

Small herbaceous species are vulnerable to water stress but adjusted to their habitat conditions and internally coordinated to avoid hydraulic failure.
Abstract
The requirements of the water transport system of small herbaceous species differ considerably from those of woody species. Despite their ecological importance for many biomes, knowledge regarding herb hydraulics remains very limited. We compared key hydraulic features (vulnerability to drought-induced hydraulic decline, pressure-volume relations, onset of cellular damage, in situ variation of water potential, and stomatal conductance) of three Ranunculus species differing in their soil humidity preferences and ecological amplitude. All species were very vulnerable to water stress (50% reduction in whole-leaf hydraulic conductance [kleaf] at −0.2 to −0.8 MPa). In species with narrow ecological amplitude, the drought-exposed Ranunculus bulbosus was less vulnerable to desiccation (analyzed via loss of kleaf and turgor loss point) than the humid-habitat Ranunculus lanuginosus. Accordingly, water stress-exposed plants from the broad-amplitude Ranunculus acris revealed tendencies toward lower vulnerability to water stress (e.g. osmotic potential at full turgor, cell damage, and stomatal closure) than conspecific plants from the humid site. We show that small herbs can adjust to their habitat conditions on interspecific and intraspecific levels in various hydraulic parameters. The coordination of hydraulic thresholds (50% and 88% loss of kleaf, turgor loss point, and minimum in situ water potential) enabled the study species to avoid hydraulic failure and damage to living cells. Reversible recovery of hydraulic conductance, desiccation-tolerant seeds, or rhizomes may allow them to prioritize toward a more efficient but vulnerable water transport system while avoiding the severe effects that water stress poses on woody species.
The resistance of terrestrial plant species to water stress is an important determinant of their spatial distribution (Engelbrecht et al., 2007), and in a globally changing climate, resistance to water stress-induced damage to the water transport system is an essential parameter for the prediction of species survival and future changes in biodiversity and species composition. The hydraulic vulnerability to drought-induced embolism in woody species is adjusted to their environmental conditions on a global scale (Choat et al., 2012), but increased hydraulic safety entails several tradeoffs, such as transport efficiency, as plants can reduce the risk of hydraulic failure by the formation of smaller and hydraulically less efficient conduits (Tyree et al., 1994; Wagner et al., 1998, Gleason et al., 2016).
While many studies have dealt with the hydraulics of woody species (Maherali et al., 2004; Choat et al., 2012), the hydraulic characteristics of small herbaceous species are largely understudied, and our knowledge regarding their water transport system, including vulnerability to drought-induced loss of conductivity, remains very limited (Kocacinar and Sage, 2003; Brodribb and Holbrook, 2004; Iovi et al., 2009; Holloway-Phillips and Brodribb, 2011a; Tixier et al., 2013). Yet, herbs play an important ecological role in many biomes, such as grasslands and the alpine zone (Billings and Mooney, 1968; Gilliam, 2007; Scholz et al., 2010), and represent a significant percentage of crop plants (Monfreda et al., 2008); therefore, they are interesting from an ecophysiological viewpoint.
The requirements of the water transport system of herbs differ in several ways from those of woody species (Mencuccini, 2003). First, transport distances and supported leaf areas are much smaller in herbs, and reversible extraxylary limitations (Brodribb and Holbrook, 2005; Kim and Steudle, 2007) may have a stronger effect on hydraulic conductivity. Second, herbs often do not feature an essential, lignified main axis designed for long-term functioning, in which hydraulic failure may mean whole-plant mortality. Third, small plants may more easily restore water transport capacity in the case of embolism formation, because, due to smaller size, a water potential (Ψ) at or near zero can be reached quickly under favorable conditions due to positive root pressure (Tyree and Sperry, 1989; Tyree and Ewers, 1991; Stiller and Sperry, 2002; Tyree and Zimmermann, 2002; Ganthaler and Mayr, 2015). Thus, herbs may not be as threatened by drought-induced failure as taller woody species.
Hydraulic failure occurs when vulnerability thresholds are exceeded under water stress and the xylem tension is suddenly released as water is replaced by air, effectively blocking water transport in the affected conduits (Tyree and Zimmermann, 2002). Vulnerability to drought-induced embolism can thus be analyzed hydraulically by measuring decreases in conductance during increasing water stress (Sperry et al., 1988a; Cochard, 2002; Cochard et al., 2013), by noninvasive imaging of xylem water content (Choat et al., 2010; Brodersen et al., 2013; Cochard et al., 2015), or indirectly by monitoring of ultrasonic acoustic emissions (UE) which occur during the spontaneous energy release at embolism formation (Milburn and Johnson, 1966; Mayr and Rosner, 2011; Ponomarenko et al., 2014). However, the usability of UE analysis to noninvasively study hydraulic vulnerability remains under discussion (Sandford and Grace, 1985; Kikuta, 2003; Wolkerstorfer et al., 2012; Cochard et al., 2013; Rosner, 2015).
On the leaf level, desiccation may induce turgor loss, decreases in mesophyll conductance (Kim and Steudle, 2007; Scoffoni et al., 2014), or cell collapse (Blackman et al., 2010; Holloway-Phillips and Brodribb, 2011a), and limit hydraulic conductance, gas exchange, and thus photosynthesis before embolism occurs. When water stress increases further, living cells may incur damage resulting in tissue and leaf mortality.
In this study, we compared key hydraulic features (vulnerability to drought-induced loss of conductance as measured via hydraulic flow and xylem staining, pressure-volume relations, and the onset of cellular damage) of three species from the genus Ranunculus, which is distributed almost worldwide and contains species adapted to dry and humid habitats as well as species with broad ecological amplitude. We hypothesized that herbaceous species would be more vulnerable to water stress than woody species but also would show interspecific and intraspecific adjustments in hydraulic vulnerability based on the water availability of their respective habitats. Namely, the hydraulics of species with narrow ecological amplitude were hypothesized to reflect their respective habitat conditions, and broad-amplitude species should show adequate intraspecific variation to enable growth in dry and humid habitats.
RESULTS
Volumetric soil water content (SWC) at field capacity was 40.4% (dry site), 51% (humid site; Ranunculus acris), and 30.4% (humid site; Ranunculus lanuginosus). Relative SWC after 7 d of rain-free weather was above 90% at the humid sites and 43% to 58% at the dry site (Table I). The low field capacity SWC of the second humid site was due to the soil composition (thin layer of humus and litter on top of gravel substrate), yet the SWC remained constant due to the humid conditions inside the gorge. The reason for a relative soil water content (SWCrel) of 100% or greater at this site may lie in water seeping through the slope from uphill soils. Ecological indicator values for the study species and cooccurring species at the sampling sites supported the observed differences in water availability: averaged humidity values were 4.4 at the dry site and notably higher at 5.5 (R. acris) and 5.7 (R. lanuginosus) at the humid sites (Table I). At the dry site, Ranunculus bulbosus (humidity index of 3; indicator for dry sites) and R. acris (humidity index of 6; indicator for humid sites but broad amplitude) were accompanied by dry-habitat species such as Bromus erectus, Knautia arvensis, and Lotus corniculatus. In contrast, at the humid sites, R. lanuginosus (humidity index of 6) and R. acris were cooccurring with other species typical for moist habitats, including Polygonum bistorta, Rumex conglomeratus, Urtica dioica, and Solanum dulcamara (Supplemental Table S1).
