

Abstract
In order to study the molecular pathways of Parkinson's disease (PD) and to develop novel therapeutic strategies, scientific investigators rely on animal models. The identification of PD-associated genes has led to the development of genetic PD models. Most transgenic α-SYN mouse models develop gradual α-SYN pathology but fail to display clear dopaminergic cell loss and dopamine-dependent behavioral deficits. This hurdle was overcome by direct targeting of the substantia nigra with viral vectors overexpressing PD-associated genes. Local gene delivery using viral vectors provides an attractive way to express transgenes in the central nervous system. Specific brain regions can be targeted (e.g. the substantia nigra), expression can be induced in the adult setting and high expression levels can be achieved. Further, different vector systems based on various viruses can be used. The protocol outlines all crucial steps to perform a viral vector injection in the substantia nigra of the rat to develop a viral vector-based alpha-synuclein animal model for Parkinson's disease.
Keywords: Medicine, Issue 108, rAAV vectors, animal model, Parkinson's disease, alpha-synuclein, stereotactic injections, brain transduction
Introduction
To study the pathophysiology of PD and to develop novel therapeutic strategies, there is an urgent need for animal models that closely resemble the neuropathology, physiology and motor symptoms of human PD. The higher the predictive value, the better we can translate new therapies from animal models to patients.
The discovery of alpha-synuclein (α-SYN) as the first PARK gene in 1997 led to the development of the first genetic PD models. Many transgenic mice overexpressing human wild-type (WT) or mutant (A30P, A53T) α-SYN have been generated over the last decade. The levels of α-SYN overexpression have proven to be crucial in the development of the pathology. Also the mouse strain, the presence or absence of endogenous α-SYN and whether the full length or a truncated form is expressed, plays a role (detailed review by Magen and Chesselet1). Overexpression of both WT and several clinical mutants of human α-SYN in transgenic mice induces pathological accumulation of α-SYN and neuronal dysfunction2-6. However, until now most transgenic α-SYN mouse models failed to display clear dopaminergic cell loss and dopamine-dependent behavioral deficits.
This hurdle was overcome by direct targeting of the substantia nigra (SN) with viral vectors overexpressing α-SYN. Viral vectors are derived from viruses that can easily infect cells, introduce genetic material into their host genome and force the host cell to replicate the viral genome in order to produce new virus particles. Viruses can be engineered to non-replicating viral vectors that retain their ability to enter cells and introduce genes. By deleting parts of the viral genome and replacing them by the genes of interest, application of the vector will result in a single round infection without replication in the host cell, generally designated as 'transduction'. Viral vectors can be used for both overexpression and gene silencing. The expressed transgene can be a reporter protein (e.g. green fluorescent protein or firefly luciferase)7, a therapeutic protein for gene therapy applications8-10 or, as we will focus on in this paper, a disease-related protein used for disease modeling11-14.
Viral vector-mediated gene delivery provides an alternative way to express transgenes in the CNS with several advantages. Using local transgene delivery, specific brain regions can be targeted. Further, transgene expression can be induced during adulthood decreasing the risk of compensatory mechanisms during development. Also, models can be created in different species and strains. And finally, different transgenes can easily be combined. Using viral vectors, high transgene expression levels can be achieved, which might be crucial since the disease onset and severity frequently depend on the level of overexpression.
Several vector systems based on different viruses have been developed. The choice of the vector system depends on the size of the gene of interest, the required duration of gene expression, the target cell and biosafety issues. For stable gene transfer in the brain, lentiviral (LV) and recombinant adeno-associated viral (rAAV) vectors are now considered the vector systems of choice since they lead to efficient and long-term gene expression in the rodent brain. For specific targeting of the dopaminergic neurons (DN) of the SN, rAAV vectors have gradually outcompeted LV vectors because of their higher titers and transduction efficiency of DN.
The best α-SYN based rodent models currently available have been developed from a combined approach using newer AAV serotypes (rAAV 1, 5, 6, 7, 8) and optimized vector constructs, titers, and purity15,16. The vector titer as well as the vector purity directly influences the phenotypic outcome of the model. Excessive vector titers or insufficiently purified vector batches may result in non-specific toxicity. Therefore, appropriate control vectors are indispensable. Considerable time investment in the viral vector production, upscaling, and purification procedures have also proven essential to obtain reproducible and high quality vector batches.
Protocol
All animal experiments are carried out in accordance with the European Communities Council Directive of 24 November 1986 (86/609/EEC) and approved by the Bioethical Committee of the University of Leuven (Belgium).
1. Recombinant AAV Production and Purification
Note: rAAV vector production and purification was performed by the Leuven Viral Vector Core (LVVC) as previously described17.
