

Abstract
Gastro-entero-pancreatic neuroendocrine neoplasms (GEP-NENs) represents a various family of rare tumours. Surgery is the first choice in GEP-NENs patients with localized disease whilst in the metastatic setting many other treatment options are available. Somatostatin analogues are indicated for symptoms control in functioning tumours. Furthermore they may be effective to inhibit tumour progression. GEP-NENs pathogenesis has been extensively studied in the last years therefore several driver mutations pathway genes have been identified as crucial factors in their tumourigenesis. GEP-NENs can over-express vascular endothelial growth factor (VEGF), basic-fibroblastic growth factor, transforming growth factor (TGF-α and -β), platelet derived growth factor (PDGF), insulin-like growth factor-1 (IGF-1) and their receptors PDGF receptor, IGF-1 receptor, epidermal growth factor receptor, VEGF receptor, and c-kit (stem cell factor receptor) that can be considered as potential targets. The availability of new targeted agents, such as everolimus and sunitinib that are effective in advanced and metastatic pancreatic neuroendocrine tumours, has provided new treatment opportunities. Many trials combing new drugs are ongoing.
Keywords: Neuroendocrine neoplasms of the gastro-entero-pancreatic system, Chemotherapy, Targeted agents, Somatostatin analogues, Everolimus, Sunitinib
Core tip: In this review, recent evidences in the biology and pathology of neuroendocrine neoplasms of the gastro-entero-pancreatic system were analysed, focusing on new biological perspectives of medical treatment. The evidence-based data of new-targeted drugs and the new molecular knowledge are summarized looking at the basis for future studies.
INTRODUCTION
Neuroendocrine neoplasms of the gastro-entero-pancreatic system (GEP-NENs) include a heterogeneous group of disease emerging from neuroendocrine cells of gastro-intestinal tract and pancreatic islets[1]. Nevertheless, despite their morphologic, clinical and prognostic heterogeneity, GEP-NENs are often considered as a single entity[2].
Although still considered a rare disease, SEER data showed an increasing incidence in the last three decades up to 3.65/100000 per years[3]. This may be due to a remarkable improvement of diagnostic technique as well as a real change in population demography[4]. GEP-NENs are more frequently detected in adult population[5] and in about 50% of cases nodal (25%) or distant (25%) metastases are already existing from the beginning[3,6]. On the basis of their morphologic features and proliferation index, NENs are currently stratified in two groups, according to WHO 2010 classification criteria[7]: Neuroendocrine carcinomas, G3 tumours with ki67 proliferation index > 20%, and neuroendocrine tumours (NETs), including G1 (ki67 < 3%) and G2 (ki67 between 3% and 20%) neoplasms. Neuroendocrine carcinomas represent a separate cluster in the family of NENs, with specific biological features and a more aggressive behavior, so chemotherapy is currently considered the standard of care in this specific set[8,9]. Conversely well and moderately-differentiated NENs do not represent a single entity and their pathogenesis has become clearer in recent years. In fact many driver mutations pathway genes have been identified as crucial factors in their tumourigenesis. Therefore altered pathways represent as a profitable therapeutic choice in neoplastic disease and also in NENs[10-13].
Despite extensive and remarkable medical exertions, therapeutic choices are still unsatisfactory, mainly due to the lack of a broad knowledge of biological mechanisms and predictive factors. This review aims to summarize the present knowledge about chemotherapy and the pathways involved in sporadic well and moderately differentiated GEP-NETs, highlighting available evidences and new biological perspectives on biological and targeted therapies.
CHEMOTHERAPY
Although most of the studies were conducted on a heterogeneous population and the relationship between response rate (RR) and proliferation index value is often not clearly defined, GEP-NENs, therapy should include cytotoxic agents, especially in symptomatic subjects, progressive disease, moderated differentiation and more aggressive features. Chemotherapy should also be evaluated when the aim is to obtain a response in case of bulky lesions. However the best sequence for chemotherapy still remains uncertain[14-18].
The most common used chemotherapy schemes include alkylating agents [streptozotocin (STZ), dacarbazine, temozolomide], antimetabolites [5-fluorouracil (5-FU), capecitabine] and platinum derivatives.
Temozolomide combined with 5-FU[19] or capecitabine[20] can represent the regimen of choice in G1 and G2 advanced P-NENs. Retrospective data showed a RR of 70% and progression-free survival (PFS) of 18 mo for temozolomide and capecitabine combination[20].
Furthermore the association of STZ and 5-FU is frequently evaluated as a first-line therapy for advanced P-NENs with RRs between 6% to 40%, with the benefit in PFS ranging between 5 and 20 mo and with a median overall survival of 16-24 mo[19].
Then, oxaliplatin in combination with capecitabine could also be considered for different setting of G1-G2 GEP-NETs[15]. None of small retrospective studies or case reports conducted with other chemotherapy regimens have demonstrated sufficient efficacy in GEP-NETs.
SOMATOSTATIN
Many studies have shown the importance of somatostatin in the regulation of NENs’ physiological functions. Currently, a cluster of five distinct somatostatin receptors (SSTRs) has been characterized in humans (SSTR1-SSTR5)[21,22].
The presence of SSTRs has been demonstrated in over 80% of well-differentiated GEP-NENs, with a clear predominance of SSTR2 both in GI-NENs (90%) and P-NETs (80%)[23,24].
Among the different SSTR subtypes, SSTR2 is usually the most prevalent in NENs, after that SSTR1 and SSTR5, whilst SSTR3 is less commonly expressed and SSTR4 almost absent[25-27].
In general, tumour dedifferentiation is usually associated with a reduction of receptor density and changes in receptor subtype profile; thus, the presence of SSTRs might be also useful as a tumour specific predictor of prognosis.
Furthermore, the presence of SSTR5 seems to correlate with a major risk of angioinvasion and distant metastasis[28]; instead, the loss expression of SSTR2 could be highly associated with the disregulation of tumour proliferation, consequently promoting tumour growth[29]. The lack of SSTR2 induces the generation of new membrane dimers, with development of different receptors, characterized by new function[29-36]. It remains unclear if only numeric reduction of SSTRs or also their down-regulation are linked with tumour dedifferentiation[37]. In pancreatic gastrinomas, glucagonomas and VIPomas, SSTRs are high expressed (80%-100% of patients). However, SSTRs seem to be expressed in 50%-70% of insulinomas, especially SSTR5 mRNA expression was demonstrated to be positively correlated with histopathological features of tumour aggressiveness in primary insulinomas[38].
Therefore, in P-NENs subtypes, which express less SSTR, short synthetic analogues of somatostatin (SSAs) show a reduced activity in symptoms’ control with a worsen hypoglycaemia[39,40]. This high and heterogeneous expression does not show any relevant correlation between the subtype(s) expressed and the primary tumour origin, or a specific hormone secretion[41-43].
The intracellular pathways activated by SSTRs appear different in several types of tumour cells and depend on the specific SSTR distribution pattern, signalling elements, as well as to receptor desensitization, internalization, and cross talk[44,45].
The activation of G-proteins regulates the different critical enzymatic proteins such as adenylyl cyclase and protein kinase A, phospho-tyrosine phosphatases (PTPs) and mitogen activated kinases (MAPKs)[22,46,47].
In particular SSTR1 induces MAPK pathway activations, SSTR2 improves SHP1 and epidermal growth factor receptor (EGFR) work, up-regulate p21 and Rb reducing MAPK switching on and blocking cellular proliferation. SSTR3 activates p53 and Bax inducing apoptosis, besides it blocks vascular EGFR (VEGFR). SSTR5 induce the activations of PTPs. Globally, these mechanisms leads to an inhibition of cellular proliferation and hormones secretion. Conversely, SSTR4 promotes cell mitosis up-regulating MAPK/ERK1/2 pathway[21,48].
Since the 80s’, several SSAs including octreotide, lanreotide, vapreotide, seglitide and pasireotide, were studied. In contrast to the endogenous somatostatin, these peptides have a more durable half-life (1.5-2 h vs 1-2 min) and activity, as they have a greater resistance to peptidase[49].
Furthermore, compared to native somatostatin, they have diverse affinity for the aforementioned receptor subtypes[25,37,50]. In particular the natural ligands of SSTR1-5 can bind all SSTRs with high affinity. Conversely different SSAs, in the same cell type, may elicit differential effects, due to the activation of different subsets of intracellular mediators[45,51,52].
The analogues octreotide, lanreotide, vapreotide and seglitide exhibit elevated affinity for SSTR2 and lower for SSTR3 and SSTR5. Multi-SSTR-targeted analogue SOM230 (pasireotide) shows higher binding capacity towards SSTR1 and activates also SSTR 2, 3 and 5[50,53].
The various SSTR binding show a different affinity with their own ligands, which is responsible for the distinct biological and clinical activity[37]. Imam et al[54] and Eriksson et al[55] demonstrated a pro-apoptotic role of SSAs. In fact they analysed tumor samples of GEP-NENs patients, who received high doses of SSAs[54,55], finding increased apoptosis processes. The antiproliferative effect of SSAs is mediated by direct and indirect mechanisms. The inhibition of SSTRs, if expressed on tumour cells’ membrane, operates directly on cell proliferation, stimulating antimitotic and apoptotic activities. SSAs induce cell growth inhibition also with indirect activities, such as angiogenesis inhibition, modulation of immune system and growth factors’ block.
The indirect antiproliferative efficacy of SSAs does not require SSTR tumour expression and is shown by an antiangiogenic or immunomodulation mechanism, mediated by stimulation of the production of natural-killer cells[56-58]. The antiproliferative activity of SSAs has been shown through various experimental models[59-64]. The indication of using SSAs as fundamental therapy in NETs derives mainly from two studies: PROMID and CLARINET trials[65,66]. The PROMID study showed a significant benefit with octreotide LAR (long-acting release) therapy in 85 subjects affected by advanced midgut NENs.
This study demonstrated an advantage in time to progression (TTP). In fact in patients treated with octreotide LAR a mTTP of 14.3 mo was observed, whilst patients in the control arm, receiving placebo, reported a mTTP of 6 mo. Sixty-four percent of subjects in the experimental arm showed stable disease (SD), which was observed only in 37.2% of subjects assuming placebo. Furthermore, patients treated with octreotide LAR experienced a 67% risk reduction of tumour progression compared with patients receiving placebo. The benefit of octreotide LAR was independent either of chromogranin level or hormone secretion.
The study did not show significant differences in OS, presumably due to the few deaths’ percentage in both treatment arms. Furthermore the failure of the demonstration of an impact of octreotide in survival could be also done to the high rate of cross-over[67].
Based on PROMID results, octreotide LAR has been approved as treatment of recurrent and advanced neuroendocrine tumors’ patients, irrespective of the site of primary tumour, functional status and symptoms’ presence. Lanreotide is another SSA with a similar in vitro hormone release inhibitory profile to octreotide[68].
Recently, the CLARINET trial focused on 204 subjects suffering of nonfunctioning GEP-NENs who were randomized to receive either depot lanreotide, 120 mg every 4 wk for 96 wk, or placebo. The study demonstrated an improvement in PFS for patients treated with lanreotide (mPFS not reached in lanreotide arm; mPFS of 18 mo in placebo arm). This benefit was confirmed both in patients with P-NENs and midgut NENs.
