

Abstract
Envenomation by Loxosceles spider is characterized by the development of dermonecrosis. In previous studies, we have demonstrated that increased expression/secretion of matrix metalloproteinases 2 and 9, induced by Loxosceles intermedia venom Class 2 SMases D (the main toxin in the spider venom), contribute to the development of cutaneous loxoscelism. In the present study we show that the more potent venom containing the Class 1 SMase D from Loxosceles laeta, in addition to increasing the expression/secretion of MMP2 and MMP9, also stimulates the expression of MMP7 (Matrilysin-1), which was associated with keratinocyte cell death. Tetracycline, a matrix metalloproteinase inhibitor, prevented cell death and reduced MMPs expression. Considering that L. laeta venom is more potent at inducing dermonecrosis than L. intermedia venom, our results suggest that MMP7 may play an important role in the severity of dermonecrosis induced by L. laeta spider venom SMase D. In addition, the inhibition of MMPs by e.g. tetracyclines may be considered for the treatment of the cutaneous loxoscelism.
Introduction
The spiders of the genus Loxosceles, Sicariidae family, commonly known in the Americas as brown or recluse spiders, can induce serious local and systemic effects in humans upon envenomation [1]. Loxosceles envenomation can locally result in extensive tissue destruction and chronic ulcer formation which take many months to heal [2–4]. Complement mediated hemolysis, disseminated intravascular coagulation, shock and renal impairment are rare systemic reactions, but are the main causes of death associated with Loxosceles envenomation.
It is generally acknowledged that sphingomyelinase D (SMase D) is the main toxin in the venoms of Loxosceles spiders and is responsible for all effects observed in loxoscelism (reviewed in 1). Substrates of SMases D include sphingomyelin and lysophosphatidylcholine, hydrolysis of which results in the release of choline and formation of ceramide-1-phosphate and lysophosphatidic acid, both of which have the potency to activate cells [5–9].
Over 190 Loxosceles SMases D have now been sequenced and at the amino acid level, they display a significant level of sequence and structural similarity [10, 11]. Based on structural aspects of the enzyme a SMases D classification was proposed whereby SMases D possessing a single disulphide bridge and containing a variable loop were grouped in Class I, while SMasesD containing an additional intra-chain disulphide bridge that links a flexible loop with a catalytic loop were grouped in Class II [10–13].
As we showed previously, L. laeta venom (containing Class 1 SMases D) has stronger biological activities (haemolysis and dermonecrosis) than L. intermedia (containing Class 2 SMases D) [14]. This is reflected in the purified toxins where SMases D belonging to Class 1 have a stronger ability to induce dermonecrosis than those belonging to Class 2 [15]. Evolutionary, the majority of the Class I SMase D enzymes belong to the same clade, suggesting that they originate from a single common ancestor [10]. In the rabbit model of cutaneous loxoscelism, as well as in human keratinocytes cultures, we previously showed that L. intermedia Class 2 SMases D induced an increase in the expression of matrix metalloproteinases- 2 and 9 (MMP2, MMP9), which likely contribute to the pathology of cutaneous loxoscelism [16, 17].
MMPs are important targets for pharmacological intervention in a variety of diseases [18]. The protein synthesis inhibiting antibiotics, tetracycline and its chemically derivatives have shown also to inhibit metalloproteinases expression and function [19–22]. We previously showed that in vitro and in vivo tetracycline inhibited the Class 2 SMase D induced increase in expression/secretion of MMP2 and MMP9 and dermonecrosis in a rabbit model of cutaneous loxoscelism [17, 23].
Since Class 1 SMases D have a higher dermonecrosis inducing potency than the Class 2 SMase D, we aimed to investigate how the venom of L. laeta, containing Class 1 SMases D, induced keratinocyte cell death and if the dermonecrosis induced by this venom could be prevented/reduced by tetracyclines, as a potential therapeutic tool.
Materials and Methods
Chemicals, reagents, and buffers
Tween 20, bovine serum albumin (BSA), formalin, gelatin, Triton X-100, 3-(4,5 dimethylthiazol-2yl)-2,5 diphenyltetrazolium bromide—MTT and tetracycline were from Sigma (St Louis, MO, USA). Goat antibodies against mouse IgG (GAM), labeled with alkaline phosphatase (IgG-AP), 5-bromo-4-chloro-3-indolyl-phosphate (BCIP) and nitroblue tetrazolium (NBT), were purchased from Promega Corp. (Madison, Wisconsin, USA). MAP Human MMP Magnetic Bead Panel 2 was purchased from Millipore (Billerica, MA, USA). Brij-35 was from Fluka–BioChemika (Werdenberg, Switzerland). Mouse monoclonal antibody (MoAb) against human MMP7 was from R&D Systems (MN, USA). Tetracycline (5% in a hydrosoluble lanolin cream) was obtained from Oficinallis Pharma handle pharmacy (São Paulo, Brazil). Coomassie brilliant blue solution: 40% methanol, 10% acetic acid and 0.1% Coomassie brilliant blue. Buffers were: Saline: 0.9% NaCl; Phosphate-Buffered Saline (PBS), pH 7.2, containing 10 mM NaH2PO4, 150 mM NaCl; Zymography buffer, pH 8.3: 50 mM Tris-HCl, 200 mM NaCl, 10 mM CaCl2, 0,05% Brij-35.