Briefly, transfect subconfluent low (<50) passage adherent HEK 293T cells using a 25kD linear polyethylenimine 150 nM NaCl transfection solution and three different plasmids in a ratio of 1:1:1 in DMEM medium 2% foetal bovine serum. After 24 hr of incubation at 37 °C in a 5% CO2, replace the medium with fresh DMEM medium 2% foetal bovine serum. Note: The plasmids include the constructs for the AAV7 serotype, the AAV transfer plasmid encoding the human A53T mutant α-SYN under the control of the CMVie enhanced synapsin1 promoter and the pAdvDeltaF6 adenoviral helper plasmid17.
Harvest the medium 5 days after transient transfection and concentrate using tangential flow filtration17.
Purify the rAAV vector particles from the concentrated medium using an iodixanol step gradient17.
Use standard techniques of real-time PCR for genomic copy (GC) determination. In this protocol, a vector titer of 3.0 E11 GC/ml was used to develop an α-SYN based rat model for PD17.
2. Stereotactic injection of rAAV α-SYN Vector in the SN of the Rat (Figure 2)
House eight weeks old female Wistar rats weighing about 200-250 g under a normal 12 hr light/dark cycle with free access to pelleted food and tap water.
Submit the rat to intraperitoneal (i.p.) anesthesia containing a mixture of ketamine (60 mg/kg) and medetomidine (0.4 mg/kg). Once the rat is anesthetized and doesn't react when squeezing the different paws, administer a micro-transponder subcutaneously on the back of the rat for further recognition using a micro-transponder implanter. Check if the micro-transponder is positioned correctly and can be read out by the reading device.
Cut the hair on top of the scalp. Apply a local anesthetic on both the scalp and the ears. Perform the rest of the surgical procedure under a laminar flow using aseptic techniques.
Place the rats in a stereotactic head frame using two ear bars, a mouth and a nose bar. Cover the body of the rat with a paper blanket to avoid a drop in body temperature. Apply an ocular lubricant to prevent the eyes from drying.
Disinfect the scalp with jodium 1% in isopropanol 70% and make a small incision in the midline of the scalp. Gently scrape away the membranes on the skull and rinse with saline. Let the skull dry for several minutes. Observe the cranial sutures and the two reference points: Bregma and Lambda.
To inject the rAAV vector into the SN, define the coordinates towards Bregma (anteroposterior: 5.3 mm; mediolateral: 2.0 mm and dorsoventral: 7.2 mm calculated from the dura). Note: The three dimensional coordinates for each region of interest can be calculated using a stereotaxic atlas of the rat brain, applying Bregma as anatomical reference point.
Fill a 10 µl microinjection syringe (30 gauge 20 mm) with rAAV vector and place it in the stereotaxic instrument connected with a motorized microinjection pump. Control the volume by releasing a drop of vector and eliminate in a polyvalent cleaning detergent pH 9 (e.g. RBS).
- Visually check if the head is fixed straight in the head frame and evaluate the left-right axis. Carefully visually define the anteroposterior and mediolateral coordinates for Bregma and Lambda and measure their height using a 30 gauge 20 mm needle in the dorsoventral arm of the stereotactic frame.
- Allow a maximum of 0.3 mm difference in height between Bregma and Lambda. Place the needle back on Bregma and apply the anteroposterior and mediolateral coordinates by moving the anteroposterior and the mediolateral arm of the stereotactic frame.
At the place of injection, measure the height of the skull and ensure that it does not differ more than 0.3 mm from the height of Bregma. Drill a hole in the skull with a diameter of approximately 2 mm. Measure the height of the dura, this will serve as a reference to apply the dorsoventral coordinate. Alternatively subtract a fixed thickness for the skull (0.9 mm).
Penetrate the dura using a 26 gauge needle and absorb the blood with a sterile tissue. Wait until all bleeding has stopped before proceeding.
Slowly insert the 10 µl microinjection syringe pre-loaded with vector solution into the brain to the pre-determined depth (dorsoventral coordinate). Wait 1 min with the needle in place. Inject 3 µl of vector solution (3.0 E11 genome copies/ml (medium vector dose) or 1.0 E12 GC/ml (high vector dose) of rAAV2/7 α-SYN or eGFP control vector) using the motorized microinjection pump with a throughput of 0.25 µl/min.
After injection, keep the needle in place for another 5 min before slowly removing it. Stitch the scalp using coated braided polyester 3.0, disinfect with 1% jodium in 70% isopropanol and gently remove the animal from the stereotactic instrument. First loosen the nose and mouth bar, then the two ear bars.
To reverse the anesthesia, inject the rat intraperitoneally with 0.5 mg/kg atipamezole and place the rat in a clean cage on a heating plate of 38 °C until it wakes up. Cover the rat with a paper blanket to prevent a drop in body temperature.
Provide easy access to food and water for the first hours. Monitor the rat for the first few days. If necessary apply analgesia. Note: There is no need to remove the stitches from the skull. After 1-2 weeks the skull is completely repaired and the stiches come loose.
