

Abstract
Key players in translational regulation such as ribosomes might represent powerful, but hitherto largely unexplored, targets to eliminate drug-refractory cancer stem cells (CSCs). A recent study by the Lisanti group has documented how puromycin, an old antibiotic derived from Streptomyces alboniger that inhibits ribosomal protein translation, can efficiently suppress CSC states in tumorspheres and monolayer cultures. We have used a closely related approach based on Biolog Phenotype Microarrays (PM), which contain tens of lyophilized antimicrobial drugs, to assess the chemosensitivity profiles of breast cancer cell lines enriched for stem cell-like properties. Antibiotics directly targeting active sites of the ribosome including emetine, puromycin and cycloheximide, inhibitors of ribosome biogenesis such as dactinomycin, ribotoxic stress agents such as daunorubicin, and indirect inhibitors of protein synthesis such as acriflavine, had the largest cytotoxic impact against claudin-low and basal-like breast cancer cells. Thus, biologically aggressive, treatment-resistant breast cancer subtypes enriched for stem cell-like properties exhibit exacerbated chemosensitivities to anti-protozoal and anti-bacterial antibiotics targeting protein synthesis. These results suggest that old/existing microbicides might be repurposed not only as new cancer therapeutics, but also might provide the tools and molecular understanding needed to develop second-generation inhibitors of ribosomal translation to eradicate CSC traits in tumor tissues.
Keywords: Antibiotics, breast cancer, basal-like, Biolog, cancer stem cells, claudin-low, drug repositioning, drug repurposing, phenotype, ribosomes
Recent work from Lisanti's group has rekindled the interest of cancer researchers in using “old” antibiotics inhibiting protein synthesis as effective drugs to eliminate cancer stem cells (CSCs).1 CSCs are implicated in disease recurrence and metastatic spread and are known to be resistant to many conventional therapies.2-7 In their hands, breast CSCs were significantly enriched for numerous ribosomal proteins and appeared to be addicted to protein synthesis. Additionally, they found that antibiotics such as puromycin, which competitively inhibits protein synthesis by mimicking the 3′ end of an aminoacylated tRNA interacting with the A-site of the ribosome and also by generating puromycylated-peptides and causing premature release of the peptide chain, completely prevented CSC-formed mammospheres in a manner strictly dependent on the blockade of nascent protein synthesis by ribosomes.1 Interestingly, puromycin preferentially targeted CSC cellular states because pre-incubation with the antibiotic in the setting of an attached monolayer fully prevented the later capacity of breast cancer cell populations to form microtumors in non-adherent non-differentiating conditions.1
Global gene expression analyses have allowed the delineation of 5 different intrinsic subtypes of breast cancer: Luminal A, Luminal B, HER2-enriched, basal-like, and the recently characterized claudin-low group.8–11 The most aggressive subtypes, claudin-low and basal-like, have the worst mortality rate among the intrinsic breast cancer subtypes due to higher grade at diagnosis, predilection for early metastasis and, importantly, lack of targeted therapy. Basal-like and claudin-low tumors are distinct from other breast cancer subtypes since they possess stem cell-like properties with high expression of mesenchymal and epithelial-to-mesenchymal (EMT) genes, which is thought to be reflective of a least-differentiated stage of epithelial development. Indeed, we are amassing evidence that the claudin-low breast cancer subtype should be viewed as the first example of adult carcinoma driven by aberrant reactivation of an embryonic-like stem cell transdifferentiation program.12 Basal-like and claudin-low characteristics, which are also common in chemotherapy-resistant breast tumors,13 might represent different extents of reprogramming due to the aberrant activation of EMT inducers in committed cells (e.g., luminal progenitors).
An intimate relationship exists between EMT, CSCs, and the claudin-low and basal-like subtypes of breast cancer, representing a cell state with aggressive and therapeutic resistant properties. Given that these breast carcinoma subtypes prominently exhibit enrichment and hyperactivation of oncogenic signaling cascades driven by c-MYC and mTOR,14–17 2 key regulators of the translational machinery and protein synthesis, respectively, we recently hypothesized that pharmacological inhibition of deregulated protein synthesis might constitute a highly conserved target in multi-drug resistant claudin-low and basal-like tumor cells. Moreover, because drug repositioning (also termed re-profiling, therapeutic switching or drug repurposing) constitutes a useful strategy to accelerate the drug development process, we envisioned that one could combine both ideas by simultaneously exploring the cytotoxic activity of “old” antibiotics, antimalarials, antiprotozoals or anticancer drugs against a small panel of selected breast cancer cell lines highly representative of human claudin-low and basal-like tumors.