Spiders, venoms and SMases D
Loxosceles spiders were bred and maintained in house. Venom was obtained by electrostimulation by the modified method of Bucherl [7, 24]. Recombinant SMases I (Class 1) from L. laeta, P1 and P2 (Class 2) from L. intermedia, were prepared as previously described [25, 13]. Permission to access the Loxosceles venom (permission n° 01/2009) was provided by the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA), a Brazilian Ministry of the Environment's enforcement agency.
Animals and ethics statement
Adult New Zealand white rabbits, weighing approximately 3 Kg, were supplied by the Central Animal Breeding Facilities of the Butantan Institute, SP, Brazil. The animals were kept in a room with controlled lighting [12-h light/dark cycle] and temperature (22°C) and housed individually with water and food provided ad libitum. All efforts were made to minimize animal suffering and to reduce the number of animals used. The experimental animals were handled in strict accordance with the ethical principles in animal research adopted by the Brazilian Society of Animal Science and the National Brazilian Legislation no. 11.794/08. The protocol was approved by the Institutional Animal Care and Use Committee from the Butantan Institute (permission no. 840/11).
Cell culture and maintenance
HaCaT cells (immortal human keratinocyte cell line; from Dr Fusenig, Heidelberg, Germany) was maintained in DMEM (Gibco-BRL, Gaithersburg, MD, USA), supplemented with 10% fetal bovine serum (FBS), penicillin (100 IU/mL) and streptomycin (100 IU/mL).
Viability assay
HaCaT cells were seeded in 96-well plates at 2x104 cells in 200 μl. One day before treatment, medium was removed and replaced with DMEM without FBS. Cells were, incubated with venom or SMase D in the presence or absence of tetracycline. After 24 to 72 hours, culture supernatants were collected for zymography, Western blot and Luminex analyses and cells assessed for viability by incubation with 100 μl of MTT (5 mg/mL) in PBS for 3 hours at 37°C, 5% CO2. Supernatants were removed and cells were incubated with 100 μl of DMSO and the absorbance was measured at 540 and 620 nm. Relative cell viability was calculated as [(sample OD (540-620nm)−background control OD (540-620nm))/ (control OD (540-620nm)] x 100%.
Gelatin zymography
Gelatinase activity was analyzed by zymography as described in detail [26]. In brief: supernatants from HaCaT cells, collected after 48 or 72 hours of incubation with SMases D or L. laeta venom, respectively, were run on a 10% polyacrylamide gel containing 0.1% gelatin. After overnight incubation at 37°C in zymography buffer, gels were stained with Coomassie brilliant blue.
Electrophoresis and Western blot
Supernatants from the HaCaT cell cultures, collected after 48 or 72 hours of incubation with SMases D or L. laeta venom, respectively, were run on 12.5% SDS-PAGE, under non-reducing conditions and blotted onto nitrocellulose. After blocking with PBS/5% BSA membranes were incubated with MoAb against human MMP7 (1 μg/mL), followed by incubation with GAM-IgG-AP (1:7500). Blots were developed using NBT/BCIP (Promega).
Quantification of MMP2, MMP7 and MMP9 in the supernatant of keratinocytes cell cultures
Total MMP2, MMP7 and MMP9 in supernatants samples, collected from HaCaT cell cultures, treated or not with SMases D or venom, were quantified using MAP Human MMP Magnetic Bead Panel 2 and read on Luminex (MAGPIX software).
Dermonecrosis and treatment with tetracycline
L. laeta venom (15 μg/mL in PBS) was injected intradermally (i.d.; 200 μl) in the shaved back of rabbits. Six hours later, tetracycline cream was administered topically twice daily over a 48 hour period on the lesion area. Negative control groups consisted of animals injected with PBS and treated with tetracycline cream, while positive controls consisted of animals injected with venom and treated with hydrosoluble lanolin (vehicle). Experiments were repeated twice using 3 animals per group. After 48 h, the animals were euthanized by intravenous overdose of anesthetic (ketamine) and skin sections were harvested for histological examination.
Histological analysis
Skin samples were prepared for histological analysis as previously described [16]. After staining with hematoxylin and eosin, sections were examined for the presence of epithelial necrosis, epithelial slough, dermal infiltrates, hemorrhage and level of collagen dissociation in the dermis and skin muscle fiber degeneration.
Statistical analysis
ANOVA followed by Bonferroni test was used for statistical analysis. P value < 0.05 was considered significant.
Results
Loxosceles laeta venom and Class 1 SMase D induce human keratinocyte cell death
We previously reported that L. intermedia venom and it’s class 2 SMase D, induced cell death by apoptosis in the human keratinocyte cell line HaCaT [17]. In order to verify if Class 1 SMase D from Loxosceles laeta venom has the same ability, HaCaT cells were incubated with L. laeta venom or Class 1 SMase D during 24, 48 and 72 hours and the cell viability was analyzed. Fig 1 (panels A and B) shows that both, venom and Class 1 SMase D from L. laeta, induced loss of cell viability in a dose and time dependent manner. Class 1 SMase D was more potent at inducing loss of cell viability than whole L. laeta venom. At higher concentrations (100–200 μg/ml), the SMase I induced 100% of keratinocyte cell death.
