mct10 and Mct8 deficiency. Endocrinology. 2014;155:315–325. [DOI] [PubMed] [Google Scholar]
- 292. Mayerl S, Müller J, Bauer R, et al. Transporters MCT8 and OATP1C1 maintain murine brain thyroid hormone homeostasis. J Clin Invest. 2014;124:1987–1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 293. Mayerl S, Visser TJ, Darras VM, Horn S, Heuer H. Impact of Oatp1c1 deficiency on thyroid hormone metabolism and action in the mouse brain. Endocrinology. 2012;153:1528–1537. [DOI] [PubMed] [Google Scholar]
- 294. Berry MJ, Grieco D, Taylor BA, et al. Physiological and genetic analyses of inbred mouse strains with a type I iodothyronine 5′ deiodinase deficiency. J Clin Invest. 1993;92:1517–1528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 295. Christoffolete MA, Arrojo e Drigo R, Gazoni F, et al. Mice with impaired extrathyroidal thyroxine to 3,5,3′-triiodothyronine conversion maintain normal serum 3,5,3′-triiodothyronine concentrations. Endocrinology. 2007;148:954–960. [DOI] [PubMed] [Google Scholar]
- 296. Schoenmakers CH, Pigmans IG, Poland A, Visser TJ. Impairment of the selenoenzyme type I iodothyronine deiodinase in C3H/He mice. Endocrinology. 1993;132:357–361. [DOI] [PubMed] [Google Scholar]
- 297. Galton VA, Schneider MJ, Clark AS, St Germain DL. Life without thyroxine to 3,5,3′-triiodothyronine conversion: studies in mice devoid of the 5′-deiodinases. Endocrinology. 2009;150:2957–2963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 298. Galton VA, Wood ET, St Germain EA, et al. Thyroid hormone homeostasis and action in the type 2 deiodinase-deficient rodent brain during development. Endocrinology. 2007;148:3080–3088. [DOI] [PubMed] [Google Scholar]
- 299. Hernandez A, Martinez ME, Fiering S, Galton VA, St Germain D. Type 3 deiodinase is critical for the maturation and function of the thyroid axis. J Clin Invest. 2006;116:476–484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 300. Hernandez A, Martinez ME, Liao XH, et al. Type 3 deiodinase deficiency results in functional abnormalities at multiple levels of the thyroid axis. Endocrinology. 2007;148:5680–5687. [DOI] [PubMed] [Google Scholar]
- 301. Flamant F, Gauthier K. Thyroid hormone receptors: the challenge of elucidating isotype-specific functions and cell-specific response. Biochim Biophys Acta. 2013;1830:3900–3907. [DOI] [PubMed] [Google Scholar]
- 302. Flamant F, Samarut J. Thyroid hormone receptors: lessons from knockout and knock-in mutant mice. Trends Endocrinol Metab. 2003;14:85–90. [DOI] [PubMed] [Google Scholar]
- 303. O'Shea PJ, Williams GR. Insight into the physiological actions of thyroid hormone receptors from genetically modified mice. J Endocrinol. 2002;175:553–570. [DOI] [PubMed] [Google Scholar]
- 304. Wikström L, Johansson C, Saltó C, et al. Abnormal heart rate and body temperature in mice lacking thyroid hormone receptor alpha 1. EMBO J. 1998;17:455–461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 305. Göthe S, Wang Z, Ng L, et al. Mice devoid of all known thyroid hormone receptors are viable but exhibit disorders of the pituitary-thyroid axis, growth, and bone maturation. Genes Dev. 1999;13:1329–1341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 306. Kindblom JM, Gevers EF, Skrtic SM, et al. Increased adipogenesis in bone marrow but decreased bone mineral density in mice devoid of thyroid hormone receptors. Bone. 2005;36:607–616. [DOI] [PubMed] [Google Scholar]
- 307. Kindblom JM, Göthe S, Forrest D, et al. GH substitution reverses the growth phenotype but not the defective ossification in thyroid hormone receptor α 1−/−β−/− mice. J Endocrinol. 2001;171:15–22. [DOI] [PubMed] [Google Scholar]
- 308. Wallis K, Dudazy S, van Hogerlinden M, Nordström K, Mittag J, Vennström B. The thyroid hormone receptor α1 protein is expressed in embryonic postmitotic neurons and persists in most adult neurons. Mol Endocrinol. 2010;24:1904–1916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 309. Ng L, Rüsch A, Amma LL, et al. Suppression of the deafness and thyroid dysfunction in Thrb-null mice by an independent mutation in the Thra thyroid hormone receptor α gene. Hum Mol Genet. 2001;10:2701–2708. [DOI] [PubMed] [Google Scholar]
- 310. Fraichard A, Chassande O, Plateroti M, et al. The T3R α gene encoding a thyroid hormone receptor is essential for post-natal development and thyroid hormone production. EMBO J. 1997;16:4412–4420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 311. Gauthier K, Chassande O, Plateroti M, et al. Different functions for the thyroid hormone receptors TRα and TRβ in the control of thyroid hormone production and post-natal development. EMBO J. 1999;18:623–631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 312. Bassett JH, Nordström K, Boyde A, et al. Thyroid status during skeletal development determines adult bone structure and mineralization. Mol Endocrinol. 2007;21:1893–1904. [DOI] [PubMed] [Google Scholar]
- 313. Gauthier K, Plateroti M, Harvey CB, et al. Genetic analysis reveals different functions for the products of the thyroid hormone receptor α locus. Mol Cell Biol. 2001;21:4748–4760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 314. Flamant F, Poguet AL, Plateroti M, et al. Congenital hypothyroid Pax8(−/−) mutant mice can be rescued by inactivating the TRα gene. Mol Endocrinol. 2002;16:24–32. [DOI] [PubMed] [Google Scholar]
- 315. Mittag J, Friedrichsen S, Heuer H, Polsfuss S, Visser TJ, Bauer K. Athyroid Pax8−/− mice cannot be rescued by the inactivation of thyroid hormone receptor α1. Endocrinology. 2005;146:3179–3184. [DOI] [PubMed] [Google Scholar]
- 316. Parrilla R, Mixson AJ, McPherson JA, McClaskey JH, Weintraub BD. Characterization of seven novel mutations of the c-erbA β gene in unrelated kindreds with generalized thyroid hormone resistance. Evidence for two “hot spot” regions of the ligand binding domain. J Clin Invest. 1991;88:2123–2130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 317. Kaneshige M, Suzuki H, Kaneshige K, et al. A targeted dominant negative mutation of the thyroid hormone α 1 receptor causes increased mortality, infertility, and dwarfism in mice. Proc Natl Acad Sci USA. 2001;98:15095–15100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 318. O'Shea PJ, Bassett JH, Cheng SY, Williams GR. Characterization of skeletal phenotypes of TRα1 and TRβ mutant mice: implications for tissue thyroid status and T3 target gene expression. Nucl Recept Signal. 2006;4:e011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 319. Bassett JH, Boyde A, Zikmund T, et al. Thyroid hormone receptor α mutation causes a severe and thyroxine-resistant skeletal dysplasia in female mice. Endocrinology. 2014;155:3699–3712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 320. Quignodon L, Vincent S, Winter H, Samarut J, Flamant F. A point mutation in the activation function 2 domain of thyroid hormone receptor α1 expressed after CRE-mediated recombination partially recapitulates hypothyroidism. Mol Endocrinol. 2007;21:2350–2360. [DOI] [PubMed] [Google Scholar]
- 321. Desjardin C, Charles C, Benoist-Lasselin C, et al. Chondrocytes play a major role in the stimulation of bone growth by thyroid hormone. Endocrinology. 2014;155:3123–3135. [DOI] [PubMed] [Google Scholar]
- 322. Segni M, Leonardi E, Mazzoncini B, Pucarelli I, Pasquino AM. Special features of Graves' disease in early childhood. Thyroid. 1999;9:871–877. [DOI] [PubMed] [Google Scholar]
