

Abstract
γ-Tocopherol increases responses to allergen challenge in allergic adult mice, but it is not known whether γ-tocopherol regulates the development of allergic disease. Development of allergic disease often occurs early in life. In clinical studies and animal models, offspring of allergic mothers have increased responsiveness to allergen challenge. Therefore, we determined whether γ-tocopherol augments development of allergic responses in offspring of allergic female mice. Allergic female mice were supplemented with γ-tocopherol starting at mating. The pups from allergic mothers developed allergic lung responses, whereas pups from saline-treated mothers did not respond to allergen challenge. The γ-tocopherol supplementation of allergic female mice increased the numbers of eosinophils twofold in the pup bronchoalveolar lavage and lungs after allergen challenge. There was also about a twofold increase in pup lung CD11b+ subsets of CD11c+ dendritic cells and in numbers of these dendritic cells expressing the transcription factor IRF4. There was no change in several CD11b− dendritic cell subsets. Furthermore, maternal supplementation with γ-tocopherol increased the number of fetal liver CD11b+CD11c+ dendritic cells twofold in utero. In the pups, γ-tocopherol increased lung expression of the inflammatory mediators CCL11, amphiregulin, activin A, and IL-5. In conclusion, maternal supplementation with γ-tocopherol increased fetal development of subsets of dendritic cells that are critical for allergic responses and increased development of allergic responses in pups from allergic mothers. These results have implications for supplementation of allergic mothers with γ-tocopherol in prenatal vitamins.
Keywords: allergic lung inflammation, γ-tocopherol, vitamin E, dendritic cells
development of allergic disease originates as complex environmental and genetic interactions that may start early in life (54), a time when the airways and the immune system are developing. It has been suggested that risk for allergic disease in humans is associated with in utero and early exposures to environmental factors (9). In reports examining whether development of allergies in offspring associate with human maternal or paternal asthma, most associations are with maternal asthma (50). In animal studies, offspring from allergic mothers are predisposed to responses to suboptimal doses of allergens (28, 30, 35, 36, 38, 39, 42, 50). Interestingly, in animal studies, dendritic cells (DCs), but not macrophages, from offspring of allergic mothers transfer the risk for allergies to naïve neonates (26). This indicates that in mice there is a functional change in neonatal DCs during development of offspring of allergic mothers (26).
The prevalence of asthma from 1950 to the present has increased in many countries (10, 29, 84). In humans, the marked differences in prevalence of asthma within countries, in migrating populations, and over relatively short periods of time, support an important role of exposure to the local environment, such as diet, in asthma inception. One environmental change over the past 50 yr has been an increase in the γ-tocopherol form of vitamin E in the diet, in infant formulas, and in vitamin supplements (22, 78). Studies with vitamin E report seemingly conflicting outcomes for allergy and other inflammatory diseases (21, 24, 70, 77, 89). We recently demonstrated that different isoforms of vitamin E have opposing functional outcomes in adult lungs in animals and humans (1, 7, 17, 18, 21, 53, 55).
Vitamin E consists of natural isoforms and synthetic racemic isoforms. The four natural tocopherol isomers are d-α-, d-β-, d-γ-, and d-δ-tocopherol. The most abundant isoforms are α-tocopherol and γ-tocopherol, which differ by one methyl group (Fig. 1). Plants synthesize the natural isoforms from tyrosine and chlorophyll (40). Mammals consume plant tocopherols in food and cooking oils, and mammals do not interconvert the isoforms. Dietary tocopherols are taken up from the intestine, transported via the lymph to the blood, and then enter the liver. In the liver, α-tocopherol transfer protein (αTTP) preferentially transfers α-tocopherol to lipoproteins, resulting in 10-fold higher level of α-tocopherol than γ-tocopherol in tissues (92). Tocopherols in plasma lipoproteins are transferred to cells by plasma phospholipid transfer protein, scavenger receptors, or the lipoprotein lipase pathway (23). After uptake, the tocopherols are found in cell membranes as they are lipid vitamins. αTTP is also expressed by trophoblasts, fetal endothelium, and amnion epithelium of the placenta (59). It is reported that basal levels of α-tocopherol are required for placentation (44, 59).
Fig. 1.
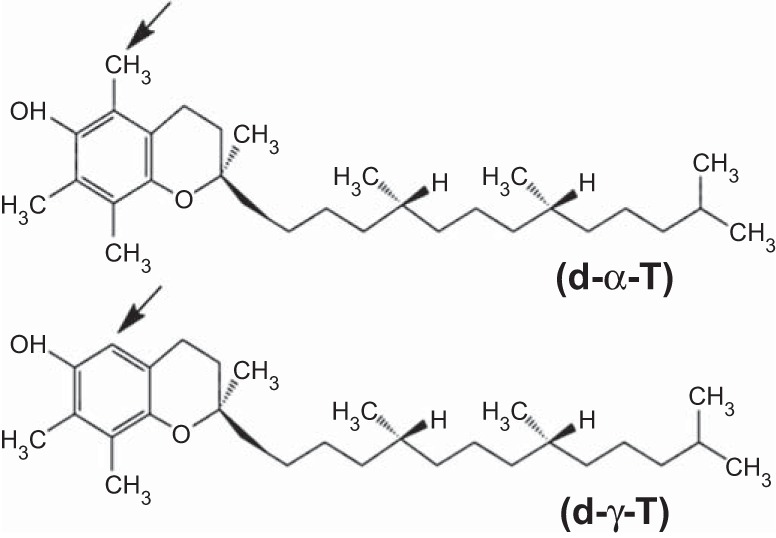
Schematic of d-γ-tocopherol and d-α-tocopherol which differ by one methyl group (arrow).
We have demonstrated in adults that α-tocopherol is anti-inflammatory and γ-tocopherol is proinflammatory (7, 22, 55). When presensitized allergic adult mice are supplemented with α-tocopherol and then challenged with antigen, allergic inflammation is decreased 65% whereas γ-tocopherol supplementation increases allergic inflammation by 175% (7). We demonstrated that α-tocopherol inhibits allergic responses and that, even at 10-fold lower concentrations than α-tocopherol, γ-tocopherol ablates the anti-inflammatory benefit of α-tocopherol during allergic responses in adult mice (7, 20). We also reported that α-tocopherol inhibits and γ-tocopherol increases leukocyte transendothelial migration, at least by, functioning as an antagonist and agonist, respectively, of endothelial cell PKCα (56). The opposing functions of these two tocopherol isoforms have implications for interpretations of studies with mixed tocopherols and we have reviewed alternative interpretations of the seemingly conflicting reports on vitamin E and the vitamin E isoforms in these studies (1, 16, 21, 22). Interestingly, countries that have reports of high human average plasma γ-tocopherol levels tend to have the highest prevalence rate of asthma (7, 24, 70, 73, 77, 87, 89). In the United States, average plasma γ-tocopherol levels are two to five times higher than those of European and Asian countries whereas the average plasma α-tocopherol levels are about the same among these countries (87). The plasma γ-tocopherol levels are high in the United States because the diet is rich in γ-tocopherol found in soy oil, corn oil, and canola oil (7, 11, 22, 60, 78). In contrast, γ-tocopherol is low in olive oil and sunflower oil used in European countries (7, 43, 87). The differences in outcomes from clinical reports on vitamin E and asthma in Europe and the Unites States may, in part, reflect the opposing functions of α-tocopherol and γ-tocopherol. We have demonstrated that in adult humans α-tocopherol associates with better lung spirometry and γ-tocopherol associates with lower lung spirometry by age 21 (53), suggesting that tocopherol isoforms may regulate development and lung responses to environmental pollutants, allergens, or infections. We demonstrated that supplementation of allergic female mice with α-tocopherol inhibited neonatal development of allergic responses (3). However, it is not known whether maternal γ-tocopherol regulates the development of allergic responses prenatally and early in life. We hypothesized that maternal supplementation with γ-tocopherol elevates the development of allergic responses in the offspring of allergic mothers.
We report here that supplementation of allergic female mice with γ-tocopherol elevated neonatal development of allergic responses. In addition, there was an increase in neonatal cytokines, chemokines, and lung CD11b+IRF4+ DC subsets, which are critical to development of allergic responses. γ-tocopherol also increased CD11b+ DC subsets in the fetal liver and in vitro in bone marrow differentiation of DCs. These data have important implications for development of allergic disease in offspring of allergic mothers consuming γ-tocopherol in their diet or prenatal vitamins and for infants consuming infant formulas supplemented with γ-tocopherol.