Fig 1. Effect of L. laeta venom and Class 1 SMase D on human keratinocytes cell viability.
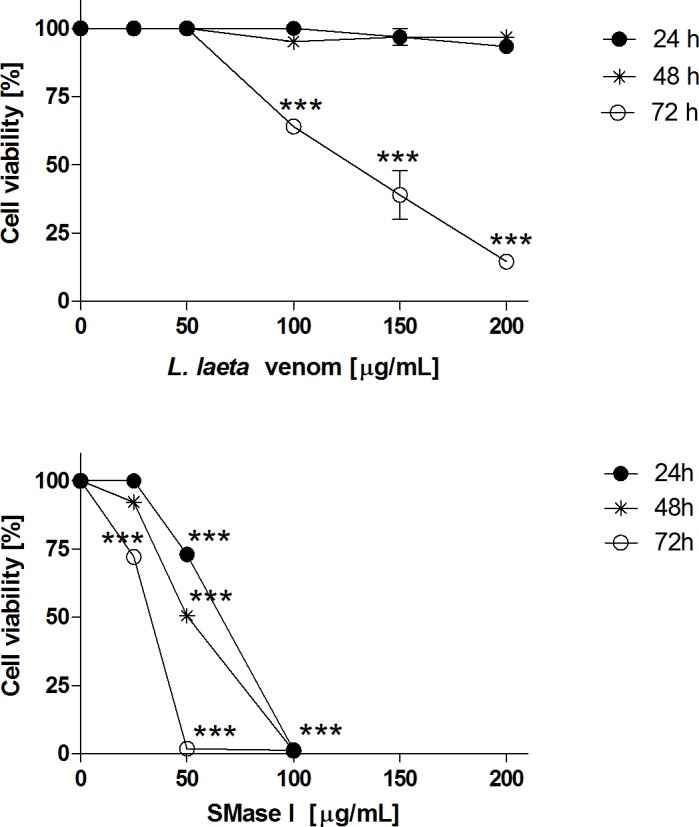
HaCaT cell cultures (2x104 cells) were incubated with increasing concentrations (25 to 200 μg/ml) of Loxosceles venom (A) or SMase I (B). After 24, 48 and 72 hours of treatment, cell viability was analyzed by MTT assay. Results are representative of three independent experiments and expressed as the mean of triplicates ± standard deviation. Significant differences (***) P<0.001 from control.
MMPs expressions in HaCaT cells supernatants
Since SMase I and L. laeta venom exhibit time and dose differences in cell toxicity, supernatants from HaCaT cells were investigated for MMPs expression after 72 hours of treatment with venom and 48 hours with Class 1 SMase D by gelatin zymography and western blot. Fig 2 (panels A and B) shows that venom or Class 1 SMase D from L. laeta increased expression of MMP2 (~56 kDa), as well as induced the expression of MMP9 (~80 kDa). This is similar to what we previously observed with L. intermedia venom and it’s Class 2 SMase D [17]. However, in addition a band with Mr around ~ 20 kDa was also observed in cells treated with L. laeta venom or Class 1 SMase D. In control cells, only pro-MMP2 was detected in the supernatants.
Fig 2. L. laeta venom and Class 1 SMase D induce the expression of matrix metaloproteinases in human keratinocytes.

Zymography analysis: HaCaT cell culture supernatants collected after 72 hours of the treatment with L. laeta venom (A) or 48 hours with the SMase D Class I (SMase I) (B) were run on gelatin containing 10% SDS-PAGE gels under non-reducing conditions. Control supernatants were harvested after 48 and 72 hours from cells incubated with medium plus saline. (C) Western blot analysis: HaCaT cell culture supernatants, collected after 24, 48 72 hours of the treatment with medium plus saline (C), venom or SMases D from Class 1 (SMase I from L. laeta) were run on 12.5% SDS-PAGE gels, blotted and developed using MoAbs against human MMP7. Figures are representative of three independent experiments.
To identify the lower molecular weight band that displayed gelatinolytic activity, western blotting was carried out. Pro-MMP7 has a Mr of about 21 kDa, similar to the Mr of the band observed. Western blot analysis showed that two bands with Mr around ~ 20 kDa were detected by the anti-MMP7 monoclonal antibody, suggesting that these bands may correspond to the pro- and active forms of MMP7 (Fig 2C). In the control supernatants, pro MMP7 and MMP7 were not detected.