Pasireotide, a new SSA, is characterized by an elevated binding affinity to four of the five SSTR sub-types[69]. Hence, due to its broad binding profile, pasireotide may represent an effective therapeutic opportunity in tumours refractory to octreotide or lanreotide[70]. However, its role in GEP-NETs still remains to be defined. In a phase III study pasireotide did not improve the control of flushing or diarrhea in patients affected by refractory carcinoid syndrome[71] (Table 1). The antiproliferative effects are being tested in several clinical studies[72,73]. Telotristat etiprate (LX1606) is an oral serotonin synthesis inhibitor used in patients with diarrhoea related to carcinoid syndrome[74].
Table 1.
Ongoing phaseIII trials in gastro-entero-pancreatic neuroendocrine tumours
| ClinicalTrials.gov Identifier | Investigated drug | Target | Type of enrolled pts |
| NCT00171873 | Octreotide LAR 30 mg | SSTR | Locally inoperable or metastatic well differentiated NETs of the midgut Naïve pts |
| NCT01524783 | Everolimus plus BSC vs PBO plus BSC | mTOR | Unresectable or metastatic G1 or G2 neuroendocrine tumours of GI or lung Treatment-naïve pts and pre-treated pts (all available treatment options are allowed) with PD |
| NCT00842348 | Lanreotide autogel 120 mg | SSTR | Non-functioning GEP-NETs |
| NCT00690430 | Pasireotide LAR 60 mg vs Octreotide LAR 40 mg | SSTR | Metastatic carcinoid tumours Pts with disease-related symptoms inadequately controlled by somatostatin analogues |
| NCT00774930 | Somatuline depot (lanreotide) vs PCB | SSTR | Carcinoid tumours with liver metastasis Treatment-naïve pts and pts pre-treated with and responsive to somatostatine analogues |
| NCT00092287 | Lanreotide autogel vs Sandostatin LAR | SSTR | Carcinoid tumours localized in lung, stomach or midgut Treatment-naïve pts and pts pre-treated with and responsive to somatostatine analogues |
| NCT00263659 | Telotristat etiprate (LX1606) vs PBO | TPH | Well-differentiated metastatic NETs with carcinoid syndrome Treatment-naïve pts |
| NCT01677910 | Telotristat etiprate (LX1606) vs PBO | TPH | Well-differentiated metastatic NETs with carcinoid syndrome Pts with disease-related symptoms inadequately controlled by somatostatin analogues |
GEP-NETs: Gastro-entero-pancreatic neuroendocrine tumours; LAR: Long acting release; SSTR: Somatostatin receptor; mTOR: Mammalian target of rapamycin; BSC: Best supportive care; PBO: Placebo; PD: Programmed death; TPH: Tryptophan hydroxylase; pts: Patients.
A recent randomized prospective single-arm study has been conducted in patients with carcinoid tumour and diarrhoea (≥ 4 bowel movements/day) inadequately controlled by octreotide. Among patients treated with telotristat etiprate, 28% experienced a ≥ 30% reduction in bowel movements frequency for more than 2 wk and 56% had a biochemical response. These results suggest a potential activity of telotristat etiprate in controlling carcinoid syndrome and diarrhoea. Pavel et al[75] made a prospective exploratory dose escalating 12-wk open label multicentre study of telotristat etiprate in metastatic well-differentiated NETs with ≥ 4-bowel movements/day. Whole patients experienced reductions in bowel movements, 74.2% mean reduction in metabolites of serotonin and 75% of patients reported adequate relief of GI symptoms (Table 1).
MAMMALIAN TARGET OF RAPAMYCIN PROTEIN KINASE B, PHOSPHOINOSITIDE 3-KINASE AND PHOSPHATASE AND TENSIN HOMOLOG PATHWAY
A considerable number of intracellular pathways seem to conditionate tumorigenesis and neoplastic spread in NENs, as receptor tyrosine kinases (RTKs) and G-protein coupled receptors (GPCRs) transduction mechanisms. Their action seems to be modulated by Ras/Raf, MAPK, phosphoinositide 3-kinase (PI3K)-protein kinase B (AKT)-mammalian target of rapamycin (mTOR) and JNK increasing cells’ growth and number. The AKT family of serine/threonine kinases is an important mediator of PI3K signaling, promoting the principal cellular functions[76]. Akt isoforms seem to be an eminent target for GEP-NENs therapy[77]. PI3K/AKT/mTOR pathway is especially activated among P-NENs[78] and their somatic mutations are detected among a minority of P-NETs[79]. Although discrete mutations in the aforementioned pathway are rarely found in GEP-NENs, overexpression of mTOR and/or its downstream targets is been individuated in a high frequency of cases and it is correlated with higher proliferative activity and adverse clinical outcomes[80,81]. mTOR is composed by two complexes working together guarantying many cells’ activities[82-91]. The importance of mTOR inhibitors results from the aforementioned considerations[92,93]. RADIANT-1 (phase II study) represents the first trial demonstrating everolimus utility in GEP-NETs[94]. The trial compared everolimus alone vs everolimus plus octreotide in 160 patients. Regarding combined therapy arm the median PFS was 16.7 mo with a quite well tolerance.
In RADIANT-2 (phase III trial) subjects affected by symptomatic well-differentiated NETs received everolimus plus octreotide vs octreotide alone. A lack of significant benefit in PFS was showed in the combination arm. The most common grade 3/4 side effects in the everolimus arm were stomatitis (6.5%), diarrhea (6%), infections (5.1%), and hyperglycemia (5.1%)[95]. RADIANT-3 (phase III trial) contemplated everolimus vs placebo[96]. The study recruited only G1-G2 P-NETs subjects. Everolimus arm was associated with a better PFS although a low ORR. Therefore everolimus was approved in the management of advanced P-NETs.
RADIANT-4 (ongoing phase III trial) investigates role of everolimus in gastrointestinal/pulmonary neuroendocrine tumors. It may lead to a better definition of the role of everolimus in patients with carcinoid tumours. Finally, other targeted therapies are being studied in NETs (Table 1). Furthermore temsirolimus, another mTOR inhibitor, was evaluated in NETs[97]. However, the results were not considered clinically relevant and further studies with this agent in NETs won’t be performed.
Another fundamental target implicated is PTEN (phosphatase and tensin homologue). Loss of PTEN is commonly individualized in a several human cancers[98] and it is related to the presence of metastases and therapy resistance towards mTOR inhibition[99-103]. PTEN is localized in the nucleus. Its activation through internalization leads to a reduction of Act[104-106]. PTEN is frequently mutated in P-NETs and a low expression of PTEN correlates with high grading[107].
PI3K pathway represents a hot point in NETs proliferation and some studies evaluating its inhibition are ongoing. BEZ235 is a PI3K inhibitor studied associated with everolimus (phase II study) (Table 2). Then a phase I study is on-going using BYL179 in combination with everolimus and exemestane in P-NETs.
Table 2.
Ongoing phase II trials in gastro-entero-pancreatic neuroendocrine tumours
| ClinicalTrials.gov Identifier | Investigated drug | Target | Type of enrolled pts |
| NCT01841736 | Pazopanib | VEGFR PDGFR FGFR c-kit | Progressive carcinoid tumours |
| NCT02399215 | Nintedanib | VEGFR FGFR PDGFR | Carcinoid tumour Metastatic carcinoid tumour Neuroendocrine neoplasm |
| NCT01994213 | Famitinib | c-kit PDGFR VEGFR Flt | Gastroenteropancreatic neuroendocrine tumour |
| NCT01121939 | Bevacizumab plus pertuzumab plus sandostatin LAR | VEGF HER2 | Advanced neuroendocrine cancers |
| NCT02259725 | Regorafenib | c-RAF BRAF VEGFR PDGFRa FGFR-1 c-kit RET Flt-3 | Gastrinoma Glucagonoma Insulinoma Metastatic gastrointestinal carcinoid tumour Pancreatic polypeptide tumour Pulmonary carcinoid tumour Recurrent gastrointestinal carcinoid tumour Recurrent Islet cell carcinoma Somatostatinoma |
| NCT01784861 | X-82 plus everolimus | mTOR | Pancreatic neuroendocrine tumours |
| NCT01508104 | BEZ235 plus everolimus | PI3K | Advanced cancers of different types |
| NCT00781911 | Cixutumumab | IGF-1R | Neuroendocrine tumours |
VEGFR: Vascular endothelial growth factor receptor; PDGFR: Platelet derived growth factor receptor; FGFR: Fibroblast growth factor receptor; mTOR: Mammalian target of rapamycin; PI3K: Phosphoinositide 3-kinase; IGF-1R: Insulin-like growth factor-1 receptor; pts: Patients.
INSULIN GROWTH FACTOR-1
Insulin growth factor 1 (IGF-1) represents a fundamental factor in tumour expansion, so its inhibition may reduce tumour proliferation. NETs have demonstrated to secrete a significant quantity of IGF-1, then its receptor (IGF-1R) shows a key role in GEP-NETs tumorigenesis[108,109].
Furthermore, many evidences have related a major IGF-1R expression with the presence of functioning and symptomatic NETs[109-118]. Cixutumumab (CIX), a monoclonal antibody competitively binding IGF-1R and then causing its degradation, is currently being evaluated in an on-going trial in association with octreotide depot (Table 2). The usefulness of CIX has already been demonstrated in combination with many other therapeutic options[119].
VEGF
Angiogenesis displays a crucial role for tumour expansion and distant spread and it’s mediated by VEGF and its receptors (VEGFRs). Four VEGF forms were identified: VEGF-A, VEGF-B, VEGF-C and VEGF-D, with a different affinity to their three own receptors[120-129]. Octreotide showed an inhibition of angiogenesis probably mediated by an interaction with VEGF pathway[130]. The tyrosine kinase inhibitor (TKI) sunitinib[131] has been demonstrated a valid targeted therapy option in NENs.
A phase II trial evaluated the efficacy of sunitinib in GEP-NETs demonstrating a significant antitumour activity in P-NETs, while among patients with carcinoid tumours OR were only 2.4%; the treatment was average well tolerated with especially gastrointestinal toxicities[132].
As a consequence of these results, a phase III trial evaluated sunitinib vs placebo in 171 low- and intermediate-grade advanced P-NETs[133]. In the experimental arm was demonstrated an improvement of PFS although the RRs associated with the drug were only 9.3%. The benefit was independent of previous treatments and concomitant administration of SSAs. Considering the importance of VEGF in pathogenesis of NENs, bevacizumab, an antibody directed against VEGF[134], has been used either alone or in combination with other drugs with favourable results[135].
CYTOTOXIC T-LYMPHOCYTE ANTIGEN-4 AND PROGRAMMED DEATH-1
Recently, immunotherapy was demonstrated to be an important treatment option in various cancers. In fact several new immune-target drugs, directed towards specific immune checkpoints, showed an important antitumoral effect.
The first developed immune agents were directed against mediator of immunity inhibition, as cytotoxic t-lymphocyte antigen-4 (CTLA-4) and programmed death-1 (PD-1). These mediators are both membrane glycoprotein, which are mainly expressed in activated T-lymphocyte.
