

Summary
Strategies for improved homing of mesenchymal stem cells (MSCs) to a place of injury are being sought and it has been shown that natural killer (NK) cells can stimulate MSC recruitment. Here, we studied the chemokines behind this recruitment. Assays were performed with bone marrow human MSCs and NK cells freshly isolated from healthy donor buffy coats. Supernatants from MSC-NK cell co-cultures can induce MSC recruitment but not to the same extent as when NK cells are present. Antibody arrays and ELISA assays confirmed that NK cells secrete RANTES (CCL5) and revealed that human NK cells secrete NAP-2 (CXCL7), a chemokine that can induce MSC migration. Inhibition with specific antagonists of CXCR2, a receptor that recognizes NAP-2, abolished NK cell-mediated MSC recruitment. This capacity of NK cells to produce chemokines that stimulate MSC recruitment points toward a role for this immune cell population in regulating tissue repair/regeneration.
Graphical Abstract
Highlights
-
•
Primary unstimulated human NK cells produce NAP-2 (CXCL7)
-
•
NAP-2 is a chemokine that can promote recruitment of bone marrow MSCs
-
•
Inhibiting the NAP-2 receptor CXCR2 abolishes NK cell-mediated MSC recruitment
In this paper, Almeida and colleagues show that human unstimulated NK cells secrete NAP-2 (CXCL7), a chemokine that promotes human bone marrow MSC migration. Furthermore, inhibiting the NAP-2 receptor CXCR2 with an antagonist blocked NK cell-mediated MSC recruitment. Strategies for improved homing of MSCs can take advantage of this capacity of NK cells to recruit MSCs.
Introduction
Mesenchymal stem/stromal cells (MSCs) are attractive for different cell-based therapies, from bone regeneration to treatment of autoimmune diseases (Singer and Caplan, 2011). However, tissue regeneration therapies that involve injection/implantation of MSCs have not been fully successful, and strategies that use soluble mediators produced by MSCs or that attract endogenous stem cells and regulate its behavior are appealing, as recruitment, and not only proliferation and differentiation of progenitor cells, is important for effective repair/regeneration (Wei et al., 2013). In this context, it is important to know which factors regulate MSC recruitment.
Mobilization and recruitment of MSCs to a bone injury has been correlated with repair (Granero-Molto et al., 2009, Kumar and Ponnazhagan, 2012). Inflammatory mediators can lead to increased MSC migration (Ren et al., 2010, Tondreau et al., 2009), and thus immune cells such as macrophages and natural killer (NK) cells can stimulate MSC recruitment (Almeida et al., 2012, Anton et al., 2012). While monocytes/macrophages can also stimulate MSC differentiation along the osteoblastic lineage (Champagne et al., 2002, Ekstrom et al., 2013), NK cells do not interfere with MSC differentiation capacity (Almeida et al., 2012). This may be of interest as cell differentiation into specific lineages can then be orchestrated by other cues from the microenvironment. Macrophages can recruit MSC by producing the chemokine RANTES (Anton et al., 2012), which is involved in recruitment of MSCs in the degenerated intervertebral disc (Pattappa et al., 2014). However, the chemokines behind NK cell-mediated MSC recruitment are still unknown.
NK cells are one of the first immune cell populations to arrive at an injury site (Agaiby and Dyson, 1999), are involved in uterine tissue remodeling in pregnancy (Moffett and Colucci, 2014), may contribute to wound healing (Liippo et al., 2009), and can trigger differentiation of monocytes into osteoclasts (Soderstrom et al., 2010). NK cells are capable of recognizing cells in different stages of the cell cycle (Nolte-'t Hoen et al., 2006), and their activation by target cells depends on the target cells activating/inhibitory ligands ratio and distribution in the cell membrane (Almeida and Davis, 2006, Endt et al., 2007, Kaplan et al., 2011). Activation of NK cells by different ligands or in different contexts may lead to degranulation of lytic granules, or cytokine or chemokine secretion (Almeida et al., 2011, Fauriat et al., 2010). NK cells produce several chemokines (Fauriat et al., 2010, Robertson, 2002), which can stimulate MSC recruitment (Anton et al., 2012, Ponte et al., 2007).
In this work, we dissected some of the chemokines involved in NK-cell-mediated MSC recruitment, focusing on NAP-2, GRO-β, GRO-γ, interleukin-8 (IL-8), and RANTES. Resting human NK cells can produce different soluble factors that may play a role in this recruitment. Of significance is secretion of NAP-2, which can stimulate MSC migration on its own. Furthermore, treating MSCs with an antagonist specific for the chemokine receptor CXCR2 abolished MSC recruitment by NK cells.