Comparison of the gelatinolytic activity induced by SMases D Class 1 and Class 2
Previously we only detected the presence of MMP2 and MMP9 in the supernatants obtained from human keratinocytes treated with the L. intermedia SMases D (Class 2) [17]. In order to verify if the pro-MMP7 expression was only induced by Class 1 SMases D, supernatants collected from cells treated with 100 μg/ml of L. laeta venom, SMases D from Class 1 (SMase I from L. laeta) or Class 2 (P1 and P2 from L. intermedia venom) were compared by zymography and western blot analysis using a specific MMP7 monoclonal antibody. Fig 3 (panels A and B) shows that MMP7 was detected in the supernatants of cells treated with L. laeta Class 1 SMase D but not with L. intermedia Class 2 SMases D. However, using Luminex technology, basal levels of MMP7 were detected in the supernatants of HaCaT control cells. Only Class I SMase D induced an increase in expression of MMP7, while P1 and P2 SMases, seemed to reduce MMP7 (Fig 3C). The Luminex technology also corroborated the zymography results: while SMase D, from both classes induced significant increase in MMP9 secretion, only the Class I SMase D induced a significant increase in MMP2 expression.
Fig 3. Class 1 SMase D (SMase I from L. laeta) but not SMase Class 2 induced expression of MMP7 in human keratinocytes.

HaCaT cell culture supernatants, collected after 48 hours of the treatment with medium plus saline (Control) or 100 μg/mL of SMases D from Class 1 (SMase I from L. laeta) or Class 2 (P1 and P2 from L. intermedia), were subjected to zymography analysis (A), Western blotting (B) using MoAbs against human MMP7, and Luminex assay (C) for human MMP2, MMP9 and MMP7. Results are representative of three independent experiments and are expressed as the mean of triplicates ± standard deviation. Significant differences (*) P<0.05; (**) P<0.01 and (***) P<0.001 from control or (#) P<0.05; (##) P<0.01 and (###) P<0.001 from SMases D treated cells.
Tetracycline reduces expression of MMPs and protects HaCaT from cell death
We have previously shown that cell death through apoptosis induced by L. intermedia venom and Class 2 SMases D in HaCaT keratinocytes could be prevented by metalloproteinase inhibitors, such as tetracycline [17]. In order to verify if tetracycline was also effective at inhibiting biological effects of Class 1 SMaseD in L. laeta venom, HaCaT cells were incubated with L. laeta venom in the presence of various concentrations of tetracycline. Indeed, tetracycline dose dependently protected HaCaT cells from venom induced cell death and complete protection was obtained at 200 μg/mL tetracycline (Fig 4A). Tetracycline did not affect HaCaT cell viability of control cells.
Fig 4. Effect of tetracycline on the cell viability and the expression of MMPs in human keratinocytes treated with L. laeta venom.

HaCaT cell cultures were incubated with 100 μg/mL of L. laeta venom (V) or control medium (C) in the presence or absence of tetracycline (T). After 72 hours of treatment, cell viability was analyzed by MTT assay (A) and cell supernatants were analyzed by zymography (B), western blotting (C) and Luminex assay (tetracycline: 150 μg/mL) (D). Results are representative of three independent experiments and are expressed as the mean of triplicates ± standard deviation. Significant differences (*) P<0.05; (**) P<0.01 and (***) P<0.001 from control or (#) P<0.05; (##) P<0.01 and (###) P<0.001 from venom treated cells.
Effects on metalloproteinases expression/secretion by tetracycline were evaluated by zymography, western blotting and luminex technology. Fig 4 (panels B, C, D) shows that tetracycline abolished the conversion of secreted pro-MMP7 into active MMP7. Western blot showed that at higher concentrations tetracycline also reduced the secretion of pro-MMP7. In addition, tetracycline significantly inhibited MMP2 and MMP9 expression induced by L. laeta venom treatment.
Tetracyclines reduces the dermonecrotic lesion
The ability of tetracycline to inhibit L. laeta venom induced dermonecrosis was assessed by topical administration of tetracycline containing cream at the sites of venom injection (Fig 5). Control animals, injected with venom but not treated with tetracycline cream, developed within a few hours the typical loxoscelic lesions, characterised by edema, erythema and mild tenderness. 24 h later, necrosis with gravitational spread and scar had developed and the lesion size increased over 48 h. Topical administration of tetracycline cream twice daily over 48 hours significantly reduced the venom induced dermonecrotic lesion while vehicle only (lanolin) did not have an effect.
Fig 5. Effect of tetracyclines on dermonecrotic lesions.

(A) Adult rabbits were injected intradermally with 3 μg of venom from L. laeta spiders. After 6 h the animals were treated topically (black triangle) with a cream containing 5% tetracycline, twice a day during 48 hours. Control animals were inoculated with venom and either not treated (black circle) or treated with lanolin cream alone (black square) 6 hours after venom injection. Results are representative for two independent experiments and are expressed as the mean of triplicates ± SD. The asterisks indicate values statistically different (***) P<0.001 from the controls, i.e., non-treated or lanolin cream treated venom injected animals.