CTLA-4, known also as CD152, owns an elevated kinship with CD28 and plays a crucial role regulating immunity’s homeostasis, through the switching-off of T-lymphocyte activation. Its expression seems to be mayor stimulated in switched-on effector T-lymphocytes (Teff cells)[136]. CTLA-4 is constitutive and represented in regulatory T lymphocytes (Treg)[137]. As aforementioned said, it joints CD28, thanks to theirs high affinity, to costimulatory proteins (CD80, CD86) represented in antigen-presenting cells (APC).
Several humanized monoclonal antibodies directed vs CTLA-4, were studied, such as ipilimumab and tremelimumab. The programmed cell death protein-1, PD-1, a membrane protein, acts inhibiting a large group of molecules owning to CD28 family of T-lymphocytes regulators. PD-1 is most represented on surface membrane of activated monocytes, T lymphocytes, and B lymphocytes. PD-1 have different ligands, the most known are PD-L1[138] and PD-L2[139].
PD-L1, a transmembrane protein notably presents in macrophages, in T-ymphocytes, B lymphocytes and dendritic cells (DCs), its concentration increases since cellular activating processes. PD-L1 may be presented also in some tissues not involved in immune system. The principal function of PD-1 seems to be reducing autoimmunity and switching off T-lymphocyte activities involved in inflammatory response to infection[140-142].
In conclusion the linkage between PD-1, mainly expressed in activated T-lymphocytes and PD-L1, principally expressed in tissue DCs, induce a switching-off of T-lymphocytes activation and a blockage of their effector activity[143]. Identifying a selected group of NENs’ patients that could benefit from immunotherapies is not still possible because no predictive biomarkers to immune drugs have been found. Further studies are needed to evaluate the exact expression of aforementioned target immune proteins (PD-1, PD-L1/L2) in the various NENs.
EGF AND TRANSFORMING GROWTH FACTOR ALPHA
EGF and transforming growth factor alpha (TGF-α) are polypeptides that bind the EGFRs regulating cellular responses to growth signals through activating signal transduction pathways (RAS-RAF-MAPK). From a biological point of view, EGF is a mitogen factor regulating growth, proliferation and differentiation of numerous cell types; abnormalities in EGF-signalling pathways have been related to tumour growth and progression[144].
The EGFR belongs to the HER receptor family. Gastrointestinal (GI) and pancreatic NETs express and activate EGFRs[145]. Papouchado et al[146] demonstrated a most elevated presence of EGFR (> 91%) in GI-NENs, (especially in rectal NETs), whilst in P-NENs its expression was lower (< 25%).
Srivastava et al[147] showed instead an elevated presence of EGFR and TGF-α, in P-NENs. Sixty-three per cent of neoplasms in fact showed positivity for TGF-α and 65% for EGFR. However the study did not demonstrate an association with measure, functional status, ability to secrete hormones, or biologic behaviour[147].
TGF-α is expressed in approximately 70%-100% of NETs depending on the technique used (immunohistochemistry or northern blot analysis)[148-150] and is commonly over-expressed in larger rectal NETs with a high Ki-67 index[150]. TGF-α binds with high affinity to the EGFR extracellular domain. Cytoplasmic substrates phosphorylation occurs and initiates a signalling cascade (RAS/RAF/MAPK-ERK) that drives pro-proliferative gene expression, cytoskeletal rearrangement, and increased cell proliferation[144].
Gefitinib is a targeted agent that selectively inhibits receptor tyrosine kinases, including EGFR. A phase II trial enrolling subjects affected by advanced NENs, gefitinib exhibited somewhat promising initial results. At 6 mo, 61% of patients affected by carcinoid tumours and 31% affected by P-NEN were progression-free; however, objective responses for each group were low, 5% and 9.6%, respectively[151].
BASIC FIBROBLASTIC GROWTH FACTOR
The basic fibroblastic growth factor (bFGF) is involved in both physiological and pathological processes by interaction with determinated receptors localized in cellular membrane[152,153].
Because overexpression of bFGF and/or its receptors is frequently detected in tumours, the development of antagonists to bFGF and its receptors has been studied as a potential strategy for cancer therapy[154-156].
Almost five isoform of transmembrane FGF receptors (FGFR), able to dimerize, are well known. The first four subtypes are characterized by a tyrosine kinase activity[157]. Chaudhry et al[158] searched for mRNA expression of 6 different transmembrane receptors (FGFR, EGFR, IGF-1R, TGF-betaR1 and betaR2), and the presence of SSTRs in determinate subtypes of GEP-NENs tissues (gastrinoma, insulinoma, tumours with carcinoid syndrome, not-functioning neoplasms) using reverse transcriptase-polymerase chain reaction. Among the four tumour subtypes, expression frequencies of the receptors aforementioned varied significantly[158]. Taken together, these studies have accounted for high growth factor abundance in GEP-NENs. Considering these results GEP-NENs seems to have an elevated growth factors concentration.
C-KIT/ PLATELET DERIVED GROWTH FACTOR
The c-kit receptor, also referred to CD117 or platelet derived growth factor receptor (PDGFR) is a type I transmembrane glycoprotein. It is usually included in the family of tyrosine kinase receptor (RTK)[159].
In tumor cells, PDGF promotes proliferation and neoplastic spread[160-163]. Various subtypes of c-kit receptor have been already identified[164] but their ligand still remains stem cell factor (SCF), a hematopoietic cytokine involved in cell survival, proliferation and differentiation[165]. Few pre-clinical studies performed of GEP-NETs have shown a variable expression of c-kit, with ranges from 0% to 38%, and PDGFRα in carcinoids[166], with a particularly high expression in gastrinomas (up to 100% of c-kit expression)[167].
MULTI-TARGETED AGENTS
Famitinib is an oral tyrosine-inhibitor agent targeting at c-kit, PDGFR, VEGFR2, VEGFR3, Flt1 and Flt3. Its efficacy in GEP-NETs is currently being evaluated (Table 2).
Regorafenib is a novel multi-kinase inhibitor (c-RAF; BRAF, VEGFR-1, 2, 3; PDGFRα, FGFR-1; c-kit; RET; Flt-3) belonging to the group of biaryl urea chemicals[168-170]. Pazopanib is an oral inhibitor of several specific cellular pathways involved in neoplastic growth and dissemination[171]. Its efficacy in NENs was demonstrated in a phase II clinical trial combining pazopanib and SSA achieving a 17% RR in G1 P-NETs[172]. Data related to ongoing trials with pazopanib and with regorafenib in NETs are summarized in Table 2.
CONCLUSION
In GEP-NETs tumourigenesis and progression are often involved SSTRs, mTOR/Akt/PI3K and PTEN, IGF-1, VEGF, EGF, TGF, FGF and c-kit/PDGF and its corresponding receptors[145,148,149,173-177] (Figure 1). The recent availability of novel drugs has provided new treatment opportunities and holds promise given the expression in GEP-NENs of this variety of targets[33,178,179].
Figure 1.
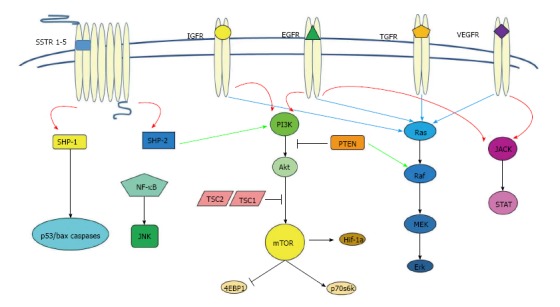
Illustration of principal pathways involved in cellular differentiation, proliferation, survival and apoptosis: Somatostatin receptors, mammalian target of rapamycin protein kinase B, phosphoinositide 3-kinase and phosphatase and tensin homolog, insulin-like growth factor 1 receptor, vascular endothelial growth factor receptor, epidermal growth factor receptor, transforming growth factor receptor, fibroblast growth factors. SSTRs: Somatostatin receptors; mTOR: Mammalian target of rapamycin; Akt: Protein kinase B; PI3K: Phosphoinositide 3-kinase; PTEN: Phosphatase and tensin homolog; IGFR: Insulin-like growth factor receptor; VEGFR: Vascular endothelial growth factor receptor; EGFR: Epidermal growth factor receptor; TGFR: Transforming growth factor receptor.