The claudin-low MDA-MB-231 and SUM-159PT10,11 cell lines and the basal-like HER2+ JIMT-118-24 cell line were used in a chemical sensitivity screen. We utilized the Phenotype Microarray (PM) system, marketed and sold by Biolog (www.biolog.com), to measure the sensitivity of these cells to a wide variety of 92 antibiotics and other growth inhibitors in microplates (PM-M11 to PM-M14). This approach enables the testing of tens of phenotypes and the identification of shared sensitivities among claudin-low and basal-like breast cancer cells to a wide variety of drugs (Fig. 1). We arbitrarily grouped chemosensitivity responses of the breast cancer cell lines into 5 categories based on both the number of drug doses able to induce a growth inhibition rate greater than 50% and the number of drug-responsive cell lines (Fig. 2). Based on these criteria, 5 drugs showed the largest cytotoxic impact on the growth of claudin-low and basal-like cell lines: 1) Emetine, an anti-protozoal alkaloid used in the treatment of ameobiasis that also displays potent anti-malarial activity, is known to interact with the E-site of the ribosomal small subunit and inhibit ribosome movement along mRNA,25 thus irreversibly blocking protein synthesis. Two) Dactinomycin (also known generically as actinomycin D), the first polypeptide antibiotic shown to have anti-cancer activity, inhibits all RNA synthesis including ribosome biogenesis, which is reflected in an inhibition of protein synthesis. Three) Puromycin interacts with the A-site of the ribosome and inhibits protein synthesis via premature chain termination. Four) Daunorubicin (daunomycin), a chemotherapeutic of the anthracycline family, is a ribotoxic stress agent that inhibits protein translation.26 Five) Acriflavine, an intercalating antimetabolite used as a topical antiseptic, has been shown to rapidly inhibit protein synthesis to exert emetine-like amebicidal actions.
Figure 1.

Rapid screening for anti-breast cancer antibiotics using Biolog Phenotype Microarrays. Representative micrographs of Phenotype Microarray analyses measuring tetrazolium dye reduction by MDA-MB-231, SUM-159PT, and JIMT-1 breast carcinoma cells cultured in the absence or presence of 92 antibiotics and other growth inhibitors. Red boxes denote highly active drugs in which at least 3 of the 4 graded concentrations decreased optical density by ≥50 % in each individual cell line. Drug names labeled in red denote those drugs exhibiting high activity in all 3 breast cancer cell lines.
Figure 2.
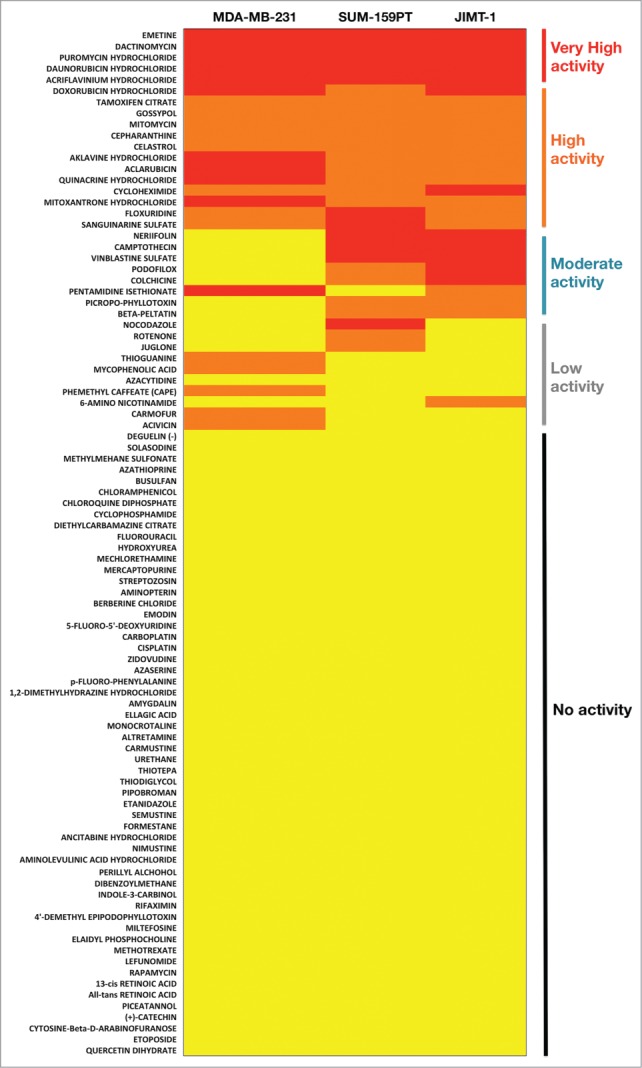
Chemosensitivity map of cell line models representative of breast cancer subtypes enriched for stem cell-like properties. A color-coding scheme assigned each drug to yellow (none of the 4 graded drug concentrations decreased cell growth by ≥50 %), orange (at least one of the 4 graded concentrations decreased cell growth by ≥50 %), or red (at least 3 of the 4 graded concentrations decreased cell growth by ≥50 %) categories. Drugs were categorized as “very high active” if the red code occurred in all 3 cell lines, “high activity” if either the red or orange codes occurred in all 3 cell lines, “moderate activity” if either the red or orange codes occurred in 2 of the 3 cell lines, “low activity” if either the red or orange codes occurred in one of the 3 cell lines, and “no activity” when none of the color-coding categories occurred in the 3 cell lines.
