

Abstract
Cyclooxygenase‐2 (COX‐2) and its downstream product prostaglandin E2 (PGE2) play a key role in generation of the inflammatory microenvironment in tumor tissues. Gastric cancer is closely associated with Helicobacter pylori infection, which stimulates innate immune responses through Toll‐like receptors (TLRs), inducing COX‐2/PGE2 pathway through nuclear factor‐κB activation. A pathway analysis of human gastric cancer shows that both the COX‐2 pathway and Wnt/β‐catenin signaling are significantly activated in tubular‐type gastric cancer, and basal levels of these pathways are also increased in other types of gastric cancer. Expression of interleukin‐11, chemokine (C‐X‐C motif) ligand 1 (CXCL1), CXCL2, and CXCL5, which play tumor‐promoting roles through a variety of mechanisms, is induced in a COX‐2/PGE2 pathway‐dependent manner in both human and mouse gastric tumors. Moreover, the COX‐2/PGE2 pathway plays an important role in the maintenance of stemness with expression of stem cell markers, including CD44, Prom1, and Sox9, which are induced in both gastritis and gastric tumors through a COX‐2/PGE2‐dependent mechanism. In contrast, disruption of Myd88 results in suppression of the inflammatory microenvironment in gastric tumors even when the COX‐2/PGE2 pathway is activated, indicating that the interplay of the COX‐2/PGE2 and TLR/MyD88 pathways is needed for inflammatory response in tumor tissues. Furthermore, TLR2/MyD88 signaling plays a role in maintenance of stemness in normal stem cells as well as gastric tumor cells. Accordingly, these results suggest that targeting the COX‐2/PGE2 pathway together with TLR/MyD88 signaling, which would suppress the inflammatory microenvironment and maintenance of stemness, could be an effective preventive or therapeutic strategy for gastric cancer.
Keywords: Gastric cancer, inflammation, innate immunity, microenvironment, stemness
Epidemiological studies have indicated that regular use of non‐steroidal anti‐inflammatory drugs is associated with a reduced risk of gastrointestinal cancer, suggesting that inflammation plays a role in cancer development.1, 2 Target molecules of non‐steroidal anti‐inflammatory drugs are COX‐1 and COX‐2, which catalyze prostaglandin biosynthesis. Genetic studies have indicated that COX‐2 and its downstream product PGE2 play a key role in gastrointestinal tumorigenesis.3, 4, 5 In cancer tissues, macrophages infiltrate and are activated with the induction of the COX‐2/PGE2 pathway, which further induces cytokine signaling.6, 7 The transcription factors NF‐κB and Stat3 are activated by inflammatory cytokines, which plays an important role in tumorigenesis.8, 9, 10 Based on these results, it has been established that inflammation promotes cancer development through induction of the COX‐2/PGE2 pathway and activation of NF‐κB and Stat3.11, 12, 13 Innate immune responses through the TLRs have also been shown to be involved in tumor promotion.14 We have shown that the COX‐2/PGE2 pathway triggers generation of inflammatory microenvironment in the stomach in a mouse gastric tumor model,5, 15 and that TLR signaling through the adaptor molecule MyD88 is required for COX‐2/PGE2‐induced gastritis and gastric tumor development.16 Accordingly, it is possible that the interplay of the COX‐2/PGE2 and TLR/MyD88 pathways is important for generation of the inflammatory microenvironment, which promotes tumorigenesis.
In this review, we define the phenotypes of our gastric tumor model (Gan mice that develop COX‐2/PGE2 pathway‐dependent gastric tumors) by comparing them to human gastric cancers and discuss the role of COX‐2/PGE2‐associated inflammation in gastric tumorigenesis and its connection to the TLR/MyD88 pathways.
Gan mice recapitulate human tubular‐type gastric cancer
Cyclooxygenase‐2 expression is induced in most cancers, and the role of the COX‐2/PGE2 pathway was first examined in intestinal tumors.12 Using Apc Δ716 mice, a model of familial adenomatous polyposis, we have shown that COX‐2/PGE2 signaling is required for intestinal tumorigenesis and that treatment with a COX‐2 inhibitor significantly suppressed intestinal tumorigenesis.3, 17, 18, 19 Thus far, a number of mechanisms of PGE2 signaling have been reported to be involved in tumor promotion, including angiogenesis,20 tumor cell survival,21 silencing of tumor suppressors by DNA methylation,22 and recruitment of MDSCs.23
Induction of COX‐2 expression is found in more than 90% of gastric cancers,24 and Helicobacter pylori infection is known to be a major cause of COX‐2 induction.25 To examine the role of the COX‐2/PGE2 pathway in the stomach, we constructed K19‐C2mE mice that express Ptgs2 and Ptges, encoding COX‐2 and microsomal prostaglandin E synthase‐1, respectively, using the Krt19 gene promoter (Data S1). The PGE2 level is significantly increased in the gastric mucosa of K19‐C2mE mice, which develop gastritis with macrophage infiltration, indicating that the COX‐2/PGE2 pathway triggers inflammatory response.15, 26
Inflammation promotes tumorigenesis in cooperation with oncogenic activation, and accumulation of β‐catenin, a hallmark of Wnt signaling activation, is found in more than 50% of gastric cancers,5 although genetic alterations in APC and the β‐catenin gene (CTNNB1) were found only in 24/223 (10.8%) and 6/223 (2.7%) of gastric cancers, respectively.27 It is possible that Wnt signaling is activated in gastric cancer also by mechanisms other than APC or CTNNB1 mutations. We thus constructed K19‐Wnt1 transgenic mice that express Wnt1, a ligand for canonical Wnt signaling, in gastric epithelial cells. Notably, K19‐Wnt1 mice develop small dysplastic lesions in the gastric mucosa; however, simultaneous activation of Wnt signaling and the COX‐2/PGE2 pathway in compound K19‐Wnt1 and K19‐C2mE mice (Gan mice) results in the development of inflammation‐associated gastric tumors.5, 15 Histologically, Gan mouse tumors consist of a glandular structure with irregular branching, and are similar to that of human tubular‐type gastric cancer (Fig. 1a, Data S1). In Gan mouse tumors, F4/80 positive macrophages and a small number of lymphocytes infiltrate the tumor stroma. Accordingly, it is possible that the COX‐2/PGE2 pathway induces the inflammatory response, but is not sufficient to induce acquired immune responses. Notably, depletion of macrophages in Gan mouse tumors caused apoptosis of tumor cells, indicating a role of macrophages in tumor cell survival.28
Figure 1.