Footnotes
Conflict-of-interest statement: All authors declare that they have no competing interests.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: May 25, 2015
First decision: September 14, 2015
Article in press: February 16, 2016
P- Reviewer: Kleeff J S- Editor: Ji FF L- Editor: A E- Editor: Lu YJ
References
- 1.Cives M, Strosberg J. An update on gastroenteropancreatic neuroendocrine tumors. Oncology (Williston Park) 2014;28:749–756, 758. [PubMed] [Google Scholar]
- 2.Yang Z, Tang LH, Klimstra DS. Gastroenteropancreatic neuroendocrine neoplasms: historical context and current issues. Semin Diagn Pathol. 2013;30:186–196. doi: 10.1053/j.semdp.2013.06.005. [DOI] [PubMed] [Google Scholar]
- 3.Yao JC, Hassan M, Phan A, Dagohoy C, Leary C, Mares JE, Abdalla EK, Fleming JB, Vauthey JN, Rashid A, et al. One hundred years after “carcinoid”: epidemiology of and prognostic factors for neuroendocrine tumors in 35,825 cases in the United States. J Clin Oncol. 2008;26:3063–3072. doi: 10.1200/JCO.2007.15.4377. [DOI] [PubMed] [Google Scholar]
- 4.van der Zwan JM, Trama A, Otter R, Larrañaga N, Tavilla A, Marcos-Gragera R, Dei Tos AP, Baudin E, Poston G, Links T. Rare neuroendocrine tumours: results of the surveillance of rare cancers in Europe project. Eur J Cancer. 2013;49:2565–2578. doi: 10.1016/j.ejca.2013.02.029. [DOI] [PubMed] [Google Scholar]
- 5.Lepage C, Bouvier AM, Faivre J. Endocrine tumours: epidemiology of malignant digestive neuroendocrine tumours. Eur J Endocrinol. 2013;168:R77–R83. doi: 10.1530/EJE-12-0418. [DOI] [PubMed] [Google Scholar]
- 6.Lawrence B, Gustafsson BI, Chan A, Svejda B, Kidd M, Modlin IM. The epidemiology of gastroenteropancreatic neuroendocrine tumors. Endocrinol Metab Clin North Am. 2011;40:1–18, vii. doi: 10.1016/j.ecl.2010.12.005. [DOI] [PubMed] [Google Scholar]
- 7.Rindi G, Arnold R, Bosman FT, Capella C, Klimstra DS, Kloppel G, Komminoth P, Solcia E. Nomenclature and classification of neuroendocrine neoplasms of the digestive system. In WHO Classification of Tumours of the Digestive System. In: Bosman FT, Carneiro F, Hruban RH, Theise N, editors. Lyon: IARC Press; 2010. pp. 13–14. [Google Scholar]
- 8.Sorbye H, Strosberg J, Baudin E, Klimstra DS, Yao JC. Gastroenteropancreatic high-grade neuroendocrine carcinoma. Cancer. 2014;120:2814–2823. doi: 10.1002/cncr.28721. [DOI] [PubMed] [Google Scholar]
- 9.La Rosa S, Sessa F. High-grade poorly differentiated neuroendocrine carcinomas of the gastroenteropancreatic system: from morphology to proliferation and back. Endocr Pathol. 2014;25:193–198. doi: 10.1007/s12022-014-9316-9. [DOI] [PubMed] [Google Scholar]
- 10.Capdevila J, Salazar R, Halperín I, Abad A, Yao JC. Innovations therapy: mammalian target of rapamycin (mTOR) inhibitors for the treatment of neuroendocrine tumors. Cancer Metastasis Rev. 2011;30 Suppl 1:27–34. doi: 10.1007/s10555-011-9290-3. [DOI] [PubMed] [Google Scholar]
- 11.Capdevila J, Tabernero J. A shining light in the darkness for the treatment of pancreatic neuroendocrine tumors. Cancer Discov. 2011;1:213–221. doi: 10.1158/2159-8290.CD-11-0151. [DOI] [PubMed] [Google Scholar]
- 12.Hilfenhaus G, Göhrig A, Pape UF, Neumann T, Jann H, Zdunek D, Hess G, Stassen JM, Wiedenmann B, Detjen K, et al. Placental growth factor supports neuroendocrine tumor growth and predicts disease prognosis in patients. Endocr Relat Cancer. 2013;20:305–319. doi: 10.1530/ERC-12-0223. [DOI] [PubMed] [Google Scholar]
- 13.Oberstein PE, Saif MW. Update on prognostic and predictive biomarkers for pancreatic neuroendocrine tumors. JOP. 2012;13:368–371. doi: 10.6092/1590-8577/965. [DOI] [PubMed] [Google Scholar]
- 14.Vilar E, Salazar R, Pérez-García J, Cortes J, Oberg K, Tabernero J. Chemotherapy and role of the proliferation marker Ki-67 in digestive neuroendocrine tumors. Endocr Relat Cancer. 2007;14:221–232. doi: 10.1677/ERC-06-0074. [DOI] [PubMed] [Google Scholar]
- 15.Bajetta E, Catena L, Procopio G, De Dosso S, Bichisao E, Ferrari L, Martinetti A, Platania M, Verzoni E, Formisano B, et al. Are capecitabine and oxaliplatin (XELOX) suitable treatments for progressing low-grade and high-grade neuroendocrine tumours? Cancer Chemother Pharmacol. 2007;59:637–642. doi: 10.1007/s00280-006-0306-6. [DOI] [PubMed] [Google Scholar]
- 16.Brixi-Benmansour H, Jouve JL, Mitry E, Bonnetain F, Landi B, Hentic O, Bedenne L, Cadiot G. Phase II study of first-line FOLFIRI for progressive metastatic well-differentiated pancreatic endocrine carcinoma. Dig Liver Dis. 2011;43:912–916. doi: 10.1016/j.dld.2011.07.001. [DOI] [PubMed] [Google Scholar]
- 17.Engstrom PF, Lavin PT, Moertel CG, Folsch E, Douglass HO. Streptozocin plus fluorouracil versus doxorubicin therapy for metastatic carcinoid tumor. J Clin Oncol. 1984;2:1255–1259. doi: 10.1200/JCO.1984.2.11.1255. [DOI] [PubMed] [Google Scholar]
- 18.Vogl TJ, Naguib NN, Zangos S, Eichler K, Hedayati A, Nour-Eldin NE. Liver metastases of neuroendocrine carcinomas: interventional treatment via transarterial embolization, chemoembolization and thermal ablation. Eur J Radiol. 2009;72:517–528. doi: 10.1016/j.ejrad.2008.08.008. [DOI] [PubMed] [Google Scholar]
- 19.Moertel CG, Hanley JA, Johnson LA. Streptozocin alone compared with streptozocin plus fluorouracil in the treatment of advanced islet-cell carcinoma. N Engl J Med. 1980;303:1189–1194. doi: 10.1056/NEJM198011203032101. [DOI] [PubMed] [Google Scholar]
- 20.Strosberg JR, Fine RL, Choi J, Nasir A, Coppola D, Chen DT, Helm J, Kvols L. First-line chemotherapy with capecitabine and temozolomide in patients with metastatic pancreatic endocrine carcinomas. Cancer. 2011;117:268–275. doi: 10.1002/cncr.25425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Patel YC. Somatostatin and its receptor family. Front Neuroendocrinol. 1999;20:157–198. doi: 10.1006/frne.1999.0183. [DOI] [PubMed] [Google Scholar]
- 22.Møller LN, Stidsen CE, Hartmann B, Holst JJ. Somatostatin receptors. Biochim Biophys Acta. 2003;1616:1–84. doi: 10.1016/s0005-2736(03)00235-9. [DOI] [PubMed] [Google Scholar]
- 23.Ferone D, Arvigo M, Semino C, Jaquet P, Saveanu A, Taylor JE, Moreau JP, Culler MD, Albertelli M, Minuto F, et al. Somatostatin and dopamine receptor expression in lung carcinoma cells and effects of chimeric somatostatin-dopamine molecules on cell proliferation. Am J Physiol Endocrinol Metab. 2005;289:E1044–E1050. doi: 10.1152/ajpendo.00209.2005. [DOI] [PubMed] [Google Scholar]
- 24.Arnold R, Trautmann ME, Creutzfeldt W, Benning R, Benning M, Neuhaus C, Jürgensen R, Stein K, Schäfer H, Bruns C, et al. Somatostatin analogue octreotide and inhibition of tumour growth in metastatic endocrine gastroenteropancreatic tumours. Gut. 1996;38:430–438. doi: 10.1136/gut.38.3.430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Reubi JC. Peptide receptors as molecular targets for cancer diagnosis and therapy. Endocr Rev. 2003;24:389–427. doi: 10.1210/er.2002-0007. [DOI] [PubMed] [Google Scholar]
- 26.Reubi JC, Waser B. Concomitant expression of several peptide receptors in neuroendocrine tumours: molecular basis for in vivo multireceptor tumour targeting. Eur J Nucl Med Mol Imaging. 2003;30:781–793. doi: 10.1007/s00259-003-1184-3. [DOI] [PubMed] [Google Scholar]
- 27.Reubi JC. Somatostatin and other Peptide receptors as tools for tumor diagnosis and treatment. Neuroendocrinology. 2004;80 Suppl 1:51–56. doi: 10.1159/000080742. [DOI] [PubMed] [Google Scholar]
- 28.Schmid HA, Lambertini C, van Vugt HH, Barzaghi-Rinaudo P, Schäfer J, Hillenbrand R, Sailer AW, Kaufmann M, Nuciforo P. Monoclonal antibodies against the human somatostatin receptor subtypes 1-5: development and immunohistochemical application in neuroendocrine tumors. Neuroendocrinology. 2012;95:232–247. doi: 10.1159/000330616. [DOI] [PubMed] [Google Scholar]
- 29.Reubi JC, Laissue J, Krenning E, Lamberts SW. Somatostatin receptors in human cancer: incidence, characteristics, functional correlates and clinical implications. J Steroid Biochem Mol Biol. 1992;43:27–35. doi: 10.1016/0960-0760(92)90184-k. [DOI] [PubMed] [Google Scholar]
- 30.Rocheville M, Lange DC, Kumar U, Sasi R, Patel RC, Patel YC. Subtypes of the somatostatin receptor assemble as functional homo- and heterodimers. J Biol Chem. 2000;275:7862–7869. doi: 10.1074/jbc.275.11.7862. [DOI] [PubMed] [Google Scholar]
- 31.Corleto VD, Falconi M, Panzuto F, Milione M, De Luca O, Perri P, Cannizzaro R, Bordi C, Pederzoli P, Scarpa A, et al. Somatostatin receptor subtypes 2 and 5 are associated with better survival in well-differentiated endocrine carcinomas. Neuroendocrinology. 2009;89:223–230. doi: 10.1159/000167796. [DOI] [PubMed] [Google Scholar]
- 32.Kim HS, Lee HS, Kim WH. Clinical significance of protein expression of cyclooxygenase-2 and somatostatin receptors in gastroenteropancreatic neuroendocrine tumors. Cancer Res Treat. 2011;43:181–188. doi: 10.4143/crt.2011.43.3.181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Srirajaskanthan R, Watkins J, Marelli L, Khan K, Caplin ME. Expression of somatostatin and dopamine 2 receptors in neuroendocrine tumours and the potential role for new biotherapies. Neuroendocrinology. 2009;89:308–314. doi: 10.1159/000179899. [DOI] [PubMed] [Google Scholar]
- 34.Zamora V, Cabanne A, Salanova R, Bestani C, Domenichini E, Marmissolle F, Giacomi N, O’Connor J, Méndez G, Roca E. Immunohistochemical expression of somatostatin receptors in digestive endocrine tumours. Dig Liver Dis. 2010;42:220–225. doi: 10.1016/j.dld.2009.07.018. [DOI] [PubMed] [Google Scholar]
- 35.Reubi JC, Kvols LK, Waser B, Nagorney DM, Heitz PU, Charboneau JW, Reading CC, Moertel C. Detection of somatostatin receptors in surgical and percutaneous needle biopsy samples of carcinoids and islet cell carcinomas. Cancer Res. 1990;50:5969–5977. [PubMed] [Google Scholar]
- 36.Oconnor JM, Belli S, Pesce V, Mendez GA, Bestani C, Marmissolle F, Giacomi N, Belli S, Dominichini E, Chacon M, et al. Somatostatin receptor (sstr) expression and proliferative index (ki 67) in 100 patients (pts) with gastroenteropancreatic neuroendocrine tumours (gep-nets). Clinical-pathological correlation. J Clin Oncol. 2012;30:Abstr e14598. [Google Scholar]