It is noteworthy that an inhibitory capacity on protein synthesis was shared by all the most effective antimicrobials that efficiently inhibited growth of claudin-low and basal-like breast carcinoma cells. Other well-known antibiotics capable of effectively inhibiting protein synthesis such as cycloheximide, an elongation inhibitor that binds to the E site of ribosomes, were among the drugs with a high activity profile against claudin-low and basal-like breast carcinomas (Fig. 2). Although cancer cell lines representative of other breast cancer subtypes (e.g.,, Luminal A/B, HER2-enriched) were not included in the sensitivity screen, it is notable that when the same Biolog Phenotype Microarray was used to profile HER2-enriched SKBR3 cells, reagents such as berberine chloride, azathioprine, celastrol, gossypol, miltefosine, and etoposide, were found to have the largest growth impact on the cell line.27 It is therefore reasonable to assume that an intrinsic hypersensitivity to antimicrobial inhibitors of protein synthesis might occur in biologically aggressive and chemoresistant claudin-low and basal-like breast cancer cells.
Cellular translation is receiving increasing attention in anti-cancer therapy, with key translation regulatory factors such as ribosomes emerging as feasible targets.28,29 In this scenario, drug repurposing can circumvent the high cost of drug discovery and development, the high failure rates and the long duration to develop novel treatments.30,31 Using Phenotype MicroArrays, which enables quantitative measurements of thousands of cellular phenotypes at the same time, we show that breast cancer subtypes enriched for stem cell-like properties display exacerbated chemical sensitivities to microbicides that target ribosomes and/or inhibit protein synthesis (Fig. 3). These findings suggest that classical anti-protozoal and anti-bacterial antibiotics, such as emetine, puromycin or acriflavine, could be used in combination with the currently used conventional cytotoxic chemotherapy in the treatment of claudin-low and basal-like breast carcinomas. Moreover, our ever-growing mechanistic understanding of ribosome functioning might guide the design of future small molecule inhibitors to target eukaryotic translation, which might be closely involved with the acquisition of molecular and functional traits of CSCs. As exemplified for the anti-protozoal alkaloid emetine, now repurposed as an anti-malarial based on the elucidation of how emetine binds to the ribosome,25 and which has already been shown to selectively inhibit glioblastoma stem cells-enriched cultures,32 it might be possible for drug manufacturers to chemically modify natural antibiotics to further enhance their potency and specificity as translation inhibitors capable of eradicating CSC traits in tumor tissues.
Figure 3.