3. Assessment of rAAV2/7 α-SYN Injected Rats Using Non-invasive PET Imaging, Behavioral Tests and Immunohistochemical Analysis
To follow up the kinetics of nigrostriatal dopaminergic neurodegeneration non-invasively over time in individual animals, quantify dopamine transporter (DAT) binding using small-animal positron emission tomography (PET) and a tracer of the DA Transporter e.g. [18F]-FECT16.
- To examine whether the level of dopaminergic neurodegeneration is sufficient to induce motor impairments in the rats, subject the rats to the cylinder test to evaluate spontaneous forelimb use.
- Place the rat in a 20 cm wide clear glass cylinder and videotape the behavior during vertical movements along the wall and landing after a rear. Score the number of contacts made by each forepaw for a total of 20 contacts. For detailed description of the scoring criteria see Schallert et al.18 Express the number of impaired forelimb contacts (e.g. left forepaw) as a percentage of total forelimb contacts (left plus right forepaw). Note: Non-lesioned control rats using both paws equally should score around 50% in this test.
- Perform immunohistochemical (IHC) analysis to assess the level of transgene expression and dopaminergic cell loss.
- At different end stages, sacrifice the rats with an overdose of sodium pentobarbital (60 mg/kg, i.p.) and perform an intracardial perfusion with cold saline followed by 4% paraformaldehyde in PBS19. Fixate the brains overnight at 4 °C and cut 50 µm thick coronal brain sections using a vibrating microtome.
- Perform IHC staining on free-floating sections using antibodies against α-SYN and tyrosine hydroxylase to analyze α-SYN expression levels and the level of neurodegeneration16.
Representative Results
The overall scheme of the experiment is depicted in Figure 1
rAAV 2/7-mediated overexpression of A53T α-SYN induces dopamine-dependent motor deficits. To examine whether the level of α-SYN overexpression is sufficient to induce motor impairments in the rats, we subjected the rats to the cylinder test to evaluate spontaneous forelimb use (Figure 3A). From 3 weeks after injection, a significant motor impairment was seen in rats that received a dose 3.0 E11 GC/ml of A53T α-SYN rAAV2/7 vector. At 4 weeks after injection a 50% decrease in spontaneous contralateral (left) forepaw use was observed, whereas the control eGFP rAAV2/7 injected animals showed no asymmetry in forepaw use (Figure 3B). Rats that received a higher A53T α-SYN rAAV2/7 vector dose showed a more pronounced impairment of forepaw use (70%) at 29 days after injection (Figure 3C). To prove that the observed motor impairment was dopamine-dependent, we administered a single dose of L-DOPA (6 mg/kg i.p.) to the rats injected with a high vector dose. When we repeated the cylinder test 45 min after L-DOPA treatment, a full recovery of the forepaw use in the A53T α-SYN rAAV2/7 injected animals was observed (Figure 3C).
PET imaging allows non-invasive imaging of α-SYN induced progressive neurodegeneration. To follow up the kinetics of nigrostriatal dopaminergic neurodegeneration non-invasively over time in individual animals, we quantified dopamine transporter (DAT) binding using small-animal positron emission tomography (PET) with [18F]-FECT as radioligand. DAT binding significantly decreased in the ipsilateral caudate-putamen of A53T α-SYN rAAV2/7 injected rats over time but remained stable in the eGFP control animal (Figure 4A-4B). Quantification of DAT binding of A53T α-SYN rAAV2/7 injected animals showed a maximal rate of nigrostriatal dopaminergic degeneration between day 7 and 21 after injection. After 32 days, a decrease in DAT binding of up to 85% was observed (Figure 4C). As a positive control, injection of the neurotoxin 6-OHDA in the SN induced 90% loss of DAT binding within 7 days (Figure 4B-4C).
Stereotactic injection of rAAV2/7 A53T α-SYN in the SN of the rat induces nigral dopaminergic cell death and formation of insoluble α-SYN positive aggregates. To analyze the level of α-SYN overexpression and dopaminergic cell loss we sacrificed the animals at different time points. IHC was performed on free-floating sections using an antibody against α-synuclein (rabbit polyclonal 1:5,000). This antibody can detect both human and rat α-synuclein, but endogenous levels of rat α-synuclein were below detection limits within nigral cell somata, owing to its predominant localization at synaptic membranes. To asses cell loss we used an antibody against TH (rabbit polyclonal 1:1,000).