- 37.Modlin IM, Pavel M, Kidd M, Gustafsson BI. Review article: somatostatin analogues in the treatment of gastroenteropancreatic neuroendocrine (carcinoid) tumours. Aliment Pharmacol Ther. 2010;31:169–188. doi: 10.1111/j.1365-2036.2009.04174.x. [DOI] [PubMed] [Google Scholar]
- 38.de Sá SV, Corrêa-Giannella ML, Machado MC, de Souza JJ, Pereira MA, Patzina RA, Siqueira SA, Machado MC, Giannella-Neto D. Somatostatin receptor subtype 5 (SSTR5) mRNA expression is related to histopathological features of cell proliferation in insulinomas. Endocr Relat Cancer. 2006;13:69–78. doi: 10.1677/erc.1.00962. [DOI] [PubMed] [Google Scholar]
- 39.Portela-Gomes GM, Stridsberg M, Grimelius L, Rorstad O, Janson ET. Differential expression of the five somatostatin receptor subtypes in human benign and malignant insulinomas - predominance of receptor subtype 4. Endocr Pathol. 2007;18:79–85. doi: 10.1007/s12022-007-0014-8. [DOI] [PubMed] [Google Scholar]
- 40.Janson ET, Oberg K. Neuroendocrine tumors--somatostatin receptor expression and somatostatin analog treatment. Cancer Chemother Biol Response Modif. 2003;21:535–546. [PubMed] [Google Scholar]
- 41.Reubi JC, Kappeler A, Waser B, Laissue J, Hipkin RW, Schonbrunn A. Immunohistochemical localization of somatostatin receptors sst2A in human tumors. Am J Pathol. 1998;153:233–245. doi: 10.1016/S0002-9440(10)65564-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Papotti M, Bongiovanni M, Volante M, Allìa E, Landolfi S, Helboe L, Schindler M, Cole SL, Bussolati G. Expression of somatostatin receptor types 1-5 in 81 cases of gastrointestinal and pancreatic endocrine tumors. A correlative immunohistochemical and reverse-transcriptase polymerase chain reaction analysis. Virchows Arch. 2002;440:461–475. doi: 10.1007/s00428-002-0609-x. [DOI] [PubMed] [Google Scholar]
- 43.Volante M, Rosas R, Allìa E, Granata R, Baragli A, Muccioli G, Papotti M. Somatostatin, cortistatin and their receptors in tumours. Mol Cell Endocrinol. 2008;286:219–229. doi: 10.1016/j.mce.2007.12.002. [DOI] [PubMed] [Google Scholar]
- 44.Lahlou H, Guillermet J, Hortala M, Vernejoul F, Pyronnet S, Bousquet C, Susini C. Molecular signaling of somatostatin receptors. Ann N Y Acad Sci. 2004;1014:121–131. doi: 10.1196/annals.1294.012. [DOI] [PubMed] [Google Scholar]
- 45.Schonbrunn A. Selective agonism in somatostatin receptor signaling and regulation. Mol Cell Endocrinol. 2008;286:35–39. doi: 10.1016/j.mce.2007.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Weckbecker G, Lewis I, Albert R, Schmid HA, Hoyer D, Bruns C. Opportunities in somatostatin research: biological, chemical and therapeutic aspects. Nat Rev Drug Discov. 2003;2:999–1017. doi: 10.1038/nrd1255. [DOI] [PubMed] [Google Scholar]
- 47.Florio T. Somatostatin/somatostatin receptor signalling: phosphotyrosine phosphatases. Mol Cell Endocrinol. 2008;286:40–48. doi: 10.1016/j.mce.2007.08.012. [DOI] [PubMed] [Google Scholar]
- 48.Reisine T, Bell GI. Molecular biology of somatostatin receptors. Endocr Rev. 1995;16:427–442. doi: 10.1210/edrv-16-4-427. [DOI] [PubMed] [Google Scholar]
- 49.Oberg K. Future aspects of somatostatin-receptor-mediated therapy. Neuroendocrinology. 2004;80 Suppl 1:57–61. doi: 10.1159/000080743. [DOI] [PubMed] [Google Scholar]
- 50.Bruns C, Lewis I, Briner U, Meno-Tetang G, Weckbecker G. SOM230: a novel somatostatin peptidomimetic with broad somatotropin release inhibiting factor (SRIF) receptor binding and a unique antisecretory profile. Eur J Endocrinol. 2002;146:707–716. doi: 10.1530/eje.0.1460707. [DOI] [PubMed] [Google Scholar]
- 51.Ben-Shlomo A, Zhou C, Pichurin O, Chesnokova V, Liu NA, Culler MD, Melmed S. Constitutive somatostatin receptor activity determines tonic pituitary cell response. Mol Endocrinol. 2009;23:337–348. doi: 10.1210/me.2008-0361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Cescato R, Loesch KA, Waser B, Mäcke HR, Rivier JE, Reubi JC, Schonbrunn A. Agonist-biased signaling at the sst2A receptor: the multi-somatostatin analogs KE108 and SOM230 activate and antagonize distinct signaling pathways. Mol Endocrinol. 2010;24:240–249. doi: 10.1210/me.2009-0321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Reubi JC, Eisenwiener KP, Rink H, Waser B, Mäcke HR. A new peptidic somatostatin agonist with high affinity to all five somatostatin receptors. Eur J Pharmacol. 2002;456:45–49. doi: 10.1016/s0014-2999(02)02651-1. [DOI] [PubMed] [Google Scholar]
- 54.Imam H, Eriksson B, Lukinius A, Janson ET, Lindgren PG, Wilander E, Oberg K. Induction of apoptosis in neuroendocrine tumors of the digestive system during treatment with somatostatin analogs. Acta Oncol. 1997;36:607–614. doi: 10.3109/02841869709001323. [DOI] [PubMed] [Google Scholar]
- 55.Eriksson B, Renstrup J, Imam H, Oberg K. High-dose treatment with lanreotide of patients with advanced neuroendocrine gastrointestinal tumors: clinical and biological effects. Ann Oncol. 1997;8:1041–1044. doi: 10.1023/a:1008205415035. [DOI] [PubMed] [Google Scholar]
- 56.Lamberts SW, Krenning EP, Reubi JC. The role of somatostatin and its analogs in the diagnosis and treatment of tumors. Endocr Rev. 1991;12:450–482. doi: 10.1210/edrv-12-4-450. [DOI] [PubMed] [Google Scholar]
- 57.Bousquet C, Puente E, Buscail L, Vaysse N, Susini C. Antiproliferative effect of somatostatin and analogs. Chemotherapy. 2001;47 Suppl 2:30–39. doi: 10.1159/000049159. [DOI] [PubMed] [Google Scholar]
- 58.Butturini G, Bettini R, Missiaglia E, Mantovani W, Dalai I, Capelli P, Ferdeghini M, Pederzoli P, Scarpa A, Falconi M. Predictive factors of efficacy of the somatostatin analogue octreotide as first line therapy for advanced pancreatic endocrine carcinoma. Endocr Relat Cancer. 2006;13:1213–1221. doi: 10.1677/erc.1.01200. [DOI] [PubMed] [Google Scholar]
- 59.Schally AV. Oncological applications of somatostatin analogues. Cancer Res. 1988;48:6977–6985. [PubMed] [Google Scholar]
- 60.Weckbecker G, Raulf F, Stolz B, Bruns C. Somatostatin analogs for diagnosis and treatment of cancer. Pharmacol Ther. 1993;60:245–264. doi: 10.1016/0163-7258(93)90009-3. [DOI] [PubMed] [Google Scholar]
- 61.Pollak MN, Schally AV. Mechanisms of antineoplastic action of somatostatin analogs. Proc Soc Exp Biol Med. 1998;217:143–152. doi: 10.3181/00379727-217-44216. [DOI] [PubMed] [Google Scholar]
- 62.Froidevaux S, Eberle AN. Somatostatin analogs and radiopeptides in cancer therapy. Biopolymers. 2002;66:161–183. doi: 10.1002/bip.10256. [DOI] [PubMed] [Google Scholar]
- 63.Schally AV, Nagy A. Chemotherapy targeted to cancers through tumoral hormone receptors. Trends Endocrinol Metab. 2004;15:300–310. doi: 10.1016/j.tem.2004.07.002. [DOI] [PubMed] [Google Scholar]
- 64.Pyronnet S, Bousquet C, Najib S, Azar R, Laklai H, Susini C. Antitumor effects of somatostatin. Mol Cell Endocrinol. 2008;286:230–237. doi: 10.1016/j.mce.2008.02.002. [DOI] [PubMed] [Google Scholar]
- 65.Rinke A, Müller HH, Schade-Brittinger C, Klose KJ, Barth P, Wied M, Mayer C, Aminossadati B, Pape UF, Bläker M, et al. Placebo-controlled, double-blind, prospective, randomized study on the effect of octreotide LAR in the control of tumor growth in patients with metastatic neuroendocrine midgut tumors: a report from the PROMID Study Group. J Clin Oncol. 2009;27:4656–4663. doi: 10.1200/JCO.2009.22.8510. [DOI] [PubMed] [Google Scholar]
- 66.Caplin ME, Pavel M, Ćwikła JB, Phan AT, Raderer M, Sedláčková E, Cadiot G, Wolin EM, Capdevila J, Wall L, et al. Lanreotide in metastatic enteropancreatic neuroendocrine tumors. N Engl J Med. 2014;371:224–233. doi: 10.1056/NEJMoa1316158. [DOI] [PubMed] [Google Scholar]
- 67.Arnold R, Wittenberg M, Rinke A, Schade-Brittinger C, Aminossadati B, Ronicke Gress TM, Mueller HH; PROMID Study Group. Placebo controlled, double blind, prospective, randomized study on the effect of octreotide LAR in the control of tumour growth in patients with metastatic neuroendocrine midgut tumours (PROMID): results on long-term survival. J Clin Oncol. 2013;31:Abstr 4030. doi: 10.1159/000443612. [DOI] [PubMed] [Google Scholar]
- 68.Hofland LJ, van Koetsveld PM, Waaijers M, Zuyderwijk J, Lamberts SW. Relative potencies of the somatostatin analogs octreotide, BIM-23014, and RC-160 on the inhibition of hormone release by cultured human endocrine tumor cells and normal rat anterior pituitary cells. Endocrinology. 1994;134:301–306. doi: 10.1210/endo.134.1.7903931. [DOI] [PubMed] [Google Scholar]
- 69.Weckbecker G, Briner U, Lewis I, Bruns C. SOM230: a new somatostatin peptidomimetic with potent inhibitory effects on the growth hormone/insulin-like growth factor-I axis in rats, primates, and dogs. Endocrinology. 2002;143:4123–4130. doi: 10.1210/en.2002-220219. [DOI] [PubMed] [Google Scholar]
- 70.Schmid HA. Pasireotide (SOM230): development, mechanism of action and potential applications. Mol Cell Endocrinol. 2008;286:69–74. doi: 10.1016/j.mce.2007.09.006. [DOI] [PubMed] [Google Scholar]
- 71.Wolin EM, Jarzab B, Eriksson B, Walter T, Toumpanakis C, Morse M, Tomassetti P, Weber M, Fogelman DR, Ramage J, et al. A multicenter, randomized, blinded, phase III study of pasireotide LAR versus octreotide LAR in patients with metastatic neuroendocrine tumours (NET) with disease-related symptoms inadequately controlled by somatostatin analogs. J Clin Oncol. 2013;31(suppl):Abstr 4031. [Google Scholar]
- 72.Lee Moffitt H. Study of pasireotide long acting release (LAR) in patients with metastatic neuroendocrine tumours. In: ClinicalTrials.gov [Internet]. Bethesda (MD): National Library of Medicine (US) Available from: http://www.clinicaltrials.gov/ct2/show/NCT01253161 NLM identifier: NCT01253161.
- 73.Novartis Pharmaceuticals. Efficacy of everolimus alone or in combination with pasireotide LAR in advanced PNET (COOPERATE-1). In: ClinicalTrials.gov [Internet]. Bethesda (MD): National Library of Medicine (US) Available from: http://www.clinicaltrials.gov/ct2/show/NCT01374451.