Inhibitors of ribosomal protein synthesis: (A)novel strategy to eradicate CSC traits in tumor tissues. Biologically aggressive, treatment-resistant breast cancer subtypes enriched for stem cell-like properties exhibit exacerbated chemosensitivities to antibiotics directly targeting active sites of the ribosome, such as emetine, puromycin and cycloheximide; inhibitors of ribosome biogenesis such as dactinomycin; ribotoxic stress agents such as daunorubicin, and indirect inhibitors of protein synthesis such as acriflavine. The repurposing of these old/existing microbicides and the development of second-generation inhibitors of ribosomal translation might constitute a valuable strategy to eradicate CSC traits in tumor tissues.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
The authors thank Dr. Kenneth McCreath for the support during editing of the manuscript.
Funding
This work was supported by grants from the Ministerio de Ciencia e Innovación (Grant SAF2012–38914), Plan Nacional de I+D+I, Spain and the Agència de Gestió d'Ajuts Universitaris i de Recerca (AGAUR) (Grant 2014 SGR229), Departament d'Economia I Coneixement, Catalonia, Spain to Javier A. Menendez.
References
- 1.Lamb R, Harrison H, Smith DL, Townsend PA, Jackson T, Ozsvari B, Martinez-Outschoorn UE, Pestell RG, Howell A, Lisanti MP, et al.. Targeting tumor-initiating cells: Eliminating anabolic cancer stem cells with inhibitors of protein synthesis or by mimicking caloric restriction. Oncotarget 2015; 6:4585-601. PMID;25671304; DOI: 10.18632/oncotarget.3278 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kakarala M, Wicha MS.. Implications of the cancer stem-cell hypothesis for breast cancer prevention and therapy. J Clin Oncol 2008; 26:2813-20; PMID:18539959; http://dx.doi.org/ 10.1200/JCO.2008.16.3931 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Scheel C, Weinberg RA.. Cancer stem cells and epithelial-mesenchymal transition: concepts and molecular links. Semin Cancer Biol. 2012; 22:396-403; PMID:22554795; http://dx.doi.org/ 10.1016/j.semcancer.2012.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Marjanovic ND, Weinberg RA, Chaffer CL.. Cell plasticity and heterogeneity in cancer. Clin Chem 2013; 59:168-79; PMID:23220226; http://dx.doi.org/ 10.1373/clinchem.2012.184655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Meacham CE, Morrison SJ.. Tumour heterogeneity and cancer cell plasticity. Nature 2013; 501:328-37; PMID:24048065; http://dx.doi.org/ 10.1038/nature12624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Pattabiraman DR, Weinberg RA.. Tackling the cancer stem cells - what challenges do they pose? Nat Rev Drug Discov 2014; 13:497-512; PMID:24981363; http://dx.doi.org/ 10.1038/nrd4253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chaffer CL, Weinberg RA.. How does multistep tumorigenesis really proceed? Cancer Discov 2015; 5:22-4; PMID:25583800; http://dx.doi.org/ 10.1158/2159-8290.CD-14-0788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Heiser LM, Sadanandam A, Kuo WL, Benz SC, Goldstein TC, Ng S, Gibb WJ, Wang NJ, Ziyad S, Tong F, et al.. Subtype and pathway specific responses to anticancer compounds in breast cancer. Proc Natl Acad Sci U S A 2012; 109:2724-9; PMID:22003129; http://dx.doi.org/ 10.1073/pnas.1018854108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Prat A, Karginova O, Parker JS, Fan C, He X, Bixby L, Harrell JC, Roman E, Adamo B, Troester M, et al.. Characterization of cell lines derived from breast cancers and normal mammary tissues for the study of the intrinsic molecular subtypes. Breast Cancer Res Treat 2013; 142:237-55; PMID:24162158; http://dx.doi.org/ 10.1007/s10549-013-2743-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Prat A, Perou CM.. Deconstructing the molecular portraits of breast cancer. Mol Oncol 2011; 5:5-23; PMID:21147047; http://dx.doi.org/ 10.1016/j.molonc.2010.