Results and Discussion
NK Cells Produce Soluble Mediators that Stimulate MSC Invasion
NK cells stimulate MSC invasion through transwell chambers coated with Matrigel, which mimics the extracellular matrix. In order to dissect regulation of MSC recruitment, invasion assays were performed with MSCs pre-treated with pertussis toxin, which inhibits all G-coupled proteins, including chemokine receptors and others. NK-cell-mediated MSC recruitment was inhibited in the presence of pertussis toxin (Figure 1A). As NK cells are known producers of chemokines, which are usually involved in recruitment of cells in an inflammatory context, this suggests chemokines receptors may have a role in NK-cell-mediated MSC recruitment.
Figure 1.

Characterization of Chemokines Involved in NK-Cell-Mediated MSC Recruitment
(A and B) MSC invasion through filters coated with Matrigel was determined after 24 hr of incubation. (A) The lower compartment of the invasion chamber was filled with serum-free medium, as a negative control, or with NK cells. MSCs were either untreated (control) or pre-treated with pertussis toxin at 100 ng/ml for 2 hr at 37°C/5% CO2 (n = 6 independent experiments with four NK cell and two MSC donors, ∗p < 0.05; Wilcoxon test). (B) The lower compartment was filled with serum-free medium, NK cells, or with supernatants (sup) from MSCs or NK cells cultured alone or collected after an invasion assay. n = 6–9 independent experiments with six NK cell and three MSC donors, ∗p < 0.05; ∗∗∗p < 0.001; Kruskal-Wallis test followed by the Dunn's multiple comparisons test to compare each sample against the negative control.
(C) The supernatants were analyzed with protein arrays detecting 38 different factors. Examples of the developed membranes with indication of factors of interest.
(D) Concentrations of RANTES, GRO-β, GRO-γ, NAP-2, and IL-8 in supernatants of NK cells, MSCs, or NK cells co-cultured with MSCs in invasion assays were determined by ELISA. Graphs are min-max whisker plots. n = 10 independent experiments with ten NK cell and three MSC donors; ∗p < 0.05; ∗∗∗p < 0.001; Friedman test followed by Dunn's multiple comparisons test to compare each sample against the negative control.
(E) Intracellular staining of NAP-2 (bottom) was performed on isolated NK cells (CD56+/CD3−, top). Plots are representative of data from four NK cell donors.
Invasion assays were performed with MSCs stimulated with NK cells or conditioned media collected from NK cells and MSCs alone or in co-culture. The number of MSCs that invaded through the Matrigel coating increased in the presence of media from co-cultures but not as much as in the presence of NK cells (Figure 1B). This indicates that NK cells continuously produce soluble factors that promote MSC invasion, which in the case of the supernatants will be consumed without being replenished. The fact that only conditioned media from co-cultures had a significant effect suggests that either MSCs also produce mediators that promote its own recruitment and/or that MSCs further stimulate NK cells to produce the factors involved in this recruitment.
The overall number of MSCs being recruited by NK cells might seem low, but it represents a fold increase higher than three. In these invasion assays, cells have to remodel the matrix to cross the transwell, and thus the number of cells that cross the membrane will be lower than in migration assays, with uncoated membranes. The number of recruited cells might increase when there are more NK cells as a stimulus (Almeida et al., 2012) or when NK cells are stimulated to produce higher amounts of mediators.
NK Cells Secrete Chemokines Such as NAP-2
To find chemokines that might be stimulating MSC recruitment, supernatants from MSC, NK cells, or co-cultures were analyzed with antibody arrays. NK cells and MSCs secreted different chemokines, including the yet undescribed NAP-2 for NK cells and GRO for MSCs (Figure 1C). Other chemokines that have been reported to have a role in regulating MSC recruitment, such as SDF-1, MIP-1α, MIP-1β, and MCP-1 (Eseonu and De Bari, 2015), were not consistently detected for all donors analyzed and thus were not further pursued.
With three independent NK cell donors, semi-quantitative analysis revealed a decrease in the amount of NAP-2 and RANTES and an increase in the amount of GRO (but not GRO-α) in the co-culture, compared with the sum of the amounts produced by NK cells and MSCs separately (Figure S1). IL-8 is a cytokine known to promote MSC recruitment (Anton et al., 2012) that was secreted at very high levels by MSCs and whose increase or decrease in the co-culture varied. Therefore, ELISA assays were performed for quantification of RANTES, GRO-β, GRO-γ, NAP-2, and IL-8 (Figure 1D). Importantly, NK cells from all donors secreted significant amounts of NAP-2, with a slight increase in the presence of MSCs. This is in agreement with the increased recruitment of MSCs by supernatants from MSC-NK cell co-cultures, and strengthens the need to confirm data obtained by antibody arrays. NAP-2 is a chemokine produced by platelets that stimulates recruitment and activation of neutrophils and has never been reported to be produced by NK cells. Intracellular staining against NAP-2 on freshly isolated NK cells from four donors confirmed that 27.3% ± 7.6% of CD56+ cells expressed NAP-2 (Figure 1E). RANTES and IL-8 have been described to be involved in macrophage-mediated MSC recruitment (Anton et al., 2012). Here, it was confirmed that NK cells secrete RANTES, producing a lower amount in the presence of MSCs. The amount of secreted IL-8 increased significantly in the MSC-NK cell co-cultures. This suggests that RANTES is being consumed in the co-culture while production of IL-8 is being further stimulated. No statistically significant differences were found for the other mediators.