Table I. SWCrel (measured in summer 2014 and 2015; n = 10), averaged humidity value (n = 10–13), kleaf max (n = 5), TLP (n = 5), diurnal Ψmin (n = 3 [Fig. 4; Supplemental Fig. S2]), and diurnal gs max (n = 5 [Fig. 4; Supplemental Fig. S2]).
Values are means ± se. Letters indicate significant differences in each row (P < 0.05).
| Species | SWCrel (2014) | SWCrel (2015) | Humidity Value | TLP | kleaf max | Ψmin | gs max |
|---|---|---|---|---|---|---|---|
| % | % | MPa | mmol m−2 s−1 MPa−1 | MPa | mmol m−2 s−1 | ||
| R. bulbosus (dry) | 57.57 ± 4.03 a | 43.29 ± 2.42 a | 4.40 ± 0.34 a | −1.43 ± 0.02 a | 28.00 ± 0.78 a | −1.70 ± 0.03 a | 537.7 ± 105.3 a |
| R. lanuginosus (humid) | 102.72 ± 6.14 b | 98.34 ± 2.31 b | 5.69 ± 0.26 b | −1.14 ± 0.05 b | 27.77 ± 1.12 a | −0.42 ± 0.12 b | 492.1 ± 86.1 a |
| R. acris (dry) | 57.57 ± 4.03 a | 43.29 ± 2.42 a | 4.40 ± 0.34 a | −1.72 ± 0.08 c | 23.40 ± 0.42 a | −2.20 ± 0.06 c | 756.6 ± 195.9 a |
| R. acris (humid) | 93.40 ± 1.87 b | 90.59 ± 1.30 c | 5.50 ± 0.23 b | −1.63 ± 0.05 a,c | 24.06 ± 0.78 a | −1.48 ± 0.15 a | 885.0 ± 37.3 a |
The turgor loss point (TLP) was least negative in R. lanuginosus at a leaf water potential (Ψleaf) of −1.1 ± 0.1 MPa and was reached at −1.4 ± 0.0 MPa in R. bulbosus and at −1.6 ± 0.1 to −1.7 ± 0.1 MPa in R. acris (Table I). The osmotic potential (Π0) followed a similar trend at −0.9 ± 0.1 MPa (R. lanuginosus), −1.3 ± 0.0 MPa (R. bulbosus), and −1.4 ± 0.1 to −1.5 ± 0.1 MPa (R. acris). In contrast, the modulus of elasticity was lowest (i.e. cells were more elastic) at 9.6 ± 1.9 MPa in R. lanuginosus and 1.5 to 2.5 times higher in the other species (R. bulbosus, 16.7 ± 1.3 MPa; and R. acris, 20.4 ± 4.4 to 24.9 ± 3.6 MPa).
Maximum whole-leaf hydraulic conductance (kleaf max) was between 23.4 and 28 mmol m−2 s−1 MPa−1 and did not differ between species (Table I). Hydraulically measured water potential at 50% loss of leaf hydraulic conductance (Pk50) was least negative in R. lanuginosus (−0.2 MPa) and more negative in R. acris and R. bulbosus (−0.6 to −0.8 MPa; Table II; Fig. 1). Similarly, water potential at 88% loss of leaf hydraulic conductance (Pk88) was highest in R. lanuginosus (−0.4 MPa) and significantly more negative in the other two species (−1.5 to −3.3 MPa; Table II; Fig. 1).
Table II. Water potential at 50 and 88% loss of leaf hydraulic conductance (Pk50 and Pk88; MPa), and 12% of cellular lysis (P12CL; MPa).
Thresholds were computed after fitting the reparameterized Weibull function (Ogle et al., 2009) to vulnerability curves. Values are means (MPa) and 95% confidence intervals. Letters indicate significant differences in each row (P < 0.05).
| Species | Leaf Hydraulic Vulnerability |
Electrolyte Leakage |
|
|---|---|---|---|
| Pk50 | Pk88 | P12CL | |
| R. bulbosus (dry) | −0.57 (−0.63, −0.48) a | −1.53 (<−1.55, −1.31) a | −1.11 (−1.53, −0.84) a |
| R. lanuginosus (humid) | −0.15 (−0.19, −0.10) b | −0.44 (−0.60, −0.35) b | −1.16 (−1.47, −0.88) a |
| R. acris (dry) | −0.82 (−1.08, −0.54) a | −3.27 (<−3.27, −2.64) c | −2.02 (−2.43, −1.64) b |
| R. acris (humid) | −0.73 (−0.93, −0.50) a | −2.84 (<−3.15, −2.25) c | −2.02 (−2.29, −1.62) b |
Figure 1.

kleaf versus Ψ in R. bulbosus, R. lanuginosus, and dry and humid R. acris populations measured with the non-steady state leaf rehydration technique. Vertical lines indicate fitted hydraulic vulnerability parameters Pk50 (solid) and Pk88 (dashed). Curves were extrapolated using the same equation and parameters used for vulnerability curves (Eq. 2).
Conductive xylem staining in R. acris (dry) showed the first notable signs of embolism in minor leaf veins and vascular bundles at −2 MPa, with significant losses of even first-order leaf veins by −2.7 MPa (Fig. 2).
Figure 2.

Stained leaf laminas and petiole cross sections of R. acris (dry) indicating loss of hydraulic conductance at increasing levels of dehydration. Conducting elements appear red, while embolized conduits remain unstained. A, Leaf lamina undersides and petioles. Bars = 20 mm. B, Leaf lamina details. Bars = 5 mm. C, Petiole cross sections approximately 3 cm from the laminas. Bars = 200 µm. D, Vascular bundle details from cross sections. Bars = 50 µm.
Significant cellular lysis started at more negative Ψ than Pk50, with water potential at 12% of cellular lysis (P12CL) between −1.1 and −2 MPa (Table II; Fig. 3), whereas thresholds for cell damage did not correspond to humidity conditions. In contrast, the onset of UE was observed at less negative Ψ than hydraulic vulnerability thresholds, with peak activity at −1.1 MPa or greater (Supplemental Table S2; Supplemental Fig. S1).
Figure 3.

Cellular lysis (percentage of maximum after autoclaving) versus Ψ in R. bulbosus, R. lanuginosus, and dry and humid R. acris populations. Vertical lines (dashed) indicate the onset of damage to living cells (P12CL). Curves were extrapolated using the same equation and parameters used for vulnerability curves (Eq. 2).