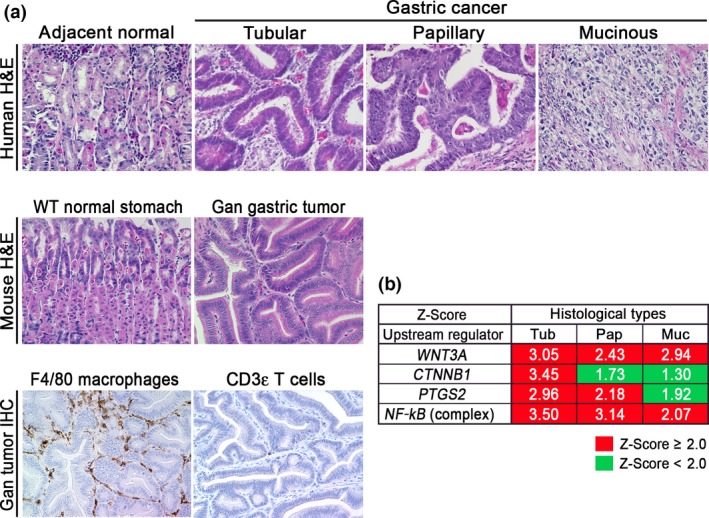
Classification of Gan mouse gastric tumors. (a) Representative histological photographs (H&E) of human gastric cancer (top) and Gan mouse gastric tumor (middle), and immunohistochemistry (IHC) for macrophages (F4/80) and T cells (CD3ε) of Gan mouse tumors (bottom). (b) Ingenuity pathway analysis of the indicated pathways in the respective histological types of human gastric cancers. Z‐scores > 2.0 were considered as significantly activated pathways.
The results of a comprehensive molecular evaluation of human gastric cancers have been reported in the TCGA project.29 Moreover, the Asian Cancer Research Group developed a novel classification of clinically relevant molecular subtypes of gastric cancer.27 In the present study, we examined the signaling pathways that are activated in the human gastric cancer subtypes using the TCGA database. We extracted expression data from the respective histological types (tubular‐type, papillary‐type, and mucinous‐type gastric cancer) and carried out an Ingenuity pathway analysis (Fig. 1b). Notably, both of the canonical Wnt signaling pathways (WNT3 and CTNNB1) were significantly activated in tubular‐type gastric cancer. In contrast, NF‐κB activation was found in all types of gastric cancer, indicating inflammatory responses in all gastric cancer types. Moreover, the PTGS2 pathway was significantly activated in tubular‐type and papillary‐type gastric cancer. Accordingly, Wnt signaling and the COX‐2/PGE2 pathways are simultaneously activated in tubular‐type gastric cancer, although the basal levels of these pathways are increased in all types compared with normal stomach.
Based on the histological characteristics and expression profiles, we concluded that Gan mice recapitulate human tubular‐type gastric cancer. That is, the combined activation of Wnt signaling and the COX‐2/PGE2 pathway can cause tubular‐type gastric cancer. This finding is consistent with our previous microarray analyses, showing that expression profiles of Gan mouse tumors are similar to intestinal‐type gastric cancer (Lauren's classification).30
Roles of COX‐2/PGE2 pathway‐associated inflammation in stemness
Recently, PGE2 has been revealed to be involved in the maintenance of stemness. Treatment of irradiated mice with PGE2 results in increased survival of hematopoietic stem cells,31 and PGE2 signaling regulates hematopoietic stem cell self‐renewal through direct interaction with Wnt signaling by cAMP/protein kinase A activation.32, 33 The PGE2/Wnt signaling interaction also regulates regeneration of other organs including liver.32 More recently, it has been shown that PGE2 signaling through the EP4 receptor induces expansion of CD133+ CD44+ cancer stem cell populations in intestinal tumors through the activation of phosphatidylinositol 3‐kinase and MAPK signaling.34
We therefore examined the COX‐2/PGE2 pathway‐dependent upregulated genes in mouse gastritis and tumors using the RNA sequencing results (GSE70135) (Data S1).16 A principal component analysis indicated the similarity of the expression profile of Gan mouse tumors to that of K19‐C2mE gastritis (Fig. 2a), in both of which COX‐2/PGE2 pathway is induced. A clustering analysis revealed that the genes encoding stem cell‐related factors including CD44, Sox2, Noxo1, Cd14, Ephb2, Cxcr4, Sox9, and Prom1, are upregulated in both gastritis and gastric tumors compared to normal stomach (Fig. 2b,c). These genes are also upregulated in Lgr5+VE normal gastric stem cells.35 Moreover, it has been shown recently that Cxcr4‐expressing innate lymphoid cells construct perivascular niche for Mist1‐expressing gastric stem cells.36 On the other hand, Kcne2, Trim50, and Hrh2 were significantly downregulated in both gastritis and gastric tumors. These molecules are involved in acid secretion from differentiated epithelia. Accordingly, it is possible that the COX‐2/PGE2 pathway promotes gastric tumorigenesis through maintenance of stemness by cell‐intrinsic mechanisms or by stem cell niche construction.