- 323. Kaneshige M, Kaneshige K, Zhu X, et al. Mice with a targeted mutation in the thyroid hormone β receptor gene exhibit impaired growth and resistance to thyroid hormone. Proc Natl Acad Sci USA. 2000;97:13209–13214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 324. Zhou YX, Xu X, Chen L, Li C, Brodie SG, Deng CX. A Pro250Arg substitution in mouse Fgfr1 causes increased expression of Cbfa1 and premature fusion of calvarial sutures. Hum Mol Genet. 2000;9:2001–2008. [DOI] [PubMed] [Google Scholar]
- 325. Ornitz DM, Marie PJ. FGF signaling pathways in endochondral and intramembranous bone development and human genetic disease. Genes Dev. 2002;16:1446–1465. [DOI] [PubMed] [Google Scholar]
- 326. Guigon CJ, Zhao L, Lu C, Willingham MC, Cheng SY. Regulation of β-catenin by a novel nongenomic action of thyroid hormone β receptor. Mol Cell Biol. 2008;28:4598–4608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 327. Tsourdi E, Rijntjes E, Köhrle J, Hofbauer LC, Rauner M. Hyperthyroidism and hypothyroidism in male mice and their effects on bone mass, bone turnover, and the Wnt inhibitors sclerostin and dickkopf-1. Endocrinology. 2015;156:3517–3527. [DOI] [PubMed] [Google Scholar]
- 328. Papadimitriou A, Papadimitriou DT, Papadopoulou A, Nicolaidou P, Fretzayas A. Low TSH levels are not associated with osteoporosis in childhood. Eur J Endocrinol. 2007;157:221–223. [DOI] [PubMed] [Google Scholar]
- 329. Kleinau G, Biebermann H. Constitutive activities in the thyrotropin receptor: regulation and significance. Adv Pharmacol. 2014;70:81–119. [DOI] [PubMed] [Google Scholar]
- 330. Vassart G, Costagliola S. G protein-coupled receptors: mutations and endocrine diseases. Nat Rev Endocrinol. 2011;7:362–372. [DOI] [PubMed] [Google Scholar]
- 331. Abramowicz MJ, Duprez L, Parma J, Vassart G, Heinrichs C. Familial congenital hypothyroidism due to inactivating mutation of the thyrotropin receptor causing profound hypoplasia of the thyroid gland. J Clin Invest. 1997;99:3018–3024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 332. Biebermann H, Schöneberg T, Krude H, Schultz G, Gudermann T, Grüters A. Mutations of the human thyrotropin receptor gene causing thyroid hypoplasia and persistent congenital hypothyroidism. J Clin Endocrinol Metab. 1997;82:3471–3480. [DOI] [PubMed] [Google Scholar]
- 333. Clifton-Bligh RJ, Gregory JW, Ludgate M, et al. Two novel mutations in the thyrotropin (TSH) receptor gene in a child with resistance to TSH. J Clin Endocrinol Metab. 1997;82:1094–1100. [DOI] [PubMed] [Google Scholar]
- 334. Sunthornthepvarakui T, Gottschalk ME, Hayashi Y, Refetoff S. Brief report: resistance to thyrotropin caused by mutations in the thyrotropin-receptor gene. N Engl J Med. 1995;332:155–160. [DOI] [PubMed] [Google Scholar]
- 335. Bretones P, Duprez L, Parma J, David M, Vassart G, Rodien P. A familial case of congenital hypothyroidism caused by a homozygous mutation of the thyrotropin receptor gene. Thyroid. 2001;11:977–980. [DOI] [PubMed] [Google Scholar]
- 336. Gagné N, Parma J, Deal C, Vassart G, Van Vliet G. Apparent congenital athyreosis contrasting with normal plasma thyroglobulin levels and associated with inactivating mutations in the thyrotropin receptor gene: are athyreosis and ectopic thyroid distinct entities? J Clin Endocrinol Metab. 1998;83:1771–1775. [DOI] [PubMed] [Google Scholar]
- 337. Paschke R, Ludgate M. The thyrotropin receptor in thyroid diseases. N Engl J Med. 1997;337:1675–1681. [DOI] [PubMed] [Google Scholar]
- 338. Führer D, Wonerow P, Willgerodt H, Paschke R. Identification of a new thyrotropin receptor germline mutation (Leu629Phe) in a family with neonatal onset of autosomal dominant nonautoimmune hyperthyroidism. J Clin Endocrinol Metab. 1997;82:4234–4238. [DOI] [PubMed] [Google Scholar]
- 339. de Roux N, Polak M, Couet J, et al. A neomutation of the thyroid-stimulating hormone receptor in a severe neonatal hyperthyroidism. J Clin Endocrinol Metab. 1996;81:2023–2026. [DOI] [PubMed] [Google Scholar]
- 340. Esapa CT, Duprez L, Ludgate M, et al. A novel thyrotropin receptor mutation in an infant with severe thyrotoxicosis. Thyroid. 1999;9:1005–1010. [DOI] [PubMed] [Google Scholar]
- 341. Holzapfel HP, Wonerow P, von Petrykowski W, Henschen M, Scherbaum WA, Paschke R. Sporadic congenital hyperthyroidism due to a spontaneous germline mutation in the thyrotropin receptor gene. J Clin Endocrinol Metab. 1997;82:3879–3884. [DOI] [PubMed] [Google Scholar]
- 342. Supornsilchai V, Sahakitrungruang T, Wongjitrat N, Wacharasindhu S, Suphapeetiporn K, Shotelersuk V. Expanding clinical spectrum of non-autoimmune hyperthyroidism due to an activating germline mutation, p.M453T, in the thyrotropin receptor gene. Clin Endocrinol (Oxf). 2009;70:623–628. [DOI] [PubMed] [Google Scholar]
- 343. Grüters A, Schöneberg T, Biebermann H, et al. Severe congenital hyperthyroidism caused by a germ-line neo mutation in the extracellular portion of the thyrotropin receptor. J Clin Endocrinol Metab. 1998;83:1431–1436. [DOI] [PubMed] [Google Scholar]
- 344. Kopp P, Jameson JL, Roe TF. Congenital nonautoimmune hyperthyroidism in a nonidentical twin caused by a sporadic germline mutation in the thyrotropin receptor gene. Thyroid. 1997;7:765–770. [DOI] [PubMed] [Google Scholar]
- 345. Chawla R, Alden TD, Bizhanova A, Kadakia R, Brickman W, Kopp PA. Squamosal suture craniosynostosis due to hyperthyroidism caused by an activating thyrotropin receptor mutation (T632I). Thyroid. 2015;25:1167–1172. [DOI] [PubMed] [Google Scholar]
- 346. Dumitrescu AM, Liao XH, Abdullah MS, et al. Mutations in SECISBP2 result in abnormal thyroid hormone metabolism. Nat Genet. 2005;37:1247–1252. [DOI] [PubMed] [Google Scholar]
- 347. Azevedo MF, Barra GB, Naves LA, et al. Selenoprotein-related disease in a young girl caused by nonsense mutations in the SBP2 gene. J Clin Endocrinol Metab. 2010;95:4066–4071. [DOI] [PubMed] [Google Scholar]
- 348. Di Cosmo C, McLellan N, Liao XH, et al. Clinical and molecular characterization of a novel selenocysteine insertion sequence-binding protein 2 (SBP2) gene mutation (R128X). J Clin Endocrinol Metab. 2009;94:4003–4009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 349. Schoenmakers E, Agostini M, Mitchell C, et al. Mutations in the selenocysteine insertion sequence-binding protein 2 gene lead to a multisystem selenoprotein deficiency disorder in humans. J Clin Invest. 2010;120:4220–4235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 350. Hamajima T, Mushimoto Y, Kobayashi H, Saito Y, Onigata K. Novel compound heterozygous mutations in the SBP2 gene: characteristic clinical manifestations and the implications of GH and triiodothyronine in longitudinal bone growth and maturation. Eur J Endocrinol. 2012;166:757–764. [DOI] [PubMed] [Google Scholar]
- 351. Refetoff S, DeWind LT, DeGroot LJ. Familial syndrome combining deaf-mutism, stuppled epiphyses, goiter and abnormally high PBI: possible target organ refractoriness to thyroid hormone. J Clin Endocrinol Metab. 1967;27:279–294. [DOI] [PubMed] [Google Scholar]