In situ, species reached minimum leaf water potential (Ψmin) of −0.4 ± 0.1 to −2.2 ± 0.1 MPa, whereby Ψmin was lower at the dry site (Table I; Fig. 4). In contrast, stomatal conductance (gs) exhibited considerable variability at more negative Ψ. Maximum stomatal conductance (gs max) was between 492.1 ± 86.1 and 885 ± 37.3 mmol m−2 s−1 and did not reflect soil humidity differences between sites (Table I; Fig. 4). The progression of Ψleaf and gs in the field (illustrated in Fig. 4 for R. acris) revealed that gs was reduced in all species when Ψmin was reached. Theoretical whole-leaf hydraulic conductance (kleaf) as calculated from measured Ψleaf and transpiration (E; based on measured gs and climate data from the closest weather station at approximately 3 km distance) agreed well with kleaf predicted from vulnerability curves. However, vapor pressure deficit and transpirational demand were expected to be lower in the lowest canopy layer than calculated here (Wohlfahrt et al., 2010). Both Ψleaf and gs were compared on representative, sunny days, but environmental conditions (wind or prolonged drought stress) may lead to more extreme values. Because of windy (R. bulbosus) or shady (R. lanuginosus; due to its understory gorge habitat) conditions during in situ measurements (Supplemental Fig. S2), diurnal variation data for only R. acris were used for further interpretation.
Figure 4.

Diurnal variation of Ψ (MPa; black circles, solid black lines) and gs (mmol m−2 s−1; red open circles, dashed red lines) in R. acris populations on dry and humid sites. Vertical lines (dashed) indicate meteorological dawn, and yellow areas indicate direct sunlight. Values are means ± se.
DISCUSSION
We found that all studied species were generally vulnerable to dehydration, but differences between species/populations reflected the humidity conditions of their respective habitats.
Sites differed in their water availability, as the SWCrel between dry and humid sites differed significantly after only 7 d without precipitation. At the dry site, SWCrel decreased to less than 60% of field capacity after only 1 week of rain-free weather, whereas the humid sites experienced hardly any reduction in soil humidity. The low SWC at the humid site of R. lanuginosus, which was due to soil composition, was compensated by the humid conditions inside the gorge, so that SWCrel did not decrease during the rain-free period. Species composition and ecological indicator values supported the distinction between dry and humid sites (Table I; Supplemental Table S1).
Herb Hydraulics
kleaf is known to vary widely across species and plant functional types, with an average kleaf of 11.5 ± 0.9 mmol m−2 s−1 MPa−1 across 107 woody and herbaceous species and with individual species ranging between 0.8 and 49 mmol m−2 s−1 MPa−1 (Sack and Holbrook, 2006). Leaf hydraulic efficiency in herbaceous crops was above average at 24.1 ± 2.2 mmol m−2 s−1 MPa−1 (Sack and Holbrook, 2006), and other, noncultivated herbaceous angiosperms in the literature had values between 2.7 and 22 mmol m−2 s−1 MPa−1 (Brodribb and Holbrook, 2004; Bunce, 2006; Galmes et al., 2007). Our study species were within the range known for herbaceous plants and, therefore, showed much higher hydraulic efficiency than woody species (Table I; Fig. 1).
Leaf hydraulic vulnerability studies on small herbaceous species found 50% reduction in kleaf between −1 and −1.8 MPa in perennial grasses (Brodribb and Holbrook, 2004; Holloway-Phillips and Brodribb, 2011a, 2011b). Woody species generally feature leaves that are less vulnerable to water stress, with an average Pk50 of −1.8 ± 0.1 MPa across 81 woody angiosperm species when measured with comparable methods (Nolf et al., 2015). Although some of these species also had very vulnerable leaves at Pk50 of −1 MPa or greater (Hao et al., 2008; Chen et al., 2009; Blackman et al., 2012; Guyot et al., 2012), the majority of previously studied species exhibited Pk50 of −1.5 MPa or less and down to −4.3 MPa (Sack and Holbrook, 2006; Blackman et al., 2010; Johnson et al., 2012; Nardini et al., 2012; Bucci et al., 2013). Accordingly, stems were typically also more vulnerable to drought-induced hydraulic failure in herbs (−1 to −3.8 MPa; Kocacinar and Sage, 2003; Rosenthal et al., 2010; Tixier et al., 2013; Nolf et al., 2014) than in woody species (−0.1 to −14.1 MPa, and −3.4 ± 0.1 MPa across 480 species; Choat et al., 2012).
Leaf hydraulic analysis in our study showed that all three Ranunculus spp. were very vulnerable to water stress, with Pk50 of −0.8 MPa or greater in kleaf across all species (Table II; Fig. 1). However, xylem staining indicated that early hydraulic conductivity losses at moderate Ψ were due to extraxylary, likely reversible, effects rather than xylem embolism. Stained petiole sections and leaves revealed loss of conductivity due to embolism and suggested an estimated 50% loss of xylem hydraulic conductivity due to embolism of −2 MPa or less in R. acris (dry; Fig. 2). Thus, despite the fact that plants were still very vulnerable to drought, it is important to stress that the leaf hydraulics method employed measured bulk hydraulic conductivity, including both xylary and extraxylary (e.g. mesophyll) pathways, and reflected the total effect on symplastic and apoplastic transport. Interestingly, stained cross sections also indicated an all-or-nothing effect, where, in most cases, individual vascular bundles lost all conductivity at once (Fig. 2C).
Compared with the staining results, ultrasonic activity was observed at much less negative Ψ than expected (Supplemental Table S2; Supplemental Fig. S1). We believe that other signal sources unrelated to water transport (e.g. parenchyma and sclerenchyma) were responsible for a considerable portion of emissions, skewing the acoustic analysis. Thus, interpretation of UE analyses on herbs is difficult and requires further methodical optimization.
With daily Ψmin being 0.3 to 1.4 MPa lower than Pk50, at least two of our species would incur significant water transport reduction during the course of the day (Tables I and II; Stiller et al., 2003), limiting transpiration and, thus, according to Ohm’s law, preventing a further decrease of Ψ. However, the staining experiment suggests that embolism plays only a minor role in hydraulic decline at the observed in situ Ψ (Fig. 2); therefore, water transport is limited mainly by reversible extraxylary effects (e.g. turgor loss or cell collapse and associated mesophyll conductance losses; Brodribb and Holbrook, 2005; Kim and Steudle, 2007; Blackman et al., 2010; Holloway-Phillips and Brodribb, 2011a; Scoffoni et al., 2014).
Like significant hydraulic transport reductions, the TLP is another critical point plants need to avoid so they can maintain photosynthesis, transport of solutes, and growth (Frensch and Hsiao, 1994; Kramer and Boyer, 1995; Brodribb and Holbrook, 2003; Bartlett et al., 2012). In our study species, the TLP was lower than hydraulic Pk50, indicating that all studied species would face notable reductions in transport capacity under limited water supply before leaves would lose turgor (Tables I and II). This indicates that the impairment of whole-leaf water transport is the first critical event during water stress. Pressure-volume analysis revealed that TLPs in this study were determined mainly by the Π0, whereas a mediating, opposite effect caused by the observed trends in bulk elasticity was small. Only in R. lanuginosus, the high elasticity may have compensated for the species’ weak water-holding capability (high Π0) to maintain turgor longer.