Figure 2.
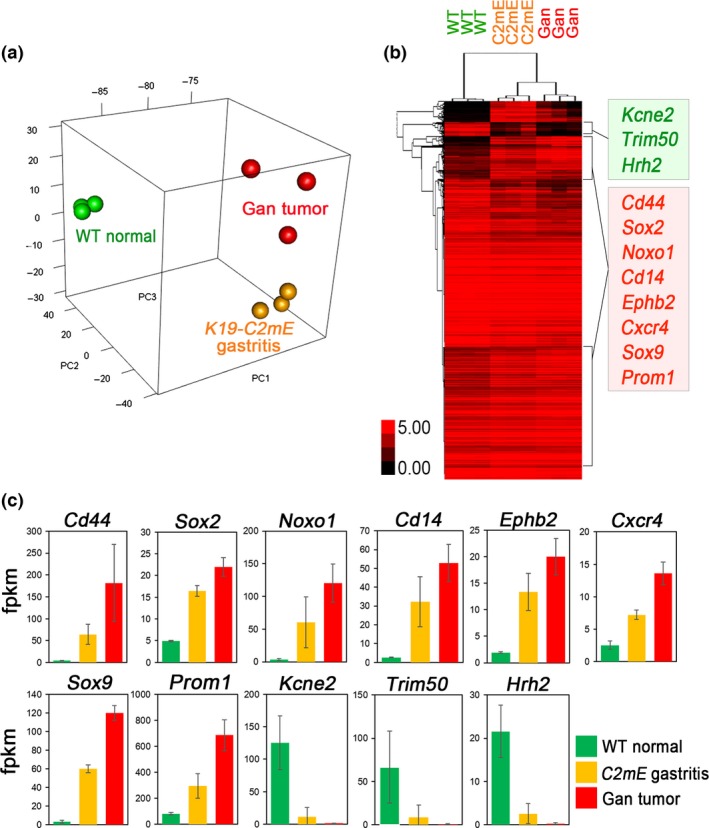
Expression analyses of mouse gastritis and gastric tumors. (a) Principal component analysis of Gan mouse gastric tumors (red), K19‐C2mE gastritis (orange), and wild‐type normal stomach (green). (b) Average‐linkage clustering analysis of genotype‐specific genes using RNA sequencing results of the wild‐type normal stomach (WT), K19‐C2mE mouse gastritis (C2mE), and Gan mouse gastric tumors (Gan). Selected genes that were upregulated (red) or downregulated (green) in both gastritis and gastric tumors are listed (right). (c) Expression levels of the selected genes in (b) are shown in bar graphs (mean ± SD). fpkm, fragments per kilobase of transcript per million fragments mapped.
It has been shown that TNF‐α, IL‐6, and CXCL12 signals activate each other, inducing the cytokine network in tumor tissues.37 Accordingly, inhibition of TNF‐α may disturb the cytokine network in tumor microenvironment. Importantly, disruption of the TNF‐α gene (Tnf) in BMDCs of Gan mice significantly suppressed gastric tumorigenesis, and expression levels of stem cell markers (CD44, Prom1, Sox9, and Noxo1) were significantly downregulated.38 Accordingly, it is possible that the TNF‐α cytokine network, which is activated by the COX‐2/PGE2 pathway, plays an important role in maintenance of tumor cell stemness.
Among the TNF‐α‐dependent upregulated genes in tumor cells, we noticed that Noxo1 expression is required for the soft‐agar colony formation of gastric cancer cells.38 Noxo1 is a component of the NOX1 complex, and plays a role in ROS production.39 NOX1 is involved in tumor promotion through inactivation of phosphatase and tensin homolog and SHP2, and ablation of NOX1 expression resulted in growth arrest of Ras‐transformed cells.39, 40, 41 Moreover, it has been shown that NOX1/ROS signaling activates NF‐κB, which contributes to maintenance of stemness in intestinal tumor cells.42 Accordingly, it is possible that the NOX1/ROS pathway is one of the key mechanisms that link the COX‐2/PGE2‐dependent inflammatory microenvironment and cancer stem cell regulation (Fig. 3).
Figure 3.
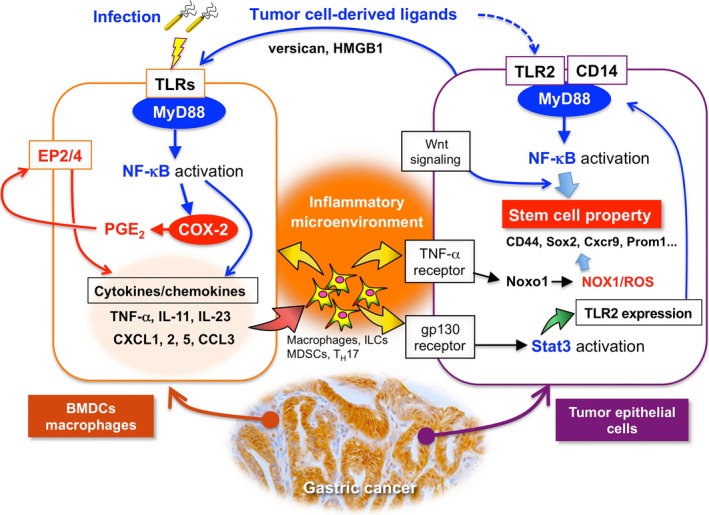
Schematic drawing of the interplay of the COX‐2/prostaglandin E2 (PGE 2) and Toll‐like receptor (TLR)/MyD88 pathways in bone marrow‐derived cells (BMDCs) for generation of inflammatory microenvironment (left), and the role of TLR2/MyD88 and nuclear factor‐κB (NF‐κB) together with the NADPH oxidase 1 (NOX1)/reactive oxygen species (ROS) pathway in tumor cells for maintenance of stemness and promotion of tumorigenesis (right). CCL, chemokine (C‐C motif) ligand; CXCL, chemokine (CXC motif) ligand; Cxcr, CXC chemokine receptor; HMGB1, high mobility group box 1; IL, interleukin; ILC, innate lymphoid cell; MDSC, myeloid‐derived suppressor cell; Stat, signal transducer and activator of transcription; TNF‐α, tumor necrosis factor‐α.