Four days after rAAV vector injection, α-SYN or eGFP expression was detected in the SN (Figure 5A-5C) of the rats. The majority (>90%) of the DN was efficiently transduced and both transgenic proteins were localized in the cell bodies and axons. At 29 days post injection (p.i.) a substantial reduction in α-SYN expression was observed in the SNpc, while it was still detectable in areas surrounding the SN (Figure 5B-5C). Next, we analysed the level of nigral cell loss. A rapid and progressive loss of up to 80% of TH-positive neurons was detected over 29 days in rats injected with A53T α-SYN rAAV2/7 (Figure 5D-5H). Of note, overexpression of wild type instead of A53T α-SYN resulted in similar dopaminergic neurodegeneration (data not shown). The loss of the DN in the SN was paralleled by a robust decrease of TH-positive nerve terminals in the striatum (STR) (Figure 5F). To rule out specific vector batch effects, different α-SYN vector preparations were tested in the SN with similar results. No reduction in TH staining was observed in the SN or STR of eGFP rAAV2/7 injected control animals (Figure 5E-5H). Next to dopaminergic neurodegeneration, the presence of α-synucleinopathy is a second important hallmark of PD. Despite the short time course of our model (four weeks), we observed both α-SYN-positive cytoplasmic aggregates in the SN and dystrophic neurites in the STR (Figure 5I). Ubiquitin immunoreactivity is a distinct feature of Lewy body pathology in the human brain20-22. We observed co-localization of α-SYN and ubiquitin at 29 days p.i. in a fraction (± 20%) of the α-SYN expressing nigral neurons (Figure 5J). The fibrillar nature of the α-SYN aggregates was evaluated by Thioflavin S (Thio S) staining23. Thio S positive cells were detected in the SN from 17 days onwards (Figure 5J).
 Figure 1: Stereotactic injection of rAAV2/7 α-SYN vector results in progressive neurodegeneration. Stereotactic injection of rAAV2/7 α-SYN vector in the SN of the rat induces dopaminergic neurodegeneration measured via behavior analysis (cylinder test), non-invasive PET imaging and IHC analysis. Please click here to view a larger version of this figure.
Figure 1: Stereotactic injection of rAAV2/7 α-SYN vector results in progressive neurodegeneration. Stereotactic injection of rAAV2/7 α-SYN vector in the SN of the rat induces dopaminergic neurodegeneration measured via behavior analysis (cylinder test), non-invasive PET imaging and IHC analysis. Please click here to view a larger version of this figure.
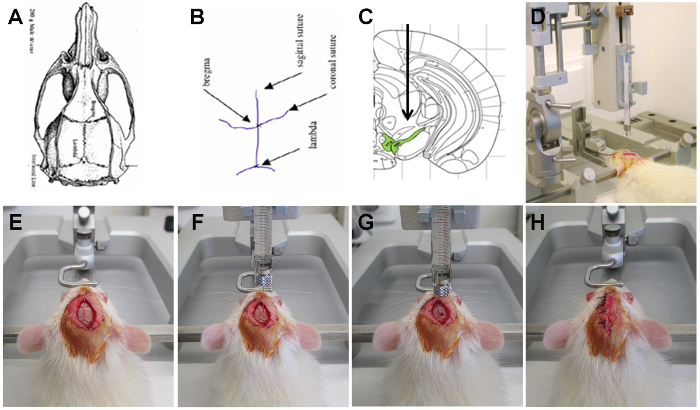 Figure 2: Stereotactic injection of rAAV2/7 vector encoding α-SYN in the SN of the rat. (A, B, E) The cranial sutures on the rat skull, defining the two reference points: Bregma and Lambda. (C) A stereotaxic atlas of the rat brain presenting the region of injection namely the SN. (D) A Wistar rat positioned in a stereotactic head frame using two ear bars, a mouth and a nose bar. (F) The 30 gauge needle filled with vector is placed in position for the substantia nigra. (G) A small whole is drilled at the site of injection and the needle is placed in position. (H) After injection the scalp is stitched and disinfected. Please click here to view a larger version of this figure.
Figure 2: Stereotactic injection of rAAV2/7 vector encoding α-SYN in the SN of the rat. (A, B, E) The cranial sutures on the rat skull, defining the two reference points: Bregma and Lambda. (C) A stereotaxic atlas of the rat brain presenting the region of injection namely the SN. (D) A Wistar rat positioned in a stereotactic head frame using two ear bars, a mouth and a nose bar. (F) The 30 gauge needle filled with vector is placed in position for the substantia nigra. (G) A small whole is drilled at the site of injection and the needle is placed in position. (H) After injection the scalp is stitched and disinfected. Please click here to view a larger version of this figure.
 Figure 3: rAAV 2/7-mediated overexpression of A53T α-SYN induces dopamine-dependent motor deficits. (A, B) Cylinder test at different time points after injection of rAAV2/7 A53T α-SYN. (Mean ± s.d., * p <0.05 versus 17 days, # p <0.05 eGFP controls by ANOVA and Tukey post hoc test, n = 5). (C) Cylinder test at different time points after injection of rAAV2/7 A53T α-SYN (high vector dose). (Mean ± s.d., * p <0.05 4 days versus 29 days by ANOVA and Tukey post hoc test, n = 5). The test was performed with or without administration of Levodopa (L-DOPA). (Mean ± s.d., * p <0.05 non-treated versus treated animals by ANOVA and Tukey post hoc test, n = 5). Reprinted from Neurobiology of aging, Vol. 36, Van der Perren et al., Longitudinal follow-up and characterization of a robust rat model for Parkinson's disease based on overexpression of alpha-synuclein with adeno-associated viral vectors, 1543-1558, (2015), with permission from Elsevier. Please click here to view a larger version of this figure.