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Prat A, Parker JS, Karginova O, Fan C, Livasy C, Herschkowitz JI, He X, Perou CM.. Phenotypic and molecular characterization of the claudin-low intrinsic subtype of breast cancer. Breast Cancer Res. 2010; 12:R68; PMID:20813035; http://dx.doi.org/ 10.1186/bcr2635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Morel AP, Hinkal GW, Thomas C, Fauvet F, Courtois-Cox S, Wierinckx A, Devouassoux-Shisheboran M, Treilleux I, Tissier A, Gras B, et al.. EMT inducers catalyze malignant transformation of mammary epithelial cells and drive tumorigenesis towards claudin-low tumors in transgenic mice. PLoS Genet 2012; 8:e1002723; PMID:22654675; http://dx.doi.org/ 10.1371/journal.pgen.1002723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Creighton CJ, Li X, Landis M, Dixon JM, Neumeister VM, Sjolund A, Rimm DL, Wong H, Rodriguez A, Herschkowitz JI, et al.. Residual breast cancers after conventional therapy display mesenchymal as well as tumor-initiating features. Proc Natl Acad Sci U S A 2009;106:13820-5; PMID:19666588; http://dx.doi.org/ 10.1073/pnas.0905718106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chandriani S, Frengen E, Cowling VH, Pendergrass SA, Perou CM, Whitfield ML, Cole MD.. A core MYC gene expression signature is prominent in basal-like breast cancer but only partially overlaps the core serum response. PLoS One 2009; 4:e6693; PMID:19690609; http://dx.doi.org/ 10.1371/journal.pone.0006693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Terunuma A, Putluri N, Mishra P, Mathé EA, Dorsey TH, Yi M, Wallace TA, Issaq HJ, Zhou M, Killian JK, et al.. MYC-driven accumulation of 2-hydroxyglutarate is associated with breast cancer prognosis. J Clin Invest 2014; 124:398-412; PMID:24316975; http://dx.doi.org/ 10.1172/JCI71180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liu JC, Voisin V, Wang S, Wang DY, Jones RA, Datti A, Uehling D, Al-awar R, Egan SE, Bader GD, et al.. Combined deletion of Pten and p53 in mammary epithelium accelerates triple-negative breast cancer with dependency on eEF2K. EMBO Mol Med 2014; 6:1542-60; PMID:25330770; http://dx.doi.org/ 10.15252/emmm.201404402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ghosh S, Varela L, Sood A, Park BH, Lotan TL.. mTOR signaling feedback modulates mammary epithelial differentiation and restrains invasion downstream of PTEN loss. Cancer Res 2013; 73:5218-31; PMID:23774212; http://dx.doi.org/ 10.1158/0008-5472.CAN-13-0429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jönsson G, Staaf J, Olsson E, Heidenblad M, Vallon-Christersson J, Osoegawa K, de Jong P, Oredsson S, Ringnér M, Höglund M, et al.. High-resolution genomic profiles of breast cancer cell lines assessed by tiling BAC array comparative genomic hybridization. Genes Chromosomes Cancer 2007; 46:543-58; PMID:17334996; http://dx.doi.org/ 10.1002/gcc.20438. [DOI] [PubMed] [Google Scholar]
- 19.Oliveras-Ferraros C, Vazquez-Martin A, Martin-Castillo B, Cufí S, Del Barco S, Lopez-Bonet E, Brunet J, Menendez JA.. Dynamic emergence of the mesenchymal CD44(pos)CD24(neg/low) phenotype in HER2-gene amplified breast cancer cells with de novo resistance to trastuzumab (Herceptin). Biochem Biophys Res Commun 2010; 397:27-33; PMID:20470755; http://dx.doi.org/ 10.1016/j.bbrc.2010.05.041. [DOI] [PubMed] [Google Scholar]
- 20.Oliveras-Ferraros C, Vazquez-Martin A, Martin-Castilló B, Pérez-Martínez MC, Cufí S, Del Barco S, Bernado L, Brunet J, López-Bonet E, Menendez JA.. Pathway-focused proteomic signatures in HER2-overexpressing breast cancer with a basal-like phenotype: new insights into de novo resistance to trastuzumab (Herceptin). Int J Oncol 2010;37:669-78; PMID:20664936. [DOI] [PubMed] [Google Scholar]