Electrolyte leakage analysis showed that desiccation-induced damage to living cells (P12CL) would only begin long after significant hydraulic decline (Pk50; Table II; Figs. 1 and 3). Since leaves were benchtop dehydrated for up to several hours, we would expect an even higher apparent vulnerability due to continuous exposure to low Ψ in these measurements, compared with shorter in situ stress peaks followed by stomatal regulation and Ψ recovery. Thus, living cells were well protected from desiccation-induced cell damage, as found previously by other authors (Vilagrosa et al., 2010; Beikircher et al., 2013). Interestingly, plants from dry habitats did not appear more resistant to cell damage than plants from humid sites (Table II; Fig. 3). While we sampled only fully developed, mature leaves, we note that R. bulbosus required measurement 1 to 2 months earlier than other species due to their summer dormancy (see “Study Sites and Plant Material”).
Interspecific Differences
Study species with narrow ecological amplitude (R. bulbosus and R. lanuginosus) showed significant differences in some key hydraulic parameters (TLP, Pk50, and Pk88), which were correlated with their respective soil humidity conditions. The corresponding trend in daily Ψmin further indicated that Ψleaf was regulated near Pk88 in both species. In contrast, kleaf max, daily gs max, and P12CL revealed no corresponding trend, which suggests that these parameters are more evolutionarily conserved.
The hydraulics of R. acris, with its broad ecological amplitude, showed characteristics closer to the drought-adapted species (e.g. Pk50 and Pk88; Table II) regarding hydraulic measurements and did not reveal the intermediate values we expected. The fact that Pk50, Pk88, and P12CL were more negative in R. acris than in R. bulbosus may suggest that, although R. acris also occurs frequently in humid sites, it is generally well adapted for dry habitats. Another possible explanation is that, by dying back to perennial tubers in early summer, R. bulbosus effectively avoids periods of intense water stress in summer and autumn (Coles, 1973; Sarukhan, 1974; Larcher, 1995).
Intraspecific Acclimation
Within-species variation in hydraulic parameters was found previously for woody species (Matzner et al., 2001; Cochard et al., 2007; Beikircher and Mayr, 2009; Charra-Vaskou et al., 2012) and herbaceous species (Holste et al., 2006; Tixier et al., 2013; Nolf et al., 2014). R. acris was chosen for its wide distribution and broad ecological amplitude and showed trends for intraspecific variation. While most of the species’ hydraulic traits indicated adjustment to dry habitats when compared with other species (Tables I and II; Figs. 1, 3, and 4), plants from the dry site tended to have more negative vulnerability thresholds (Pk50 and Pk88), TLP, and daily Ψmin than conspecific plants from the humid site. Thus, we suggest that the intraspecific variability in hydraulic parameters of R. acris allows the species to retain fitness over a range of humidity conditions (Rejmanek, 1999) and is a major factor determining its wide distribution, as found for other herbaceous species (Hao et al., 2013; Nolf et al., 2014). Interestingly, the onset of desiccation-induced cellular damage (P12CL; Table II; Fig. 3) was constant across sites, indicating that this parameter was not subject to adjustment.
CONCLUSION
Overall, all of our study species were very vulnerable to water stress, even R. bulbosus, which is a typical species of dry meadows. However, the first effect of drought in these species was found to be a loss of kleaf at moderate Ψ based on extraxylary pathways rather than embolism formation. This may be an effective, easily reversible way for plants to avoid critical decreases in Ψ and subsequent hydraulic failure.
Yet, also in these herbs, differences in vulnerability to water stress were observed on interspecific and intraspecific levels: in narrow-amplitude species, some key hydraulic parameters were correlated to the humidity conditions of their respective habitats (TLP, daily Ψmin, Pk50, and Pk88), while other parameters (kleaf max, gs max, and P12CL) did not reveal such trends. In contrast, the broad-amplitude species showed correlations or trends in intraspecific adjustment to different habitat conditions. Due to the ecological importance of herbaceous species in many ecosystems, better knowledge regarding their hydraulics will be essential to improve evaluations of water stress vulnerability on vegetation and ecosystem levels.
MATERIALS AND METHODS
Study Sites and Plant Material
The study was conducted at three sites in Innsbruck, Austria (47.267°N, 11.393°E), with 896.5 mm of precipitation and an average air temperature of 8.5°C in the 30-year mean (ZAMG Zentralanstalt für Meteorologie und Geodynamik, 2002). Three species of Ranunculus, which occur frequently within the study area, were chosen for analysis due to their differing habitat conditions. Ranunculus bulbosus is a typical species of dry meadows and nutrient-poor grasslands and grows up to 15 to 40 cm tall (Hegi, 1974; Fischer et al., 2008). This species dies back to perennial bulb-like corms in July and remains dormant until the following spring (Coles, 1973; Sarukhan, 1974). Sample plants were taken from a south-exposed, nonintensively managed meadow (Hötting). In contrast, Ranunculus lanuginosus is found in humid, shaded environments such as riparian forests or gorges and grows up to 50 to 70 cm (Hegi, 1974; Fischer et al., 2008). Plants were collected in a small gorge (Sillschlucht) at the south end of the city. The third species, Ranunculus acris, occurs in a range of humidity conditions from dry to humid meadows and reaches intermediate heights of 30 to 60 cm (Hegi, 1974; Polatschek, 2000; Fischer et al., 2008). For this species, a dry population cooccurring with the sampled R. bulbosus population and a humid population occurring on a separate high-humidity site (Völs) were used for analysis.
Soil humidity, species composition, and diurnal variation of gs and Ψleaf were recorded on site in late spring and summer (May to August) in two consecutive years. For all other analyses, whole plants including major roots were collected from the field sites between May and August, immediately placed in dark plastic bags to limit water loss, and transported to the laboratory within 30 min. Plants were rehydrated overnight with their roots in water and covered with a dark plastic bag. Measurements were made on healthy, fully developed leaves including petioles (pressure-volume relations, leaf hydraulic vulnerability, and conductive xylem staining), leaf petioles with intact laminas (acoustic emissions), and leaf discs cut from the leaf laminas (electrolyte leakage).
Soil Humidity
Soil humidity was analyzed in situ in 2 years during dry periods with the Hydrosense II Soil Moisture Measurement System (Campbell Scientific), whereby volumetric SWC was measured 24 h after strong rainfall (SWCfield capacity) and after 7 d without rain (SWCdry) in 10 samples per site and date. The SWCrel (percentage of field capacity) after 7 d without rain was calculated as:
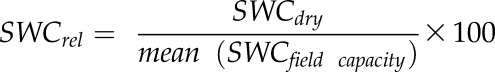 |
(1) |
Additionally, we recorded species composition at the three sites and characterized water availability using averaged ecological indicator values (humidity value; Ellenberg et al., 1992; Schaffers and Sykora, 2000).
Ψ Determination
Ψleaf was measured with a pressure chamber (Model 1000 Pressure Chamber; PMS Instrument) by applying pressure to an excised leaf sealed in the chamber, with the petiole protruding out through a rubber gasket (Scholander et al., 1965). Air pressure inside the chamber was increased slowly until the xylem sap meniscus became visible at the cutting site.