- 352. Sakurai A, Takeda K, Ain K, et al. Generalized resistance to thyroid hormone associated with a mutation in the ligand-binding domain of the human thyroid hormone receptor β. Proc Natl Acad Sci USA. 1989;86:8977–8981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 353. Usala SJ, Tennyson GE, Bale AE, et al. A base mutation of the C-erbA β thyroid hormone receptor in a kindred with generalized thyroid hormone resistance. Molecular heterogeneity in two other kindreds. J Clin Invest. 1990;85:93–100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 354. Dumitrescu AM, Refetoff S. The syndromes of reduced sensitivity to thyroid hormone. Biochim Biophys Acta. 2013;1830:3987–4003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 355. Refetoff S, Weiss RE, Usala SJ. The syndromes of resistance to thyroid hormone. Endocr Rev. 1993;14:348–399. [DOI] [PubMed] [Google Scholar]
- 356. Weiss RE. “They have ears but do not hear” (Psalms 135:17): non-thyroid hormone receptor β (non-TRβ) resistance to thyroid hormone. Thyroid. 2008;18:3–5. [DOI] [PubMed] [Google Scholar]
- 357. Magner JA, Petrick P, Menezes-Ferreira MM, Stelling M, Weintraub BD. Familial generalized resistance to thyroid hormones: report of three kindreds and correlation of patterns of affected tissues with the binding of [125I] triiodothyronine to fibroblast nuclei. J Endocrinol Invest. 1986;9:459–470. [DOI] [PubMed] [Google Scholar]
- 358. Smallridge RC, Parker RA, Wiggs EA, Rajagopal KR, Fein HG. Thyroid hormone resistance in a large kindred: physiologic, biochemical, pharmacologic, and neuropsychologic studies. Am J Med. 1989;86:289–296. [DOI] [PubMed] [Google Scholar]
- 359. Magalhães PK, Rodrigues Dare GL, Rodrigues Dos Santos S, Nogueira CR, de Castro M, Zanini Maciel LM. Clinical features and genetic analysis of four Brazilian kindreds with resistance to thyroid hormone. Clin Endocrinol (Oxf). 2007;67:748–753. [DOI] [PubMed] [Google Scholar]
- 360. Bode HH, Danon M, Weintraub BD, Maloof F, Crawford JD. Partial target organ resistance to thyroid hormone. J Clin Invest. 1973;52:776–782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 361. Weiss RE, Refetoff S. Resistance to thyroid hormone. Rev Endocr Metab Disord. 2000;1:97–108. [DOI] [PubMed] [Google Scholar]
- 362. Refetoff S, Dumitrescu AM. Syndromes of reduced sensitivity to thyroid hormone: genetic defects in hormone receptors, cell transporters and deiodination. Best Pract Res Clin Endocrinol Metab. 2007;21:277–305. [DOI] [PubMed] [Google Scholar]
- 363. Brucker-Davis F, Skarulis MC, Grace MB, et al. Genetic and clinical features of 42 kindreds with resistance to thyroid hormone. The National Institutes of Health Prospective Study. Ann Intern Med. 1995;123:572–583. [DOI] [PubMed] [Google Scholar]
- 364. Kvistad PH, Løvås K, Boman H, Myking OL. Retarded bone growth in thyroid hormone resistance. A clinical study of a large family with a novel thyroid hormone receptor mutation. Eur J Endocrinol. 2004;150:425–430. [DOI] [PubMed] [Google Scholar]
- 365. Refetoff S. Syndromes of thyroid hormone resistance. Am J Physiol. 1982;243:E88–E98. [DOI] [PubMed] [Google Scholar]
- 366. Furlanetto TW, Kopp P, Peccin S, Gu WX, Jameson JL. A novel mutation (M310L) in the thyroid hormone receptor β causing resistance to thyroid hormone in a Brazilian kindred and a neonate. Mol Genet Metab. 2000;71:520–526. [DOI] [PubMed] [Google Scholar]
- 367. Phillips SA, Rotman-Pikielny P, Lazar J, et al. Extreme thyroid hormone resistance in a patient with a novel truncated TR mutant. J Clin Endocrinol Metab. 2001;86:5142–5147. [DOI] [PubMed] [Google Scholar]
- 368. Ono S, Schwartz ID, Mueller OT, Root AW, Usala SJ, Bercu BB. Homozygosity for a dominant negative thyroid hormone receptor gene responsible for generalized resistance to thyroid hormone. J Clin Endocrinol Metab. 1991;73:990–994. [DOI] [PubMed] [Google Scholar]
- 369. Ortiga-Carvalho TM, Sidhaye AR, Wondisford FE. Thyroid hormone receptors and resistance to thyroid hormone disorders. Nat Rev Endocrinol. 2014;10:582–591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 370. Usala SJ, Menke JB, Watson TL, et al. A homozygous deletion in the c-erbA β thyroid hormone receptor gene in a patient with generalized thyroid hormone resistance: isolation and characterization of the mutant receptor. Mol Endocrinol. 1991;5:327–335. [DOI] [PubMed] [Google Scholar]
- 371. Weiss RE, Refetoff S. Effect of thyroid hormone on growth. Lessons from the syndrome of resistance to thyroid hormone. Endocrinol Metab Clin North Am. 1996;25:719–730. [DOI] [PubMed] [Google Scholar]
- 372. Rodrigues Dare GL, Ribeiro Magalhaes PK, de Castro M, Zanini Maciel LM. Peripheral parameters of thyroid hormone action in resistance to thyroid hormone syndrome: a focus on mineral metabolism. Thyroid. 2009;19:785–787. [DOI] [PubMed] [Google Scholar]
- 373. Weiss RE, Dumitrescu A, Refetoff S. Approach to the patient with resistance to thyroid hormone and pregnancy. J Clin Endocrinol Metab. 2010;95:3094–3102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 374. Cardoso LF, de Paula FJ, Maciel LM. Resistance to thyroid hormone due to mutations in the THRB gene impairs bone mass and affects calcium and phosphorus homeostasis. Bone. 2014;67:222–227. [DOI] [PubMed] [Google Scholar]
- 375. Menzaghi C, Balsamo A, Di Paola R, et al. Association between an R338L mutation in the thyroid hormone receptor-β gene and thyrotoxic features in two unrelated kindreds with resistance to thyroid hormone. Thyroid. 1999;9:1–6. [DOI] [PubMed] [Google Scholar]
- 376. Behr M, Ramsden DB, Loos U. Deoxyribonucleic acid binding and transcriptional silencing by a truncated c-erbA β 1 thyroid hormone receptor identified in a severely retarded patient with resistance to thyroid hormone. J Clin Endocrinol Metab. 1997;82:1081–1087. [DOI] [PubMed] [Google Scholar]
- 377. Mäenpää J, Liewendahl K. Peripheral insensitivity to thyroid hormones in a euthyroid girl with goitre. Arch Dis Child. 1980;55:207–212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 378. Catargi B, Monsaingeon M, Bex-Bachellerie V, et al. A novel thyroid hormone receptor-β mutation, not anticipated to occur in resistance to thyroid hormone, causes variable phenotypes. Horm Res. 2002;57:137–142. [DOI] [PubMed] [Google Scholar]
- 379. Sato H, Koike Y, Honma M, Yagame M, Ito K. Evaluation of thyroid hormone action in a case of generalized resistance to thyroid hormone with chronic thyroiditis: discovery of a novel heterozygous missense mutation (G347A). Endocr J. 2007;54:727–732. [DOI] [PubMed] [Google Scholar]
- 380. Refetoff S, Bassett JH, Beck-Peccoz P, et al. Classification and proposed nomenclature for inherited defects of thyroid hormone action, cell transport, and metabolism. J Clin Endocrinol Metab. 2014;99:768–770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 381. Vennström B, Mittag J, Wallis K. Severe psychomotor and metabolic damages caused by a mutant thyroid hormone receptor α 1 in mice: can patients with a similar mutation be found and treated? Acta Paediatrica. 2008;97:1605–1610. [DOI] [PubMed] [Google Scholar]
- 382. Bochukova E, Schoenmakers N, Agostini M, et al. A mutation in the thyroid hormone receptor alpha gene. N Engl J Med. 2012;366:243–249. [DOI] [PubMed] [Google Scholar]
- 383. Moran C, Schoenmakers N, Agostini M, et al. An adult female with resistance to thyroid hormone mediated by defective thyroid hormone receptor α. J Clin Endocrinol Metab. 2013;98:4254–4261. [DOI] [PubMed] [Google Scholar]
- 384. van Mullem A, van Heerebeek R, Chrysis D, et al. Clinical phenotype and mutant TRα1. N Engl J Med. 2012;366:1451–1453. [DOI] [PubMed] [Google Scholar]