Figure 3: rAAV 2/7-mediated overexpression of A53T α-SYN induces dopamine-dependent motor deficits. (A, B) Cylinder test at different time points after injection of rAAV2/7 A53T α-SYN. (Mean ± s.d., * p <0.05 versus 17 days, # p <0.05 eGFP controls by ANOVA and Tukey post hoc test, n = 5). (C) Cylinder test at different time points after injection of rAAV2/7 A53T α-SYN (high vector dose). (Mean ± s.d., * p <0.05 4 days versus 29 days by ANOVA and Tukey post hoc test, n = 5). The test was performed with or without administration of Levodopa (L-DOPA). (Mean ± s.d., * p <0.05 non-treated versus treated animals by ANOVA and Tukey post hoc test, n = 5). Reprinted from Neurobiology of aging, Vol. 36, Van der Perren et al., Longitudinal follow-up and characterization of a robust rat model for Parkinson's disease based on overexpression of alpha-synuclein with adeno-associated viral vectors, 1543-1558, (2015), with permission from Elsevier. Please click here to view a larger version of this figure.
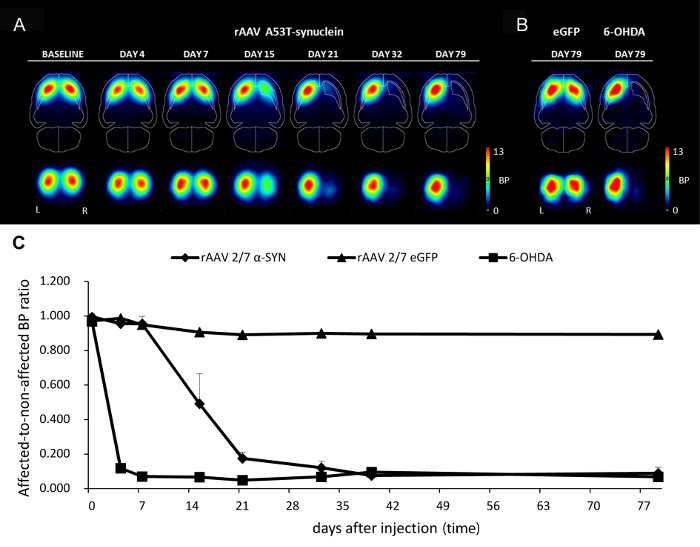 Figure 4: Non-invasive imaging of A53T α-SYN induced dopaminergic cell death using DAT PET imaging. (A-B) Series of horizontal and coronal slices depicting mean striatal DAT binding of (A) rAAV2/7 A53T α-SYN injected animals at different time points after injection (n = 7) and (B) rAAV2/7 eGFP injected (n = 1) or 6-OHDA treated control animals (n =1) 79 days after injection. Color bars indicate binding potentials for the DAT. (C) Quantification of the DAT binding of rAAV2/7 A53T α-SYN, rAAV2/7 eGFP and 6-OHDA injected animals measured at different time points (data represent mean ± s.d.). Reprinted from Neurobiology of aging, Vol. 36, Van der Perren et al., Longitudinal follow-up and characterization of a robust rat model for Parkinson's disease based on overexpression of alpha-synuclein with adeno-associated viral vectors, 1543-1558, (2015), with permission from Elsevier. Please click here to view a larger version of this figure.
Figure 4: Non-invasive imaging of A53T α-SYN induced dopaminergic cell death using DAT PET imaging. (A-B) Series of horizontal and coronal slices depicting mean striatal DAT binding of (A) rAAV2/7 A53T α-SYN injected animals at different time points after injection (n = 7) and (B) rAAV2/7 eGFP injected (n = 1) or 6-OHDA treated control animals (n =1) 79 days after injection. Color bars indicate binding potentials for the DAT. (C) Quantification of the DAT binding of rAAV2/7 A53T α-SYN, rAAV2/7 eGFP and 6-OHDA injected animals measured at different time points (data represent mean ± s.d.). Reprinted from Neurobiology of aging, Vol. 36, Van der Perren et al., Longitudinal follow-up and characterization of a robust rat model for Parkinson's disease based on overexpression of alpha-synuclein with adeno-associated viral vectors, 1543-1558, (2015), with permission from Elsevier. Please click here to view a larger version of this figure.