Pressure-Volume Analysis
Pressure-volume analyses (Tyree and Hammel, 1972; Sack and Pasquet-Kok, 2011; Bartlett et al., 2012) were carried out on five leaves per species/population to determine TLP, Π0 at full turgor, and modulus of elasticity. Leaves were fully saturated before dehydration and regular measurements of Ψleaf and fresh weight. Relative water content was calculated from saturated, fresh, and dry weights and plotted against the inverse of Ψleaf to find the TLP (transition point between the linear and curved portions of the graph), Π0 (negative inverse of the linear graph portion’s intercept with the y axis), and modulus of elasticity (slope of the curve above and including the TLP). Parameters were calculated individually per leaf and then averaged per species/population.
Leaf Hydraulic Vulnerability
Declines in kleaf due to increasing desiccation were analyzed with a modified non-steady-state leaf rehydration technique (Brodribb and Cochard, 2009; Blackman and Brodribb, 2011) and benchtop dehydration. For 39 to 45 leaves per species/population, we measured the decay of initial water uptake into the leaf from a small water container placed on a digital balance (CP64; Sartorius), which was connected to the leaf petiole via hydraulic tubing (see below).
At increasing levels of dehydration, plants were equilibrated in dark plastic bags and plant Ψ was determined on two leaves per plant. If Ψ between both leaves differed by 10% or less, the decay of initial flow was measured by excising a third leaf under water (including approximately 3 cm of the petiole), immediately connecting the leaf to hydraulic tubing, and covering the leaf with a moist paper towel to prevent transpiration. Thus, hydraulic flow was measured on non-light-acclimated, nontranspiring leaves. Flow rates (kleaf [mmol m−2 s−1 MPa−1]) were determined by linear regression of decreasing mass readings (water uptake) during the first 10 s and adjusted for temperature and leaf area. kleaf max was calculated as the average of five maximum conductances observed per species/population. No cutting artifact (Wheeler et al., 2013) was expected for leaves excised under water (Scoffoni and Sack, 2015).
The reparameterized Weibull function (Ogle et al., 2009) was fitted to plots of relative kleaf versus Ψ using the fitplc package for R version 3.1.1 (Duursma, 2014; R Core Team, 2014):
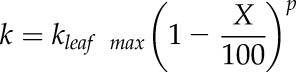 |
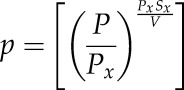 |
(2) |
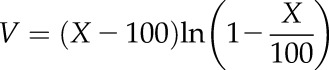 |
where k is the expected hydraulic conductance at X% loss of conductance, kleaf max is the k at full saturation (i.e. k at 0 MPa), P is the positive-valued Ψ (P = −Ψ), Px is the Ψ at X% loss of conductance, and Sx is the slope of the vulnerability curve at P = Px. Pk50 and Pk88 were defined as above.
Staining of Conducting Xylem Elements
To confirm the observed high levels of leaf hydraulic vulnerability, we stained conducting xylem elements in leaves of R. acris from the dry site. At increasing levels of dehydration, whole plants were equilibrated in dark plastic bags and plant Ψ was determined on one leaf per plant. Two additional leaves of the same plant were excised under water and immediately placed in a beaker containing (1) 1% (w/v) phloxine B (modified after Nardini et al., 2003) to reveal ongoing water transport in the lamina and (2) 0.05% (w/v) safranin (modified after Sperry et al., 1988b) to stain water-conducting conduits in the petioles. Samples were exposed to sunlight for about 10 min. Transpiration was favored by partially removing the cuticle and confirmed with an SC-1 Leaf Porometer (Decagon Devices). We analyzed images of the lamina taken with a digital camera and petiole cross sections (approximately 3 cm from the lamina) taken with a light microscope (Olympus BX41; Olympus Austria) and interfaced digital camera (ProgRes CT3; Jenoptik). To prevent artifacts by passive dye diffusion, only samples with unstained parenchyma and phloem were used.
Electrolyte Leakage
The vulnerability of living tissues to dehydration (Morin et al., 2007; Hu et al., 2010; Vilagrosa et al., 2010; Beikircher et al., 2013) was analyzed by benchtop dehydrating plants and comparing the leakage of electrolytes from 52 to 82 leaves per species at increasing levels of dehydration. After Ψ determination, three to 10 circular discs (0.6 cm in diameter) were cut from each leaf by use of a cork borer while avoiding major leaf veins. Leaf discs were pooled for each sample leaf, submerged in 15 mL of distilled water in individual centrifuge tubes, shaken for 24 h on a horizontal shaker (ST5 Bidimensional Shaker; CAT) at 5°C, followed by electrolytic conductivity measurement at room temperature (after 1 h of temperature equilibration) using a four-electrode conductivity sensor (Tetracon 325; WTW). Samples were then autoclaved at 120°C for 30 min and shaken overnight at room temperature, before final electrolytic conductivity was measured.
Relative electrolyte leakage (REL) was calculated as:
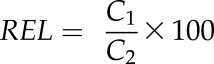 |
(3) |
where C1 and C2 are the initial and final electrolytic conductivities, respectively. REL was then standardized to the percentage of cellular lysis by relating each sample’s REL to the REL of the control (RELc; Morin et al., 2007); in this case, samples were measured at Ψ of −0.2 MPa or greater:
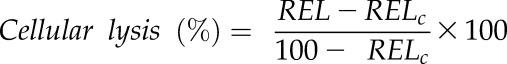 |
(4) |
Vulnerability thresholds were again calculated by curve fitting to Equation 2, where k is the percentage of intact cells (k = 100 − the percentage of cellular lysis). The onset of damage to living cells (P12CL) was defined as above.
Diurnal Variation of gs and Ψleaf
We monitored the diurnal progression of gs (using an SC-1 Leaf Porometer; Decagon Devices) and corresponding Ψleaf between sunrise and sunset on one clear, sunny day per species following a period of rainfall. For each data point, gs was measured on the lower leaf side of four to five undamaged, fully developed leaves of different plants, whereas Ψleaf was determined on three undamaged, fully developed leaves of different plants.
Statistical Analyses
Data for curve fitting were pooled per species/population and method before further analysis. Vulnerability thresholds were statistically tested for differences using bootstrapped confidence intervals (n = 999 bootstrap estimates). Other data were normally distributed (QQ plots and Shapiro-Wilk test) and tested for differences using Student’s t test (homoscedastic data as determined by the robust Brown-Forsythe Levene-type test) or Welch’s two-sample t test (heteroscedastic data). All analyses were done in R version 3.1.1 (R Core Team, 2014) at a significance level of P < 0.05, whereas P was Bonferroni corrected for multiple comparisons. Values are presented as means ± se or mean (lower confidence interval, upper confidence interval).
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. Relative cumulative UE versus Ψ in R. bulbosus, R. lanuginosus, and dry and humid R. acris populations.
Supplemental Figure S2. Diurnal variation of Ψ and gs in R. bulbosus and R. lanuginosus.
Supplemental Table S1. Species composition and ecological indicator values.