However, it has been shown that the mechanism by which a variant‐form CD44 protects against ROS is important for gastric tumorigenesis.43 Accordingly, it is conceivable that high ROS levels are detrimental to tumor cells and that the antioxidant mechanism is important for tumorigenesis, while ROS signaling may also promote early‐stage tumorigenesis.44
“Infection‐associated” and “tumor‐elicited” inflammation
Approximately 20% of malignant cancers are associated with infection.45, 46 Gastric cancer, liver cancer, and cervical cancer are associated with H. pylori, hepatitis C virus, and human papilloma virus infection, respectively, and are included in this group. Helicobacter pylori promotes gastric cancer by injecting CagA into host cells, which binds cellular SHP2 or partitioning‐defective 1, resulting in disturbance of cell function.47 At the same time, H. pylori stimulates TLRs, leading to activation of innate immunity to induce “infection‐associated inflammation” and generate the inflammatory microenvironment.48
Chronic inflammation, conversely, is associated with most cancers that are not related to infection. In cancer tissues, tumor cell‐derived factors can also stimulate TLRs as damage‐associated molecular patterns to induce “tumor‐elicited inflammation”.49, 50
We therefore compared the expression profiles of cytokines and chemokines in human gastric cancer (TCGA database) with those of H. pylori infection‐associated human gastritis (GSE60662) (Table 1).51 Interestingly, expression patterns of inflammation‐related factors were not always consistent in gastritis and gastric cancer. For example, expression of IL‐8 and IL‐11 is predominantly induced in gastric cancer, whereas TNF‐α expression is mostly increased in gastritis. It is possible that “tumor‐elicited inflammation” is also induced in gastric cancer, which may result in cytokine profiles that differ from H. pylori infection‐induced gastritis. However, CXCL1, CXCL2, CXCL5, CCL3, CCL4, and TLR2 are upregulated in both gastritis and gastric cancer (Table 1). Notably, these genes are induced also in both K19‐C2mE gastritis and Gan gastric tumors. It is therefore possible that these inflammatory factors are induced in a COX‐2/PGE2 pathway‐dependent manner, regardless of the mechanisms that activate TLRs. Although IL‐11 is not significantly induced in human gastritis, it is upregulated in mouse gastritis, suggesting that it is induced through the COX‐2/PGE2 pathway. Importantly, these listed cytokines and chemokines have tumor‐promoting functions as follows.
Table 1.
Expression of cytokine and chemokines in human and mouse gastritis and gastric cancer tissues (log2 values of fold changes)
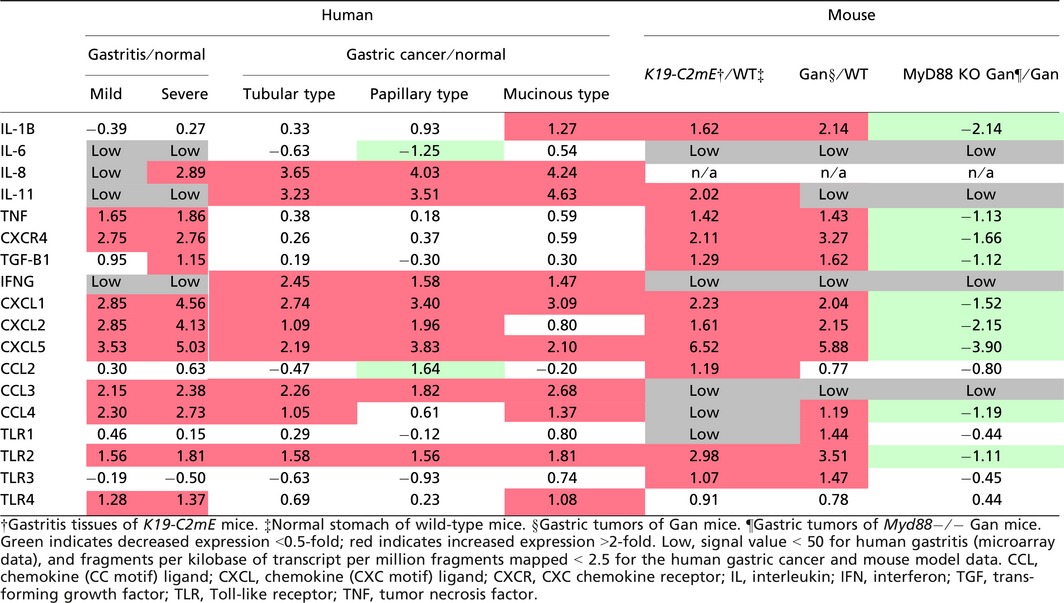
Interleukin‐11 signaling by the gp130 receptor promotes gastric tumorigenesis through Stat3 activation.23, 52 Moreover, PGE2 induces expression of CXCL1 and CXCL2, which further induces colitis‐associated colon cancer through recruitment of CXC chemokine receptor 2‐expressing MDSCs.23 A similar mechanism was shown for CXCL5, which recruits TGF‐β‐expressing MDSCs to tumor tissues, resulting in malignant progression by immune suppression and the M2‐type host reaction.53 Accordingly, both “infection‐associated” and “tumor‐elicited” inflammation may promote gastric tumorigenesis through induction of tumor‐promoting cytokines, and the COX‐2/PGE2 pathway is involved in this induction process.