MATERIALS AND METHODS
Animals.
C57BL/6 mice were from Jackson Laboratories (Bar Harbor, ME). The studies are approved by the Northwestern University Institutional Review Committee for animals.
Tocopherol reagents.
d-γ-Tocopherol (>98% pure) from Alpha Chemistry was sent to Dyets (Bethlehem, PA) to generate the diets with 250 mg γ-tocopherol/kg diet (catalog no. 103742; Table 1). As in standard rodent chow, the basal diet contained 45 mg γ-tocopherol and 45 mg α-tocopherol/kg diet (catalog no. 101591; Dyets). The purity of these tocopherols that were used to make the diets was confirmed by HPLC with electrochemical detection as described below.
Table 1.
γ-Tocopherol-supplemented diet
| Diet, g/kg | |
|---|---|
| Casein, high nitrogen | 200 |
| l-Cystine | 3 |
| Sucrose | 100 |
| Cornstarch | 397 |
| Dyetrose | 132 |
| Corn oil | 70 |
| Cellulose | 50 |
| Mineral mix no. 210025 | 35 |
| Vitamin mix no. 210025 | 10 |
| Choline bitartrate | 2.5 |
| d-γ-Tocopherol | 0.25 |
The γ-tocopherol-supplemented diet (250 mg γ- tocopherol and 45 mg α-tocopherol/kg of diet; catalog no. 103742; modified AIN-93G Purified Rodent Diet with corn oil replacing soybean oil) and the basal control diet without supplementation (45 mg γ-tocopherol and 45 mg α-tocopherol/kg diet; no. 101591) were from Dyets. Corn oil, which was commonly used in rodent diet in the past, was used in this basal diet instead of the more recent formulas with soybean oil that contain high levels of γ-tocopherol.
Ovalbumin/tocopherol administration and inflammation.
C57BL/6 female mice were maintained on chow diet. Female mice at 4 to 6 wk old were sensitized by intraperitoneal injection (200 μl) of ovalbumin (OVA) grade V (5 μg)/alum (1 mg) or saline/alum (1 mg) on days 0 and 7 (28, 30, 50). The mice were exposed to nebulized saline or 3% (wt/vol) OVA in saline for 20 min three times a week during weeks 4, 8, 12, and 16. At the last challenge on week 16, the female mice were fed basal diet or diet supplemented with d-γ-tocopherol (250 mg γ-tocopherol/kg diet; Table 1) (48, 57, 79). Females were given the diets and mated with normal males. Three-day-old pups from five to six mothers per group were suboptimally sensitized by treating with only one 50-μl intraperitoneal injection (rather than two injections) of 5 μg OVA/1 mg alum (28, 30, 39). At 10, 11, and 12 days old, the pups were challenged for 15 min with 3% OVA (Fig. 2). At 13 days old, the pups were weighed and euthanized and tissues were collected. Serum was collected for analysis of IgE, liver was for tocopherol analysis, lung lavage was for leukocytes and cytokines, and lung tissue was for histology, flow cytometry, and quantitative PCR of mediators of allergic inflammation. A separate set of lungs were collected, minced, and cultured in HBSS with 0.2 mM CaCl2, 0.1 mM MgCl2, and 2 mM glutamine for 2 h. From these cultures, the supernatants were used for protein analysis by the 308L RayBiotech Array. Liver was extracted and examined for tocopherols by high-pressure liquid chromatography with electrochemical detection (HPLC/ECD) as we previously described (7, 55). Bronchoalveolar lavage (BAL) cells were stained and counted as previously described (4). OVA-specific IgE was determined by ELISA as previously described (14). Lung tissue sections were fixed in cold methanol for 15 min, rehydrated for 60 min with PBS and stained with eosin and methyl green as we previously described (6).
Fig. 2.
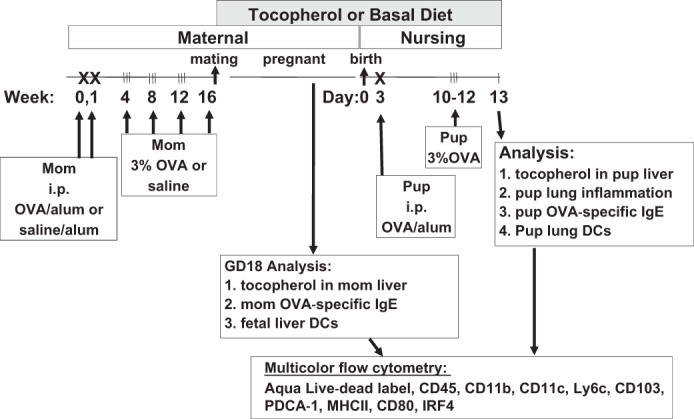
Schematic of the timeline for d-α-tocopherol diet and ovalbumin (OVA) treatment of mothers and pups. Mothers were sensitized and challenged with OVA or saline, and then at the time of breeding, they were given basal diet or diet supplemented with 250 mg d-γ-tocopherol (d-γ-T)/kg of diet. On day 3 after birth, the pups were sensitized once with OVA/alum by intraperitoneal injection and then challenged with aerosolized 3% OVA on days 10–12. On postnatal day (PND) 13, the pup livers were analyzed for tocopherol isoforms by HPLC and the pup lungs were analyzed for bronchoalveolar lavage (BAL) inflammation and lung tissue dendritic cell (DC) subsets by multicolor flow cytometry. With another set of mothers, on gestational day (GD) 18, tocopherols were determined in the liver of the mothers by HPLC and the fetal liver was analyzed for DC subsets.
Tocopherol measurement.
Diets or livers were weighed and homogenized in absolute ethanol with 5% ascorbic acid on ice. The internal standard tocol was added to each lung to determine recovery. The homogenate or plasma was extracted with an equal volume of hexane with 0.37 wt% butylated hydroxytoluene to prevent oxidation and increase recovery of tocopherol. The samples were vortexed and then centrifuged for 10 min at 9,000 g at 4°C. The hexane layer was dried under nitrogen and stored at −20°C. The samples were reconstituted in methanol and then tocopherols were separated using a reverse phase C18 HPLC column (Hewlett Packard) and HPLC (Waters) with 99% methanol-1% water as a mobile phase with detection with an electrochemical detector (potential 0.7 V; Waters).
Cytokines.
Supernatants from 2-h lung cultures were examined for cytokines by the L308 protein array (RayBiotech). Cytokines in lung lavage or 2-h lung supernatants were measured by ELISA (RayBiotech).
Flow cytometry analysis of pup lung, fetal liver, and cultured DCs.
Following bronchoalveolar lavage of pups, lungs were removed for analysis of DC types by flow cytometry. In the fetus, hematopoiesis occurs in the fetal liver. Gestational day 18 fetal livers were collected for analysis of fetal liver DCs. Briefly, tissues were minced and suspended in 5 ml of RPMI solution containing 1 mg/ml collagenase D (Roche) and 0.2 mg/ml DNAse I (Roche) 37°C with agitation. After 1 h, samples were filtered through sterile 70-μm nylon mesh, centrifuged, and resuspended in 10% FBS-RPMI media. Red blood cells were lysed in 1× BD PharmLyse Lysing Buffer (BD Biosciences) and the cells were washed once in 10% FBS-RPMI media. All centrifugation steps were carried out at 300 g for 5 min. Three million cells were used per sample for immunolabeling.
For bone marrow-derived DCs, a single cell suspension of bone marrow from 10-day-old mice was placed in RPMI-1640, glutamine, HEPES, gentamycin sulfate, pen/strep, and 10% heat-inactivated FCS. The bone marrow cells were treated with the DMSO solvent control (0.01%) or d-γ-tocopherol (2 μM) as we previously described for in vitro treatment with d-γ-tocopherol (7). The cells were stimulated with 5 ng granulocyte macrophage-colony-stimulating factor (GM-CSF)/ml. The culture medium with tocopherol and GM-CSF was replaced on days 2, 4, and 7. On day 8, the cells were suspended with Accutase (catalog no. SCR005; EMD Millipore), counted, plated (1 ml of 6.2 × 105 cells/ml in 12-well plates), and nonstimulated or stimulated overnight with 20 μg/ml house dust mite extract (catalog no. XPB82D3A2.5; Greer). On day 9, the cells were counted, immunolabeled with the markers described below for DCs, and examined by flow cytometry.