- 74.Kulke MH, O’Dorisio T, Phan A, Bergsland E, Law L, Banks P, Freiman J, Frazier K, Jackson J, Yao JC, et al. Telotristat etiprate, a novel serotonin synthesis inhibitor, in patients with carcinoid syndrome and diarrhea not adequately controlled by octreotide. Endocr Relat Cancer. 2014;21:705–714. doi: 10.1530/ERC-14-0173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Pavel M, Hörsch D, Caplin M, Ramage J, Seufferlein T, Valle J, Banks P, Lapuerta P, Sands A, Zambrowicz B, et al. Telotristat etiprate for carcinoid syndrome: a single-arm, multicenter trial. J Clin Endocrinol Metab. 2015;100:1511–1519. doi: 10.1210/jc.2014-2247. [DOI] [PubMed] [Google Scholar]
- 76.Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. doi: 10.1016/j.cell.2011.02.013. [DOI] [PubMed] [Google Scholar]
- 77.Chen WS, Xu PZ, Gottlob K, Chen ML, Sokol K, Shiyanova T, Roninson I, Weng W, Suzuki R, Tobe K, et al. Growth retardation and increased apoptosis in mice with homozygous disruption of the Akt1 gene. Genes Dev. 2001;15:2203–2208. doi: 10.1101/gad.913901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Missiaglia E, Dalai I, Barbi S, Beghelli S, Falconi M, della Peruta M, Piemonti L, Capurso G, Di Florio A, delle Fave G, et al. Pancreatic endocrine tumors: expression profiling evidences a role for AKT-mTOR pathway. J Clin Oncol. 2010;28:245–255. doi: 10.1200/JCO.2008.21.5988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Humar R, Kiefer FN, Berns H, Resink TJ, Battegay EJ. Hypoxia enhances vascular cell proliferation and angiogenesis in vitro via rapamycin (mTOR)-dependent signaling. FASEB J. 2002;16:771–780. doi: 10.1096/fj.01-0658com. [DOI] [PubMed] [Google Scholar]
- 80.Alonso-Gordoa T, Capdevila J, Grande E. GEP-NETs update: Biotherapy for neuroendocrine tumours. Eur J Endocrinol. 2015;172:R31–R46. doi: 10.1530/EJE-14-0354. [DOI] [PubMed] [Google Scholar]
- 81.Jiao Y, Shi C, Edil BH, de Wilde RF, Klimstra DS, Maitra A, Schulick RD, Tang LH, Wolfgang CL, Choti MA, et al. DAXX/ATRX, MEN1, and mTOR pathway genes are frequently altered in pancreatic neuroendocrine tumors. Science. 2011;331:1199–1203. doi: 10.1126/science.1200609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Edinger AL, Thompson CB. Akt maintains cell size and survival by increasing mTOR-dependent nutrient uptake. Mol Biol Cell. 2002;13:2276–2288. doi: 10.1091/mbc.01-12-0584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Jaeschke A, Dennis PB, Thomas G. mTOR: a mediator of intracellular homeostasis. Curr Top Microbiol Immunol. 2004;279:283–298. doi: 10.1007/978-3-642-18930-2_17. [DOI] [PubMed] [Google Scholar]
- 84.Shaw RJ, Cantley LC. Ras, PI(3)K and mTOR signalling controls tumour cell growth. Nature. 2006;441:424–430. doi: 10.1038/nature04869. [DOI] [PubMed] [Google Scholar]
- 85.Wang X, Proud CG. The mTOR pathway in the control of protein synthesis. Physiology (Bethesda) 2006;21:362–369. doi: 10.1152/physiol.00024.2006. [DOI] [PubMed] [Google Scholar]
- 86.Herman MA, Kahn BB. Glucose transport and sensing in the maintenance of glucose homeostasis and metabolic harmony. J Clin Invest. 2006;116:1767–1775. doi: 10.1172/JCI29027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Motoshima H, Goldstein BJ, Igata M, Araki E. AMPK and cell proliferation--AMPK as a therapeutic target for atherosclerosis and cancer. J Physiol. 2006;574:63–71. doi: 10.1113/jphysiol.2006.108324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Guertin DA, Sabatini DM. Defining the role of mTOR in cancer. Cancer Cell. 2007;12:9–22. doi: 10.1016/j.ccr.2007.05.008. [DOI] [PubMed] [Google Scholar]
- 89.Sarbassov DD, Ali SM, Sabatini DM. Growing roles for the mTOR pathway. Curr Opin Cell Biol. 2005;17:596–603. doi: 10.1016/j.ceb.2005.09.009. [DOI] [PubMed] [Google Scholar]
- 90.Ellisen LW. Growth control under stress: mTOR regulation through the REDD1-TSC pathway. Cell Cycle. 2005;4:1500–1502. doi: 10.4161/cc.4.11.2139. [DOI] [PubMed] [Google Scholar]
- 91.Kaper F, Dornhoefer N, Giaccia AJ. Mutations in the PI3K/PTEN/TSC2 pathway contribute to mammalian target of rapamycin activity and increased translation under hypoxic conditions. Cancer Res. 2006;66:1561–1569. doi: 10.1158/0008-5472.CAN-05-3375. [DOI] [PubMed] [Google Scholar]
- 92.Croce CM. Oncogenes and cancer. N Engl J Med. 2008;358:502–511. doi: 10.1056/NEJMra072367. [DOI] [PubMed] [Google Scholar]
- 93.Cingarlini S, Bonomi M, Corbo V, Scarpa A, Tortora G. Profiling mTOR pathway in neuroendocrine tumors. Target Oncol. 2012;7:183–188. doi: 10.1007/s11523-012-0226-9. [DOI] [PubMed] [Google Scholar]
- 94.Yao JC, Lombard-Bohas C, Baudin E, Kvols LK, Rougier P, Ruszniewski P, Hoosen S, St Peter J, Haas T, Lebwohl D, et al. Daily oral everolimus activity in patients with metastatic pancreatic neuroendocrine tumors after failure of cytotoxic chemotherapy: a phase II trial. J Clin Oncol. 2010;28:69–76. doi: 10.1200/JCO.2009.24.2669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Pavel ME, Hainsworth JD, Baudin E, Peeters M, Hörsch D, Winkler RE, Klimovsky J, Lebwohl D, Jehl V, Wolin EM, et al. Everolimus plus octreotide long-acting repeatable for the treatment of advanced neuroendocrine tumours associated with carcinoid syndrome (RADIANT-2): a randomised, placebo-controlled, phase 3 study. Lancet. 2011;378:2005–2012. doi: 10.1016/S0140-6736(11)61742-X. [DOI] [PubMed] [Google Scholar]
- 96.Yao JC, Shah MH, Ito T, Bohas CL, Wolin EM, Van Cutsem E, Hobday TJ, Okusaka T, Capdevila J, de Vries EG, et al. Everolimus for advanced pancreatic neuroendocrine tumors. N Engl J Med. 2011;364:514–523. doi: 10.1056/NEJMoa1009290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Duran I, Kortmansky J, Singh D, Hirte H, Kocha W, Goss G, Le L, Oza A, Nicklee T, Ho J, et al. A phase II clinical and pharmacodynamic study of temsirolimus in advanced neuroendocrine carcinomas. Br J Cancer. 2006;95:1148–1154. doi: 10.1038/sj.bjc.6603419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Maehama T, Dixon JE. PTEN: a tumour suppressor that functions as a phospholipid phosphatase. Trends Cell Biol. 1999;9:125–128. doi: 10.1016/s0962-8924(99)01519-6. [DOI] [PubMed] [Google Scholar]
- 99.Hafsi S, Pezzino FM, Candido S, Ligresti G, Spandidos DA, Soua Z, McCubrey JA, Travali S, Libra M. Gene alterations in the PI3K/PTEN/AKT pathway as a mechanism of drug-resistance (review) Int J Oncol. 2012;40:639–644. doi: 10.3892/ijo.2011.1312. [DOI] [PubMed] [Google Scholar]
- 100.Zhang S, Yu D. PI(3)king apart PTEN’s role in cancer. Clin Cancer Res. 2010;16:4325–4330. doi: 10.1158/1078-0432.CCR-09-2990. [DOI] [PubMed] [Google Scholar]
- 101.Ramaswamy S, Nakamura N, Vazquez F, Batt DB, Perera S, Roberts TM, Sellers WR. Regulation of G1 progression by the PTEN tumor suppressor protein is linked to inhibition of the phosphatidylinositol 3-kinase/Akt pathway. Proc Natl Acad Sci USA. 1999;96:2110–2115. doi: 10.1073/pnas.96.5.2110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Myers MP, Pass I, Batty IH, Van der Kaay J, Stolarov JP, Hemmings BA, Wigler MH, Downes CP, Tonks NK. The lipid phosphatase activity of PTEN is critical for its tumor supressor function. Proc Natl Acad Sci USA. 1998;95:13513–13518. doi: 10.1073/pnas.95.23.13513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Furnari FB, Huang HJ, Cavenee WK. The phosphoinositol phosphatase activity of PTEN mediates a serum-sensitive G1 growth arrest in glioma cells. Cancer Res. 1998;58:5002–5008. [PubMed] [Google Scholar]
- 104.Putz U, Howitt J, Doan A, Goh CP, Low LH, Silke J, Tan SS. The tumor suppressor PTEN is exported in exosomes and has phosphatase activity in recipient cells. Sci Signal. 2012;5:ra70. doi: 10.1126/scisignal.2003084. [DOI] [PubMed] [Google Scholar]
- 105.Tamura M, Gu J, Takino T, Yamada KM. Tumor suppressor PTEN inhibition of cell invasion, migration, and growth: differential involvement of focal adhesion kinase and p130Cas. Cancer Res. 1999;59:442–449. [PubMed] [Google Scholar]
- 106.Kim JS, Xu X, Li H, Solomon D, Lane WS, Jin T, Waldman T. Mechanistic analysis of a DNA damage-induced, PTEN-dependent size checkpoint in human cells. Mol Cell Biol. 2011;31:2756–2771. doi: 10.1128/MCB.01323-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Wang L, Ignat A, Axiotis CA. Differential expression of the PTEN tumor suppressor protein in fetal and adult neuroendocrine tissues and tumors: progressive loss of PTEN expression in poorly differentiated neuroendocrine neoplasms. Appl Immunohistochem Mol Morphol. 2002;10:139–146. doi: 10.1097/00129039-200206000-00008. [DOI] [PubMed] [Google Scholar]
- 108.Samani AA, Yakar S, LeRoith D, Brodt P. The role of the IGF system in cancer growth and metastasis: overview and recent insights. Endocr Rev. 2007;28:20–47. doi: 10.1210/er.2006-0001. [DOI] [PubMed] [Google Scholar]
- 109.Furukawa M, Raffeld M, Mateo C, Sakamoto A, Moody TW, Ito T, Venzon DJ, Serrano J, Jensen RT. Increased expression of insulin-like growth factor I and/or its receptor in gastrinomas is associated with low curability, increased growth, and development of metastases. Clin Cancer Res. 2005;11:3233–3242. doi: 10.1158/1078-0432.CCR-04-1915. [DOI] [PubMed] [Google Scholar]