- 385. van Mullem AA, Chrysis D, Eythimiadou A, et al. Clinical phenotype of a new type of thyroid hormone resistance caused by a mutation of the TRα1 receptor: consequences of LT4 treatment. J Clin Endocrinol Metab. 2013;98:3029–3038. [DOI] [PubMed] [Google Scholar]
- 386. Schoenmakers N, Moran C, Peeters RP, Visser T, Gurnell M, Chatterjee K. Resistance to thyroid hormone mediated by defective thyroid hormone receptor α. Biochim Biophys Acta. 2013;1830:4004–4008. [DOI] [PubMed] [Google Scholar]
- 387. van Mullem AA, Visser TJ, Peeters RP. Clinical consequences of mutations in thyroid hormone receptor-α1. Eur Thyroid J. 2014;3:17–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 388. Tylki-Szymańńska A, Acuna-Hidalgo R, Krajewska-Walasek M, et al. Thyroid hormone resistance syndrome due to mutations in the thyroid hormone receptor alpha gene (THRA). J Med Genet. 2015;52:312–316. [DOI] [PubMed] [Google Scholar]
- 389. Moran C, Agostini M, Visser WE, et al. Resistance to thyroid hormone caused by a mutation in thyroid hormone receptor (TR)α1 and TRα2: clinical, biochemical, and genetic analyses of three related patients. Lancet Diabetes Endocrinol. 2014;2:619–626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 390. Espiard S, Savagner F, Flamant F, et al. A novel mutation in THRA gene associated with an atypical phenotype of resistance to thyroid hormone. J Clin Endocrinol Metab. 2015;100:2841–2848. [DOI] [PubMed] [Google Scholar]
- 391. Williams GR. Role of thyroid hormone receptor-α1 in endochondral ossification. Endocrinology. 2014;155:2747–2750. [DOI] [PubMed] [Google Scholar]
- 392. Hüffmeier U, Tietze HU, Rauch A. Severe skeletal dysplasia caused by undiagnosed hypothyroidism. Eur J Med Genet. 2007;50:209–215. [DOI] [PubMed] [Google Scholar]
- 393. Salerno M, Lettiero T, Esposito-del Puente A, et al. Effect of long-term L-thyroxine treatment on bone mineral density in young adults with congenital hypothyroidism. Eur J Endocrinol. 2004;151:689–694. [DOI] [PubMed] [Google Scholar]
- 394. Salerno M, Micillo M, Di Maio S, et al. Longitudinal growth, sexual maturation and final height in patients with congenital hypothyroidism detected by neonatal screening. Eur J Endocrinol. 2001;145:377–383. [DOI] [PubMed] [Google Scholar]
- 395. Rasmussen SA, Yazdy MM, Carmichael SL, Jamieson DJ, Canfield MA, Honein MA. Maternal thyroid disease as a risk factor for craniosynostosis. Obstet Gynecol. 2007;110:369–377. [DOI] [PubMed] [Google Scholar]
- 396. Fröhlich E, Wahl R. Mechanisms in endocrinology: impact of isolated TSH levels in and out of normal range on different tissues. Eur J Endocrinol. 2015;174:R29–R41. [DOI] [PubMed] [Google Scholar]
- 397. Heemstra KA, Hamdy NA, Romijn JA, Smit JW. The effects of thyrotropin-suppressive therapy on bone metabolism in patients with well-differentiated thyroid carcinoma. Thyroid. 2006;16:583–591. [DOI] [PubMed] [Google Scholar]
- 398. Murphy E, Williams GR. The thyroid and the skeleton. Clin Endocrinol (Oxf). 2004;61:285–298. [DOI] [PubMed] [Google Scholar]
- 399. Taylor PN, Razvi S, Pearce SH, Dayan CM. Clinical review: a review of the clinical consequences of variation in thyroid function within the reference range. J Clin Endocrinol Metab. 2013;98:3562–3571. [DOI] [PubMed] [Google Scholar]
- 400. Chin KY, Ima-Nirwana S, Mohamed IN, Aminuddin A, Johari MH, Ngah WZ. Thyroid-stimulating hormone is significantly associated with bone health status in men. Int J Med Sci. 2013;10:857–863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 401. El Hadidy el HM, Ghonaim M, El Gawad SSh, El Atta MA. Impact of severity, duration, and etiology of hyperthyroidism on bone turnover markers and bone mineral density in men. BMC Endocr Disord. 2011;11:15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 402. Kim BJ, Lee SH, Bae SJ, et al. The association between serum thyrotropin (TSH) levels and bone mineral density in healthy euthyroid men. Clin Endocrinol (Oxf). 2010;73:396–403. [DOI] [PubMed] [Google Scholar]
- 403. Kim MK, Yun KJ, Kim MH, et al. The effects of thyrotropin-suppressing therapy on bone metabolism in patients with well-differentiated thyroid carcinoma. Bone. 2015;71:101–105. [DOI] [PubMed] [Google Scholar]
- 404. Lakatos P, Foldes J, Horvath C, et al. Serum interleukin-6 and bone metabolism in patients with thyroid function disorders. J Clin Endocrinol Metab. 1997;82:78–81. [DOI] [PubMed] [Google Scholar]
- 405. Kim S, Jung J, Jung JH, Kim SK, Kim RB, Hahm JR. Risk factors of bone mass loss at the lumbar spine: a longitudinal study in healthy Korean pre- and perimenopausal women older than 40 years. PLoS One. 2015;10:e0136283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 406. Baqi L, Payer J, Killinger Z, et al. Thyrotropin versus thyroid hormone in regulating bone density and turnover in premenopausal women. Endocr Regul. 2010;44:57–63. [DOI] [PubMed] [Google Scholar]
- 407. Baqi L, Payer J, Killinger Z, et al. The level of TSH appeared favourable in maintaining bone mineral density in postmenopausal women. Endocr Regul. 2010;44:9–15. [DOI] [PubMed] [Google Scholar]
- 408. Christy AL, D'Souza V, Babu RP, et al. Utility of C-terminal telopeptide in evaluating levothyroxine replacement therapy-induced bone loss. Biomarker Insights. 2014;9:1–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 409. Lee MY, Park JH, Bae KS, et al. Bone mineral density and bone turnover markers in patients on long-term suppressive levothyroxine therapy for differentiated thyroid cancer. Ann Surg Treat Res. 2014;86:55–60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 410. Sugitani I, Fujimoto Y. Effect of postoperative thyrotropin suppressive therapy on bone mineral density in patients with papillary thyroid carcinoma: a prospective controlled study. Surgery. 2011;150:1250–1257. [DOI] [PubMed] [Google Scholar]
- 411. Tournis S, Antoniou JD, Liakou CG, et al. Volumetric bone mineral density and bone geometry assessed by peripheral quantitative computed tomography in women with differentiated thyroid cancer under TSH suppression. Clin Endocrinol (Oxf). 2015;82:197–204. [DOI] [PubMed] [Google Scholar]
- 412. Moon JH, Jung KY, Kim KM, et al. The effect of thyroid stimulating hormone suppressive therapy on bone geometry in the hip area of patients with differentiated thyroid carcinoma. Bone. 2016;83:104–110. [DOI] [PubMed] [Google Scholar]
- 413. Eriksen EF, Mosekilde L, Melsen F. Kinetics of trabecular bone resorption and formation in hypothyroidism: evidence for a positive balance per remodeling cycle. Bone. 1986;7:101–108. [DOI] [PubMed] [Google Scholar]
- 414. Mosekilde L, Eriksen EF, Charles P. Effects of thyroid hormones on bone and mineral metabolism. Endocrinol Metab Clin North Am. 1990;19:35–63. [PubMed] [Google Scholar]
- 415. Mosekilde L, Melsen F. Effect of antithyroid treatment on calcium-phosphorus metabolism in hyperthyroidism. II: Bone histomorphometry. Acta Endocrinol (Copenh). 1978;87:751–758. [DOI] [PubMed] [Google Scholar]
- 416. Mosekilde L, Melsen F. Morphometric and dynamic studies of bone changes in hypothyroidism. Acta Pathol Microbiol Scand A. 1978;86:56–62. [DOI] [PubMed] [Google Scholar]
- 417. Senturk T, Kozaci LD, Kok F, Kadikoylu G, Bolaman Z. Proinflammatory cytokine levels in hyperthyroidism. Clin Invest Med. 2003;26:58–63. [PubMed] [Google Scholar]
- 418. Garnero P, Vassy V, Bertholin A, Riou JP, Delmas PD. Markers of bone turnover in hyperthyroidism and the effects of treatment. J Clin Endocrinol Metab. 1994;78:955–959. [DOI] [PubMed] [Google Scholar]