The cells were stained for live/dead exclusion in 500 μl PBS containing 0.25 μl of Aqua fluorescent reactive dye (no. L34957; Molecular Probes, Invitrogen) for 20 min at room temperature in the dark. The cells were washed and fixed by resuspension in 2% paraformaldehyde in PBS on ice for 30 min. Then, the cells were centrifuged and resuspended in permeabilization buffer (catalog no. 00-8333-56; eBioscience). Then, FC receptors were blocked by incubating the cells in 50 μl permeabilization buffer with 0.5 μl purified rat anti-mouse CD16/CD32 Mouse BD FC Block (no. 553142; BD Pharmingen) for 10 min at 4°C.
To prepare the antibodies for immunolabeling, for each sample equivalent, an antibody stock was prepared by adding the following antibodies to 50 μl of flow cytometry buffer: 1) CD45, rat anti-mouse Alexa 700-conjugated, 0.5 mg/ml, clone 30-F11 (no. 103127; Biolegend), 0.5 μl per sample; 2) CD11b, rat anti-mouse PE-CF594-conjugated, 0.2 mg/ml, clone M1/70 (no. 562317; BD Biosciences), 0.05 μl per sample; 3) Ly-6C, rat anti-mouse APC/Cy7-conjugated, 0.2 mg/ml, clone HK1.4 (no. 128025; Biolegend), 0.4 μl per sample; 4) CD11c, Armenian hamster anti-mouse PE/Cy7-conjugated, 0.2 mg/ml, clone N418 (no. 117317; Biolegend), 0.2 μl per sample; 5) I-A/I-E [myosin heavy chain II (MHCII)], rat anti-mouse PerCP/Cy5.5-conjugated, 0.2 mg/ml, clone M5/114.15.2 (no. 107625; Biolegend), 0.1 μl per sample; 6) CD103, Armenian hamster anti-mouse Brilliant Violet 421-conjugated, clone 2E7 (Biolegend no. 121421), 0.2 μl per sample; 7) CD317 (PDCA-1), rat anti-mouse APC-conjugated, 0.2 mg/ml, clone 927 (no. 127015; Biolegend), and 0.5 μl per sample; 8) CD80, Armenian hamster anti-mouse PE-conjugated, 0.2 mg/ml, clone 16-10A1 (no. 104707; Biolegend), 0.5 μl per sample; and 9) IRF4, rat anti-mouse FITC-conjugated, 0.5 mg/ml, clone 3E4 (no. 11-9858-82; eBioscience), 0.2 μl per sample. Then, 50 μl of the antibody stock were added to the samples that were treated with FC block. The samples were incubated for 30 min at 4°C in the dark. The cells were washed in flow cytometry staining buffer containing bovine calf serum and sodium azide (catalog no. 420201; Biolegend) and then resuspended and stored in 2% formaldehyde in PBS for analysis by flow cytometry.
The cells were examined with a BD LSRII flow cytometer (BD Biosciences). Analysis was performed using FlowJo VX software (TreeStar). Compensation was done using FlowJo compensation wizard based on single color control staining of compensation beads (eBioscience). Nonstained controls were used to assess boundaries of live and dead populations. Only live, singlet (based on FSC-H vs. FSC-A gating), hematopoietic (CD45+) cells were used for subsequent gating of all populations. Fluorescence Minus One (FMO) staining controls were used as negative controls to identify gates for populations of interest. The following subpopulations of DCs were analyzed: 1) resident cDCs: CD11b+Ly6C−CD11chighMHCIIhigh; 2) CD11b+ alveolar DCs: CD11b+Ly6C−CD11c+MHCII−; 3) monocyte-derived DCs: CD11b+Ly6C+CD11c+MHCII+; 4) plasmacytoid DCs: CD11b−Ly6C−CD11clowPDCA-1+MHCII−; 5) resident CD103+ DCs: CD11b−Ly6C−CD11c+CD103+MHCII−; and 6) CD11b− alveolar DCs: CD11b−Ly6C−CD11c+MHCII+/−. The flow cytometry gating strategy is as we previously described (3).
Statistics.
Data were analyzed by a one-way ANOVA followed by Tukey's or Dunnett's multiple comparisons test (SigmaStat; Jandel Scientific, San Ramon, CA). Presented are the means ± SE.
RESULTS
d-γ-Tocopherol supplementation of allergic female mice increases allergic inflammation in neonates.
It was determined whether d-γ-tocopherol supplementation of mothers during pregnancy/lactation increases development of allergic inflammation in their pups. In these studies, allergic responses were induced in adult female mice as in the timeline in Fig. 2. Then, at the time of mating (Fig. 2), the mothers were given a standard basal diet (45 mg d-γ-tocopherol and 45 mg d-α-tocopherol/kg diet) or supplemented with 250 mg d-γ-tocopherol/kg diets (Table 1). A basal tocopherol diet was used as the control diet instead of a tocopherol deficient diet because d-α-tocopherol is required for placenta development (44, 59). A dose of 250 mg d-γ-tocopherol/kg of diet is well within the range to elevate tissue tocopherols in adult rodents (57, 58, 76); this dose is 30 times lower than the maternal tocopherol rodent diet dose that reduces rodent hippocampus function (8). Moreover, the dose yields a two- to threefold increase in liver γ-tocopherol in pups and adult mice (Fig. 3) which is similar to the fold changes achievable in the human population (2, 7, 20, 22, 55, 56). All pups from the mothers were suboptimally sensitized with OVA-alum and then received three OVA challenges (Fig. 3). It was determined whether d-γ-tocopherol-supplemented diets increased liver d-γ-tocopherol concentrations in mothers and pups because, in the liver, d-γ-tocopherol is loaded onto lipoproteins that then enter circulation for delivery to peripheral tissues. The d-γ-tocopherol-supplemented diet significantly increased liver d-γ-tocopherol in the saline-treated mothers twofold compared with basal diet controls (Fig. 3A). OVA treatment reduced the d-γ-tocopherol tissue concentrations in the d-γ-tocopherol-supplemented mothers (Fig. 3A); this is consistent with lower tocopherol levels in asthmatics compared with nonasthmatic saline mothers and this was reduced in pups from allergic mothers (Fig. 3B). With maternal d-γ-tocopherol supplementation, there were up to half as many mated females that gave birth (Fig. 4A) and for those with litters, there was not a significant effect on litter sizes or pup body weight (Fig. 4, B and C).
Fig. 3.
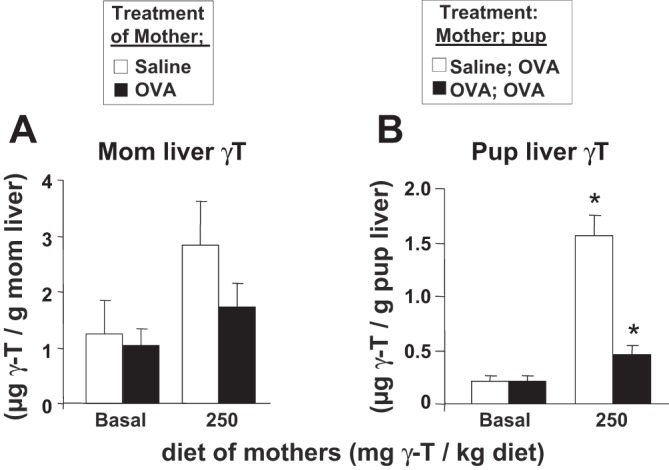
Mother and pup liver γ-tocopherol levels. Mice were treated as in Fig. 2. A: on GD18, concentrations of γ-tocopherol (γ-T) were determined in the liver of the mothers by HPLC. B: on PND13, the pup livers were analyzed for γ-tocopherol isoforms by HPLC. *P < 0.05, compared with all groups; n = 8−10 mice/group.
Fig. 4.

Maternal γ-tocopherol reduced the numbers of mated females that had pups but did not alter pup numbers per mother or pup body weight on day 13. Mice were treated as in Fig. 2. The females were mated for 48 h. A: percentage of mated females that had pups. B: average number of pups per mother that had pups. C: pup body weight at PND13. *P < 0.05, compared with corresponding basal diet group; n = 4 experiments.