NAP-2 Can Promote MSC Migration
To clarify the role of chemokines, which are expected to stimulate cell migration but not matrix remodeling, migration assays were performed, with chambers that were not coated with Matrigel, instead of the more physiological invasion assays. MSC migration was tested with recombinant human RANTES, GRO-β, GRO-γ, NAP-2, IL-8, or all these factors combined. As in migration assays, cells do not have to remodel a matrix; recruitment occurs with a faster kinetics and thus chambers were incubated for 6 hr as opposed to the 24 hr necessary to analyze invasion.
Data showed that GRO-γ at 1 ng/ml and NAP-2 at 10 ng/ml stimulated MSC recruitment (Figure 2). Others have shown that high doses of NAP-2 can also stimulate MSC recruitment and that treatment with these doses leads to increased production of other chemokines, including all the ones that bind to CXCR2, the receptor for NAP-2 (Kalwitz et al., 2009).
Figure 2.

Number of MSC Migrating when Stimulated with Recombinant Human RANTES, GRO-β, GRO-γ, NAP-2, IL-8, or a Combination of All These Chemokines
A migration assay was performed with an uncoated transwell for 6 hr when MSCs were stimulated with 1 or 10 ng/ml of recombinant proteins. Graphs are min-max whisker plots. n = 5–7 independent experiments with three MSC donors; ∗p < 0.05; data were compared with the Friedman test followed by Dunn's multiple comparisons test.
All other factors analyzed showed a tendency to stimulate MSC migration, but to levels that were not statistically significant. A characteristic bell-shaped response curve could be observed with GRO-γ and IL-8. It has been reported that both RANTES and IL-8 stimulate MSC migration, but for 18 hr migration assays performed in the presence of 10% fetal bovine serum (FBS) (Anton et al., 2012). Here experiments were performed without serum as it is capable on its own of stimulating migration, and may thus mask the effect that NK cells have on MSC recruitment. When combining all proteins, there was a slight but not statistically significant increase in the number of MSCs that migrated. It can be speculated that, when combining all the proteins together, an equilibrium between chemoattractants and chemorepellents is reached. Chemokines at high concentrations can become chemorepellent, and fugetaxis, the movement of leukocytes away from chemotactic agents, has been described (Vianello et al., 2005). When testing the recombinant proteins on their own, it was clear that NAP-2 induced migration at the higher concentration of 10 ng/ml and GRO-γ only at the lower concentration of 1 ng/ml, with a decrease in migration for higher concentrations. As these chemokines can bind to the same receptor, it is possible that the amount of chemokine binding to CXCR2 is such that there is no chemoattraction. Thus, the effect of NAP-2 might depend on the microenvironment (and the other chemokines) the cells will encounter.
Blocking CXCR2 Abolishes NK-Cell-Mediated MSC Recruitment
Table 1 summarizes the published data on MSC expression of chemokine receptors for the chemokines analyzed here. Contradictory results on expression of the chemokine receptors CCR1, CCR3, CCR5, CXCR1, and CXCR2 may be due to differences in isolation techniques, culture conditions, harvesting protocols, methods used for detection of these molecules, or even in differences between donors. But interestingly, GRO-β, GRO-γ, NAP-2, IL-8 can all be recognized by the chemokine receptor CXCR2, whose expression on MSCs has been described (Kalwitz et al., 2009, Ringe et al., 2007) and confirmed here by imaging flow cytometry and immunocytochemistry (Figure S2). Therefore, invasion assays were performed with MSCs treated with a CXCR2 antagonist, SB225002 (Figure 3A). It was found that inhibiting CXCR2 abolished NK-cell-mediated MSC recruitment.
Table 1.