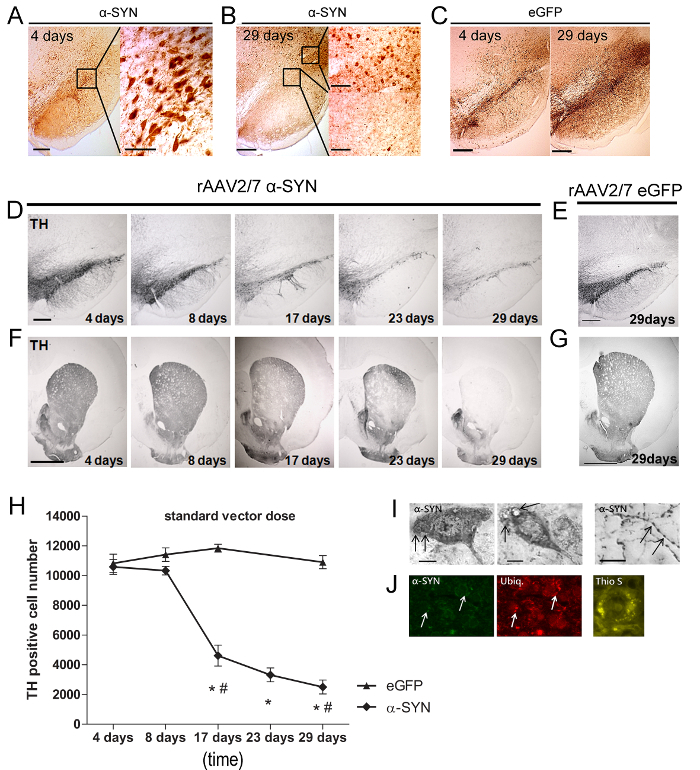 Figure 5: rAAV 2/7-mediated overexpression of A53T α-SYN induces dopaminergic cell death and formation of insoluble α-SYN positive aggregates. (A-B) IHC staining demonstrating α-SYN overexpression 4 days and 29 days after rAAV mediated transfer in rat SN. Inserts show magnifications of the selected area. Scale bar = 400 µm (overview picture left), 70 µm and 200 µm (inserts right). (C) IHC staining demonstrating eGFP overexpression 4 days and 29 days after rAAV mediated transfer in rat SN. Scale bar = 400 µm. (D-G) IHC staining for TH in the SN and STR at different time points after injection of (D, F) rAAV2/7 α-SYN or 29 days after injection of (E, G) rAAV2/7 eGFP in the SN. Scale bar a, c = 400 µm, b, d = 1,000 µm. (H) Stereological quantification of the number of TH-positive neurons in the SN over time after rAAV2/7 A53T α-SYN injection or rAAV2/7 eGFP control vector (Mean ± s.d., * p <0.05 versus 8 days, # p <0.05 versus eGFP controls by ANOVA and Tukey post hoc test, n = 5). (I) IHC staining demonstrating α-SYN pathology, including cytoplasmic aggregates in the SN and dystrophic and bulging neurites in the STR, after intranigral rAAV2/7 A53T α-SYN injection. (J) Representative confocal images of fluorescent double immunostainings for α-SYN (green) and ubiquitin (red) show an increase in co-localization over time (arrows). Scale bar c = 50 µm. Thioflavin S staining of SN 29 days after injection of rAAV2/7 A53T α-SYN. Scale bar D = 30 µm. Reprinted from Neurobiology of aging, Vol. 36, Van der Perren et al., Longitudinal follow-up and characterization of a robust rat model for Parkinson's disease based on overexpression of alpha-synuclein with adeno-associated viral vectors, 1543-1558, (2015), with permission from Elsevier. Please click here to view a larger version of this figure.
Figure 5: rAAV 2/7-mediated overexpression of A53T α-SYN induces dopaminergic cell death and formation of insoluble α-SYN positive aggregates. (A-B) IHC staining demonstrating α-SYN overexpression 4 days and 29 days after rAAV mediated transfer in rat SN. Inserts show magnifications of the selected area. Scale bar = 400 µm (overview picture left), 70 µm and 200 µm (inserts right). (C) IHC staining demonstrating eGFP overexpression 4 days and 29 days after rAAV mediated transfer in rat SN. Scale bar = 400 µm. (D-G) IHC staining for TH in the SN and STR at different time points after injection of (D, F) rAAV2/7 α-SYN or 29 days after injection of (E, G) rAAV2/7 eGFP in the SN. Scale bar a, c = 400 µm, b, d = 1,000 µm. (H) Stereological quantification of the number of TH-positive neurons in the SN over time after rAAV2/7 A53T α-SYN injection or rAAV2/7 eGFP control vector (Mean ± s.d., * p <0.05 versus 8 days, # p <0.05 versus eGFP controls by ANOVA and Tukey post hoc test, n = 5). (I) IHC staining demonstrating α-SYN pathology, including cytoplasmic aggregates in the SN and dystrophic and bulging neurites in the STR, after intranigral rAAV2/7 A53T α-SYN injection. (J) Representative confocal images of fluorescent double immunostainings for α-SYN (green) and ubiquitin (red) show an increase in co-localization over time (arrows). Scale bar c = 50 µm. Thioflavin S staining of SN 29 days after injection of rAAV2/7 A53T α-SYN. Scale bar D = 30 µm. Reprinted from Neurobiology of aging, Vol. 36, Van der Perren et al., Longitudinal follow-up and characterization of a robust rat model for Parkinson's disease based on overexpression of alpha-synuclein with adeno-associated viral vectors, 1543-1558, (2015), with permission from Elsevier. Please click here to view a larger version of this figure.