Leukocytes were examined in the OVA-challenged pup lungs. Compared with OVA-challenged pups from nonallergic mothers (saline-treated) fed basal diet, the OVA-treated pups from allergic mothers with basal diet had a significant increase in number of BAL eosinophils and monocytes and an increase in the relatively low numbers of BAL lymphocytes and neutrophils (Fig. 5A); this is consistent with previous reports (3, 50). Interestingly, with γ-tocopherol supplementation of the allergic mothers, there was a twofold increase in allergic inflammation in the OVA-challenged pups compared with OVA-challenged pups from allergic mothers with basal diet (Fig. 5A). d-γ-Tocopherol supplementation of allergic female mice also increased the numbers of eosinophils in the lung tissue of OVA-challenged pups (Fig. 5D). The allergic mothers and OVA-challenged pups from allergic mothers had increased serum IgE, but serum IgE was not affected by d-γ-tocopherol supplementation (Fig. 5, B and C). Because inflammation is regulated by Th2 cytokines and chemokines, we analyzed protein expression of IL-5 and eotaxins in the lung lavage. OVA-challenged pups from allergic mothers with basal diet had a two- to threefold increase in CCL11, CCL24, and IL-5 compared with OVA-challenged pups from nonallergic mothers with basal diet (Fig. 6). CCL11, CCL24, and IL-5 was increased twofold in the lung lavage from the OVA-challenged pups from nonallergic mothers with γ-tocopherol-supplemented diet compared with OVA-challenged pups from nonallergic mothers with basal diet (Fig. 6). The γ-tocopherol supplementation of allergic mothers did not further elevate the lung lavage CCL11, CCL24, or IL-5 in their pups compared with that for pups from allergic mothers with basal diet (Fig. 6), suggesting that a maximum response may have been achieved in pups from allergic mothers.
Fig. 5.

γ-Tocopherol supplementation increased lung eosinophils in OVA-challenged pups from allergic mothers. Mice were treated as in Fig. 2. A: bronchoalveolar lavage (BAL) leukocytes of PND13 OVA-challenged pups from allergic and saline (nonallergic) control mothers with basal diet or γ-tocopherol-supplemented diet. *P < 0.05 compared with OVA/basal group. **P < 0.05, compared with all groups; n = 8–10 pups/group. B: PND13 OVA-specific serum IgE as determined by ELISA; n = 8–10 mice/group. *P < 0.05, compared with the corresponding basal groups; n = 8–10 pups/group. C: mother OVA-specific serum IgE on gestational day 18 as determined by ELISA; n = 10–15 mothers/group. *P < 0.05, compared with the corresponding basal groups. D: representative micrographs of perivascular regions in pup lung tissue stained with eosin and methyl green. Images were obtained with a ×20 objective on an Olympus Microscope. Arrows on the images indicate some of the eosin-labeled perivascular eosinophils. V, vessel lumen.
Fig. 6.
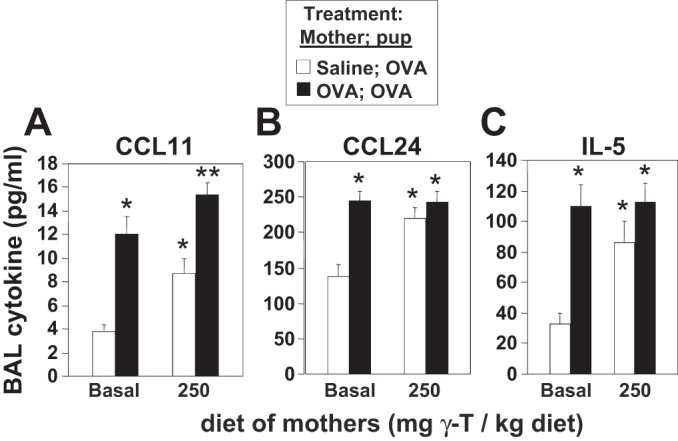
Maternal supplementation with d-γ-tocopherol increased CCL11, CCL24, and IL-5 in the lung lavage of OVA-challenged pups. BAL supernatants were collected from PND13 OVA-challenged pups from allergic and saline (nonallergic) control mothers with basal diet or γ-tocopherol-supplemented diet. The indicated cytokines and chemokines were analyzed by ELISAs. A: CCL11. B: CCL24. C: IL-5l. *P < 0.05, compared with basal-saline group. **P < 0.05, compared with all saline groups; n = 8–10 mice/group.
We also cultured minced lungs from the pups in Fig. 5 for 2 h and collected the supernatants for analysis of expression of inflammatory mediators. Using the L308 RayBiotech protein array, we analyzed protein expression in the lung culture supernatants from OVA-challenged pups from allergic mothers with γ-tocopherol-supplemented diet compared with lung supernatants of OVA-challenged pups from allergic mothers with basal diet. d-γ-Tocopherol supplementation of allergic mothers elevated OVA-induced pup lung protein expression of several mediators in Fig. 7A. Ratios above 1.5 are considered by the assay as significant for further analysis. Therefore, we focused on secreted mediators for ELISA analysis. ELISA analysis of amphiregulin in the supernatants demonstrated a 150% increase in amphiregulin in OVA-challenged pups from allergic mothers with γ-tocopherol supplementation compared with OVA-challenged pups from allergic mothers with basal diet (Fig. 7B). Amphiregulin was increased in lungs of OVA-challenged pups from allergic mothers with basal diet compared with OVA-challenged pups from nonallergic mothers with basal diet (Fig. 7B). Activin A was increased 145% in groups with d-γ-tocopherol supplementation compared with groups with basal diet as determined by ELISA (Fig. 7C). In the GM-CSF ELISA, lung supernatant GM-CSF was near the detection limits of ELISAs. Nevertheless, there was increased GM-CSF in lung supernatants of OVA-challenged pups from allergic mothers with γ-tocopherol-supplemented diet compared with OVA-challenged pups from nonallergic mothers with basal diet (Fig. 7D). Therefore, d-γ-tocopherol supplementation of allergic mothers elevated OVA-stimulated pup lung allergic inflammation (Fig. 5) and several cytokine/chemokine mediators of allergic inflammation (Figs. 6 and 7).
Fig. 7.

Maternal supplementation with γ-tocopherol increased mediators in OVA-challenged pup lungs from allergic mothers. Supernatants from 2-h cultures were from 3 OVA-challenged PND13 pup lungs from allergic mothers with γ-tocopherol-supplemented or basal diet. A: supernatants were analyzed with the RayBiotech L-308 array that detects 308 growth factors/cytokines/chemokines. Shown is fold change: the relative expression of the protein from lungs of OVA-challenged pups from γ-tocopherol-supplemented allergic moms divided by the relative expression of the protein for lungs of OVA-challenged pups from allergic moms with basal diet. A value above 1.5 are considered by RayBiotech as candidates for ELISA analysis; n = 6 arrays (3 mice × 2 groups). B: supernatants from the 2-h lung cultures were analyzed for amphiregulin protein by ELISA. C: supernatants from the 2-h lung cultures were analyzed for activin A protein by ELISA. D: supernatants from the 2-h lung cultures were analyzed for GM-CSF protein by ELISA. *P < 0.05, compared with basal-saline group. **P < 0.05, compared with all groups; n = 8–10 mice/group.
d-γ-Tocopherol supplementation of allergic female mice increases numbers of CD11b+ DC subsets in the fetal liver and in neonates.