Summary of Published Data on Human Bone Marrow MSC Expression of Chemokine Receptors that Recognize the Chemokines that Were Analyzed
| Chemokine | Other Names | Receptor | Expression of Receptors by MSCs |
|---|---|---|---|
| RANTES | CCL5 | CCR1, CCR3, CCR5 | No expression of CCR1, while CCR3 and CCR5 expression increase after TNF-α stimulation (Hemeda et al., 2010, Ponte et al., 2007); only CCR3 detected, expression increases after TNF-α and interferon-γ stimulation (Croitoru-Lamoury et al., 2007); positive for CCR1, negative for CCR3 and CCR5 (Honczarenko et al., 2006); positive for intracellular CCR1 and CCR3 and negative for CCR5 expression (Brooke et al., 2008); small percentage expresses CCR1, no mRNA detected for CCR3 or CCR5 (Sordi et al., 2005); positive for CCR1 and CCR3 (Djouad et al., 2007) |
| GRO-β | CXCL2, MIP-2α | CXCR2 | No expression (Honczarenko et al., 2006, Ponte et al., 2007, Sordi et al., 2005); MSCs positive for both CXCR2 and CXCR1 (Kalwitz et al., 2009, Ringe et al., 2007) |
| GRO-γ | CXCL3, MIP-2β | CXCR2 | |
| NAP-2 | CXCL7 | CXCR2 | |
| IL-8 | CXCL8 | CXCR1, CXCR2 |
Figure 3.

NK-Cell-Mediated MSC Recruitment Involves the Chemokine Receptor CXCR2
(A) MSCs were treated for 1 hr with 0.13 μM of the CXCR2 antagonist SB225002 before performing a 24 hr invasion assay through Matrigel-coated transwells when stimulated with medium only or with NK cells. The chemotactic index is calculated by dividing the number of MSCs migrating when stimulated with NK cells by the negative control. Graphs are min-max whisker plots. n = 8 independent experiments with eight NK cell and two MSC donors; ∗p < 0.05; ∗∗p < 0.01; Kruskal-Wallis test followed by Dunn's multiple comparisons test to compare all samples.
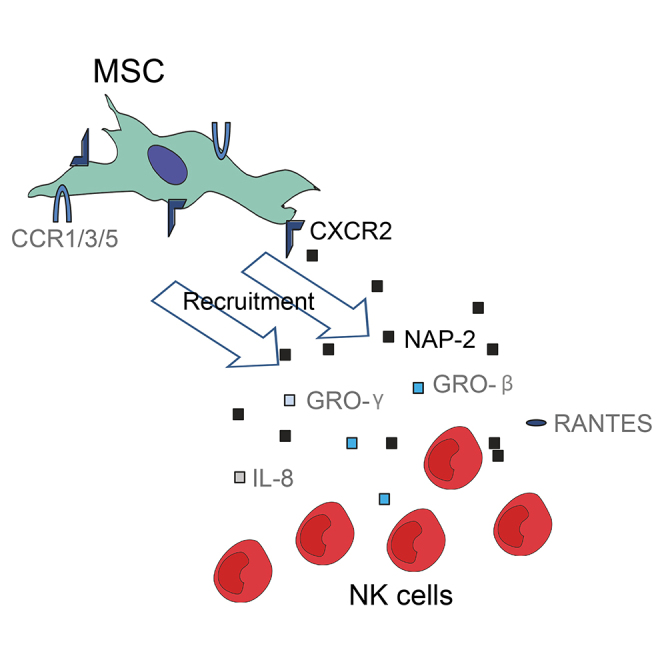
(B) Resting human NK cells produce NAP-2, which stimulates MSC recruitment via CXCR2. Furthermore, NK cells are capable of producing other chemokines that can bind to CXCR2, such as GRO-β, GRO-γ, and IL-8. NK cells also produce RANTES, which might be recognized by CCR1, CCR3, or CCR5. On the other hand, MSCs seem to produce unknown paracrine factors that affect NK cells and thus affect stimulation of MSC recruitment.
Here, it was shown that freshly isolated primary NK cells can produce NAP-2. In an injury scenario, NK cells will be recruited and if reaching a sufficiently high number of cells, they might establish an NAP-2 gradient. It is not clear whether NK cells in an injury/inflammation site become activated as the cytokines that are well known to activate NK cells (IL-2, IL-15, IL-18, and IL-12) are not common at an injury site. However, cytokines characteristic of tissue repair such as tumor necrosis factor alpha (TNF-α), IL-6, and IL-1, can affect NK cell maturation, differentiation, cytotoxicity, and cytokine secretion (Cooper et al., 2001, Lee et al., 2009, Passos et al., 2010). NK cells can also become activated to secrete chemokines by binding to different ligands (Fauriat et al., 2010). Thus, future strategies encompassing biomaterials that either stimulate NAP-2 production or that arrest NK cells to function as factories of chemokines will be of interest.
To conclude, here we propose that NAP-2 secreted by human NK cells can bind to CXCR2 on MSCs leading to stimulation of its recruitment (Figure 3B). Besides NAP-2, other chemokines that bind to CXCR2 can be produced by NK cells, albeit to different extents by different donors. NK cells can also produce RANTES, which may also have a role in mediating MSC recruitment. The fact that NK cells isolated from different donors produce different CXCR2-binding chemokines suggests that CXCR2 may be a key chemokine receptor.