Histological analysis of skin lesion
Histopathological analysis, of rabbit skin 48 h after intradermal L. laeta venom injection, showed a normal epidermis, dissociation of the collagen fibers due to the edema and deep intradermal intense neutrophil infiltration (Fig 6B), degeneration of muscle fibers, and discrete neutrophil infiltration in the muscle layer. Topical treatment with tetracycline after venom injection largely prevented edema and neutrophil infiltration and neutrophil invasion into the muscle layer (Fig 6D). Lanolin (vehicle) treatment did not prevent the edema and neutrophil infiltration, although the latter was more dispersed, possibly caused by the physical action of the massaging of the lanolin in the affected area. (Fig 6C). Skins of rabbits subjected to the venom plus tetracycline treatment were histologically similar to that of PBS-inoculated animals (Fig 6, panels A and D) and far lower infiltration of neutrophils than in the venom injected animals was observed (Fig 6, panels B and D).
Fig 6. Histological analysis of the dermonecrotic lesion induced by L. laeta venom after tetracycline treatment.

Rabbits were injected with 3 μg/mL of L. laeta venom and treated, twice a day during 48 hours, with tetracycline containing cream. Control sites were injected with an equal volume of PBS. Panels correspond to the panoramic view of skin sections from rabbits injected with PBS (A), L. laeta venom (B), L. laeta venom and treated with lanolin cream vehicle (C), L. laeta venom and treated with lanolin cream containing tetracycline (D). Arrows indicate areas of leukocyte infiltration. Panels 1–2 show details of collagenous area of the dermis of the same sections. Black bars at the top of each panel indicate 100 μm.
Discussion
Dermonecrosis is a well-documented consequence of envenomation by Loxosceles spiders in humans. SMase D is the principle enzyme in Loxosceles venom responsible for the major pathological effects. In human keratinocytes Loxosceles intermedia Class 2 SMases D induced the expression of matrix metalloproteinases-2 and 9 (MMP2, MMP9), concomitant with the cutaneous loxoscelism lesion, as we previously reported [17].
In Brazil, medically important species of Loxosceles include L. laeta and L. intermedia, L. laeta being the most potent in inducing dermonecrosis [14]. We show here that L. laeta venom and it’s recombinant Class 1 SMase D, in addition to an increased expression of MMP2 and -9 as previously shown for Class 2 SMase D, also induced the expression of a molecule with gelatinolytic activity with Mr around 20 kDa in the HaCaT keratinocytic cell line (Figs 2 and 3). Using western blotting and MMP7 specific Luminex assay, we identified this molecule as MMP7/Matrilysin-1. The increase of MMPs expression/secretion coincided with a reduction of cell viability. The increase in expression of MMP2 and induction of expression/activation of MMP7 and MMP9 was inhibited by tetracycline. While tetracycline only slightly inhibited the induction of pro-MMP7, it largely prevented the conversion into mature MMP7. In vivo, tetracycline inhibited the development of the lesion and enhances recovery. Concomitantly, it reduced the tissue destruction and the influx of neutrophils.
SMases D displays a typical TIM (α/β)8-barrel fold and its active-site cleft is surrounded by the catalytic loop, variable loop, flexible loop and other short hydrophobic loops. While Class 1 SMases D contain one disulphide bridge, SMases D Class 2 contain an additional disulphide bridge which connects the catalytic loop to flexible loop, diminishes the active-site volume and also alters the inherent flexibility exhibited by the flexible loop [10–12]. This difference may explain the differences in potency of the L. laeta class 1 SMases D (Class I) and the L. intermedia (class 2) SMases D, resulting in a high expression of MMP7, MMP2 and MMP9, compared to class 1 SMases D that induced MMP9 and not MMP7.
The observation that SMase D can induce the high expression of MMP7 is novel. MMP7/matrilysin, secreted as a 28 kDa proenzyme and becomes activated after enzymatic cleavage of its N-terminal 9-kDa prodomain [27, 28]. MMP7 belongs to the group of matrilysins, which also includes MMP-26. Matrilysins are responsible for degrading extracellular matrix components and MMP7 substrates include elastin, proteoglycans, type IV collagen, fibronectin, and nidogen [29–31]. MMP7 expression in keratinocytes is increased in pro-inflammatory conditions e.g. such as caused by tissue trauma resulting in acute and chronic wounds [32], autoimmune diseases such as lupus [33] and by ultraviolet light in HaCaT cells [34] and is also increased in skin carcinomas [35, 36].
Non-healing chronic wounds are characterised by uncontrolled proteolytic tissue destruction in which MMPs play an important role. In addition to inhibition of MMP expression, tetracycline also inhibits the activation from pro-MMPs to mature MMPs, as well as the enzymatic activity of MMPs [37]. Previously, we have shown that the increased expression of MMP2, MMP9, keratinocyte cell death and the in vivo dermonecrotic lesion formation induced by L. intermedia venom could be prevented by the use of tetracycline [17, 23]. We show here that tetracycline can also modulate the action of L. laeta venom in vitro and in vivo. And as shown here for the first time, this inhibitor also prevents the expression/secretion and activation of MMP7 induced by L. laeta venom in HaCaT cells (Fig 4).