It is reported that DCs, but not macrophages, are critical for the development of allergic responses in the pups from allergic mothers (26). Therefore, we determined whether d-γ-tocopherol regulates the development of DCs in postnatal day 13 neonates and gestational day 18 fetuses from allergic mothers in the timeline in Fig. 2. The cells of the postnatal day 13 neonate lungs or gestational day 18 fetal livers were dissociated and immunolabeled as previously described (3, 64). The flow cytometry analysis scheme for pup lung DCs was as we previously described (3). There were CD11b+ and CD11b− DC subsets in the neonatal lungs but d-γ-tocopherol supplementation only altered the CD11b+ DC subsets. Therefore, these two DC lineages are described here. Briefly, the following CD11b+ DC subsets were present in the postnatal day 13 OVA-challenged neonate lungs (Fig. 8A) (3): resident DCs, monocyte-derived DCs, and alveolar DCs. The following CD11b− DC subsets were present in the postnatal day 13, OVA-challenged neonate lungs (Fig. 8A) (3): plasmacytoid DCs, resident CD103+ DCs, alveolar DCs, and alveolar macrophages. We also examined IRF4 expression in these DC subsets because allergen challenge of the lung induces DC expression of IRF4 that drives the Th2 phenotype that is critical for allergic lung responses (90). Interestingly, compared with the OVA-challenged pups from nonallergic mothers with basal diet, the OVA-challenged pups from allergic mothers with basal diet had increased lung tissue numbers of mDCs, alveolar DCs, IRF4+ alveolar DCs, resident DCs, and IRF4+ resident DCs (Fig. 8B). Supplementation of allergic mothers with d-γ-tocopherol significantly increased twofold the OVA-challenged pup lung tissue number of mDCs, IRF4+ mDCs, alveolar DCs, IRF4+ alveolar DCs, resident DCs, and IRF4+ resident DCs (Fig. 8B). In contrast to the CD11c+CD11b+ DCs, d-γ-tocopherol supplementation of the mothers did not alter the pup lung CD11c+CD11b− subsets of cells, including pDCs, CD103+ resident DCs, CD11b− alveolar DCs, and alveolar macrophages (Fig. 8C). For CD11c+CD11b+ DC subsets (Fig. 9) and CD11c+CD11b− DC subsets (data not shown), the mean fluorescence intensity for MHCII, CD80, and IRF4 was not different among the groups.
Fig. 8.

d-γ-Tocopherol supplementation of allergic female mice increased the numbers of CD11c+CD11b+ DCs in the pup lung. The lung tissues were from the pups treated as in Fig. 2. Lung DCs were isolated, immunolabeled, and examined by flow cytometry. A: chart of lung CD11c+ subsets analyzed in the pup lungs. B: numbers of CD11b+ DC subsets per million lung cells. C: numbers of CD11b− DC subsets per million lung cells. Presented is the mean ± SE; n = 8–10 mice/group. *P < 0.05, compared with basal-saline group. **P < 0.05, compared with all other groups.
Fig. 9.
d-γ-Tocopherol supplementation of allergic female mice did not alter the expression of MHCII, CD80, or IRF4 on the CD11c+CD11b+ pup lung DCs. The pup lung cells were those analyzed in Fig. 8. Presented is the mean fluorescence intensity (MFI) for MHCII, CD80, or IRF4. A: MFI of MHCII on mDCs. B: MFI of MHCII on alveolar DCs. C: MFI of MHCII on resident DCs. D: MFI of CD80 on mDCs. E: MFI of CD80 on alveolar DCs. F: MFI of CD80 on resident DCs. G: MFI of IRF4 in mDCs. H: MFI of IRF4 in alveolar DCs. I: MFI of IRF4 on resident DCs. Presented is the mean ± SE; n = 8−10 mice/group.
Dendritic cell subsets in the fetal liver were determined on gestational day 18 as in the timeline in Fig. 2. Gestational day 18 was analyzed because mouse fetal livers begin hematopoiesis at about gestational day 16 and pups are born gestational day 21. The flow cytometry analysis scheme for fetal liver DCs is as previously described (3). The following CD11c+CD11b+ DC phenotypes were present in the gestational day 18 fetal livers (Fig. 10A) (3): monocyte-derived DCs and resident DCs. The following CD11c+CD11b− cell phenotypes were present in the gestational day 18 fetal livers (Fig. 10A) (3): Ly6c+CD103-PDCA+MHCII+ cells and Ly6c-CD103low PDCA+MHCII+ cells. Interestingly, d-γ-tocopherol supplementation of allergic mothers resulted in twofold increases in the number of CD11c+CD11b+ DCs in gestational day 18 fetal livers, including those of the phenotype of mDCs (Fig. 10, Ca-Cc) and phenotype of resident DCs (Fig. 10, Dg-Di). Maternal supplementation of d-γ-tocopherol did not alter the expression of the stimulatory receptors (MHCII and CD80) and the transcription factor IRF4 in the mDCs or resident DCs (Fig. 10, Cd-Cf, and Dj-Dl). Maternal supplementation of d-γ-tocopherol reduced by 50% the numbers of fetal liver CD11c+CD11b-Ly6c+CD103-PDCA+MHCII+ cells in the fetal liver of allergic and nonallergic mothers (Fig. 10B).
Fig. 10.

d-γ-Tocopherol supplementation of allergic female mice increased the numbers of CD11c+CD11b+ DCs in the fetal liver. The mice were treated as in Fig. 2 and fetal livers were collected on GD18. The liver cells were isolated, immunolabeled and examined by flow cytometry. A: chart of lung CD11c+ subsets analyzed in the GD18 fetal liver. B: numbers of CD45+CD11c+CD11b-Ly6c+CD103-PDCA+MHCII+ plasmacytoid DC phenotype in the fetal liver. *P < 0.05, compared with corresponding mothers with basal diet. Ca-Cc: numbers of mDC subsets per million fetal liver cells. Cd-Cf: MFI of MHCII, CD80 and IRF4 on fetal liver cells with the mDC phenotype. Dg-Di: numbers of resident DC subsets per million fetal liver cells. Dj-Dl: MFI of MHCII, CD80, and IRF4 on fetal liver cells with the resident DC phenotype. Presented is the mean ± SE. *P < 0.05, compared with basal groups. **P < 0.05 compared with all other groups; n = 8–10 mice/group.
To determine whether d-γ-tocopherol can directly regulate differentiation of DCs, we determined whether d-γ-tocopherol supplementation of bone marrow in vitro increased development of bone marrow-derived CD11b+ DCs. For these studies, bone marrow from 10-day-old neonates from mothers with basal diet was incubated with GM-CSF for 8 days in vitro in the presence of d-γ-tocopherol or the solvent control DMSO. Then the cells were stimulated with house dust mite extract or nonstimulated. The treatments did not affect the percentage of viable cells in the culture as determined by live/dead Aqua fluorescent staining and flow cytometry (data not shown). In nonstimulated cultures, d-γ-tocopherol increased the number of IRF4+CD45+ CD11b+CD11c+ MHCII+ DCs (Fig. 11). In the d-γ-tocopherol-treated house dust mite-stimulated group, the number of IRF4+CD45+ CD11b+CD11c+ MHCII+ DCs was highly elevated by threefold compared with the other groups (Fig. 11). In summary, d-γ-tocopherol increased the number of CD11b+CD11c+ DCs in vitro, in the fetal liver, and in the neonate.
Fig. 11.
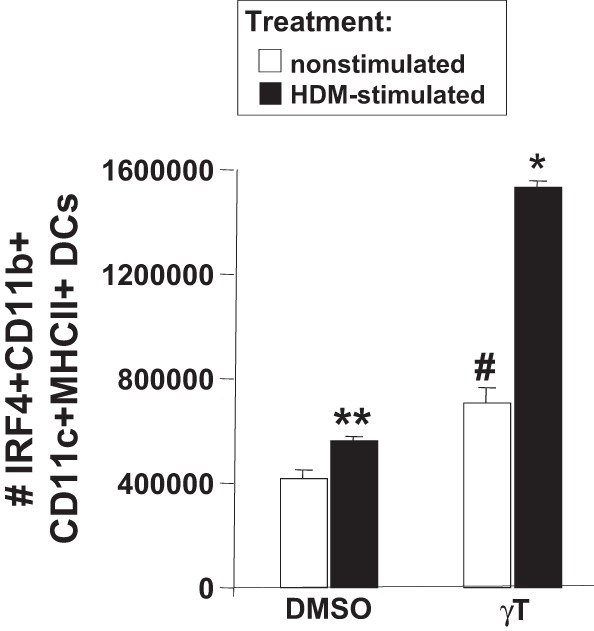
d-γ-Tocopherol increased the generation of bone marrow-derived IRF4+CD11c+CD11b+ DCs in vitro. Bone marrow from PND10 pups with basal diet was cultured for 10 days with GM-CSF in the presence of 0.01% DMSO (solvent control) or 2 μM d-γ-tocopherol (as we previously described for in vitro cell loading with d-α-tocopherol) (7). The cells were nonstimulated or stimulated with 20 μg/ml house dust mite (HDM) extract overnight. The DCs were analyzed by immunolabeling and flow cytometry. Presented is the number of IRF4+CD45+CD11b+CD11c+ MHCII+ DCs. There was no difference in the %live cells between the 2 groups (data not shown). There were no cells from the culture with the monocyte-derived phenotype (CD45+CD11c+CD11b+Ly6c+, MHCII high; data not shown); n = 5–6 from a representative experiment of 2 experiments. Presented is the mean ± SE. *P < 0.05, compared with all groups. **P < 0.05, compared with nonstimulated-DMSO group. #P < 0.05, compared with nonstimulated-DMSO control group and compared with HDM-stimulated γ-tocopherol-treated group.