Experimental Procedures
MSC and NK Cell Isolation
MSCs and NK cells were isolated as described elsewhere (Almeida et al., 2012). All experimental protocols were conducted following the approval and recommendations of the Ethics Committee for Health from Centro Hospitalar S. João. MSCs were isolated from human bone marrow samples collected by Serviço de Ortopedia e Traumatologia from Hospital de São João after ethics committee approval and informed consent. Isolation of MSCs was confirmed by surface staining of CD105, CD73, CD90, CD45, CD34, CD14, CD19, and HLA-DR and by testing differentiation in osteoblasts, chondroblasts, or adipocytes (Almeida et al., 2012). Prior to each experiment, frozen aliquots of MSCs were thawed and cultured in DMEM with low glucose and with Glutamax plus 10% selected inactivated FBS and 1% penicillin/streptomycin (all from Invitrogen). After reaching about 80% confluence, cells were detached with 0.05% trypsin/EDTA. Experiments were performed with MSCs from four donors, in passages 6–8.
NK cells were obtained from human healthy donors buffy coats, kindly provided by Hospital de São João. Briefly, a peripheral blood mononuclear cell suspension was prepared by density gradient centrifugation, and NK cells were purified by negative selection using the EasySep human NK cell enrichment kit (STEMCELL Technologies), according to the manufacturer's instructions. The percentage of CD56+/CD3- cells was higher than 80% (average 90%). NK cells were isolated from 12 different donors.
Migration and Invasion Assays
Studies on migration and invasion of MSCs were performed using a transwell chamber system. Membrane filters with a pore size of 8 μm were used, and coated with Matrigel (Corning) for invasion assays. For migration assays, the top chamber was incubated with 100 μl of bovine gelatin 0.1% for 1 hr at 37°C/5% CO2 and washed with PBS to promote cell adhesion. For invasion assays, Matrigel-coated inserts were pre-incubated for 1 hr with serum-free DMEM. The lower compartments of the chamber were filled with 750 μl of DMEM medium, as a negative control, or serum-free DMEM with NK cells at an NK/MSC ratio of 15:1. MSCs (4 × 104 cells per well) in 500 μl of serum-free DMEM medium were seeded into the upper compartment. Migration and invasion chambers were incubated at 37°C/5% CO2 for 6 hr and 24 hr, respectively. After incubation, inserts were washed with PBS and cells were fixed in 4% paraformaldehyde for 15 min at room temperature (RT). Supernatants from invasion assays were spun at 300 × g for 10 min, collected, and frozen at −80°C for other assays. Inserts were washed with PBS and kept at 4°C until analysis. Cells on the top surfaces of the filters were wiped off with cotton swabs, and the membrane was mounted in a slide with Vectashield and DAPI. Cells that had migrated and attached to the lower surface of the filter were counted in a fluorescence microscope (Zeiss Axiovert). Cell nuclei were counted in ten 200× fields of view for each membrane. The number of migrated cells was estimated by taking into account the area of a field of view and the total area of the membrane.
Assays with supernatants were performed by adding 750 μl of collected supernatant to the bottom well. When specified, MSCs were pre-treated with pertussis toxin at 100 ng/ml for 2 hr at 37°C/5% CO2, harvested, and added to the invasion chambers. Migration assays with recombinant proteins were performed with 1 or 10 ng/ml RANTES, GRO-β, GRO-γ, NAP-2, or IL-8 (Peprotech). For treatment with antagonist, MSCs were pre-incubated with 0.13 μM SB225002 (Sigma) for 1 hr in the invasion chamber. Controls were performed with the vehicle DMSO.
Protein Arrays
The presence of different chemokines was analyzed in supernatants collected from invasion assays with the human chemokine antibody array 1 (RayBiotech) following the manufacturer's instructions.
ELISA Assays
ELISA kits were used to quantify RANTES (RayBiotech), NAP-2 (RayBiotech), and IL-8 (BioLegend). For GRO-β and GRO-γ, plates were incubated at 4°C with agitation for 24 hr with 100 μl of serially diluted standard in PBS or samples. Plates were washed four times with 300 μl of wash buffer (0.05% Tween 20 in PBS) per well and blocked for 2 hr at RT with 300 μl of blocking buffer (1% BSA, BSA, in PBS). Plates were washed prior to adding 1 μg/ml biotinylated anti-GRO-β (rabbit IgG) or anti-GRO-γ (rabbit IgG) antibodies (both from Peprotech) in 0.05% Tween 20/0.1% BSA in PBS. Plates were incubated for 2 hr and washed. Then, 100 μl of avidin-horseradish peroxidase conjugate (1:2,000) was added, and plates were incubated for 30 min at RT. Plates were washed and 100 μl of substrate solution (1 mg/ml o-phenylenediamine dihydrochloride with 0.03% H2O2 in phosphate citrate buffer 0.05 M [pH 5.0]) was added. Plates were incubated at RT for color development and 100 μl of H2SO4 1 M was added. Plates were read at 492 nm with wavelength correction at 630 nm.