Toll‐like receptor signaling in generation of inflammatory microenvironment
Inflammation is triggered when innate immune cells detect infection or tissue injury through TLRs.54 As described above, TLR/MyD88 signaling plays a role in the generation of the tumor inflammatory microenvironment.14 Moreover, disruption of the MyD88 gene (Myd88) in mouse models suppressed sporadic intestinal tumors,55, 56 colitis‐associated colon cancer,57, 58 gastric cancer,59 liver cancer,60 skin tumors, and sarcomas.61 Notably, disruption of Myd88 in Apc Min mice suppressed expression of COX‐2 and other cytokines.55 These results suggest that TLR/MyD88 signaling promotes tumorigenesis through induction of the COX‐2/PGE2 pathway, which is required for generation of the inflammatory microenvironment (Fig. 3).
Importantly, however, we recently found that disruption of Myd88 in the BMDCs of Gan mice significantly suppressed gastritis and gastric tumors,16 and that cytokine expression levels were dramatically decreased, even though the COX‐2/PGE2 pathway was stably activated (Table 1). Accordingly, it is possible that the cooperation of the COX‐2/PGE2 and TLR/MyD88 pathways is required for generation of the inflammatory microenvironment (Fig. 3).
In intestinal tumor tissues, epithelial barrier function is impaired, and thus microbial products can stimulate the TLRs of myeloid‐derived cells to express IL‐23, which further activates Th17 response.62 Consistently, expression of IL‐23 and IL‐17 is upregulated in Gan mouse tumors in a MyD88‐dependent manner.16 It has been shown that IL‐17 expression is required for inflammatory response and intestinal polyposis in Apc Min mice.63 Such a mechanism may activate the TLRs of BMDCs in gastric cancer tissues (Fig. 3).
“Cell‐intrinsic” role of TLR/MyD88 signaling in tumorigenesis
It has been shown that TLR2 and TLR4 signals through MyD88 are important for intestinal mucosal homeostasis and regeneration.64 Moreover, NF‐κB signaling in epithelial cells promotes inflammation‐associated intestinal tumorigenesis.8 These results suggest a “cell‐intrinsic” role of TLR/MyD88 signaling through NF‐κB activation in epithelial homeostasis and tumorigenesis.
Importantly, TLR2/MyD88 signaling in intestinal epithelial cells plays a critical role in expansion of the stem cell pool in damaged mucosa and is required for tumor growth in mouse models.65 Interestingly, TLR2 is the most upregulated TLR in gastric tumors in both human and mouse models (Table 1),66 and its expression is predominantly detected in tumor epithelial cells.16 Notably, disruption of the TLR2 gene (Tlr2) in a gp130F/F gastric tumor mouse model in which the hyperactivation of Stat3 causes gastric tumorigenesis,67 results in significant suppression of tumor formation without inhibition of inflammation.66 This suggests that TLR2/MyD88 signaling in tumor cells does not contribute to the inflammatory microenvironment but that it is important for tumorigenesis through the maintenance of stemness of tumor cells by a cell‐intrinsic mechanism (Fig. 3). Expression of TLR2 has consistently been reported to be associated with maintenance of cancer stem cells in mammary and ovarian cancer.68, 69
Expression of TLR2 is also induced in gastritis of both humans and mouse models, and its expression in Gan mouse tumors was significantly decreased by disruption of Myd88.16 Moreover, it has been shown that Stat3 regulates TLR2 expression in gastric cancer cells.66 It is therefore conceivable that TLR2 expression is induced in tumor cells by IL‐11 signaling through gp130/Stat3 pathway, which is activated in the inflammatory microenvironment of gastric tumors.
How does TLR2/MyD88 regulate stemness in tumor cells? It has been reported that activation of NF‐κB together with Wnt signaling promotes acquisition of stem cell properties, inducing dedifferentiation of intestinal epithelia.70 Accordingly, it is possible that activation of TLR2/MyD88 signaling through NF‐κB, together with Wnt signaling, increases self‐renewal activity of gastric tumor cells (Fig. 3). Activation of the NOX1/ROS pathway by TNF‐α and the cytokine network may also contribute to the cell‐intrinsic role of TLR2/MyD88 signaling in regulation of stemness as described above.
The ligands that are responsible for activation of TLR signaling in BMDCs and tumor epithelial cells is a matter of interest. In gastric cancer, H. pylori is an important ligand to activate TLRs, but other tumor cell‐derived factors, such as versican or high mobility group box 1 are other possible candidates for TLR activation as damage‐associated molecular patterns;71, 72 however, this remains to be investigated.
Conclusions
It has been established that the COX‐2/PGE2 pathway plays a tumor‐promoting role through generation of the inflammatory microenvironment. Maintenance of stemness is one of the important COX‐2/PGE2‐dependent tumor promotion mechanisms. It has also been shown that TLR/MyD88 signaling is important for tumorigenesis through activation of innate immunity, which also induces generation of the inflammatory microenvironment. As discussed in this review, the interplay of these two distinct pathways is required for generation of the inflammatory microenvironment, which has a critical role in tumor promotion. Moreover, it has been shown that the cell‐intrinsic role of epithelial TLR2/MyD88 signaling is important for tumor cell stemness and gastric tumorigenesis, and such innate immune responses in tumor cells are activated by the inflammatory microenvironment (Fig. 3). Accordingly, targeting the COX‐2/PGE2 pathway together with TLR/MyD88 signaling may suppress the inflammatory microenvironment as well as the stemness of gastric tumor cells and could be an effective preventive and therapeutic strategy against gastric cancer.
Disclosure Statement
The authors have no conflicts of interest.