- 110.Nilsson O, Wängberg B, Theodorsson E, Skottner A, Ahlman H. Presence of IGF-I in human midgut carcinoid tumours--an autocrine regulator of carcinoid tumour growth? Int J Cancer. 1992;51:195–203. doi: 10.1002/ijc.2910510206. [DOI] [PubMed] [Google Scholar]
- 111.Nilsson O, Wängberg B, McRae A, Dahlström A, Ahlman H. Growth factors and carcinoid tumours. Acta Oncol. 1993;32:115–124. doi: 10.3109/02841869309083899. [DOI] [PubMed] [Google Scholar]
- 112.Wulbrand U, Wied M, Zöfel P, Göke B, Arnold R, Fehmann H. Growth factor receptor expression in human gastroenteropancreatic neuroendocrine tumours. Eur J Clin Invest. 1998;28:1038–1049. doi: 10.1046/j.1365-2362.1998.00397.x. [DOI] [PubMed] [Google Scholar]
- 113.von Wichert G, Jehle PM, Hoeflich A, Koschnick S, Dralle H, Wolf E, Wiedenmann B, Boehm BO, Adler G, Seufferlein T. Insulin-like growth factor-I is an autocrine regulator of chromogranin A secretion and growth in human neuroendocrine tumor cells. Cancer Res. 2000;60:4573–4581. [PubMed] [Google Scholar]
- 114.Höpfner M, Baradari V, Huether A, Schöfl C, Scherübl H. The insulin-like growth factor receptor 1 is a promising target for novel treatment approaches in neuroendocrine gastrointestinal tumours. Endocr Relat Cancer. 2006;13:135–149. doi: 10.1677/erc.1.01090. [DOI] [PubMed] [Google Scholar]
- 115.Vitale L, Lenzi L, Huntsman SA, Canaider S, Frabetti F, Casadei R, Facchin F, Carinci P, Zannotti M, Coppola D, et al. Differential expression of alternatively spliced mRNA forms of the insulin-like growth factor 1 receptor in human neuroendocrine tumors. Oncol Rep. 2006;15:1249–1256. [PubMed] [Google Scholar]
- 116.Richardson PG, Mitsiades CS, Laubach JP, Lonial S, Chanan-Khan AA, Anderson KC. Inhibition of heat shock protein 90 (HSP90) as a therapeutic strategy for the treatment of myeloma and other cancers. Br J Haematol. 2011;152:367–379. doi: 10.1111/j.1365-2141.2010.08360.x. [DOI] [PubMed] [Google Scholar]
- 117.Gloesenkamp C, Nitzsche B, Lim AR, Normant E, Vosburgh E, Schrader M, Ocker M, Scherübl H, Höpfner M. Heat shock protein 90 is a promising target for effective growth inhibition of gastrointestinal neuroendocrine tumors. Int J Oncol. 2012;40:1659–1667. doi: 10.3892/ijo.2012.1328. [DOI] [PubMed] [Google Scholar]
- 118.Gilbert JA, Adhikari LJ, Lloyd RV, Halfdanarson TR, Muders MH, Ames MM. Molecular markers for novel therapeutic strategies in pancreatic endocrine tumors. Pancreas. 2013;42:411–421. doi: 10.1097/MPA.0b013e31826cb243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Rowinsky EK, Youssoufian H, Tonra JR, Solomon P, Burtrum D, Ludwig DL. IMC-A12, a human IgG1 monoclonal antibody to the insulin-like growth factor I receptor. Clin Cancer Res. 2007;13:5549s–5555s. doi: 10.1158/1078-0432.CCR-07-1109. [DOI] [PubMed] [Google Scholar]
- 120.Folkman J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat Med. 1995;1:27–31. doi: 10.1038/nm0195-27. [DOI] [PubMed] [Google Scholar]
- 121.Berger DP, Herbstritt L, Dengler WA, Marmé D, Mertelsmann R, Fiebig HH. Vascular endothelial growth factor (VEGF) mRNA expression in human tumor models of different histologies. Ann Oncol. 1995;6:817–825. doi: 10.1093/oxfordjournals.annonc.a059322. [DOI] [PubMed] [Google Scholar]
- 122.Lawnicka H, Stepień H, Wyczółkowska J, Kolago B, Kunert-Radek J, Komorowski J. Effect of somatostatin and octreotide on proliferation and vascular endothelial growth factor secretion from murine endothelial cell line (HECa10) culture. Biochem Biophys Res Commun. 2000;268:567–571. doi: 10.1006/bbrc.2000.2119. [DOI] [PubMed] [Google Scholar]
- 123.Kuo CJ, Farnebo F, Yu EY, Christofferson R, Swearingen RA, Carter R, von Recum HA, Yuan J, Kamihara J, Flynn E, et al. Comparative evaluation of the antitumor activity of antiangiogenic proteins delivered by gene transfer. Proc Natl Acad Sci USA. 2001;98:4605–4610. doi: 10.1073/pnas.081615298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Matsumoto T, Claesson-Welsh L. VEGF receptor signal transduction. Sci STKE. 2001;2001:re21. doi: 10.1126/stke.2001.112.re21. [DOI] [PubMed] [Google Scholar]
- 125.Klagsbrun M, Takashima S, Mamluk R. The role of neuropilin in vascular and tumor biology. Adv Exp Med Biol. 2002;515:33–48. doi: 10.1007/978-1-4615-0119-0_3. [DOI] [PubMed] [Google Scholar]
- 126.Ferrara N, Gerber HP, LeCouter J. The biology of VEGF and its receptors. Nat Med. 2003;9:669–676. doi: 10.1038/nm0603-669. [DOI] [PubMed] [Google Scholar]
- 127.Treiber G, Wex T, Röcken C, Fostitsch P, Malfertheiner P. Impact of biomarkers on disease survival and progression in patients treated with octreotide for advanced hepatocellular carcinoma. J Cancer Res Clin Oncol. 2006;132:699–708. doi: 10.1007/s00432-006-0118-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Dimitroulopoulos D, Xinopoulos D, Tsamakidis K, Zisimopoulos A, Andriotis E, Panagiotakos D, Fotopoulou A, Chrysohoou C, Bazinis A, Daskalopoulou D, et al. Long acting octreotide in the treatment of advanced hepatocellular cancer and overexpression of somatostatin receptors: randomized placebo-controlled trial. World J Gastroenterol. 2007;13:3164–3170. doi: 10.3748/wjg.v13.i23.3164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Ho QT, Kuo CJ. Vascular endothelial growth factor: biology and therapeutic applications. Int J Biochem Cell Biol. 2007;39:1349–1357. doi: 10.1016/j.biocel.2007.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Modlin IM, Oberg K, Chung DC, Jensen RT, de Herder WW, Thakker RV, Caplin M, Delle Fave G, Kaltsas GA, Krenning EP, et al. Gastroenteropancreatic neuroendocrine tumours. Lancet Oncol. 2008;9:61–72. doi: 10.1016/S1470-2045(07)70410-2. [DOI] [PubMed] [Google Scholar]
- 131.Faivre S, Delbaldo C, Vera K, Robert C, Lozahic S, Lassau N, Bello C, Deprimo S, Brega N, Massimini G, et al. Safety, pharmacokinetic, and antitumor activity of SU11248, a novel oral multitarget tyrosine kinase inhibitor, in patients with cancer. J Clin Oncol. 2006;24:25–35. doi: 10.1200/JCO.2005.02.2194. [DOI] [PubMed] [Google Scholar]
- 132.Kulke MH, Lenz HJ, Meropol NJ, Posey J, Ryan DP, Picus J, Bergsland E, Stuart K, Tye L, Huang X, et al. Activity of sunitinib in patients with advanced neuroendocrine tumors. J Clin Oncol. 2008;26:3403–3410. doi: 10.1200/JCO.2007.15.9020. [DOI] [PubMed] [Google Scholar]
- 133.Raymond E, Dahan L, Raoul JL, Bang YJ, Borbath I, Lombard-Bohas C, Valle J, Metrakos P, Smith D, Vinik A, et al. Sunitinib malate for the treatment of pancreatic neuroendocrine tumors. N Engl J Med. 2011;364:501–513. doi: 10.1056/NEJMoa1003825. [DOI] [PubMed] [Google Scholar]
- 134.Yao JC, Phan A, Hoff PM, Chen HX, Charnsangavej C, Yeung SC, Hess K, Ng C, Abbruzzese JL, Ajani JA. Targeting vascular endothelial growth factor in advanced carcinoid tumor: a random assignment phase II study of depot octreotide with bevacizumab and pegylated interferon alpha-2b. J Clin Oncol. 2008;26:1316–1323. doi: 10.1200/JCO.2007.13.6374. [DOI] [PubMed] [Google Scholar]
- 135.Faivre S, Sablin MP, Dreyer C, Raymond E. Novel anticancer agents in clinical trials for well-differentiated neuroendocrine tumors. Endocrinol Metab Clin North Am. 2010;39:811–826. doi: 10.1016/j.ecl.2010.09.006. [DOI] [PubMed] [Google Scholar]
- 136.Linsley PS, Bradshaw J, Greene J, Peach R, Bennett KL, Mittler RS. Intracellular trafficking of CTLA-4 and focal localization towards sites of TCR engagement. Immunity. 1996;4:535–543. doi: 10.1016/s1074-7613(00)80480-x. [DOI] [PubMed] [Google Scholar]
- 137.Takahashi T, Tagami T, Yamazaki S, Uede T, Shimizu J, Sakaguchi N, Mak TW, Sakaguchi S. Immunologic self-tolerance maintained by CD25(+)CD4(+) regulatory T cells constitutively expressing cytotoxic T lymphocyte-associated antigen 4. J Exp Med. 2000;192:303–310. doi: 10.1084/jem.192.2.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Freeman GJ, Long AJ, Iwai Y, Bourque K, Chernova T, Nishimura H, Fitz LJ, Malenkovich N, Okazaki T, Byrne MC, et al. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J Exp Med. 2000;192:1027–1034. doi: 10.1084/jem.192.7.1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Latchman Y, Wood CR, Chernova T, Chaudhary D, Borde M, Chernova I, Iwai Y, Long AJ, Brown JA, Nunes R, et al. PD-L2 is a second ligand for PD-1 and inhibits T cell activation. Nat Immunol. 2001;2:261–268. doi: 10.1038/85330. [DOI] [PubMed] [Google Scholar]
- 140.Ishida Y, Agata Y, Shibahara K, Honjo T. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death. EMBO J. 1992;11:3887–3895. doi: 10.1002/j.1460-2075.1992.tb05481.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Keir ME, Liang SC, Guleria I, Latchman YE, Qipo A, Albacker LA, Koulmanda M, Freeman GJ, Sayegh MH, Sharpe AH. Tissue expression of PD-L1 mediates peripheral T cell tolerance. J Exp Med. 2006;203:883–895. doi: 10.1084/jem.20051776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Okazaki T, Honjo T. PD-1 and PD-1 ligands: from discovery to clinical application. Int Immunol. 2007;19:813–824. doi: 10.1093/intimm/dxm057. [DOI] [PubMed] [Google Scholar]