- 419. Guo CY, Weetman AP, Eastell R. Longitudinal changes of bone mineral density and bone turnover in postmenopausal women on thyroxine. Clin Endocrinol (Oxf). 1997;46:301–307. [DOI] [PubMed] [Google Scholar]
- 420. Harvey RD, McHardy KC, Reid IW, et al. Measurement of bone collagen degradation in hyperthyroidism and during thyroxine replacement therapy using pyridinium cross-links as specific urinary markers. J Clin Endocrinol Metab. 1991;72:1189–1194. [DOI] [PubMed] [Google Scholar]
- 421. Toivonen J, Tähtelä R, Laitinen K, Risteli J, Välimäki MJ. Markers of bone turnover in patients with differentiated thyroid cancer with and following withdrawal of thyroxine suppressive therapy. Eur J Endocrinol. 1998;138:667–673. [DOI] [PubMed] [Google Scholar]
- 422. Mikosch P, Kerschan-Schindl K, Woloszczuk W, et al. High cathepsin K levels in men with differentiated thyroid cancer on suppressive L-thyroxine therapy. Thyroid. 2008;18:27–33. [DOI] [PubMed] [Google Scholar]
- 423. Moser E, Sikjaer T, Mosekilde L, Rejnmark L. Bone indices in thyroidectomized patients on long-term substitution therapy with levothyroxine assessed by DXA and HR-pQCT. J Thyroid Res. 2015;2015:796871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 424. Williams GR. Does serum TSH level have thyroid hormone independent effects on bone turnover? Nat Clin Pract Endocrinol Metab. 2009;5:10–11. [DOI] [PubMed] [Google Scholar]
- 425. Giusti M, Cecoli F, Ghiara C, et al. Recombinant human thyroid stimulating hormone does not acutely change serum osteoprotegerin and soluble receptor activator of nuclear factor-κβ ligand in patients under evaluation for differentiated thyroid carcinoma. Hormones (Athens). 2007;6:304–313. [DOI] [PubMed] [Google Scholar]
- 426. Martini G, Gennari L, De Paola V, et al. The effects of recombinant TSH on bone turnover markers and serum osteoprotegerin and RANKL levels. Thyroid. 2008;18:455–460. [DOI] [PubMed] [Google Scholar]
- 427. Mazziotti G, Sorvillo F, Piscopo M, et al. Recombinant human TSH modulates in vivo C-telopeptides of type-1 collagen and bone alkaline phosphatase, but not osteoprotegerin production in postmenopausal women monitored for differentiated thyroid carcinoma. J Bone Miner Res. 2005;20:480–486. [DOI] [PubMed] [Google Scholar]
- 428. Karga H, Papaioannou G, Polymeris A, et al. The effects of recombinant human TSH on bone turnover in patients after thyroidectomy. J Bone Miner Metab. 2010;28:35–41. [DOI] [PubMed] [Google Scholar]
- 429. Tsourdi E, Wallaschofski H, Rauner M, et al. Thyrotropin serum levels are differentially associated with biochemical markers of bone turnover and stiffness in women and men: results from the SHIP cohorts. Osteoporos Int. 2016;27:719–727. [DOI] [PubMed] [Google Scholar]
- 430. Zofkova I, Hill M. Biochemical markers of bone remodeling correlate negatively with circulating TSH in postmenopausal women. Endocr Regul. 2008;42:121–127. [PubMed] [Google Scholar]
- 431. Lin JD, Pei D, Hsia TL, et al. The relationship between thyroid function and bone mineral density in euthyroid healthy subjects in Taiwan. Endocr Res. 2011;36:1–8. [DOI] [PubMed] [Google Scholar]
- 432. Murphy E, Glüer CC, Reid DM, et al. Thyroid function within the upper normal range is associated with reduced bone mineral density and an increased risk of nonvertebral fractures in healthy euthyroid postmenopausal women. J Clin Endocrinol Metab. 2010;95:3173–3181. [DOI] [PubMed] [Google Scholar]
- 433. van Rijn LE, Pop VJ, Williams GR. Low bone mineral density is related to high physiological levels of free thyroxine in peri-menopausal women. Eur J Endocrinol. 2014;170:461–468. [DOI] [PubMed] [Google Scholar]
- 434. Noh HM, Park YS, Lee J, Lee W. A cross-sectional study to examine the correlation between serum TSH levels and the osteoporosis of the lumbar spine in healthy women with normal thyroid function. Osteoporos Int. 2015;26:997–1003. [DOI] [PubMed] [Google Scholar]
- 435. Morris MS. The association between serum thyroid-stimulating hormone in its reference range and bone status in postmenopausal American women. Bone. 2007;40:1128–1134. [DOI] [PubMed] [Google Scholar]
- 436. Kim DJ, Khang YH, Koh JM, Shong YK, Kim GS. Low normal TSH levels are associated with low bone mineral density in healthy postmenopausal women. Clin Endocrinol (Oxf). 2006;64:86–90. [DOI] [PubMed] [Google Scholar]
- 437. Hwangbo Y, Kim JH, Kim SW, et al. High-normal free thyroxine levels are associated with low trabecular bone scores in euthyroid postmenopausal women. Osteoporos Int. 2016;27:457–462. [DOI] [PubMed] [Google Scholar]
- 438. van der Deure WM, Uitterlinden AG, Hofman A, et al. Effects of serum TSH and FT4 levels and the TSHR-Asp727Glu polymorphism on bone: the Rotterdam Study. Clin Endocrinol (Oxf). 2008;68:175–181. [DOI] [PubMed] [Google Scholar]
- 439. Grimnes G, Emaus N, Joakimsen RM, Figenschau Y, Jorde R. The relationship between serum TSH and bone mineral density in men and postmenopausal women: the Tromsø study. Thyroid. 2008;18:1147–1155. [DOI] [PubMed] [Google Scholar]
- 440. Roef G, Lapauw B, Goemaere S, et al. Thyroid hormone status within the physiological range affects bone mass and density in healthy men at the age of peak bone mass. Eur J Endocrinol. 2011;164:1027–1034. [DOI] [PubMed] [Google Scholar]
- 441. Leader A, Ayzenfeld RH, Lishner M, Cohen E, Segev D, Hermoni D. Thyrotropin levels within the lower normal range are associated with an increased risk of hip fractures in euthyroid women, but not men, over the age of 65 years. J Clin Endocrinol Metab. 2014;99:2665–2673. [DOI] [PubMed] [Google Scholar]
- 442. Svare A, Nilsen TI, Asvold BO, et al. Does thyroid function influence fracture risk? Prospective data from the HUNT2 study, Norway. Eur J Endocrinol. 2013;169:845–852. [DOI] [PubMed] [Google Scholar]
- 443. Mazziotti G, Porcelli T, Patelli I, Vescovi PP, Giustina A. Serum TSH values and risk of vertebral fractures in euthyroid post-menopausal women with low bone mineral density. Bone. 2010;46:747–751. [DOI] [PubMed] [Google Scholar]
- 444. Finigan J, Greenfield DM, Blumsohn A, et al. Risk factors for vertebral and nonvertebral fracture over 10 years: a population-based study in women. J Bone Miner Res. 2008;23:75–85. [DOI] [PubMed] [Google Scholar]
- 445. Eriksen EF, Mosekilde L, Melsen F. Trabecular bone remodeling and bone balance in hyperthyroidism. Bone. 1985;6:421–428. [DOI] [PubMed] [Google Scholar]
- 446. Stamato FJ, Amarante EC, Furlanetto RP. Effect of combined treatment with calcitonin on bone densitometry of patients with treated hypothyroidism. Rev Assoc Med Bras. 2000;46:177–181. [DOI] [PubMed] [Google Scholar]
- 447. Vestergaard P, Mosekilde L. Fractures in patients with hyperthyroidism and hypothyroidism: a nationwide follow-up study in 16,249 patients. Thyroid. 2002;12:411–419. [DOI] [PubMed] [Google Scholar]
- 448. González-Rodríguez LA, Felici-Giovanini ME, Haddock L. Thyroid dysfunction in an adult female population: a population-based study of Latin American Vertebral Osteoporosis Study (LAVOS) - Puerto Rico site. P R Health Sci J. 2013;32:57–62. [PMC free article] [PubMed] [Google Scholar]
- 449. Paul TL, Kerrigan J, Kelly AM, Braverman LE, Baran DT. Long-term L-thyroxine therapy is associated with decreased hip bone density in premenopausal women. JAMA. 1988;259:3137–3141. [PubMed] [Google Scholar]