Discussion
There are several critical steps within the protocol. The vector titer as well as the vector purity directly influences the phenotypic outcome of the model. Excessive vector titers or insufficiently purified vector batches may result in non-specific toxicity. Therefore, the use of high quality vector batches and appropriate control vectors is indispensable. Further, the exact positioning of the rat's head in the stereotaxic frame and the accurate determination of the coordinates is essential in targeting the substantia nigra. After drilling the hole in the skull at the site of injection, it is important to insert the needle straight into the rat's brain without touching any margins. The needle should be slowly removed after injecting the viral vector, to prevent vector leakage. Lastly, after stitching, the scalp should be disinfected with 1% jodium in 70% isopropanol to avoid biting of the stiches by other animals. Alternatively other antiseptic reagents can be used.
The method described can also be used to develop a rAAV2/7 α-SYN based mice model for Parkinson's disease24. In mice we inject a volume of 2 µl rAAV vector in the SN. Compared to rats, mice DN seem to be somewhat less sensitive to α-SYN overexpression, resulting in a delayed manifestation of neurodegeneration. Furthermore, other regions in the brain (e.g. striatum, hippocampus, cortex, etc.) can be targeted. The coordinates for the different brain regions can be found in the stereotaxic atlas. Optimization of the coordinates can be done by Chinese ink or by using a viral vector encoding a reporter gene (e.g. eGFP). Different vectors systems (rAAV, LV, etc.) can be used depending on the application.
This technique has the limitation that each animal has to be injected individually. Therefore a trained person should perform the injections in order to minimize variations between different animals. Another limitation is that the method is time consuming (when executed by a trained person it takes around 45 min per animal). Only eight to ten animals can be injected in one day.
Viral vector-mediated gene delivery allows for specific targeting of brain regions. Using viral vectors, high transgene expression levels can be achieved, which is crucial since disease onset and severity depends on the level of α-SYN expression. Also, different doses can be applied which will result in an animal model displaying slower or faster kinetics of neurodegeneration. Lastly, this technique can be used to create models in different animal species and strains using the same vector preparation.
This procedure can be used to deliver viral vectors as well as toxins (e.g. 6-OHDA) into different regions of the brain.The transgene encoded by the vector can be a reporter protein, a therapeutic protein for gene therapy applications8-10 or a disease-related protein used for disease modeling11-14. This technique can be used to develop novel animal models which allow for preclinical drug testing and can be beneficial in studying the molecular mechanism of Parkinson's disease as well as many other neurodegenerative disorders.
Disclosures
The authors declare that there is no actual or potential conflict of interest.
Acknowledgments
The authors thank Joris Van Asselberghs and Ann Van Santvoort for their excellent technical assistance. Research was funded by the IWT-Vlaanderen (IWT SBO/80020), the FWO Vlaanderen (G.0768.10), by the EC-FP6 program 'DiMI' (LSHB-CT-2005-512146), the FP7 RTD project MEFOPA (HEALTH-2009-241791), the FP7 program 'INMiND' (HEALTH-F2-2011-278850), the KU Leuven (IOF-KP/07/001, OT/08/052A, IMIR PF/10/017), and the MJFox Foundation (Target validation 2010). A. Van der Perren and C. Casteels are a postdoctoral fellows of the Flemish Fund of Scientific Research. K. Van Laere is a senior clinical fellow of the Flemish Fund of Scientific Research.