Supplemental Table S2. Water potential at 50 and 88% of total cumulative ultrasonic emissions above -3 MPa.
Supplementary Material
Acknowledgments
We thank Dr. Georg Wohlfahrt for providing valuable microclimate data and Birgit Dämon for excellent technical assistance.
Glossary
- Ψ
water potential
- UE
ultrasonic acoustic emissions
- SWC
soil water content
- SWCrel
relative soil water content
- TLP
turgor loss point
- Ψleaf
leaf water potential
- Π0
osmotic potential
- kleaf max
maximum whole-leaf hydraulic conductance
- Pk50
water potential at 50% loss of leaf hydraulic conductance
- Pk88
water potential at 88% loss of leaf hydraulic conductance
- P12CL
water potential at 12% of cellular lysis
- Ψmin
minimum leaf water potential
- gs
stomatal conductance
- gs max
maximum stomatal conductance
- kleaf
whole-leaf hydraulic conductance
Footnotes
This work was supported by the Austrian Science Fund (FWF project nos. I826–B25 and T–667) and the Austrian Academy of Sciences (DOC fellowship to M.N.).
Articles can be viewed without a subscription.
References
- Bartlett MK, Scoffoni C, Sack L (2012) The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: a global meta-analysis. Ecol Lett 15: 393–405 [DOI] [PubMed] [Google Scholar]
- Beikircher B, De Cesare C, Mayr S (2013) Hydraulics of high-yield orchard trees: a case study of three Malus domestica cultivars. Tree Physiol 33: 1296–1307 [DOI] [PubMed] [Google Scholar]
- Beikircher B, Mayr S (2009) Intraspecific differences in drought tolerance and acclimation in hydraulics of Ligustrum vulgare and Viburnum lantana. Tree Physiol 29: 765–775 [DOI] [PubMed] [Google Scholar]
- Billings WD, Mooney HA (1968) Ecology of arctic and alpine plants. Biol Rev Camb Philos Soc 43: 481–529 [Google Scholar]
- Blackman CJ, Brodribb TJ (2011) Two measures of leaf capacitance: insights into the water transport pathway and hydraulic conductance in leaves. Funct Plant Biol 38: 118–126 [DOI] [PubMed] [Google Scholar]
- Blackman CJ, Brodribb TJ, Jordan GJ (2010) Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms. New Phytol 188: 1113–1123 [DOI] [PubMed] [Google Scholar]
- Blackman CJ, Brodribb TJ, Jordan GJ (2012) Leaf hydraulic vulnerability influences species’ bioclimatic limits in a diverse group of woody angiosperms. Oecologia 168: 1–10 [DOI] [PubMed] [Google Scholar]
- Brodersen CR, McElrone AJ, Choat B, Lee EF, Shackel KA, Matthews MA (2013) In vivo visualizations of drought-induced embolism spread in Vitis vinifera. Plant Physiol 161: 1820–1829 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodribb TJ, Cochard H (2009) Hydraulic failure defines the recovery and point of death in water-stressed conifers. Plant Physiol 149: 575–584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodribb TJ, Holbrook NM (2003) Stomatal closure during leaf dehydration, correlation with other leaf physiological traits. Plant Physiol 132: 2166–2173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodribb TJ, Holbrook NM (2004) Stomatal protection against hydraulic failure: a comparison of coexisting ferns and angiosperms. New Phytol 162: 663–670 [DOI] [PubMed] [Google Scholar]
- Brodribb TJ, Holbrook NM (2005) Water stress deforms tracheids peripheral to the leaf vein of a tropical conifer. Plant Physiol 137: 1139–1146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bucci SJ, Scholz FG, Peschiutta ML, Arias NS, Meinzer FC, Goldstein G (2013) The stem xylem of Patagonian shrubs operates far from the point of catastrophic dysfunction and is additionally protected from drought-induced embolism by leaves and roots. Plant Cell Environ 36: 2163–2174 [DOI] [PubMed] [Google Scholar]
- Bunce JA. (2006) How do leaf hydraulics limit stomatal conductance at high water vapour pressure deficits? Plant Cell Environ 29: 1644–1650 [DOI] [PubMed] [Google Scholar]
- Charra-Vaskou K, Charrier G, Wortemann R, Beikircher B, Cochard H, Ameglio T, Mayr S (2012) Drought and frost resistance of trees: a comparison of four species at different sites and altitudes. Ann For Sci 69: 325–333 [Google Scholar]
- Chen JW, Zhang Q, Li XS, Cao KF (2009) Independence of stem and leaf hydraulic traits in six Euphorbiaceae tree species with contrasting leaf phenology. Planta 230: 459–468 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choat B, Drayton WM, Brodersen C, Matthews MA, Shackel KA, Wada H, McElrone AJ (2010) Measurement of vulnerability to water stress-induced cavitation in grapevine: a comparison of four techniques applied to a long-vesseled species. Plant Cell Environ 33: 1502–1512 [DOI] [PubMed] [Google Scholar]
- Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Feild TS, Gleason SM, Hacke UG, et al. (2012) Global convergence in the vulnerability of forests to drought. Nature 491: 752–755 [DOI] [PubMed] [Google Scholar]
- Cochard H. (2002) A technique for measuring xylem hydraulic conductance under high negative pressures. Plant Cell Environ 25: 815–819 [Google Scholar]
- Cochard H, Badel E, Herbette S, Delzon S, Choat B, Jansen S (2013) Methods for measuring plant vulnerability to cavitation: a critical review. J Exp Bot 64: 4779–4791 [DOI] [PubMed] [Google Scholar]
- Cochard H, Casella E, Mencuccini M (2007) Xylem vulnerability to cavitation varies among poplar and willow clones and correlates with yield. Tree Physiol 27: 1761–1767 [DOI] [PubMed] [Google Scholar]
- Cochard H, Delzon S, Badel E (2015) X-ray microtomography (micro-CT): a reference technology for high-resolution quantification of xylem embolism in trees. Plant Cell Environ 38: 201–206 [DOI] [PubMed] [Google Scholar]
- Coles S. (1973) Ranunculus bulbosus L in Europe. Watsonia 9: 207–228 [Google Scholar]
- Duursma R (2014) Fit vulnerability curves in R (R package). Bitbucket. https://bitbucket.org/remkoduursma/fitplc/ (January 10, 2015)
- Ellenberg H, Weber EW, Wirth V, Werner W, Paulißen D (1992) Zeigerwerte von Pflanzen in Mitteleuropa. Scripta Geobotanica 18: 1–258 [Google Scholar]
- Engelbrecht BMJ, Comita LS, Condit R, Kursar TA, Tyree MT, Turner BL, Hubbell SP (2007) Drought sensitivity shapes species distribution patterns in tropical forests. Nature 447: 80–82 [DOI] [PubMed] [Google Scholar]
- Fischer M, Oswald K, Adler W (2008) Exkursionsflora für Österreich, Liechtenstein und Südtirol, Ed 3 OÖ Landesmuseen, Land Öberösterreich, Linz, Austria [Google Scholar]