NAP-2 Intracellular Staining
NK cells were cultured at 106 cells/ml for 5 hr at 37°C/5% CO2 with 10 μg/ml brefeldin A (Sigma). Then, cells were collected and fixed with paraformaldehyde 4% for 15 min at RT, washed, and permeabilized with 0.5% BSA/0.2% Tween/PBS on ice for 15 min. Cells were incubated with 10 μg/ml anti-human NAP-2 antibody (# 59418, R&D Systems) in 0.5% BSA/0.2% Tween/PBS for 1 hr on ice. Cells were washed before incubating with anti-mouse Alexa Fluor 647 antibody (1:500; Invitrogen) for 45 min and washed again. Cells were then stained with anti-CD56-PE (AF12-7H3, Miltenyi Biotec) and anti-CD3-FITC (MEM-57, Immunotools) on ice for 30 min. Cells were washed and 10,000 events were acquired in a Flow Cytometer (FACSCanto II; Becton Dickinson) and analyzed with FlowJo. Appropriate isotype controls were used.
Statistical Analysis
After verifying that the data did not follow a normal distribution, differences between samples were determined with the Wilcoxon, Kruskal-Wallis, and Friedman non-parametric tests. The Wilcoxon test was performed to compare two related samples; the Kruskal-Wallis and Friedman tests were used when there were more than two conditions to compare unpaired and paired data, respectively, and were followed by the Dunn's multiple comparisons test. Statistical analyses were performed with GraphPad Prism 5 for Windows.
Author Contributions
C.A., conception and design, financial support, collection and assembly of data, data analysis and interpretation, manuscript writing, final approval of manuscript; H.C. and D.V., collection and assembly of data, data analysis and interpretation, final approval of manuscript; M.B. conception and design, financial support, data interpretation, final approval of manuscript.
Acknowledgments
We would like to thank Hospital de São João for bone marrow samples and buffy coats. This work had the financial support of FCT/MEC through National Funds and, when applicable, co-financed by the FEDER through the PT2020 Partnership Agreement under the 4293 Unit I&D.
Published: March 24, 2016
Footnotes
This is an open access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/).
Supplemental Information includes two figures and can be found with this article online at http://dx.doi.org/10.1016/j.stemcr.2016.02.012.
Supplemental Information
References
- Agaiby A.D., Dyson M. Immuno-inflammatory cell dynamics during cutaneous wound healing. J. Anat. 1999;195:531–542. doi: 10.1046/j.1469-7580.1999.19540531.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida C.R., Davis D.M. Segregation of HLA-C from ICAM-1 at NK cell immune synapses is controlled by its cell surface density. J. Immunol. 2006;177:6904–6910. doi: 10.4049/jimmunol.177.10.6904. [DOI] [PubMed] [Google Scholar]
- Almeida C.R., Ashkenazi A., Shahaf G., Kaplan D., Davis D., Mehr R. Human NK cells differ more in their KIR2DL1-dependent thresholds for HLA-Cw6-mediated inhibition than in their maximal killing capacity. PLoS One. 2011;6:e24927. doi: 10.1371/journal.pone.0024927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida C.R., Vasconcelos D.P., Gonçalves R.M., Barbosa M.A. Enhanced mesenchymal stromal cell recruitment via natural killer cells by incorporation of inflammatory signals in biomaterials. J. R. Soc. Interf. 2012;9:261–271. doi: 10.1098/rsif.2011.0357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anton K., Banerjee D., Glod J. Macrophage-associated mesenchymal stem cells assume an activated, migratory, pro-inflammatory phenotype with increased IL-6 and CXCL10 secretion. PLoS One. 2012;7:e35036. doi: 10.1371/journal.pone.0035036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooke G., Tong H., Levesque J.P., Atkinson K. Molecular trafficking mechanisms of multipotent mesenchymal stem cells derived from human bone marrow and placenta. Stem Cells Dev. 2008;17:929–940. doi: 10.1089/scd.2007.0156. [DOI] [PubMed] [Google Scholar]
- Champagne C.M., Takebe J., Offenbacher S., Cooper L.F. Macrophage cell lines produce osteoinductive signals that include bone morphogenetic protein-2. Bone. 2002;30:26–31. doi: 10.1016/s8756-3282(01)00638-x. [DOI] [PubMed] [Google Scholar]