- 143.Yokosuka T, Takamatsu M, Kobayashi-Imanishi W, Hashimoto-Tane A, Azuma M, Saito T. Programmed cell death 1 forms negative costimulatory microclusters that directly inhibit T cell receptor signaling by recruiting phosphatase SHP2. J Exp Med. 2012;209:1201–1217. doi: 10.1084/jem.20112741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Jorissen RN, Walker F, Pouliot N, Garrett TP, Ward CW, Burgess AW. Epidermal growth factor receptor: mechanisms of activation and signalling. Exp Cell Res. 2003;284:31–53. doi: 10.1016/s0014-4827(02)00098-8. [DOI] [PubMed] [Google Scholar]
- 145.Shah T, Hochhauser D, Frow R, Quaglia A, Dhillon AP, Caplin ME. Epidermal growth factor receptor expression and activation in neuroendocrine tumours. J Neuroendocrinol. 2006;18:355–360. doi: 10.1111/j.1365-2826.2006.01425.x. [DOI] [PubMed] [Google Scholar]
- 146.Papouchado B, Erickson LA, Rohlinger AL, Hobday TJ, Erlichman C, Ames MM, Lloyd RV. Epidermal growth factor receptor and activated epidermal growth factor receptor expression in gastrointestinal carcinoids and pancreatic endocrine carcinomas. Mod Pathol. 2005;18:1329–1335. doi: 10.1038/modpathol.3800427. [DOI] [PubMed] [Google Scholar]
- 147.Srivastava A, Alexander J, Lomakin I, Dayal Y. Immunohistochemical expression of transforming growth factor alpha and epidermal growth factor receptor in pancreatic endocrine tumors. Hum Pathol. 2001;32:1184–1189. doi: 10.1053/hupa.2001.28959. [DOI] [PubMed] [Google Scholar]
- 148.Krishnamurthy S, Dayal Y. Immunohistochemical expression of transforming growth factor alpha and epidermal growth factor receptor in gastrointestinal carcinoids. Am J Surg Pathol. 1997;21:327–333. doi: 10.1097/00000478-199703000-00009. [DOI] [PubMed] [Google Scholar]
- 149.Nilsson O, Wängberg B, Kölby L, Schultz GS, Ahlman H. Expression of transforming growth factor alpha and its receptor in human neuroendocrine tumours. Int J Cancer. 1995;60:645–651. doi: 10.1002/ijc.2910600514. [DOI] [PubMed] [Google Scholar]
- 150.Shimizu T, Tanaka S, Haruma K, Kitadai Y, Yoshihara M, Sumii K, Kajiyama G, Shimamoto F. Growth characteristics of rectal carcinoid tumors. Oncology. 2000;59:229–237. doi: 10.1159/000012166. [DOI] [PubMed] [Google Scholar]
- 151.Hobday TJ, Holen K, Donehower RC, Camoriano J, Kim G, Picus J, Philip P, Lloyd R, Mahoney M, Erlichman C. A phase II trial of gefitinib in patients (pts) with progressive metastatic neuroendocrine tumours (NET): a phase II consortium (P2C) study. J Clin Oncol. 2006;24:189S. [Google Scholar]
- 152.Beenken A, Mohammadi M. The FGF family: biology, pathophysiology and therapy. Nat Rev Drug Discov. 2009;8:235–253. doi: 10.1038/nrd2792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Korc M, Friesel RE. The role of fibroblast growth factors in tumor growth. Curr Cancer Drug Targets. 2009;9:639–651. doi: 10.2174/156800909789057006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Cronauer MV, Schulz WA, Seifert HH, Ackermann R, Burchardt M. Fibroblast growth factors and their receptors in urological cancers: basic research and clinical implications. Eur Urol. 2003;43:309–319. doi: 10.1016/s0302-2838(03)00005-8. [DOI] [PubMed] [Google Scholar]
- 155.Rusnati M, Presta M. Fibroblast growth factors/fibroblast growth factor receptors as targets for the development of anti-angiogenesis strategies. Curr Pharm Des. 2007;13:2025–2044. doi: 10.2174/138161207781039689. [DOI] [PubMed] [Google Scholar]
- 156.Wu X, Yan Q, Huang Y, Huang H, Su Z, Xiao J, Zeng Y, Wang Y, Nie C, Yang Y, et al. Isolation of a novel basic FGF-binding peptide with potent antiangiogenetic activity. J Cell Mol Med. 2010;14:351–356. doi: 10.1111/j.1582-4934.2008.00506.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Trueb B. Biology of FGFRL1, the fifth fibroblast growth factor receptor. Cell Mol Life Sci. 2011;68:951–964. doi: 10.1007/s00018-010-0576-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Chaudhry A, Funa K, Oberg K. Expression of growth factor peptides and their receptors in neuroendocrine tumors of the digestive system. Acta Oncol. 1993;32:107–114. doi: 10.3109/02841869309083898. [DOI] [PubMed] [Google Scholar]
- 159.Yarden Y, Kuang WJ, Yang-Feng T, Coussens L, Munemitsu S, Dull TJ, Chen E, Schlessinger J, Francke U, Ullrich A. Human proto-oncogene c-kit: a new cell surface receptor tyrosine kinase for an unidentified ligand. EMBO J. 1987;6:3341–3351. doi: 10.1002/j.1460-2075.1987.tb02655.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Andrae J, Gallini R, Betsholtz C. Role of platelet-derived growth factors in physiology and medicine. Genes Dev. 2008;22:1276–1312. doi: 10.1101/gad.1653708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Anderberg C, Li H, Fredriksson L, Andrae J, Betsholtz C, Li X, Eriksson U, Pietras K. Paracrine signaling by platelet-derived growth factor-CC promotes tumor growth by recruitment of cancer-associated fibroblasts. Cancer Res. 2009;69:369–378. doi: 10.1158/0008-5472.CAN-08-2724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Jain RK, Booth MF. What brings pericytes to tumor vessels? J Clin Invest. 2003;112:1134–1136. doi: 10.1172/JCI20087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Dong J, Grunstein J, Tejada M, Peale F, Frantz G, Liang WC, Bai W, Yu L, Kowalski J, Liang X, et al. VEGF-null cells require PDGFR alpha signaling-mediated stromal fibroblast recruitment for tumorigenesis. EMBO J. 2004;23:2800–2810. doi: 10.1038/sj.emboj.7600289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Crosier PS, Ricciardi ST, Hall LR, Vitas MR, Clark SC, Crosier KE. Expression of isoforms of the human receptor tyrosine kinase c-kit in leukemic cell lines and acute myeloid leukemia. Blood. 1993;82:1151–1158. [PubMed] [Google Scholar]
- 165.Chen SQ, Xiong AQ. The progress and implication of stem cell factor. Jichuyixue Yu Linchuang. 2002;22:385–390. [Google Scholar]
- 166.Yao JC, Zhang JX, Rashid A, Yeung SC, Szklaruk J, Hess K, Xie K, Ellis L, Abbruzzese JL, Ajani JA. Clinical and in vitro studies of imatinib in advanced carcinoid tumors. Clin Cancer Res. 2007;13:234–240. doi: 10.1158/1078-0432.CCR-06-1618. [DOI] [PubMed] [Google Scholar]
- 167.Lankat-Buttgereit B, Hörsch D, Barth P, Arnold R, Blöcker S, Göke R. Effects of the tyrosine kinase inhibitor imatinib on neuroendocrine tumor cell growth. Digestion. 2005;71:131–140. doi: 10.1159/000084647. [DOI] [PubMed] [Google Scholar]
- 168.Wilhelm SM, Carter C, Tang L, Wilkie D, McNabola A, Rong H, Chen C, Zhang X, Vincent P, McHugh M, et al. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004;64:7099–7109. doi: 10.1158/0008-5472.CAN-04-1443. [DOI] [PubMed] [Google Scholar]
- 169.Fabian MA, Biggs WH, Treiber DK, Atteridge CE, Azimioara MD, Benedetti MG, Carter TA, Ciceri P, Edeen PT, Floyd M, et al. A small molecule-kinase interaction map for clinical kinase inhibitors. Nat Biotechnol. 2005;23:329–336. doi: 10.1038/nbt1068. [DOI] [PubMed] [Google Scholar]
- 170.Wilhelm SM, Dumas J, Adnane L, Lynch M, Carter CA, Schütz G, Thierauch KH, Zopf D. Regorafenib (BAY 73-4506): a new oral multikinase inhibitor of angiogenic, stromal and oncogenic receptor tyrosine kinases with potent preclinical antitumor activity. Int J Cancer. 2011;129:245–255. doi: 10.1002/ijc.25864. [DOI] [PubMed] [Google Scholar]
- 171.Sonpavde G, Hutson TE. Pazopanib: a novel multitargeted tyrosine kinase inhibitor. Curr Oncol Rep. 2007;9:115–119. doi: 10.1007/s11912-007-0007-2. [DOI] [PubMed] [Google Scholar]
- 172.Phan AT, Yao JC, Fogelman DR, Hess KR, Ng CS, Bullock SA, Malinoski P, Regan E, Kulke M. A prospective, multi-institutional phase II study of GW786034 (pazopanib) and depot octreotide (Sandostatin LAR) in advanced low-grade neuroendocrine carcinoma (LGNEC) J Clin Oncol. 2010;28(suppl 15):abstr 4001. [Google Scholar]
- 173.Chaudhry A, Papanicolaou V, Oberg K, Heldin CH, Funa K. Expression of platelet-derived growth factor and its receptors in neuroendocrine tumors of the digestive system. Cancer Res. 1992;52:1006–1012. [PubMed] [Google Scholar]
- 174.Chaudhry A, Oberg K, Gobl A, Heldin CH, Funa K. Expression of transforming growth factors beta 1, beta 2, beta 3 in neuroendocrine tumors of the digestive system. Anticancer Res. 1994;14:2085–2091. [PubMed] [Google Scholar]
- 175.Christofori G, Naik P, Hanahan D. Vascular endothelial growth factor and its receptors, flt-1 and flk-1, are expressed in normal pancreatic islets and throughout islet cell tumorigenesis. Mol Endocrinol. 1995;9:1760–1770. doi: 10.1210/mend.9.12.8614412. [DOI] [PubMed] [Google Scholar]
- 176.La Rosa S, Chiaravalli AM, Capella C, Uccella S, Sessa F. Immunohistochemical localization of acidic fibroblast growth factor in normal human enterochromaffin cells and related gastrointestinal tumours. Virchows Arch. 1997;430:117–124. doi: 10.1007/BF01008032. [DOI] [PubMed] [Google Scholar]
- 177.Van Gompel JJ, Chen H. Insulin-like growth factor 1 signaling in human gastrointestinal carcinoid tumor cells. Surgery. 2004;136:1297–1302. doi: 10.1016/j.surg.2004.06.061. [DOI] [PubMed] [Google Scholar]
- 178.Hofland LJ, Lamberts SW. Somatostatin receptors and disease: role of receptor subtypes. Baillieres Clin Endocrinol Metab. 1996;10:163–176. doi: 10.1016/s0950-351x(96)80362-4. [DOI] [PubMed] [Google Scholar]
- 179.Welin S, Fjällskog ML, Saras J, Eriksson B, Janson ET. Expression of tyrosine kinase receptors in malignant midgut carcinoid tumors. Neuroendocrinology. 2006;84:42–48. doi: 10.1159/000096294. [DOI] [PubMed] [Google Scholar]