DISCUSSION
In this report, the development of allergic inflammation in pups from allergic mothers was amplified by supplementation of the mother with d-γ-tocopherol. d-γ-tocopherol did not induce allergic inflammation because it did not increase allergic inflammation in the pups from nonallergic mothers. In pups from allergic mothers, maternal d-γ-tocopherol supplementation increased inflammatory mediators including the Th2 mediator amphiregulin, IL-5, CCL11, CCL24, activin A, and GM-CSF. d-γ-Tocopherol supplementation of the allergic mothers increased generation of IRF4+CD11c+CD11b+ DCs in the fetal liver and in antigen-challenged pup lungs. d-γ-tocopherol has, at least, a direct effect on hematopoietic development of DCs because d-γ-tocopherol increased the generation of IRF4+CD11c+CD11b+ bone marrow-derived DCs in vitro. In addition, d-γ-tocopherol decreased the numbers of inhibitory PDCA+ plasmacytoid CD11c+CD11b− DCs in the fetal liver. There was no effect of d-γ-tocopherol on the level of expression of MHCII, CD80, or IRF4 by the fetal liver and pup lung DCs. Maternal d-α-tocopherol supplementation also decreased the numbers of mated females that had pups but did not affect numbers of pups per litter or pup body weight. This is the first report that maternal d-γ-tocopherol supplementation of allergic mothers increased development of allergic responses in offspring and regulates numbers of select DC subsets. These results have important implications for vitamin E regulation of the development of allergic disease in future generations.
It has been suggested that early exposures to environmental factors increase risk of allergic disease (9). In reports examining human maternal and paternal asthma associations with development of allergies in offspring, most associations are with maternal asthma (50). In animal studies, offspring of allergic mothers have increased responses to suboptimal doses of allergens (28, 30, 35, 36, 38, 39, 42). The sensitization to allergens and allergic responses are dependent on DCs, which produce regulatory cytokines (80, 90). Interestingly, there are functional changes in the DCs from offspring of allergic mothers (26). Maternal exposure to environmental factors, including high-fat diets, can alter neonatal hematopoietic or metabolic function (15, 27, 41, 51, 61, 66, 83, 93). In our studies, maternal supplementation with d-γ-tocopherol increased development of CD11c+CD11b+ DCs in the fetus and in neonates. In our studies, maternal supplementation with d-γ-tocopherol raised the maternal liver γ-tocopherol level twofold and the pup liver γ-tocopherol fivefold, consistent with the fold tocopherol changes in human and mouse tissues after supplementation (2, 7, 20, 22, 55, 56). In the past 50 yr, there has been an increase in the γ-tocopherol form of vitamin E in the diet, in infant formulas, and in vitamin supplements (22, 78). This may, at least in part, contribute to the increase in incidence of allergic disease during this time period.
Basal levels of α-tocopherol are required for placentation (44, 59), but it is not known whether γ-tocopherol influences placentation. In our studies, although basal α-tocopherol was in all diets, supplementation with γ-tocopherol decreased the numbers of mated females with pups, suggesting that γ-tocopherol has an opposing function during development. Whether γ-tocopherol regulates placenta development or other functions during development is not known and is being addressed in ongoing studies. Nevertheless, the reduced numbers of mothers with pups with γ-tocopherol supplementation have potential important implications for γ-tocopherol in diet and supplements during pregnancy.
In our studies, maternal d-γ-tocopherol supplementation increased the number of pup lung CD11b+ DC subsets but not number of pup lung CD11b− DC subsets. This specificity of γ-tocopherol regulation of the CD11b+ DC subsets suggests that tocopherols may regulate signals for DC differentiation of these DC subsets or regulate signals for expression of CD11b. The generation of CD11b+ DCs is regulated by signals and several transcription factors that bind the CD11b promoter (82). Some signals that regulate expression of CD11b on DCs include GM-CSF (94), ERK5 (88), aldehyde dehydrogenase (94), and retinoic acid (5, 47, 71, 74, 94). There was increased GM-CSF expression in the OVA-challenged pup lungs from d-γ-tocopherol-supplemented allergic mothers. The CD11b promoter has binding sites for the transcription factors Sp1, Vav1/PU.1, ets, AP-2, and RAR (13, 37). Whether γ-tocopherol regulates GM-CSF signals or regulates transcription factors that activate the CD11b promoter in DCs is under investigation.
There was also altered CD11b− CD11c+ pDCs in the fetal liver with γ-tocopherol supplementation. In the gestational day 18 fetal liver of γ-tocopherol-supplemented mothers, there were fewer regulatory pDCs suggesting that with γ-tocopherol supplementation, there may be a reduced control of magnitude of responses to allergen challenge early in life. However, in the postnatal day 13 OVA-challenged pup lung, the number of pDCs was not altered with γ-tocopherol supplementation. We also observed that supplementation with γ-tocopherol partially increased mediators of allergic inflammation and DC subsets in offspring of nonallergic mothers. Maternal d-γ-tocopherol supplementation partially increased numbers of resident DCs in the fetus and OVA-challenged pup lung from nonallergic mothers but not to the extent as in the OVA-challenged pups from d-γ-tocopherol-supplemented allergic mothers. This is consistent with the increased GM-CSF with γ-tocopherol supplementation in pup lungs from allergic mothers. d-γ-Tocopherol also increased activin A in pups from nonallergic and allergic mothers. Activin A is a member of the TGF-β superfamily of cytokines and regulates allergic inflammation (32). Activin A is produced by several cell types including epithelium, endothelium, mast cells, fibroblasts, and DCs and it can induce differentiation of monocytes to mDCs and recruitment of DCs (33). Therefore, activin A may function in concert with other mediators to increase numbers of DCs and allergic inflammation. d-γ-Tocopherol increased CCL11 in pups from allergic and nonallergic mothers and it increased CCL24, and IL-5 in the pups from nonallergic mothers. OVA challenge in pups from allergic mothers with d-γ-tocopherol did not result in further increases in CCL24 or IL-5, which may indicate that a maximum response was achieved with OVA challenge. Nevertheless, the pups from the allergic mothers with d-γ-tocopherol had elevated CCL11 and amphiregulin suggesting that in combination these signals as well as the presence of GM-CSF, CCL24, IL-5, and activin A resulted in amplification of recruitment of eosinophils in pups from allergic mothers with d-γ-tocopherol-supplemented diets. Our ongoing studies are determining mechanisms for γ-tocopherol regulation of DCs and signals for allergic inflammation during development.