- Frensch J, Hsiao TC (1994) Transient responses of cell turgor and growth of maize roots as affected by changes in water potential. Plant Physiol 104: 247–254 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galmes J, Flexas J, Save R, Medrano H (2007) Water relations and stomatal characteristics of Mediterranean plants with different growth forms and leaf habits: responses to water stress and recovery. Plant Soil 290: 139–155 [Google Scholar]
- Ganthaler A, Mayr S (2015) Dwarf shrub hydraulics: two Vaccinium species (V. myrtillus, V. vitis‐idaea) of the European Alps compared. Physiol Plant 155: 424–434 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilliam FS. (2007) The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience 57: 845–858 [Google Scholar]
- Gleason SM, Westoby M, Jansen S, Choat B, Hacke UG, Pratt RB, Bhaskar R, Brodribb TJ, Bucci SJ, Cao KF, et al. (2016) Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species. New Phytol 209: 123–136 [DOI] [PubMed] [Google Scholar]
- Guyot G, Scoffoni C, Sack L (2012) Combined impacts of irradiance and dehydration on leaf hydraulic conductance: insights into vulnerability and stomatal control. Plant Cell Environ 35: 857–871 [DOI] [PubMed] [Google Scholar]
- Hao GY, Hoffmann WA, Scholz FG, Bucci SJ, Meinzer FC, Franco AC, Cao KF, Goldstein G (2008) Stem and leaf hydraulics of congeneric tree species from adjacent tropical savanna and forest ecosystems. Oecologia 155: 405–415 [DOI] [PubMed] [Google Scholar]
- Hao GY, Lucero ME, Sanderson SC, Zacharias EH, Holbrook NM (2013) Polyploidy enhances the occupation of heterogeneous environments through hydraulic related trade-offs in Atriplex canescens (Chenopodiaceae). New Phytol 197: 970–978 [DOI] [PubMed] [Google Scholar]
- Hegi G (1974) Illustrierte Flora von Mitteleuropa, Band III/3, Ed 2. Carl Hanser Verlag, Munich, Germany [Google Scholar]
- Holloway-Phillips MM, Brodribb TJ (2011a) Minimum hydraulic safety leads to maximum water-use efficiency in a forage grass. Plant Cell Environ 34: 302–313 [DOI] [PubMed] [Google Scholar]
- Holloway-Phillips MM, Brodribb TJ (2011b) Contrasting hydraulic regulation in closely related forage grasses: implications for plant water use. Funct Plant Biol 38: 594–605 [DOI] [PubMed] [Google Scholar]
- Holste EK, Jerke MJ, Matzner SL (2006) Long-term acclimatization of hydraulic properties, xylem conduit size, wall strength and cavitation resistance in Phaseolus vulgaris in response to different environmental effects. Plant Cell Environ 29: 836–843 [DOI] [PubMed] [Google Scholar]
- Hu L, Wang Z, Du H, Huang B (2010) Differential accumulation of dehydrins in response to water stress for hybrid and common bermudagrass genotypes differing in drought tolerance. J Plant Physiol 167: 103–109 [DOI] [PubMed] [Google Scholar]
- Iovi K, Kolovou C, Kyparissis A (2009) An ecophysiological approach of hydraulic performance for nine Mediterranean species. Tree Physiol 29: 889–900 [DOI] [PubMed] [Google Scholar]
- Johnson DM, McCulloh KA, Woodruff DR, Meinzer FC (2012) Evidence for xylem embolism as a primary factor in dehydration-induced declines in leaf hydraulic conductance. Plant Cell Environ 35: 760–769 [DOI] [PubMed] [Google Scholar]
- Kikuta SB. (2003) Ultrasound acoustic emissions from bark samples differing in anatomical characteristics. Phyton 43: 161–178 [Google Scholar]
- Kim YX, Steudle E (2007) Light and turgor affect the water permeability (aquaporins) of parenchyma cells in the midrib of leaves of Zea mays. J Exp Bot 58: 4119–4129 [DOI] [PubMed] [Google Scholar]
- Kocacinar F, Sage RF (2003) Photosynthetic pathway alters xylem structure and hydraulic function in herbaceous plants. Plant Cell Environ 26: 2015–2026 [DOI] [PubMed] [Google Scholar]
- Kramer PJ, Boyer JS (1995) Water Relations of Plants and Soils. Academic Press, San Diego [Google Scholar]
- Larcher W. (1995) Physiological Plant Ecology. Ed 3 Springer, Berlin [Google Scholar]
- Maherali H, Pockman WT, Jackson RB (2004) Adaptive variation in the vulnerability of woody plants to xylem cavitation. Ecology 85: 2184–2199 [Google Scholar]
- Matzner SL, Rice KJ, Richards JH (2001) Intra-specific variation in xylem cavitation in interior live oak (Quercus wislizenii A. DC.). J Exp Bot 52: 783–789 [DOI] [PubMed] [Google Scholar]
- Mayr S, Rosner S (2011) Cavitation in dehydrating xylem of Picea abies: energy properties of ultrasonic emissions reflect tracheid dimensions. Tree Physiol 31: 59–67 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mencuccini M. (2003) The ecological significance of long‐distance water transport: short‐term regulation, long‐term acclimation and the hydraulic costs of stature across plant life forms. Plant Cell Environ 26: 163–182 [Google Scholar]
- Milburn JA, Johnson RPC (1966) The conduction of sap. II. Detection of vibrations produced by sap cavitation in Ricinus xylem. Planta 69: 43–52 [DOI] [PubMed] [Google Scholar]
- Monfreda C, Ramankutty N, Foley JA (2008) Farming the planet. 2. Geographic distribution of crop areas, yields, physiological types, and net primary production in the year 2000. Global Biogeochem Cycles 22: Gb1022 [Google Scholar]
- Morin X, Améglio T, Ahas R, Kurz-Besson C, Lanta V, Lebourgeois F, Miglietta F, Chuine I (2007) Variation in cold hardiness and carbohydrate concentration from dormancy induction to bud burst among provenances of three European oak species. Tree Physiol 27: 817–825 [DOI] [PubMed] [Google Scholar]
- Nardini A, Pedà G, La Rocca N (2012) Trade-offs between leaf hydraulic capacity and drought vulnerability: morpho-anatomical bases, carbon costs and ecological consequences. New Phytol 196: 788–798 [DOI] [PubMed] [Google Scholar]
- Nardini A, Salleo S, Raimondo F (2003) Changes in leaf hydraulic conductance correlate with leaf vein embolism in Cercis siliquastrum L. Trees (Berl) 17: 529–534 [Google Scholar]
- Nolf M, Creek D, Duursma R, Holtum J, Mayr S, Choat B (2015) Stem and leaf hydraulic properties are finely coordinated in three tropical rain forest tree species. Plant Cell Environ 38: 2652–2661 [DOI] [PubMed] [Google Scholar]
- Nolf M, Pagitz K, Mayr S (2014) Physiological acclimation to drought stress in Solidago canadensis. Physiol Plant 150: 529–539 [DOI] [PubMed] [Google Scholar]