Interestingly, MMP7 has been reported to cleave pro-forms of MMP2, MMP9 [38] and activate ADAM28 [39]. Furthermore, Li and collaborators showed that MMP7 is needed for transepithelial neutrophil migration in acute lung injury. In its absence, neutrophils cannot cross the lung epithelium, and accumulate in the interstitial space between the capillaries and the epithelium, which demonstrates that MMP7 is needed for directed neutrophil migration in injury [40, 41]. Neutrophils are a major contributor to dermonecrosis after envenomation [42, 16, 23] and considering the importance of MMP7 for neutrophil migration, our observation that the class 1 SMase D containing L. laeta venom induced MMP7 high secretion, may explain why it is more potent than the Class 2 SMases D in inducing dermonecrosis. Furthermore, our observation that tetracycline inhibits the activation and expression of MMP7 (Fig 4) may explain our additional observation that tetracycline inhibits the infiltration of neutrophils after venom injection (Figs 5 and 6).
Recently, lesion formation induced by N. nomurai jellyfish envenomation was found also to be associated with increased MMP2 and MMP9 expression; this MMPs and lesion formation were also inhibited by tetracycline [43]. Although the substance responsible for induction of these lesions and MMP-expression have not been identified, our and Kang’s observations suggest that tetracycline therapy may be useful in the treatment of dermonecrotic lesions of various origins. In addition, due to its antimicrobial action, tetracycline treatment of cutaneous loxoscelism has the added advantage that it may prevent or treat concurrent wound infection.
In conclusion, we show here that Loxosceles laeta venom and its Class 1 SMase D induce and/or enhance the expression of matrix metalloproteinases (MMP2, -7 and -9) and cause cell death. The induction of MMP7 in high quantities, as a consequence of L. laeta venom and Class I SMase D action on HaCaT, may explain the enhanced severity of the envenomation induced by L. laeta compared to that of L. intermedia. Reduction of dermonecrotic lesion and inhibition of MMPs using tetracycline as shown here may be a suitable therapeutic intervention after envenomation by different Loxosceles species.
Data Availability
All relevant data are within the paper.
Funding Statement
This work was supported by funds from Fundação de Amparo à Pesquisa do Estado de São Paulo, Brazil (FAPESP), CeTICS Program FAPESP (2013/07467-1) and Conselho Nacional de Desenvolvimento Científico e Tecnológico, Brazil (CNPq). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Tambourgi DV, Goncalves-de-Andrade RM, van den Berg CW. Loxoscelism: From basic research to the proposal of new therapies. Toxicon: official journal of the International Society on Toxinology 2010; 56: 1113–1119. [DOI] [PubMed] [Google Scholar]
- 2.Futrell JM. Loxoscelism. The American journal of the medical sciences 1992; 304: 261–267. [DOI] [PubMed] [Google Scholar]
- 3.Atkins JA, Wingo CW, Sodeman WA, Flynn JE. Necrotic arachnidism. The American journal of tropical medicine and hygiene 1958; 7: 165–184. [DOI] [PubMed] [Google Scholar]
- 4.Wasserman GS, Anderson PC. Loxoscelism and necrotic arachnidism. Journal of toxicology Clinical toxicology 1983; 21: 451–472. [DOI] [PubMed] [Google Scholar]
- 5.Forrester LJ, Barrett JT, Campbell BJ. Red blood cell lysis induced by the venom of the brown recluse spider: the role of sphingomyelinase D. Archives of biochemistry and biophysics 1978; 187: 355–365. [DOI] [PubMed] [Google Scholar]
- 6.Kurpiewski GE, Forrester LJ, Campbell BJ, Barrett JT. Platelet aggregation and sphingomyelinase D activity of a purified toxin from the venom of Loxosceles reclusa. Biochimica et biophysica acta 1981; 678: 467–476. [DOI] [PubMed] [Google Scholar]
- 7.Tambourgi DV, Magnoli FC, van den Berg CW, Morgan BP, de Araujo PS, Alves EW, et al. Sphingomyelinases in the venom of the spider Loxosceles intermedia are responsible for both dermonecrosis and complement-dependent hemolysis. Biochemical and biophysical research communications 1998; 251: 366–373. [DOI] [PubMed] [Google Scholar]
- 8.van Meeteren LA, Frederiks F, Giepmans BN, Fernandes-Pedrosa MF, Billington SJ, Jost BH, et al. Spider and bacterial sphingomyelinases D target cellular lysophosphatidic acid receptors by hydrolyzing lysophosphatidylcholine. The Journal of biological chemistry 2004; 279: 10833–10836. [DOI] [PubMed] [Google Scholar]