- 21.Oliveras-Ferraros C, Vazquez-Martin A, Cufí S, Torres-Garcia VZ, Sauri-Nadal T, Barco SD, Lopez-Bonet E, Brunet J, Martin-Castillo B, Menendez JA.. Inhibitor of Apoptosis (IAP) survivin is indispensable for survival of HER2 gene-amplified breast cancer cells with primary resistance to HER1/2-targeted therapies. Biochem Biophys Res Commun 2011; 407:412-9; PMID:21402055; http://dx.doi.org/ 10.1016/j.bbrc.2011.03.039. [DOI] [PubMed] [Google Scholar]
- 22.Cufí S, Vazquez-Martin A, Oliveras-Ferraros C, Martin-Castillo B, Vellon L, Menendez JA.. Autophagy positively regulates the CD44(+) CD24(−/low) breast cancer stem-like phenotype. Cell Cycle 2011. November 15;10:3871-85; http://dx.doi.org/ 10.4161/cc.10.22.17976. [DOI] [PubMed] [Google Scholar]
- 23.Cufi S, Corominas-Faja B, Vazquez-Martin A, Oliveras-Ferraros C, Dorca J, Bosch-Barrera J, Martin-Castillo B, Menendez JA.. Metformin-induced preferential killing of breast cancer initiating CD44+CD24-/low cells is sufficient to overcome primary resistance to trastuzumab in HER2+ human breast cancer xenografts. Oncotarget 2012;3:395-8; PMID:22565037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Martin-Castillo B, Oliveras-Ferraros C, Vazquez-Martin A, Cufí S, Moreno JM, Corominas-Faja B, Urruticoechea A, Martín ÁG, López-Bonet E, Menendez JA.. Basal/HER2 breast carcinomas: integrating molecular taxonomy with cancer stem cell dynamics to predict primary resistance to trastuzumab (Herceptin). Cell Cycle 2013; 12:225-45; PMID:23255137; http://dx.doi.org/ 10.4161/cc.23274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wong W, Bai XC, Brown A, Fernandez IS, Hanssen E, Condron M, Tan YH, Baum J, Scheres SH.. Cryo-EM structure of the Plasmodium falciparum 80S ribosome bound to the anti-protozoan drug emetine. Elife 2014; 3; http://dx.doi.org/ 10.7554/eLife.03080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Sauter KA, Wood LJ, Wong J, Iordanov M, Magun BE.. Doxorubicin and daunorubicin induce processing and release of interleukin-1β through activation of the NLRP3 inflammasome. Cancer Biol Ther 2011; 11:1008-16; PMID:21464611; http://dx.doi.org/ 10.4161/cbt.11.12.15540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Parmar N, Wetton N, Alvarado S, Kennedy S.. Profiling of human normal and cancer cell lines using phenotype microarray analysis. FASEB J 2014; 28:613.5. [Google Scholar]
- 28.Silvera D, Formenti SC, Schneider RJ.. Translational control in cancer. Nat Rev Cancer 2010; 10:254-66; PMID:20332778; http://dx.doi.org/ 10.1038/nrc2824. [DOI] [PubMed] [Google Scholar]
- 29.van Riggelen J, Yetil A, Felsher DW.. MYC as a regulator of ribosome biogenesis and protein synthesis. Nat Rev Cancer 2010; 10:301-9; PMID:20332779; http://dx.doi.org/ 10.1038/nrc2819. [DOI] [PubMed] [Google Scholar]
- 30.Ashburn TT, Thor KB.. Drug repositioning: identifying and developing new uses for existing drugs. Nat Rev Drug Discov 2004; 3:673-83; PMID:15286734; http://dx.doi.org/ 10.1038/nrd1468. [DOI] [PubMed] [Google Scholar]
- 31.Vazquez-Martin A, López-Bonetc E, Cufí S, Oliveras-Ferraros C, Del Barco S, Martin-Castillo B, Menendez JA.. Repositioning chloroquine and metformin to eliminate cancer stem cell traits in pre-malignant lesions. Drug Resist Updat 2011; 14:212-23; PMID:21600837; http://dx.doi.org/ 10.1016/j.drup.2011.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Visnyei K, Onodera H, Damoiseaux R, Saigusa K, Petrosyan S, De Vries D, Ferrari D, Saxe J, Panosyan EH, Masterman-Smith M, et al.. A molecular screening approach to identify and characterize inhibitors of glioblastoma stem cells. Mol Cancer Ther 2011; 10:1818-1828; PMID:21859839; http://dx.doi.org/ 10.1158/1535-7163.MCT-11-0268. [DOI] [PMC free article] [PubMed] [Google Scholar]