Reports conflict as to whether vitamin E modulates inflammation. We have demonstrated opposing functions of d-α-tocopherol and d-γ-tocopherol in vivo and in vitro and demonstrated that d-γ-tocopherol is an agonist of PKCα, whereas d-α-tocopherol is an antagonist of PKCα upon binding to the C1A regulatory domain of PKCα (56). Interestingly, γ-tocopherol negates the anti-inflammatory benefit of α-tocopherol (7, 20). This strong opposing function of γ-tocopherol occurs even though γ-tocopherol is about 5–10 times lower in concentration in vivo than α-tocopherol. The proinflammatory effects of γ-tocopherol in adult allergic mice are partially reversed by switching from γ-tocopherol to α-tocopherol (55). Thus we have demonstrated opposing and competing functions of α-tocopherol and γ-tocopherol in vivo. Okamoto et al. (62) found that feeding mice α-tocopherol starting 2 wk before antigen sensitization did not affect IgE levels but did reduce the number of eosinophils in the BAL, but the form and purity of α-tocopherol were not indicated. Our studies with maternal supplementation of d-γ-tocopherol also did not alter pup OVA-specific IgE, suggesting that the regulatory function of tocopherols in allergic inflammation is likely through mechanisms other than regulation of production of IgE. Mabalirajan et al. (52) reported that oral administration of α-tocopherol in ethanol after antigen sensitization blocked OVA-induced lung inflammation and airway hyperresponsiveness. In a report by Suchankova et al. (72), purified α-tocopherol was administered in soy oil by gavage and they found no major effect of α-tocopherol on immune parameters or lung airway responsiveness in mice challenged with OVA. However, the soy oil vehicle in this study contains an abundance of γ-tocopherol, which can oppose the function of α-tocopherol. In their study, neither tissue tocopherol levels nor vehicle tocopherol levels were reported. Besides d-γ-tocopherol scavenging of reactive oxygen species and d-γ-tocopherol agonist function for PKCα, d-γ-tocopherol can also scavenge reactive nitrogen species (RNS) (63). γ-Tocopherol scavenging of RNS may be beneficial for inflammation with increases in RNS such as neutrophilic inflammation that is induced by ozone or endotoxin in mice (25). Consistent with this, reports indicate that supplementation with a mixture of tocopherols enriched for γ-tocopherol blocks acute endotoxin-stimulated or ozone-stimulated neutrophil inflammation in the rat and human lung (34, 85, 91). In another study, γ-tocopherol supplementation reduced antigen induction of rat lung inflammation in which there was severalfold more neutrophils than eosinophils (86). It is also reported that nebulized γ-tocopherol reduces neutrophilia in burn and smoke inhalation injury in sheep (31). Therefore, γ-tocopherol may be of benefit for acute neutrophilic inflammation. In contrast, it is reported that chronic higher plasma γ-tocopherol associates with lower lung function in humans (53) and with higher lung eosinophilia and airway hyperresponsiveness in mouse models of allergic asthma (53). We have also reported in adult mice that switching allergic mice from d-γ-tocopherol to d-α-tocopherol partially reverse the elevated inflammation of the allergic mice with d-γ-tocopherol (55). Therefore, differences among the reports for tocopherol regulation of eosinophilic lung inflammation reflect differences in the intake of tocopherol isoforms and acute vs. chronic inflammation.
Most clinical studies on vitamin E include mixed forms of natural and synthetic tocopherols from supplementation or diet. We recently demonstrated in adult mice that γ-tocopherol increases allergic responses and that α-tocopherol inhibits allergic responses (7, 20). We also recently reported in a 20-yr prospective study with 4,500 individuals that α-tocopherol associates positively with adult human lung spirometry [forced expiratory volume in 1 s (FEV1)] and γ-tocopherol associates negatively with human lung spirometry (FEV1), with high levels of γ-tocopherol associating with up to 300 ml lower FEV1 by age 20 (53), suggesting tocopherol regulation during development. We have demonstrated that α-tocopherol supplementation of allergic adult female mice during pregnancy and nursing reduces development of allergic responses in neonatal mice (3). Our studies herein demonstrate that d-γ-tocopherol supplementation of allergic adult female mice increased the generation of allergic disease in offspring exposed to allergen and that d-γ-tocopherol increased hematopoiesis of CD11c+CD11b+ DCs in the offspring. Thus the opposing regulatory effects of these two tocopherol isoforms have implications on interpretation of clinical studies with mixed tocopherols and we have written reviews discussing these implications (1, 16, 18, 21, 22).
In summary, we identified novel regulatory functions for d-γ-tocopherol. Maternal supplementation with d-γ-tocopherol increased allergic responses in offspring from allergic mothers and increased development of CD11c+CD11b+ DC types in utero, in antigen-challenged neonate lungs, and in bone marrow cultures Moreover, there was specificity of regulation of DCs by d-γ-tocopherol because d-γ-tocopherol supplementation increased the number of CD11c+CD11b+ but not CD11c+CD11b− DC types in pup lungs. There was also specificity of regulation of DCs in utero because maternal d-γ-tocopherol increased fetal liver CD11c+CD11b+ DCs and decreased numbers of plasmacytoid regulatory DCs in the fetal liver. The d-γ-tocopherol supplementation also elevated several mediators of inflammation without altering OVA-specific IgE. These studies of d-γ-tocopherol regulation of inflammation provide a basis towards designing drugs, supplements, and diets that more effectively modulate these pathways in allergic disease. Information from our studies will have potential impact on design of future clinical studies with vitamin E isoforms and on our understanding of vitamin E isoform regulation of DC function during allergic inflammation. Moreover, these studies may have implications for dietary impact on risk for allergic disease in future generations. More studies are needed in humans to examine short term vs. long term outcomes of the range of plasma concentrations of tocopherol isoforms.
GRANTS
These studies were supported by National Institutes of Health Grants R01-AT-004837 and R01-HL-111624 (both to J. M. Cook-Mills).
DISCLOSURES
No conflicts of interest, financial or otherwise are declared by the author(s).
AUTHOR CONTRIBUTIONS
Author contributions: H.A.-V., F.S., and J.M.C.-M. performed experiments; H.A.-V., F.S., and J.M.C.-M. analyzed data; H.A.-V., F.S., and J.M.C.-M. interpreted results of experiments; H.A.-V. and J.M.C.-M. prepared figures; H.A.-V., F.S., and J.M.C.-M. edited and revised manuscript; H.A.-V., F.S., and J.M.C.-M. approved final version of manuscript; J.M.C.-M. conception and design of research; J.M.C.-M. drafted manuscript.
REFERENCES
- 1.Abdala-Valencia H, Berdnikovs S, Cook-Mills JM. Vitamin E isoforms as modulators of lung inflammation. Nutrients 5: 4347–4363, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Abdala-Valencia H, Berdnikovs S, Cook-Mills JM. Vitamin E isoforms differentially regulate intercellular adhesion molecule-1 activation of PKCalpha in human microvascular endothelial cells. PLoS One 7: e41054, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Abdala-Valencia H, Berdnikovs S, Soveg F, Cook-Mills JM. α-Tocopherol supplementation of allergic female mice inhibits development of CD11c+CD11b+ dendritic cells in utero and allergic inflammation in neonates. Am J Physiol Lung Cell Mol Physiol 307: L482–L496, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Abdala-Valencia H, Earwood J, Bansal S, Jansen M, Babcock G, Garvy B, Wills-Karp M, Cook-Mills JM. Nonhematopoietic NADPH oxidase regulation of lung eosinophilia and airway hyperresponsiveness in experimentally induced asthma. Am J Physiol Lung Cell Mol Physiol 292: L1111–L1125, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Beijer MR, Kraal G, den Haan JM. Vitamin A and dendritic cell differentiation. Immunology 142: 39–45, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Berdnikovs S, Abdala-Valencia H, Cook-Mills JM. Endothelial cell PTP1B regulates leukocyte recruitment during allergic inflammation. Am J Physiol Lung Cell Mol Physiol 304: L240–L249, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Berdnikovs S, Abdala-Valencia H, McCary C, Somand M, Cole R, Garcia A, Bryce P, Cook-Mills J. Isoforms of vitamin e have opposing immunoregulatory functions during inflammation by regulating leukocyte recruitment. J Immunol 182: 4395–4405, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Betti M, Ambrogini P, Minelli A, Floridi A, Lattanzi D, Ciuffoli S, Bucherelli C, Prospero E, Frontini A, Santarelli L, Baldi E, Benetti F, Galli F, Cuppini R. Maternal dietary loads of alpha-tocopherol depress protein kinase C signaling and synaptic plasticity in rat postnatal developing hippocampus and promote permanent deficits in adult offspring. J Nutr Biochem 22: 60–70, 2011. [DOI] [PubMed] [Google Scholar]