- 9.Rivera IG, Ordoñez M, Presa N, Gomez-Larrauri A, Simón J, Trueba M, et al. Sphingomyelinase D/ceramide 1-phosphate in cell survival and inflammation. Toxins 2015; 7: 1457–1466. 10.3390/toxins7051457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pedroso A, Matioli SR, Murakami MT, Pidde-Queiroz G, Tambourgi DV. Adaptive evolution in the toxicity of a spider's venom enzymes. BMC evolutionary biology 2015;15: 290 10.1186/s12862-015-0561-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Murakami MT, Fernandes-Pedrosa MF, Tambourgi DV, Arni RK. Structural basis for metal ion coordination and the catalytic mechanism of sphingomyelinases D. The Journal of biological chemistry 2005; 280: 13658–13664. [DOI] [PubMed] [Google Scholar]
- 12.Murakami MT, Fernandes-Pedrosa MF, de Andrade SA, Gabdoulkhakov A, Betzel C, Tambourgi DV, et al. Structural insights into the catalytic mechanism of sphingomyelinases D and evolutionary relationship to glycerophosphodiester phosphodiesterases. Biochemical and biophysical research communications 2006; 342: 323–329. [DOI] [PubMed] [Google Scholar]
- 13.Tambourgi DV, Fernandes Pedrosa MF, van den Berg CW, Gonçalves-de-Andrade RM, Ferracini M, Paixão-Cavalcante D, et al. Molecular cloning, expression, function and immunoreactivities of members of a gene family of sphingomyelinases from Loxosceles venom glands. Molecular immunology 2004; 41: 831–840. [DOI] [PubMed] [Google Scholar]
- 14.de Oliveira KC, Gonçalves-de-Andrade RM, Piazza RM, Ferreira JM, van den Berg CW, Tambourgi DV. Variations in Loxosceles spider venom composition and toxicity contribute to the severity of envenomation. Toxicon: official journal of the International Society on Toxinology 2005; 45: 421–429. [DOI] [PubMed] [Google Scholar]
- 15.de Santi-Ferrara GI, Fernandes-Pedrosa MF, Junqueira-de-Azevedo IL, Gonçalves-de-Andrade RM, Portaro FC, Manzoni-de-Almeida D, et al. SMase II, a new sphingomyelinase D from Loxosceles laeta venom gland: molecular cloning, expression, function and structural analysis. Toxicon: official journal of the International Society on Toxinology 2009; 53: 743–753. [DOI] [PubMed] [Google Scholar]
- 16.Tambourgi DV, Paixão-Cavalcante D, Gonçalves-de-Andrade RM, Fernandes-Pedrosa MF, Magnoli FC, Morgan BP, et al. Loxosceles sphingomyelinase induces complement-dependent dermonecrosis, neutrophil infiltration, and endogenous gelatinase expression. The Journal of investigative dermatology 2005; 124: 725–731. [DOI] [PubMed] [Google Scholar]
- 17.Paixão-Cavalcante D, van den Berg CW, Fernandes-Pedrosa MF, Gonçalves-de-Andrade RM, Tambourgi DV. Role of matrix metalloproteinases in HaCaT keratinocytes apoptosis induced by Loxosceles venom sphingomyelinase D. The Journal of investigative dermatology 2006; 126: 61–68. [DOI] [PubMed] [Google Scholar]
- 18.Egeblad M, Werb Z. New functions for the matrix metalloproteinases in cancer progression. Nature reviews Cancer 2002; 2: 161–174. [DOI] [PubMed] [Google Scholar]
- 19.Golub LM, Ramamurthy NS, McNamara TF, Greenwald RA, Rifkin BR. Tetracyclines inhibit connective tissue breakdown: new therapeutic implications for an old family of drugs. Critical reviews in oral biology and medicine: an official publication of the American Association of Oral Biologists 1991; 2: 297–321. [DOI] [PubMed] [Google Scholar]
- 20.Nieman GF, Zerler BR. A role for the anti-inflammatory properties of tetracyclines in the prevention of acute lung injury. Current medicinal chemistry 2001; 8: 317–325. [DOI] [PubMed] [Google Scholar]
- 21.Golub LM, Lee HM, Ryan ME, Giannobile WV, Payne J, Sorsa T. Tetracyclines inhibit connective tissue breakdown by multiple non-antimicrobial mechanisms. Advances in dental research 1998; 12: 12–26. [DOI] [PubMed] [Google Scholar]
- 22.Golub LM. Introduction and background. Pharmacological research: the official journal of the Italian Pharmacological Society 2011; 63: 99–101. [DOI] [PubMed] [Google Scholar]
- 23.Paixão-Cavalcante D, van den Berg CW, Gonçalves-de-Andrade RM, Fernandes-Pedrosa MF, Okamoto CK, Tambourgi DV. Tetracycline protects against dermonecrosis induced by Loxosceles spider venom. The Journal of investigative dermatology 2007; 127: 1410–1418. [DOI] [PubMed] [Google Scholar]
- 24.Bucherl W. Biology and venoms of the most important South American spiders of the genera Phoneutria, Loxosceles, Lycosa, and Latrodectus. American zoologist 1969; 9: 157–159. [DOI] [PubMed] [Google Scholar]
- 25.Fernandes Pedrosa MF, Junqueira de Azevedo IL, Gonçalves-de-Andrade RM, van den Berg CW, Ramos CR, Ho PL, et al. Molecular cloning and expression of a functional dermonecrotic and haemolytic factor from Loxosceles laeta venom. Biochemical and biophysical research communications 2002; 298: 638–645. [DOI] [PubMed] [Google Scholar]