- Ogle K, Barber JJ, Willson C, Thompson B (2009) Hierarchical statistical modeling of xylem vulnerability to cavitation. New Phytol 182: 541–554 [DOI] [PubMed] [Google Scholar]
- Polatschek A. (2000) Flora von Nordtirol, Osttirol und Vorarlberg: Bd 3. Samenpflanzen: Fabaceae bis Rosaceae. Kartenteil. Tiroler Landesmuseum Ferdinandeum, Innsbruck, Austria [Google Scholar]
- Ponomarenko A, Vincent O, Pietriga A, Cochard H, Badel É, Marmottant P (2014) Ultrasonic emissions reveal individual cavitation bubbles in water-stressed wood. J R Soc Interface 11: 20140480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- R Core Team (2014) R: A Language and Environment for Statistical Computing. http://www.R-project.org/
- Rejmanek M. (1999) Invasive plant species and invasible ecosystems. In Sandlund OT, Schei PJ, Viken A, eds, Invasive Species and Biodiversity Management. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp 79–102 [Google Scholar]
- Rosenthal DM, Stiller V, Sperry JS, Donovan LA (2010) Contrasting drought tolerance strategies in two desert annuals of hybrid origin. J Exp Bot 61: 2769–2778 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosner S. (2015) A new type of vulnerability curve: is there truth in vine? Tree Physiol 35: 410–414 [DOI] [PubMed] [Google Scholar]
- Sack L, Holbrook NM (2006) Leaf hydraulics. Annu Rev Plant Biol 57: 361–381 [DOI] [PubMed] [Google Scholar]
- Sack L, Pasquet-Kok J (2011) Leaf pressure-volume curve parameters. PrometheusWiki. http://www.publish.csiro.au/prometheuswiki/tiki-pagehistory.php?page=Leaf%20pressure-volume%20curve%20parameters&preview=16 (January 22, 2015)
- Sandford AP, Grace J (1985) The measurement and interpretation of ultrasound from woody stems. J Exp Bot 36: 298–311 [Google Scholar]
- Sarukhan J. (1974) Studies on plant demography: Ranunculus repens L., R. bulbosus L. and R. acris L. II. Reproductive strategies and seed population dynamics. J Ecol 62: 151–177 [Google Scholar]
- Schaffers AP, Sykora KV (2000) Reliability of Ellenberg indicator values for moisture, nitrogen and soil reaction: a comparison with field measurements. J Veg Sci 11: 225–244 [Google Scholar]
- Scholander PF, Bradstreet ED, Hemmingsen EA, Hammel HT (1965) Sap pressure in vascular plants: negative hydrostatic pressure can be measured in plants. Science 148: 339–346 [DOI] [PubMed] [Google Scholar]
- Scholz FG, Bucci SJ, Hoffmann WA, Meinzer FC, Goldstein G (2010) Hydraulic lift in a neotropical savanna: experimental manipulation and model simulations. Agric For Meteorol 150: 629–639 [Google Scholar]
- Scoffoni C, Sack L (2015) Are leaves ‘freewheelin’? Testing for a Wheeler-type effect in leaf xylem hydraulic decline. Plant Cell Environ 38: 534–543 [DOI] [PubMed] [Google Scholar]
- Scoffoni C, Vuong C, Diep S, Cochard H, Sack L (2014) Leaf shrinkage with dehydration: coordination with hydraulic vulnerability and drought tolerance. Plant Physiol 164: 1772–1788 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sperry JS, Donnelly JR, Tyree MT (1988a) A method for measuring hydraulic conductivity and embolism in xylem. Plant Cell Environ 11: 35–40 [Google Scholar]
- Sperry JS, Donnelly JR, Tyree MT (1988b) Seasonal occurrence of xylem embolism in sugar maple (Acer saccharum). Am J Bot 75: 1212–1218 [Google Scholar]
- Sperry JS, Holbrook NM, Zimmermann MH, Tyree MT (1987) Spring filling of xylem vessels in wild grapevine. Plant Physiol 83: 414–417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiller V, Lafitte HR, Sperry JS (2003) Hydraulic properties of rice and the response of gas exchange to water stress. Plant Physiol 132: 1698–1706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiller V, Sperry JS (2002) Cavitation fatigue and its reversal in sunflower (Helianthus annuus L.). J Exp Bot 53: 1155–1161 [DOI] [PubMed] [Google Scholar]
- Tixier A, Cochard H, Badel E, Dusotoit-Coucaud A, Jansen S, Herbette S (2013) Arabidopsis thaliana as a model species for xylem hydraulics: does size matter? J Exp Bot 64: 2295–2305 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tyree MT, Davis SD, Cochard H (1994) Biophysical perspectives of xylem evolution: is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction. IAWA J 15: 335–360 [Google Scholar]
- Tyree MT, Ewers FW (1991) The hydraulic architecture of trees and other woody plants. New Phytol 119: 345–360 [Google Scholar]
- Tyree MT, Fiscus EL, Wullschleger SD, Dixon MA (1986) Detection of xylem cavitation in corn under field conditions. Plant Physiol 82: 597–599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tyree MT, Hammel HT (1972) Measurement of turgor pressure and water relations of plants by pressure-bomb technique. J Exp Bot 23: 267–282 [Google Scholar]
- Tyree MT, Sperry JS (1989) Vulnerability of xylem to cavitation and embolism. Annu Rev Plant Physiol 40: 19–38 [Google Scholar]
- Tyree MT, Zimmermann MH (2002) Xylem Structure and the Ascent of Sap, Ed 2 Springer, Berlin [Google Scholar]
- Vilagrosa A, Morales F, Abadia A, Bellot J, Cochard H, Gil-Pelegrin E (2010) Are symplast tolerance to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species. Environ Exp Bot 69: 233–242 [Google Scholar]
- Wagner KR, Ewers FW, Davis SD (1998) Tradeoffs between hydraulic efficiency and mechanical strength in the stems of four co-occurring species of chaparral shrubs. Oecologia 117: 53–62 [DOI] [PubMed] [Google Scholar]
- Wheeler JK, Huggett BA, Tofte AN, Rockwell FE, Holbrook NM (2013) Cutting xylem under tension or supersaturated with gas can generate PLC and the appearance of rapid recovery from embolism. Plant Cell Environ 36: 1938–1949 [DOI] [PubMed] [Google Scholar]
- Wohlfahrt G, Irschick C, Thalinger B, Hörtnagl L, Obojes N, Hammerle A (2010) Insights from independent evapotranspiration estimates for closing the energy balance: a grassland case study. Vadose Zone J 9: 1025–1033 [PMC free article] [PubMed] [Google Scholar]
- Wolkerstorfer SV, Rosner S, Hietz P (2012) An improved method and data analysis for ultrasound acoustic emissions and xylem vulnerability in conifer wood. Physiol Plant 146: 184–191 [DOI] [PubMed] [Google Scholar]
- ZAMG Zentralanstalt für Meteorologie und Geodynamik (2002) Klimadaten von Österreich 1971-2000. http://www.zamg.ac.at/fix/klima/oe71-00/klima2000/klimadaten_oesterreich_1971 _ frame1.htm (January 22, 2015)
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.