Abbreviations
- BMDC
bone marrow‐derived cell
- CCL
chemokine (CC motif) ligand
- COX
cyclooxygenase
- CXCL
chemokine (CXC motif) ligand
- IL
interleukin
- MDSC
myeloid‐derived suppressor cell
- NOX1
NADPH oxidase 1
- NF‐κB
nuclear factor‐κB
- PG
prostaglandin
- PGE2
prostaglandin E2
- ROS
reactive oxygen species
- Stat
signal transducer and activator of transcription
- TCGA
The Cancer Genome Atlas
- TNF‐α
tumor necrosis factor‐α
- TLR
Toll‐like receptor
Supporting information
Data S1. Methods for construction of mouse models, histological analyses of mouse and human gastric tumors, and gene expression analyses (Ingenuity pathway analysis, principal component analysis, and clustering analysis) are described.
Acknowledgments
We thank Dr. Yoichi Yamada for discussions about bioinformatics, and Ms. Manami Watanabe and Ms. Ayako Tsuda for their excellent technical assistance in Gan mouse studies.
Cancer Sci 107 (2016) 391–397
Funding Information
Japan Agency for Medical Research and Development, (Grant/Award Number: ‘AMED‐CREST (Chronic Inflammation)’) Ministry of Education, Culture, Sports, Science, and Technology, (Grant/Award Number: ‘Grants‐in‐Aids for Young Scientists (15K18405)’)
References
- 1. Thun MJ, Namboodiri MM, Heath CW. Aspirin use and reduced risk of fatal colon cancer. N Engl J Med 1991; 325: 1593–6. [DOI] [PubMed] [Google Scholar]
- 2. Thun MJ, Jacobs EJ, Patrono C. The role of aspirin in cancer prevention. Nat Rev Clin Oncol 2012; 9: 259–67. [DOI] [PubMed] [Google Scholar]
- 3. Oshima M, Dinchuk JE, Kargman SL et al Suppression of intestinal polyposis in Apc Δ716 knockout mice by inhibition of cyclooxygenase 2 (COX‐2). Cell 1996; 87: 803–9. [DOI] [PubMed] [Google Scholar]
- 4. Chulada PC, Thompson MB, Mahler JF et al Genetic disruption of Ptgs‐1, as well as Ptgs‐2, reduces intestinal tumorigenesis in Min mice. Cancer Res 2000; 60: 4705–8. [PubMed] [Google Scholar]
- 5. Oshima H, Matsunaga A, Fujimura T et al Carcinogenesis in mouse stomach by simultaneous activation of the Wnt signaling and prostaglandin E2 pathway. Gastroenterology 2006; 131: 1086–95. [DOI] [PubMed] [Google Scholar]
- 6. Pollard JW. Tumour‐educated macrophages promotes tumour progression and metastasis. Nat Rev Cancer 2004; 4: 71–8. [DOI] [PubMed] [Google Scholar]
- 7. Kitamura T, Qian BZ, Pollard JW. Immune cell promotion of metastasis. Nat Rev Immunol 2015; 15: 73–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Greten FR, Eckmann L, Greten TF et al IKKβ links inflammation and tumorigenesis in a mouse model of colitis‐associated cancer. Cell 2004; 118: 285–96. [DOI] [PubMed] [Google Scholar]
- 9. Grivennikov S, Karin E, Terzic J et al IL‐6 and Stat3 are required for survival of intestinal epithelial cells and development of colitis‐associated cancer. Cancer Cell 2009; 15: 7103–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Bollrath J, Phesse TJ, von Burstin VA et al gp130‐mediated Stat3 activation in enterocytes regulates cell survival and cell‐cycle progression during colitis‐associated tumorigenesis. Cancer Cell 2009; 15: 91–102. [DOI] [PubMed] [Google Scholar]
- 11. Coussens LM, Werb Z. Inflammation and cancer. Nature 2002; 420: 860–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Wang D, DuBois RN. The role of COX‐2 in intestinal inflammation and colorectal cancer. Oncogene 2010; 29: 781–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Grivennikov SI, Greten FR, Karin M. Immunity, inflammation, and cancer. Cell 2010; 140: 883–99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Pradere JP, Dapito DH, Schwabe RF. The Yin and Yang of Toll‐like receptors in cancer. Oncogene 2014; 33: 3485–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Oshima H, Oguma K, Du YC et al Prostaglandin E2, Wnt and BMP in gastric tumor mouse models. Cancer Sci 2009; 100: 1779–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Maeda Y, Echizen K, Oshima H et al Myeloid differentiation factor 88 signaling in bone marrow‐derived cells promotes tumorigenesis by generation of inflammatory microenvironment. Cancer Prev Res 2016; 9: 253–63. [DOI] [PubMed] [Google Scholar]
- 17. Sonoshita M, Takaku K, Sasaki N et al Acceleration of intestinal polyposis through prostaglandin receptor EP2 in Apc Δ716 knockout mice. Nat Med 2001; 7: 1048–51. [DOI] [PubMed] [Google Scholar]
- 18. Oshima M, Murai N, Kargman S et al Chemoprevention of intestinal polyposis in the Apc Δ716 mouse by rofecoxib, a specific cyclooxygenase‐2 inhibitor. Cancer Res 2001; 61: 1733–40. [PubMed] [Google Scholar]
- 19. Oshima M, Taketo MM. COX selectivity and animal models for colon cancer. Curr Pharm Des 2002; 8: 1021–34. [DOI] [PubMed] [Google Scholar]
- 20. Seno H, Oshima M, Ishikawa TO et al Cyclooxygenase 2‐and prostaglandin E2 receptor EP2‐dependent angiogenesis in Apc Δ716 mouse intestinal polyps. Cancer Res 2002; 62: 506–11. [PubMed] [Google Scholar]
- 21. Wang D, Wang H, Shi Q et al Prostaglandin E2 promotes colorectal adenoma growth via transactivation of the nuclear peroxisome proliferator‐activated receptor δ. Cancer Cell 2004; 6: 285–95. [DOI] [PubMed] [Google Scholar]