- Cooper M.A., Fehniger T.A., Ponnappan A., Mehta V., Wewers M.D., Caligiuri M.A. Interleukin-1beta costimulates interferon-gamma production by human natural killer cells. Eur. J. Immunol. 2001;31:792–801. doi: 10.1002/1521-4141(200103)31:3<792::aid-immu792>3.0.co;2-u. [DOI] [PubMed] [Google Scholar]
- Croitoru-Lamoury J., Lamoury F.M., Zaunders J.J., Veas L.A., Brew B.J. Human mesenchymal stem cells constitutively express chemokines and chemokine receptors that can be upregulated by cytokines, IFN-beta, and Copaxone. J. Interferon Cytokine Res. 2007;27:53–64. doi: 10.1089/jir.2006.0037. [DOI] [PubMed] [Google Scholar]
- Djouad F., Delorme B., Maurice M., Bony C., Apparailly F., Louis-Plence P., Canovas F., Charbord P., Noel D., Jorgensen C. Microenvironmental changes during differentiation of mesenchymal stem cells towards chondrocytes. Arthritis Res. Ther. 2007;9:R33. doi: 10.1186/ar2153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ekstrom K., Omar O., Graneli C., Wang X., Vazirisani F., Thomsen P. Monocyte exosomes stimulate the osteogenic gene expression of mesenchymal stem cells. PLoS One. 2013;8:e75227. doi: 10.1371/journal.pone.0075227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Endt J., McCann F.E., Almeida C.R., Urlaub D., Leung R., Pende D., Davis D.M., Watzl C. Inhibitory receptor signals suppress ligation-induced recruitment of NKG2D to GM1-rich membrane domains at the human NK cell immune synapse. J. Immunol. 2007;178:5606–5611. doi: 10.4049/jimmunol.178.9.5606. [DOI] [PubMed] [Google Scholar]
- Eseonu O.I., De Bari C. Homing of mesenchymal stem cells: mechanistic or stochastic? Implications for targeted delivery in arthritis. Rheumatology (Oxford) 2015;54:210–218. doi: 10.1093/rheumatology/keu377. [DOI] [PubMed] [Google Scholar]
- Fauriat C., Long E.O., Ljunggren H.-G., Bryceson Y.T. Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood. 2010;115:2167–2176. doi: 10.1182/blood-2009-08-238469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Granero-Molto F., Weis J.A., Miga M.I., Landis B., Myers T.J., O'Rear L., Longobardi L., Jansen E.D., Mortlock D.P., Spagnoli A. Regenerative effects of transplanted mesenchymal stem cells in fracture healing. Stem Cells. 2009;27:1887–1898. doi: 10.1002/stem.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hemeda H., Jakob M., Ludwig A.K., Giebel B., Lang S., Brandau S. Interferon-gamma and tumor necrosis factor-alpha differentially affect cytokine expression and migration properties of mesenchymal stem cells. Stem Cells Dev. 2010;19:693–706. doi: 10.1089/scd.2009.0365. [DOI] [PubMed] [Google Scholar]
- Honczarenko M., Le Y., Swierkowski M., Ghiran I., Glodek A.M., Silberstein L.E. Human bone marrow stromal cells express a distinct set of biologically functional chemokine receptors. Stem Cells. 2006;24:1030–1041. doi: 10.1634/stemcells.2005-0319. [DOI] [PubMed] [Google Scholar]
- Kalwitz G., Endres M., Neumann K., Skriner K., Ringe J., Sezer O., Sittinger M., Häupl T., Kaps C. Gene expression profile of adult human bone marrow-derived mesenchymal stem cells stimulated by the chemokine CXCL7. Int. J. Biochem. Cell Biol. 2009;41:649–658. doi: 10.1016/j.biocel.2008.07.011. [DOI] [PubMed] [Google Scholar]
- Kaplan A., Kotzer S., Almeida C.R., Kohen R., Halpert G., Salmon-Divon M., Kohler K., Hoglund P., Davis D.M., Mehr R. Simulations of the NK cell immune synapse reveal that activation thresholds can be established by inhibitory receptors acting locally. J. Immunol. 2011;187:760–773. doi: 10.4049/jimmunol.1002208. [DOI] [PubMed] [Google Scholar]
- Kumar S., Ponnazhagan S. Mobilization of bone marrow mesenchymal stem cells in vivo augments bone healing in a mouse model of segmental bone defect. Bone. 2012;50:1012–1018. doi: 10.1016/j.bone.2012.01.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J., Lee S.H., Shin N., Jeong M., Kim M.S., Kim M.J., Yoon S.R., Chung J.W., Kim T.D., Choi I. Tumor necrosis factor-alpha enhances IL-15-induced natural killer cell differentiation. Biochem. Biophys. Res. Commun. 2009;386:718–723. doi: 10.1016/j.bbrc.2009.06.120. [DOI] [PubMed] [Google Scholar]