References
- Magen I, Chesselet MF. Genetic mouse models of Parkinson's disease The state of the art. Prog Brain Res. 2010;183:53–87. doi: 10.1016/S0079-6123(10)84004-X. [DOI] [PubMed] [Google Scholar]
- Masliah E, et al. Dopaminergic loss and inclusion body formation in alpha-synuclein mice: implications for neurodegenerative disorders. Science. 2000;287:1265–1269. doi: 10.1126/science.287.5456.1265. [DOI] [PubMed] [Google Scholar]
- Freichel C, et al. Age-dependent cognitive decline and amygdala pathology in alpha-synuclein transgenic mice. Neurobiol Aging. 2007;28:1421–1435. doi: 10.1016/j.neurobiolaging.2006.06.013. [DOI] [PubMed] [Google Scholar]
- Fleming SM, Fernagut PO, Chesselet MF. Genetic mouse models of parkinsonism: strengths and limitations. NeuroRx. 2005;2:495–503. doi: 10.1602/neurorx.2.3.495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahle PJ, et al. Selective insolubility of alpha-synuclein in human Lewy body diseases is recapitulated in a transgenic mouse model. Am J Pathol. 2001;159:2215–2225. doi: 10.1016/s0002-9440(10)63072-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chesselet MF, Richter F. Modelling of Parkinson's disease in mice. Lancet Neurol. 2011;10:1108–1118. doi: 10.1016/S1474-4422(11)70227-7. [DOI] [PubMed] [Google Scholar]
- Deroose CM, Reumers V, Debyser Z, Baekelandt V. Seeing genes at work in the living brain with non-invasive molecular imaging. Curr Gene Ther. 2009;9:212–238. doi: 10.2174/156652309788488596. [DOI] [PubMed] [Google Scholar]
- Manfredsson FP, et al. rAAV-mediated nigral human parkin over-expression partially ameliorates motor deficits via enhanced dopamine neurotransmission in a rat model of Parkinson's disease. Exp Neurol. 2007;207:289–301. doi: 10.1016/j.expneurol.2007.06.019. [DOI] [PubMed] [Google Scholar]
- Vercammen L, et al. Parkin protects against neurotoxicity in the 6-hydroxydopamine rat model for Parkinson's disease. Mol Ther. 2006;14:716–723. doi: 10.1016/j.ymthe.2006.06.009. [DOI] [PubMed] [Google Scholar]
- Winklhofer KF. The parkin protein as a therapeutic target in Parkinson's disease. Expert opinion on therapeutic targets. 2007;11:1543–1552. doi: 10.1517/14728222.11.12.1543. [DOI] [PubMed] [Google Scholar]
- Kirik D, et al. Parkinson-like neurodegeneration induced by targeted overexpression of alpha-synuclein in the nigrostriatal system. J Neurosci. 2002;22:2780–2791. doi: 10.1523/JNEUROSCI.22-07-02780.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirik D, et al. Nigrostriatal alpha-synucleinopathy induced by viral vector-mediated overexpression of human alpha-synuclein: a new primate model of Parkinson's disease. Proc Natl Acad Sci U S A. 2003;100:2884–2889. doi: 10.1073/pnas.0536383100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lauwers E, et al. Neuropathology and neurodegeneration in rodent brain induced by lentiviral vector-mediated overexpression of alpha-synuclein. Brain pathology. 2003;13:364–372. doi: 10.1111/j.1750-3639.2003.tb00035.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein RL, King MA, Hamby ME, Meyer EM. Dopaminergic cell loss induced by human A30P alpha-synuclein gene transfer to the rat substantia nigra. Hum Gene Ther. 2002;13:605–612. doi: 10.1089/10430340252837206. [DOI] [PubMed] [Google Scholar]
- Vander Perren A, Van den Haute C, Baekelandt V. Viral Vector-Based Models of Parkinson's Disease. Curr Top Beh Neurosci. 2014. [DOI] [PubMed]
- Van der Perren A, et al. Longitudinal follow-up and characterization of a robust rat model for Parkinson's disease based on overexpression of alpha-synuclein with adeno-associated viral vectors. Neurobiol Aging. 2014. [DOI] [PubMed]
- Van der Perren A, et al. Efficient and stable transduction of dopaminergic neurons in rat substantia nigra by rAAV 2/1, 2/2, 2/5, 2/6.2, 2/7, 2/8 and 2/9. Gene Ther. 2011. [DOI] [PubMed]
- Schallert T, Fleming SM, Leasure JL, Tillerson JL, Bland ST. CNS plasticity and assessment of forelimb sensorimotor outcome in unilateral rat models of stroke, cortical ablation, parkinsonism and spinal cord injury. Neuropharmacology. 2000;39:777–787. doi: 10.1016/s0028-3908(00)00005-8. [DOI] [PubMed] [Google Scholar]
- Soueid J, Nokkari A, Makoukji J. Techniques and Methods of Animal Brain Surgery: Perfusion, Brain Removal, and Histological Techniques. Brain Neurotrauma: Molecular, Neuropsychological, and Rehabilitation Aspects. Frontiers in Neuroengineering. 2015. [PubMed]
- Dale GE, et al. Relationships between Lewy bodies and pale bodies in Parkinson's disease. Acta Neuropathol. 1992;83:525–529. doi: 10.1007/BF00310030. [DOI] [PubMed] [Google Scholar]
- Dawson VL. Neurobiology of flies and mice. Science. 2000;288:631–632. doi: 10.1126/science.288.5466.631. [DOI] [PubMed] [Google Scholar]
- Dawson T, Mandir A, Lee M. Animal models of PD: pieces of the same puzzle? Neuron. 2002;35:219–222. doi: 10.1016/s0896-6273(02)00780-8. [DOI] [PubMed] [Google Scholar]
- LeVine H., 3rd Quantification of beta-sheet amyloid fibril structures with thioflavin T. Methods Enzymol. 1999;309:274–284. doi: 10.1016/s0076-6879(99)09020-5. [DOI] [PubMed] [Google Scholar]
- Oliveras-Salva M, et al. rAAV2/7 vector-mediated overexpression of alpha-synuclein in mouse substantia nigra induces protein aggregation and progressive dose-dependent neurodegeneration. Mol Neurodegener. 2013;8 doi: 10.1186/1750-1326-8-44. [DOI] [PMC free article] [PubMed] [Google Scholar]