- 450. Kung AW, Pun KK. Bone mineral density in premenopausal women receiving long-term physiological doses of levothyroxine. JAMA. 1991;265:2688–2691. [PubMed] [Google Scholar]
- 451. Cummings SR, Nevitt MC, Browner WS, et al. Risk factors for hip fracture in white women. Study of Osteoporotic Fractures Research Group. N Engl J Med. 1995;332:767–773. [DOI] [PubMed] [Google Scholar]
- 452. Melton LJ, 3rd, Ardila E, Crowson CS, O'Fallon WM, Khosla S. Fractures following thyroidectomy in women: a population-based cohort study. Bone. 2000;27:695–700. [DOI] [PubMed] [Google Scholar]
- 453. Van Den Eeden SK, Barzilay JI, Ettinger B, Minkoff J. Thyroid hormone use and the risk of hip fracture in women > or = 65 years: a case-control study. J Womens Health (Larchmt). 2003;12:27–31. [DOI] [PubMed] [Google Scholar]
- 454. Ahmed LA, Schirmer H, Berntsen GK, Fønnebø V, Joakimsen RM. Self-reported diseases and the risk of non-vertebral fractures: the Tromsø study. Osteoporos Int. 2006;17:46–53. [DOI] [PubMed] [Google Scholar]
- 455. Flynn RW, Bonellie SR, Jung RT, MacDonald TM, Morris AD, Leese GP. Serum thyroid-stimulating hormone concentration and morbidity from cardiovascular disease and fractures in patients on long-term thyroxine therapy. J Clin Endocrinol Metab. 2010;95:186–193. [DOI] [PubMed] [Google Scholar]
- 456. Vestergaard P, Rejnmark L, Mosekilde L. Influence of hyper- and hypothyroidism, and the effects of treatment with antithyroid drugs and levothyroxine on fracture risk. Calcif Tissue Int. 2005;77:139–144. [DOI] [PubMed] [Google Scholar]
- 457. Vestergaard P, Weeke J, Hoeck HC, et al. Fractures in patients with primary idiopathic hypothyroidism. Thyroid. 2000;10:335–340. [DOI] [PubMed] [Google Scholar]
- 458. Mazziotti G, Mormando M, Cristiano A, et al. Association between l-thyroxine treatment, GH deficiency, and radiological vertebral fractures in patients with adult-onset hypopituitarism. Eur J Endocrinol. 2014;170:893–899. [DOI] [PubMed] [Google Scholar]
- 459. Ko YJ, Kim JY, Lee J, et al. Levothyroxine dose and fracture risk according to the osteoporosis status in elderly women. J Prev Med Public Health. 2014;47:36–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 460. Meier C, Beat M, Guglielmetti M, Christ-Crain M, Staub JJ, Kraenzlin M. Restoration of euthyroidism accelerates bone turnover in patients with subclinical hypothyroidism: a randomized controlled trial. Osteoporos Int. 2004;15:209–216. [DOI] [PubMed] [Google Scholar]
- 461. Garin MC, Arnold AM, Lee JS, Robbins J, Cappola AR. Subclinical thyroid dysfunction and hip fracture and bone mineral density in older adults: the cardiovascular health study. J Clin Endocrinol Metab. 2014;99:2657–2664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 462. Waring AC, Harrison S, Fink HA, et al. A prospective study of thyroid function, bone loss, and fractures in older men: the MrOS study. J Bone Miner Res. 2013;28:472–479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 463. Lee JS, Buzková P, Fink HA, et al. Subclinical thyroid dysfunction and incident hip fracture in older adults. Arch Intern Med. 2010;170:1876–1883. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 464. Blum MR, Bauer DC, Collet TH, et al. Subclinical thyroid dysfunction and fracture risk: a meta-analysis. JAMA. 2015;313:2055–2065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 465. Karner I, Hrgović Z, Sijanović S, et al. Bone mineral density changes and bone turnover in thyroid carcinoma patients treated with supraphysiologic doses of thyroxine. Eur J Med Res. 2005;10:480–488. [PubMed] [Google Scholar]
- 466. Reverter JL, Holgado S, Alonso N, Salinas I, Granada ML, Sanmartí A. Lack of deleterious effect on bone mineral density of long-term thyroxine suppressive therapy for differentiated thyroid carcinoma. Endocr Relat Cancer. 2005;12:973–981. [DOI] [PubMed] [Google Scholar]
- 467. Tauchmanovà L, Nuzzo V, Del Puente A, et al. Reduced bone mass detected by bone quantitative ultrasonometry and DEXA in pre- and postmenopausal women with endogenous subclinical hyperthyroidism. Maturitas. 2004;48:299–306. [DOI] [PubMed] [Google Scholar]
- 468. Franklyn JA, Betteridge J, Daykin J, et al. Long-term thyroxine treatment and bone mineral density. Lancet. 1992;340:9–13. [DOI] [PubMed] [Google Scholar]
- 469. Kung AW, Yeung SS. Prevention of bone loss induced by thyroxine suppressive therapy in postmenopausal women: the effect of calcium and calcitonin. J Clin Endocrinol Metab. 1996;81:1232–1236. [DOI] [PubMed] [Google Scholar]
- 470. Jódar E, Begoña López M, García L, Rigopoulou D, Martínez G, Hawkins F. Bone changes in pre- and postmenopausal women with thyroid cancer on levothyroxine therapy: evolution of axial and appendicular bone mass. Osteoporos Int. 1998;8:311–316. [DOI] [PubMed] [Google Scholar]
- 471. Vadiveloo T, Donnan PT, Cochrane L, Leese GP. The Thyroid Epidemiology, Audit, and Research Study (TEARS): morbidity in patients with endogenous subclinical hyperthyroidism. J Clin Endocrinol Metab. 2011;96:1344–1351. [DOI] [PubMed] [Google Scholar]
- 472. Turner MR, Camacho X, Fischer HD, et al. Levothyroxine dose and risk of fractures in older adults: nested case-control study. BMJ. 2011;342:d2238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 473. Bauer DC, Ettinger B, Nevitt MC, Stone KL. Risk for fracture in women with low serum levels of thyroid-stimulating hormone. Ann Intern Med. 2001;134:561–568. [DOI] [PubMed] [Google Scholar]
- 474. Abrahamsen B, Jørgensen HL, Laulund AS, Nybo M, Brix TH, Hegedüs L. Low serum thyrotropin level and duration of suppression as a predictor of major osteoporotic fractures-the OPENTHYRO register cohort. J Bone Miner Res. 2014;29:2040–2050. [DOI] [PubMed] [Google Scholar]
- 475. Abrahamsen B, Jørgensen HL, Laulund AS, et al. The excess risk of major osteoporotic fractures in hypothyroidism is driven by cumulative hyperthyroid as opposed to hypothyroid time: an observational register-based time-resolved cohort analysis. J Bone Miner Res. 2015;30:898–905. [DOI] [PubMed] [Google Scholar]
- 476. Jamal SA, Leiter RE, Bayoumi AM, Bauer DC, Cummings SR. Clinical utility of laboratory testing in women with osteoporosis. Osteoporos Int. 2005;16:534–540. [DOI] [PubMed] [Google Scholar]
- 477. Wang LY, Smith AW, Palmer FL, et al. Thyrotropin suppression increases the risk of osteoporosis without decreasing recurrence in ATA low- and intermediate-risk patients with differentiated thyroid carcinoma. Thyroid. 2015;25:300–307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 478. Faber J, Galløe AM. Changes in bone mass during prolonged subclinical hyperthyroidism due to L-thyroxine treatment: a meta-analysis. Eur J Endocrinol. 1994;130:350–356. [DOI] [PubMed] [Google Scholar]
- 479. Quan ML, Pasieka JL, Rorstad O. Bone mineral density in well-differentiated thyroid cancer patients treated with suppressive thyroxine: a systematic overview of the literature. J Surg Oncol. 2002;79:62–69; discussion 69–70. [DOI] [PubMed] [Google Scholar]
- 480. Uzzan B, Campos J, Cucherat M, Nony P, Boissel JP, Perret GY. Effects on bone mass of long term treatment with thyroid hormones: a meta-analysis. J Clin Endocrinol Metab. 1996;81:4278–4289. [DOI] [PubMed] [Google Scholar]