- 22. Xia D, Wang D, Kim SH et al Prostaglandin E2 promotes intestinal tumor growth via DNA methylation. Nat Med 2012; 18: 224–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Katoh H, Wang D, Daikoku T et al CXCR2‐expressing myeloid‐derived suppressor cells are essential to promote colitis‐associated tumorigenesis. Cancer Cell 2013; 24: 631–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Saukkonen K, RIntahaka J, Sivula A et al Cyclooxigenase‐2 and gastric carcinogenesis. APIMS 2003; 111: 915–25. [DOI] [PubMed] [Google Scholar]
- 25. Sung JJ, Leung WK, Go MY et al Cyclooxigenase‐2 expression in Helicobacter pylori‐associated premalignant and malignant gastric lesions. Am J Pathol 2000; 157: 729–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Oshima H, Oshima M, Inaba K et al Hyperplastic gastric tumors induced by activated macrophages in COX‐2/mPGES‐1 transgenic mice. EMBO J 2004; 23: 1669–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Cristescu R, Lee J, Nebozhyn M et al Molecular analysis of gastric cancer identifies subtypes associated with distinct clinical outcomes. Nat Med 2015; 21: 449–56. [DOI] [PubMed] [Google Scholar]
- 28. Oshima H, Hioki K, Popivanova BK et al Prostaglandin E2 signaling and bacterial infection recruit tumor‐promoting macrophages to mouse gastric tumors. Gastroenterology 2011; 140: 596–607. [DOI] [PubMed] [Google Scholar]
- 29. The Cancer Genome Atlas Research Network . Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014; 513: 202–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Itadani H, Oshima H, Oshima M et al Mouse gastric tumor models with prostaglandin E2 pathway activation show similar gene expression profiles to intestinal‐type human gastric cancer. BMC Genom 2009; 10: 615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Porter RL, Georger MA, Bromberg O et al Prostaglandin E2 increases hematopoietic stem cell survival and accelerates hematopoietic recovery after radiation injury. Stem Cells 2013; 31: 372–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Goessling W, North TE, Loewer S et al Genetic interaction of PGE2 and Wnt signaling regulates developmental specification of stem cells and regeneration. Cell 2009; 136: 1136–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Evans T. Fishing for a WNT‐PGE2 link: β‐catenin is caught in the stem cell net‐work. Cell Stem Cell 2009; 4: 280–2. [DOI] [PubMed] [Google Scholar]
- 34. Wang D, Fu L, Sun H et al Prostaglandin E2 promotes colorectal cancer stem cell expansion and metastasis in mice. Gastroenterology 2015; 149: 1884–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Barker N, Huch M, Kujala P et al Lgr5+ve stem cells drive self‐renewal in the stomach and build long‐lived gastric units in vitro. Cell Stem Cell 2010; 6: 25–36. [DOI] [PubMed] [Google Scholar]
- 36. Kulbe H, Chakravarty P, Leinster DA et al A dynamic cytokine network in the human ovarian cancer microenvironment. Cancer Res 2012; 72: 66–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Oshima H, Ishikawa T, Yoshida GJ et al TNF‐α/TNFR1 signaling promotes gastric tumorigenesis through induction of Noxo1 and Gna14 in tumor cells. Oncogene 2014; 33: 3820–9. [DOI] [PubMed] [Google Scholar]
- 38. Block K, Gorin Y. Aiding and abetting roles of NOX oxidases in cellular transformation. Nat Rev Cancer 2012; 12: 627–37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Laurent E, McCoy JW 3rd, Macina RA et al Nox1 is over‐expressed in human colon cancers and correlates with activating mutations in K‐Ras. Int J Cancer 2008; 123: 100–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Kodama R, Kato M, Furuta S et al ROS‐generating oxidases Nox1 and Nox4 contribute to oncogenic Ras‐induced premature senescence. Genes Cells 2013; 18: 32–41. [DOI] [PubMed] [Google Scholar]
- 41. Myant KB, Cammareri P, McGhee EJ et al ROS production and NF‐κB activation triggered by RAC1 facillitate WNT‐driven intestinal stem cell proliferation and colorectal cancer initiation. Cell Stem Cell 2013; 12: 761–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Ishimoto T, Nagano O, Yae T et al CD44 variant regulates redox status in cancer cells by stabilizing the xCT subunit of system xc(‐) and thereby promotes tumor growth. Cancer Cell 2011; 19: 387–400. [DOI] [PubMed] [Google Scholar]
- 43. Gorrini C, Harris IS, Mak TW. Modulation of oxidative stress as an anticancer stratedy. Nat Rev Drug Discov 2013; 12: 931–47. [DOI] [PubMed] [Google Scholar]
- 44. Parkin DM. The global health burden of infection‐associated cancers in the year 2002. Int J Cancer 2006; 118: 3030–44. [DOI] [PubMed] [Google Scholar]
- 45. de Martel C, Ferlay J, Franceschi S et al Global burden of cancers attributable to infections in 2008: a review and synthetic analysis. Lancet Oncol 2012; 13: 607–15. [DOI] [PubMed] [Google Scholar]
- 46. Hatakeyama M. Helicobacter pylori CagA and gastric cancer: a paradigm for hit‐and‐run carcinogenesis. Cell Host Microbe 2014; 15: 306–16. [DOI] [PubMed] [Google Scholar]
- 47. Pachathundikandi SK, Lind J, Tegtmeyer N et al Interplay of the gastric pathogen Helicobacter pylori with Toll‐like receptors. Biomed Res Int 2015; 2015: 192420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Wang K, Karin M. Tumor‐elicited inflammation and colorectal cancer. Adv Cancer Res 2015; 128: 173–96. [DOI] [PubMed] [Google Scholar]