- Liippo J., Toriseva M., Kähäri V.-M. Natural killer cell in wound healing. In: Lotze M., Thomson A., editors. Natural Killer Cells - Basic Science and Clinical Application. Academic Press; 2009. pp. 519–525. [Google Scholar]
- Moffett A., Colucci F. Uterine NK cells: active regulators at the maternal-fetal interface. J. Clin. Invest. 2014;124:1872–1879. doi: 10.1172/JCI68107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nolte-'t Hoen E., Almeida C., Cohen N., Nedvetzki S., Yarwood H., Davis D. Increased surveillance of cells in mitosis by human NK cells suggests a novel strategy for limiting tumor growth and viral replication. Blood. 2006;109:670–673. doi: 10.1182/blood-2006-07-036509. [DOI] [PubMed] [Google Scholar]
- Passos S.T., Silver J.S., O’Hara A.C., Sehy D., Stumhofer J.S., Hunter C.A. Il-6 promotes NK cell production of IL-17 during toxoplasmosis. J. Immunol. 2010;184:1776–1783. doi: 10.4049/jimmunol.0901843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pattappa G., Peroglio M., Sakai D., Mochida J., Benneker L.M., Alini M., Grad S. CCL5/RANTES is a key chemoattractant released by degenerative intervertebral discs in organ culture. Eur. Cell. Mater. 2014;27:124–136. doi: 10.22203/ecm.v027a10. discussion 136. [DOI] [PubMed] [Google Scholar]
- Ponte A.L., Marais E., Gallay N., Langonne A., Delorme B., Herault O., Charbord P., Domenech J. The in vitro migration capacity of human bone marrow mesenchymal stem cells: comparison of chemokine and growth factor chemotactic activities. Stem Cells. 2007;25:1737–1745. doi: 10.1634/stemcells.2007-0054. [DOI] [PubMed] [Google Scholar]
- Ren G., Zhao X., Zhang L., Zhang J., L'Huillier A., Ling W., Roberts A.I., Le A.D., Shi S., Shao C. Inflammatory cytokine-induced intercellular adhesion Molecule-1 and vascular cell adhesion Molecule-1 in mesenchymal stem cells are critical for immunosuppression. J. Immunol. 2010;184:2321–2328. doi: 10.4049/jimmunol.0902023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ringe J., Strassburg S., Neumann K., Endres M., Notter M., Burmester G.R., Kaps C., Sittinger M. Towards in situ tissue repair: human mesenchymal stem cells express chemokine receptors CXCR1, CXCR2 and CCR2, and migrate upon stimulation with CXCL8 but not CCL2. J. Cell. Biochem. 2007;101:135–146. doi: 10.1002/jcb.21172. [DOI] [PubMed] [Google Scholar]
- Robertson M.J. Role of chemokines in the biology of natural killer cells. J. Leukoc. Biol. 2002;71:173–183. [PubMed] [Google Scholar]
- Singer N.G., Caplan A.I. Mesenchymal stem cells: mechanisms of inflammation. Annu. Rev. Pathol. 2011;6:457–478. doi: 10.1146/annurev-pathol-011110-130230. [DOI] [PubMed] [Google Scholar]
- Soderstrom K., Stein E., Colmenero P., Purath U., Muller-Ladner U., de Matos C.T., Tarner I.H., Robinson W.H., Engleman E.G. Natural killer cells trigger osteoclastogenesis and bone destruction in arthritis. Proc. Natl. Acad. Sci. USA. 2010;107:13028–13033. doi: 10.1073/pnas.1000546107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sordi V., Malosio M.L., Marchesi F., Mercalli A., Melzi R., Giordano T., Belmonte N., Ferrari G., Leone B.E., Bertuzzi F. Bone marrow mesenchymal stem cells express a restricted set of functionally active chemokine receptors capable of promoting migration to pancreatic islets. Blood. 2005;106:419–427. doi: 10.1182/blood-2004-09-3507. [DOI] [PubMed] [Google Scholar]
- Tondreau T., Meuleman N., Stamatopoulos B., De Bruyn C., Delforge A., Dejeneffe M., Martiat P., Bron D., Lagneaux L. In vitro study of matrix metalloproteinase/tissue inhibitor of metalloproteinase production by mesenchymal stromal cells in response to inflammatory cytokines: the role of their migration in injured tissues. Cytotherapy. 2009;11:559–569. doi: 10.1080/14653240903051541. [DOI] [PubMed] [Google Scholar]
- Vianello F., Olszak I.T., Poznansky M.C. Fugetaxis: active movement of leukocytes away from a chemokinetic agent. J. Mol. Med. (Berl) 2005;83:752–763. doi: 10.1007/s00109-005-0675-z. [DOI] [PubMed] [Google Scholar]
- Wei X., Yang X., Han Z.P., Qu F.F., Shao L., Shi Y.F. Mesenchymal stem cells: a new trend for cell therapy. Acta Pharmacol. Sin. 2013;34:747–754. doi: 10.1038/aps.2013.50. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.