- 481. Wirth CD, Blum MR, da Costa BR, et al. Subclinical thyroid dysfunction and the risk for fractures: a systematic review and meta-analysis. Ann Intern Med. 2014;161:189–199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 482. Udayakumar N, Chandrasekaran M, Rasheed MH, Suresh RV, Sivaprakash S. Evaluation of bone mineral density in thyrotoxicosis. Singapore Med J. 2006;47:947–950. [PubMed] [Google Scholar]
- 483. Bours SP, van Geel TA, Geusens PP, et al. Contributors to secondary osteoporosis and metabolic bone diseases in patients presenting with a clinical fracture. J Clin Endocrinol Metab. 2011;96:1360–1367. [DOI] [PubMed] [Google Scholar]
- 484. Patel KV, Brennan KL, Brennan ML, Jupiter DC, Shar A, Davis ML. Association of a modified frailty index with mortality after femoral neck fracture in patients aged 60 years and older. Clin Orthop Relat Res. 2014;472:1010–1017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 485. Bauer DC, Nevitt MC, Ettinger B, Stone K. Low thyrotropin levels are not associated with bone loss in older women: a prospective study. J Clin Endocrinol Metab. 1997;82:2931–2936. [DOI] [PubMed] [Google Scholar]
- 486. Franklyn JA, Maisonneuve P, Sheppard MC, Betteridge J, Boyle P. Mortality after the treatment of hyperthyroidism with radioactive iodine. N Engl J Med. 1998;338:712–718. [DOI] [PubMed] [Google Scholar]
- 487. Wejda B, Hintze G, Katschinski B, Olbricht T, Benker G. Hip fractures and the thyroid: a case-control study. J Intern Med. 1995;237:241–247. [DOI] [PubMed] [Google Scholar]
- 488. Leese GP, Jung RT, Guthrie C, Waugh N, Browning MC. Morbidity in patients on L-thyroxine: a comparison of those with a normal TSH to those with a suppressed TSH. Clin Endocrinol (Oxf). 1992;37:500–503. [DOI] [PubMed] [Google Scholar]
- 489. Vestergaard P, Mosekilde L. Hyperthyroidism, bone mineral, and fracture risk–a meta-analysis. Thyroid. 2003;13:585–593. [DOI] [PubMed] [Google Scholar]
- 490. Estrada K, Styrkarsdottir U, Evangelou E, et al. Genome-wide meta-analysis identifies 56 bone mineral density loci and reveals 14 loci associated with risk of fracture. Nat Genet. 2012;44:491–501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 491. Ralston SH, Uitterlinden AG. Genetics of osteoporosis. Endocr Rev. 2010;31:629–662. [DOI] [PubMed] [Google Scholar]
- 492. Zhang L, Choi HJ, Estrada K, et al. Multistage genome-wide association meta-analyses identified two new loci for bone mineral density. Hum Mol Genet. 2014;23:1923–1933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 493. Liu RD, Chen RX, Li WR, et al. The Glu727 allele of thyroid stimulating hormone receptor gene is associated with osteoporosis. N Am J Med Sci. 2012;4:300–304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 494. Heemstra KA, van der Deure WM, Peeters RP, et al. Thyroid hormone independent associations between serum TSH levels and indicators of bone turnover in cured patients with differentiated thyroid carcinoma. Eur J Endocrinol. 2008;159:69–76. [DOI] [PubMed] [Google Scholar]
- 495. Yerges LM, Klei L, Cauley JA, et al. Candidate gene analysis of femoral neck trabecular and cortical volumetric bone mineral density in older men. J Bone Miner Res. 2010;25:330–338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 496. Yerges LM, Klei L, Cauley JA, et al. High-density association study of 383 candidate genes for volumetric BMD at the femoral neck and lumbar spine among older men. J Bone Miner Res. 2009;24:2039–2049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 497. Medici M, Rivadeneira F, van der Deure WM, et al. A large-scale population-based analysis of common genetic variation in the thyroid hormone receptor α locus and bone. Thyroid. 2012;22:223–224. [DOI] [PubMed] [Google Scholar]
- 498. Gogakos A, Logan JG, Waung JA, et al. THRA and DIO2 mutations are unlikely to be a common cause of increased bone mineral density in euthyroid post-menopausal women. Eur J Endocrinol. 2014;170:637–644. [DOI] [PubMed] [Google Scholar]
- 499. Heemstra KA, Hoftijzer H, van der Deure WM, et al. The type 2 deiodinase Thr92Ala polymorphism is associated with increased bone turnover and decreased femoral neck bone mineral density. J Bone Miner Res. 2010;25:1385–1391. [DOI] [PubMed] [Google Scholar]
- 500. Verloop H, Dekkers OM, Peeters RP, Schoones JW, Smit JW. Genetics in endocrinology: genetic variation in deiodinases: a systematic review of potential clinical effects in humans. Eur J Endocrinol. 2014;171:R123–R135. [DOI] [PubMed] [Google Scholar]
- 501. Goldring MB. Insight into the function of DIO2, a susceptibility gene in human osteoarthritis, as an inducer of cartilage damage in a rat model: is there a role for chondrocyte hypertrophy? Osteoarthritis Cartilage. 2013;21:643–645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 502. Meulenbelt I, Min JL, Bos S, et al. Identification of DIO2 as a new susceptibility locus for symptomatic osteoarthritis. Hum Mol Genet. 2008;17:1867–1875. [DOI] [PubMed] [Google Scholar]
- 503. Callegaro A, Meulenbelt I, Kloppenburg M, Slagboom PE, Houwing-Duistermaat JJ. Allele-sharing statistics using information on family history. Ann Hum Genet. 2010;74:547–554. [DOI] [PubMed] [Google Scholar]
- 504. Meulenbelt I, Bos SD, Chapman K, et al. Meta-analyses of genes modulating intracellular T3 bio-availability reveal a possible role for the DIO3 gene in osteoarthritis susceptibility. Ann Rheum Dis. 2011;70:164–167. [DOI] [PubMed] [Google Scholar]
- 505. Kerkhof HJ, Lories RJ, Meulenbelt I, et al. A genome-wide association study identifies an osteoarthritis susceptibility locus on chromosome 7q22. Arthritis Rheum. 2010;62:499–510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 506. Evangelou E, Kerkhof HJ, Styrkarsdottir U, et al. A meta-analysis of genome-wide association studies identifies novel variants associated with osteoarthritis of the hip. Ann Rheum Dis. 2014;73:2130–2136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 507. Rodriguez-Fontenla C, Calaza M, Evangelou E, et al. Assessment of osteoarthritis candidate genes in a meta-analysis of nine genome-wide association studies. Arthritis Rheumatol. 2014;66:940–949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 508. Bos SD, Bovée JV, Duijnisveld BJ, et al. Increased type II deiodinase protein in OA-affected cartilage and allelic imbalance of OA risk polymorphism rs225014 at DIO2 in human OA joint tissues. Ann Rheum Dis. 2012;71:1254–1258. [DOI] [PubMed] [Google Scholar]
- 509. Bomer N, den Hollander W, Ramos YF, et al. Underlying molecular mechanisms of DIO2 susceptibility in symptomatic osteoarthritis. Ann Rheum Dis. 2015;74:1571–1579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 510. Nagase H, Nagasawa Y, Tachida Y, et al. Deiodinase 2 upregulation demonstrated in osteoarthritis patients cartilage causes cartilage destruction in tissue-specific transgenic rats. Osteoarthritis Cartilage. 2013;21:514–523. [DOI] [PubMed] [Google Scholar]
- 511. Waarsing JH, Kloppenburg M, Slagboom PE, et al. Osteoarthritis susceptibility genes influence the association between hip morphology and osteoarthritis. Arthritis Rheum. 2011;63:1349–1354. [DOI] [PubMed] [Google Scholar]
- 512. Waung JA, Bassett JH, Williams GR. Adult mice lacking the type 2 iodothyronine deiodinase have increased subchondral bone but normal articular cartilage. Thyroid. 2015;25:269–277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 513. Bomer N, Cornelis FM, Ramos YF, et al. The effect of forced exercise on knee joints in Dio2-/- mice: type II iodothyronine deiodinase-deficient mice are less prone to develop OA-like cartilage damage upon excessive mechanical stress. Ann Rheum Dis. 2016;75:571–577. [DOI] [PubMed] [Google Scholar]