- 49. Rakoff‐Nahoum S, Medzhitov R. Toll‐like receptors and cancer. Nat Rev Cancer 2009; 9: 57–63. [DOI] [PubMed] [Google Scholar]
- 50. Hanada K, Uchida T, Tsukamoto Y et al Helicobacter pylori infection introduces DNA double‐strand breaks in host cells. Infect Immun 2014; 82: 4182–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Howlett M, Giraud AS, Lescesen H et al The interleukin‐6 family cytokine interleukin‐11 regulates homeostatic epithelial cell turnover and promotes gastric tumor development. Gastroenterology 2009; 136: 967–77. [DOI] [PubMed] [Google Scholar]
- 52. Putoczki TL, Thiem S, Loving A et al Interleukin‐11 is the dominant IL‐6 family cytokine during gastrointestinal tumorigenesis and can be targeted therapeutically. Cancer Cell 2013; 24: 257–71. [DOI] [PubMed] [Google Scholar]
- 53. Yang L, Huang J, Ren X et al Abrogation of TGF‐β signaling in mammary carcinomas recruits Gr‐1 + CD11b+ myeloid cells that promote metastasis. Cancer Cell 2008; 13: 23–35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54. Newton K, Dixit VM. Signaling in innate immunity and inflammation. Cold Spring Harb Perspect Biol 2012; 4: a006049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Rakoff‐Nahoum S, Medzhitov R. Regulation of spontaneous intestinal tumorigenesis through the adaptor protein MyD88. Science 2007; 317: 124–7. [DOI] [PubMed] [Google Scholar]
- 56. Lee SH, Ju LL, Ganzales‐Navajas J et al ERK activation drives intestinal tumorigenesis in Apc min/+ mice. Nat Med 2010; 16: 665–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Uronis JM, Muhlbauer M, Herfarth HH et al Modulation of the intestinal microbiota alters colitis‐associated colorectal cancer susceptibility. PLoS ONE 2009; 4: e6026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Schiechl G, Bauer B, Fuss I et al Tumor development in murine ulcerative colitis depends on MyD88 signaling of colonic F4/80+CD11bhighGr1low macrophages. J Clin Invest 2011; 121: 1692–708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Kennedy CL, Najdovska M, Tye H et al Differential role of MyD88 and Mal/TIRAP in TLR2‐mediated gastric tumorigenesis. Oncogene 2014; 33: 2540–6. [DOI] [PubMed] [Google Scholar]
- 60. Naugler WE, Sakurai T, Kim S et al Gender disparity in liver cancer due to sex differences in MyD88‐dependent IL‐6 production. Science 2007; 317: 121–4. [DOI] [PubMed] [Google Scholar]
- 61. Swann JB, Vesely MD, Silva A et al Demonstration of inflammation‐induced cancer and cancer immunoediting during primary tumorigenesis. Proc Natl Acad Sci USA 2008; 105: 652–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Grivennikov SI, Wang K, Mucida D et al Adenoma‐linked barrier and microbial products drive IL‐23/IL‐17‐medicated tumour growth. Nature 2012; 491: 254–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Chae WJ, Bothwell AL. IL‐17F deficiency inhibits small intestinal tumorigenesis in ApcMin/+ mice. Biochem Biophys Res Commun 2011; 414: 31–6. [DOI] [PubMed] [Google Scholar]
- 64. Rakoff‐Nahoum S, Paglino J, Esiami‐Varzaneh F et al Recognition of commensal microflora by toll‐like receptors is required for intestinal homeostasis. Cell 2004; 118: 229–41. [DOI] [PubMed] [Google Scholar]
- 65. Sheeren FA, Kuo AH, van Weele LJ et al A cell‐intrinsic role for TLR2‐MYD88 in intestinal and breast epithelia and oncogenesis. Nat Cell Biol 2014; 16: 1238–48. [DOI] [PubMed] [Google Scholar]
- 66. Tye H, Kennedy CL, Najdovska M et al STAT3‐driven upregulation of TLR2 promotes gastric tumorigenesis independent of tumor inflammation. Cancer Cell 2012; 22: 466–78. [DOI] [PubMed] [Google Scholar]
- 67. Jenkins BJ, Grail D, Nheu T et al Hyperactivation of Stat3 in gp130 mutant mice promotes gastric hyperproliferation and desensitizes TGF‐β signaling. Nat Med 2005; 11: 845–52. [DOI] [PubMed] [Google Scholar]
- 68. Chefetz I, Alvero AB, Holmberg JC et al TLR2 enhances ovarian cancer stem cell self‐renewal and promotes tumor repair and recurrence. Cell Cycle 2013; 12: 511–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Conti L, Lanzardo S, Arigoni M et al The noninflammatory role of high mobility group box 1/toll‐like receptor 2 axis in the self‐renewal of mammary cancer stem cells. FASEB J 2013; 27: 4731–44. [DOI] [PubMed] [Google Scholar]
- 70. Schwitalla S, Fingerle AA, Cammareri P et al Intestinal tumorigenesis initiated by dedifferentiation and acquisition of stem‐cell‐like properties. Cell 2013; 152: 25–38. [DOI] [PubMed] [Google Scholar]
- 71. Kim S, Takahashi H, Lin WW et al Carcinoma‐produced factors activate myeloid cells through TLR2 to stimulate metastasis. Nature 2009; 457: 102–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Abe A, Kuwata T, Yamauchi C et al High Mobility Group Box1 (HMGB1) released from cancer cells induces the expression of pro‐inflammatory cytokines in peritoneal fibroblasts. Pathol Int 2014; 64: 58–66. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data S1. Methods for construction of mouse models, histological analyses of mouse and human gastric tumors, and gene expression analyses (Ingenuity pathway analysis, principal component analysis, and clustering analysis) are described.