- 26.Kleiner DE, Stetler-Stevenson WG. Quantitative zymography: detection of picogram quantities of gelatinases. Analytical biochemistry 1994; 218: 325–329. [DOI] [PubMed] [Google Scholar]
- 27.Burke B. The role of matrix metalloproteinase 7 in innate immunity. Immunobiology 2004; 209: 51–56. [DOI] [PubMed] [Google Scholar]
- 28.Ii M, Yamamoto H, Adachi Y, Maruyama Y, Shinomura Y. Role of matrix metalloproteinase-7 (matrilysin) in human cancer invasion, apoptosis, growth, and angiogenesis. Experimental biology and medicine 2006; 231: 20–27. [DOI] [PubMed] [Google Scholar]
- 29.Murphy G, Cockett MI, Ward RV, Docherty AJ. Matrix metalloproteinase degradation of elastin, type IV collagen and proteoglycan. A quantitative comparison of the activities of 95 kDa and 72 kDa gelatinases, stromelysins-1 and -2 and punctuated metalloproteinase (PUMP). The Biochemical journal 1991; 277 (Pt 1): 277–279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sires UI, Griffin GL, Broekelmann TJ, Mecham RP, Murphy G, Chung AE, et al. Degradation of entactin by matrix metalloproteinases. Susceptibility to matrilysin and identification of cleavage sites. The Journal of biological chemistry 1993; 268: 2069–2074. [PubMed] [Google Scholar]
- 31.Halpert I, Sires UI, Roby JD, Potter-Perigo S, Wight TN, Shapiro SD, et al. Matrilysin is expressed by lipid-laden macrophages at sites of potential rupture in atherosclerotic lesions and localizes to areas of versican deposition, a proteoglycan substrate for the enzyme. Proceedings of the National Academy of Sciences of the United States of America 1996; 93: 9748–9753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Vaalamo M, Weckroth M, Puolakkainen P, Kere J, Saarinen P, Lauharanta J, et al. Patterns of matrix metalloproteinase and TIMP-1 expression in chronic and normally healing human cutaneous wounds. The British journal of dermatology 1996; 135: 52–59. [PubMed] [Google Scholar]
- 33.Järvinen TM, Kanninen P, Jeskanen L, Koskenmies S, Panelius J, Hasan T, et al. Matrix metalloproteinases as mediators of tissue injury in different forms of cutaneous lupus erythematosus. The British journal of dermatology 2007; 157: 970–980. [DOI] [PubMed] [Google Scholar]
- 34.Skiba B, Neill B, Piva TJ. Gene expression profiles of TNF-alpha, TACE, furin, IL-1beta and matrilysin in UVA- and UVB-irradiated HaCat cells. Photodermatology, photoimmunology & photomedicine 2005; 21: 173–182. [DOI] [PubMed] [Google Scholar]
- 35.Hartmann-Petersen S, Tammi RH, Tammi MI, Kosma VM. Depletion of cell surface CD44 in nonmelanoma skin tumours is associated with increased expression of matrix metalloproteinase 7. The British journal of dermatology 2009; 160: 1251–1257. 10.1111/j.1365-2133.2009.09031.x [DOI] [PubMed] [Google Scholar]
- 36.Mitsui H, Suárez-Fariñas M, Gulati N, Shah KR, Cannizzaro MV, Coats I, et al. Gene expression profiling of the leading edge of cutaneous squamous cell carcinoma: IL-24-driven MMP-7. The Journal of investigative dermatology 2014; 134: 1418–1427. 10.1038/jid.2013.494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hanemaaijer R, Visser H, Koolwijk P, Sorsa T, Salo T, Golub LM, et al. Inhibition of MMP synthesis by doxycycline and chemically modified tetracyclines (CMTs) in human endothelial cells. Advances in dental research 1998; 12: 114–118. [DOI] [PubMed] [Google Scholar]
- 38.Wilson CL, Matrisian LM. Matrilysin: an epithelial matrix metalloproteinase with potentially novel functions. The international journal of biochemistry & cell biology 1996; 28: 123–136. [DOI] [PubMed] [Google Scholar]
- 39.Mochizuki S, Shimoda M, Shiomi T, Fujii Y, Okada Y. ADAM28 is activated by MMP-7 (matrilysin-1) and cleaves insulin-like growth factor binding protein-3. Biochemical and biophysical research communications 2004: 315: 79–84. [DOI] [PubMed] [Google Scholar]
- 40.Li Q, Park PW, Wilson CL, Parks WC. Matrilysin shedding of syndecan-1 regulates chemokine mobilization and transepithelial efflux of neutrophils in acute lung injury. Cell 2002; 111: 635–646. [DOI] [PubMed] [Google Scholar]
- 41.Shapiro SD. Immunology: Mobilizing the army. Nature 2003; 421: 223–224. [DOI] [PubMed] [Google Scholar]
- 42.Smith CW, Micks DW. The role of polymorphonuclear leukocytes in the lesion caused by the venom of the brown spider, Loxosceles reclusa. Laboratory investigation; a journal of technical methods and pathology 1970; 22: 90–93. [PubMed] [Google Scholar]
- 43.Kang C, Jin YB, Kwak J, Jung H, Yoon WD, Yoon TJ, et al. Protective effect of tetracycline against dermal toxicity induced by Jellyfish venom. PloS one 2013; 8: e57658 10.1371/journal.pone.0057658 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.