- 514. Carvalho-Bianco SD, Kim BW, Zhang JX, et al. Chronic cardiac-specific thyrotoxicosis increases myocardial β-adrenergic responsiveness. Mol Endocrinol. 2004;18:1840–1849. [DOI] [PubMed] [Google Scholar]
- 515. Cheng AW, Bolognesi M, Kraus VB. DIO2 modifies inflammatory responses in chondrocytes. Osteoarthritis Cartilage. 2012;20:440–445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 516. Roman-Blas JA, Jimenez SA. NF-κB as a potential therapeutic target in osteoarthritis and rheumatoid arthritis. Osteoarthritis Cartilage. 2006;14:839–848. [DOI] [PubMed] [Google Scholar]
- 517. Zeöld A, Pormüller L, Dentice M, et al. Metabolic instability of type 2 deiodinase is transferable to stable proteins independently of subcellular localization. J Biol Chem. 2006;281:31538–31543. [DOI] [PubMed] [Google Scholar]
- 518. Dreier R. Hypertrophic differentiation of chondrocytes in osteoarthritis: the developmental aspect of degenerative joint disorders. Arthritis Res Ther. 2010;12:216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 519. Lin AC, Seeto BL, Bartoszko JM, et al. Modulating hedgehog signaling can attenuate the severity of osteoarthritis. Nat Med. 2009;15:1421–1425. [DOI] [PubMed] [Google Scholar]
- 520. Glasson SS, Askew R, Sheppard B, et al. Deletion of active ADAMTS5 prevents cartilage degradation in a murine model of osteoarthritis. Nature. 2005;434:644–648. [DOI] [PubMed] [Google Scholar]
- 521. Stanton H, Rogerson FM, East CJ, et al. ADAMTS5 is the major aggrecanase in mouse cartilage in vivo and in vitro. Nature. 2005;434:648–652. [DOI] [PubMed] [Google Scholar]
- 522. Billinghurst RC, Dahlberg L, Ionescu M, et al. Enhanced cleavage of type II collagen by collagenases in osteoarthritic articular cartilage. J Clin Invest. 1997;99:1534–1545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 523. Chen-An P, Andreassen KV, Henriksen K, Li Y, Karsdal MA, Bay-Jensen AC. The inhibitory effect of salmon calcitonin on tri-iodothyronine induction of early hypertrophy in articular cartilage. PLoS One. 2012;7:e40081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 524. Chen-An P, Andreassen KV, Henriksen K, Karsdal MA, Bay-Jensen AC. Investigation of chondrocyte hypertrophy and cartilage calcification in a full-depth articular cartilage explants model. Rheumatol Int. 2013;33:401–411. [DOI] [PubMed] [Google Scholar]
- 525. Randau TM, Schildberg FA, Alini M, et al. The effect of dexamethasone and triiodothyronine on terminal differentiation of primary bovine chondrocytes and chondrogenically differentiated mesenchymal stem cells. PLoS One. 2013;8:e72973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 526. Rosenthal AK, Heinkel D, Gohr CM. Thyroxine stimulates transglutaminase activity in articular chondrocytes. Osteoarthritis Cartilage. 2003;11:463–470. [DOI] [PubMed] [Google Scholar]
- 527. Jiang J, Leong NL, Mung JC, Hidaka C, Lu HH. Interaction between zonal populations of articular chondrocytes suppresses chondrocyte mineralization and this process is mediated by PTHrP. Osteoarthritis Cartilage. 2008;16:70–82. [DOI] [PubMed] [Google Scholar]
- 528. Canani LH, Capp C, Dora JM, et al. The type 2 deiodinase A/G (Thr92Ala) polymorphism is associated with decreased enzyme velocity and increased insulin resistance in patients with type 2 diabetes mellitus. J Clin Endocrinol Metab. 2005;90:3472–3478. [DOI] [PubMed] [Google Scholar]
- 529. Stein SA, Oates EL, Hall CR, et al. Identification of a point mutation in the thyrotropin receptor of the hyt/hyt hypothyroid mouse. Mol Endocrinol. 1994;8:129–138. [DOI] [PubMed] [Google Scholar]
- 530. Tinnikov A, Nordström K, Thorén P, et al. Retardation of post-natal development caused by a negatively acting thyroid hormone receptor α1. EMBO J. 2002;21:5079–5087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 531. Liu YY, Schultz JJ, Brent GA. A thyroid hormone receptor α gene mutation (P398H) is associated with visceral adiposity and impaired catecholamine-stimulated lipolysis in mice. J Biol Chem. 2003;278:38913–38920. [DOI] [PubMed] [Google Scholar]
- 532. Hayashi Y, Xie J, Weiss RE, Pohlenz J, Refetoff S. Selective pituitary resistance to thyroid hormone produced by expression of a mutant thyroid hormone receptor β gene in the pituitary gland of transgenic mice. Biochem Biophys Res Commun. 1998;245:204–210. [DOI] [PubMed] [Google Scholar]
- 533. Abel ED, Kaulbach HC, Campos-Barros A, et al. Novel insight from transgenic mice into thyroid hormone resistance and the regulation of thyrotropin. J Clin Invest. 1999;103:271–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 534. Wong R, Vasilyev VV, Ting YT, et al. Transgenic mice bearing a human mutant thyroid hormone β 1 receptor manifest thyroid function anomalies, weight reduction, and hyperactivity. Mol Med. 1997;3:303–314. [PMC free article] [PubMed] [Google Scholar]
- 535. Zhu XG, Kaneshige M, Parlow AF, et al. Expression of the mutant thyroid hormone receptor PV in the pituitary of transgenic mice leads to weight reduction. Thyroid. 1999;9:1137–1145. [DOI] [PubMed] [Google Scholar]
- 536. Schneider MJ, Fiering SN, Pallud SE, Parlow AF, St Germain DL, Galton VA. Targeted disruption of the type 2 selenodeiodinase gene (DIO2) results in a phenotype of pituitary resistance to T4. Mol Endocrinol. 2001;15:2137–2148. [DOI] [PubMed] [Google Scholar]
- 537. Dumitrescu AM, Liao XH, Weiss RE, Millen K, Refetoff S. Tissue-specific thyroid hormone deprivation and excess in monocarboxylate transporter (mct) 8-deficient mice. Endocrinology. 2006;147:4036–4043. [DOI] [PubMed] [Google Scholar]
- 538. Morte B, Ceballos A, Diez D, et al. Thyroid hormone-regulated mouse cerebral cortex genes are differentially dependent on the source of the hormone: a study in monocarboxylate transporter-8- and deiodinase-2-deficient mice. Endocrinology. 2010;151:2381–2387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 539. Kopp P, van Sande J, Parma J, et al. Brief report: congenital hyperthyroidism caused by a mutation in the thyrotropin-receptor gene. N Engl J Med. 1995;332:150–154. [DOI] [PubMed] [Google Scholar]
- 540. Schneider G, Keiser HR, Bardin CW. Peripheral resistance to thyroxine: a cause of short stature in a boy without goitre. Clin Endocrinol (Oxf). 1975;4:111–118. [DOI] [PubMed] [Google Scholar]
- 541. Takeda K, Sakurai A, DeGroot LJ, Refetoff S. Recessive inheritance of thyroid hormone resistance caused by complete deletion of the protein-coding region of the thyroid hormone receptor-β gene. J Clin Endocrinol Metab. 1992;74:49–55. [DOI] [PubMed] [Google Scholar]
- 542. Sakurai A, Miyamoto T, Hughes IA, DeGroot LJ. Characterization of a novel mutant human thyroid hormone receptor β in a family with hereditary thyroid hormone resistance. Clin Endocrinol (Oxf). 1993;38:29–38. [DOI] [PubMed] [Google Scholar]
- 543. Adams M, Matthews C, Collingwood TN, Tone Y, Beck-Peccoz P, Chatterjee KK. Genetic analysis of 29 kindreds with generalized and pituitary resistance to thyroid hormone. Identification of thirteen novel mutations in the thyroid hormone receptor β gene. J Clin Invest. 1994;94:506–515. [DOI] [PMC free article] [PubMed] [Google Scholar]