- 9.Bousquet J, Anto J, Auffray C, Akdis M, Cambon-Thomsen A, Keil T, Haahtela T, Lambrecht BN, Postma DS, Sunyer J, Valenta R, Akdis CA, Annesi-Maesano I, Arno A, Bachert C, Ballester F, Basagana X, Baumgartner U, Bindslev-Jensen C, Brunekreef B, Carlsen KH, Chatzi L, Crameri R, Eveno E, Forastiere F, Garcia-Aymerich J, Guerra S, Hammad H, Heinrich J, Hirsch D, Jacquemin B, Kauffmann F, Kerkhof M, Kogevinas M, Koppelman GH, Kowalski ML, Lau S, Lodrup-Carlsen KC, Lopez-Botet M, Lotvall J, Lupinek C, Maier D, Makela MJ, Martinez FD, Mestres J, Momas I, Nawijn MC, Neubauer A, Oddie S, Palkonen S, Pin I, Pison C, Rance F, Reitamo S, Rial-Sebbag E, Salapatas M, Siroux V, Smagghe D, Torrent M, Toskala E, van Cauwenberge P, van Oosterhout AJ, Varraso R, von Hertzen L, Wickman M, Wijmenga C, Worm M, Wright J, Zuberbier T. MeDALL (Mechanisms of the Development of ALLergy): an integrated approach from phenotypes to systems medicine. Allergy 66: 596–604, 2011. [DOI] [PubMed] [Google Scholar]
- 10.Bousquet J, Bousquet PJ, Godard P, Daures JP. The public health implications of asthma. Bull World Health Organ 83: 548–554, 2005. [PMC free article] [PubMed] [Google Scholar]
- 11.Boyle FG, Yuhas RJ, Lien EL. Red blood cell and tissue phospholipid fatty acid profiles of weanling rats fed infant formula fat blends containing soy and/or corn oil. Ann Nutr Metab 40: 234–242, 1996. [DOI] [PubMed] [Google Scholar]
- 12.Brigelius-Flohe R, Traber MG. Vitamin E: function and metabolism. FASEB J 13: 1145–1155, 1999. [PubMed] [Google Scholar]
- 13.Brugnoli F, Lambertini E, Varin-Blank N, Piva R, Marchisio M, Grassilli S, Miscia S, Capitani S, Bertagnolo V. Vav1 and PU1 are recruited to the CD11b promoter in APL-derived promyelocytes: role of Vav1 in modulating PU1-containing complexes during ATRA-induced differentiation. Exp Cell Res 316: 38–47, 2010. [DOI] [PubMed] [Google Scholar]
- 14.Bryce PJ, Geha R, Oettgen HC. Desloratadine inhibits allergen-induced airway inflammation and bronchial hyperresponsiveness and alters T-cell responses in murine models of asthma. J Allergy Clin Immunol 112: 149–158, 2003. [DOI] [PubMed] [Google Scholar]
- 15.Burke KT, Colvin PL, Myatt L, Graf GA, Schroeder F, Woollett LA. Transport of maternal cholesterol to the fetus is affected by maternal plasma cholesterol concentrations in the golden Syrian hamster. J Lipid Res 50: 1146–1155, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cook-Mills JM. Eosinophil-endothelial cell interactions. Eosinophils in health and disease. In: Eosinophils in Health and Disease, edited by Lee JJ. New York: Elsevier, 2012, p. 139–153. [Google Scholar]
- 17.Cook-Mills JM. Isoforms of vitamin E differentially regulate PKC alpha and inflammation: a review. J Clin Cell Immunol 4: 1000137, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cook-Mills JM, Abdala-Valencia H, Hartert T. Two faces of vitamin E in the lung. Am J Respir Crit Care Med 188: 279–284. 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cook-Mills JM, Avila PC. Vitamin E and D regulation of allergic asthma immunopathogenesis. Int Immunopharmacol 29: 007, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cook-Mills JM, Marchese M, Abdala-Valencia H. Vascular cell adhesion molecule-1 expression and signaling during disease: regulation by reactive oxygen species and antioxidants. Antioxid Redox Signal 15: 1607–1638, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Cook-Mills JM, Marchese ME, Abdala-Valencia H. Vascular cell adhesion molecule-1 expression and signaling during disease: regulation by reactive oxygen species and antioxidants. Antioxid Redox Signal 15: 1607–1638, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Cook-Mills JM, McCary CA. Isoforms of vitamin E differentially regulate inflammation. Endocr Metab Immune Disord Drug Targets 10: 348–366, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Desrumaux C, Deckert V, Athias A, Masson D, Lizard G, Palleau V, Gambert P, Lagrost L. Plasma phospholipid transfer protein prevents vascular endothelium dysfunction by delivering alpha-tocopherol to endothelial cells. FASEB J 13: 883–892, 1999. [DOI] [PubMed] [Google Scholar]
- 24.Dow L, Tracey M, Villar A, Coggon D, Margetts BM, Campbell MJ, Holgate ST. Does dietary intake of vitamins C and E influence lung function in older people? Am J Respir Crit Care Med 154: 1401–1404, 1996. [DOI] [PubMed] [Google Scholar]
- 25.Fakhrzadeh L, Laskin JD, Laskin DL. Ozone-induced production of nitric oxide and TNF-α and tissue injury are dependent on NF-κB p50. Am J Physiol Lung Cell Mol Physiol 287: L279–L285, 2004. [DOI] [PubMed] [Google Scholar]
- 26.Fedulov AV, Kobzik L. Allergy risk is mediated by dendritic cells with congenital epigenetic changes. Am J Respir Cell Mol Biol 44: 285–292, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Fedulov AV, Leme A, Yang Z, Dahl M, Lim R, Mariani TJ, Kobzik L. Pulmonary exposure to particles during pregnancy causes increased neonatal asthma susceptibility. Am J Respir Cell Mol Biol 38: 57–67, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Fedulov AV, Leme AS, Kobzik L. Duration of allergic susceptibility in maternal transmission of asthma risk. Am J Reprod Immunol 58: 120–128, 2007. [DOI] [PubMed] [Google Scholar]
- 29.Friebele E. The attack of asthma. Environ Health Perspect 104: 22–25, 1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Hamada K, Suzaki Y, Goldman A, Ning YY, Goldsmith C, Palecanda A, Coull B, Hubeau C, Kobzik L. Allergen-independent maternal transmission of asthma susceptibility. J Immunol 170: 1683–1689, 2003. [DOI] [PubMed] [Google Scholar]
- 31.Hamahata A, Enkhbaatar P, Kraft ER, Lange M, Leonard SW, Traber MG, Cox RA, Schmalstieg FC, Hawkins HK, Whorton EB, Horvath EM, Szabo C, Traber LD, Herndon DN, Traber DL. gamma-Tocopherol nebulization by a lipid aerosolization device improves pulmonary function in sheep with burn and smoke inhalation injury. Free Radic Biol Med 45: 425–433, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hardy CL, Rolland JM, O'Hehir RE. The immunoregulatory and fibrotic roles of activin A in allergic asthma. Clin Exp Allergy 45: 1510–1522, 2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hedger MP, Winnall WR, Phillips DJ, de Kretser DM. The regulation and functions of activin and follistatin in inflammation and immunity. Vitam Horm 85: 255–97, 2011. [DOI] [PubMed] [Google Scholar]
- 34.Hernandez ML, Wagner JG, Aline Kala R, Mills K, Wells HB, Alexis NE, Lay JC, Jiang Q, Zhang H, Zhou H, Peden DB. Vitamin E gamma-tocopherol reduces airway neutrophil recruitment after inhaled endotoxin challenge in rats and in healthy volunteers. Free Radic Biol Med 8: 00054–00053, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Herz U, Joachim R, Ahrens B, Scheffold A, Radbruch A, Renz H. Allergic sensitization and allergen exposure during pregnancy favor the development of atopy in the neonate. Int Arch Allergy Immunol 124: 193–196, 2001. [DOI] [PubMed] [Google Scholar]
- 36.Herz U, Joachim R, Ahrens B, Scheffold A, Radbruch A, Renz H. Prenatal sensitization in a mouse model. Am J Respir Crit Care Med 162: S62–65, 2000. [DOI] [PubMed] [Google Scholar]
- 37.Hickstein DD, Baker DM, Gollahon KA, Back AL. Identification of the promoter of the myelomonocytic leukocyte integrin CD11b. Proc Natl Acad Sci USA 89: 2105–2109, 1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hubeau C, Apostolou I, Kobzik L. Adoptively transferred allergen-specific T cells cause maternal transmission of asthma risk. Am J Pathol 168: 1931–1939, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hubeau C, Apostolou I, Kobzik L. Targeting of CD25 and glucocorticoid-induced TNF receptor family-related gene-expressing T cells differentially modulates asthma risk in offspring of asthmatic and normal mother mice. J Immunol 178: 1477–1487, 2007. [DOI] [PubMed] [Google Scholar]
- 40.Hunter SC, Cahoon EB. Enhancing vitamin E in oilseeds: unraveling tocopherol and tocotrienol biosynthesis. Lipids 42: 97–108, 2007. [DOI] [PubMed] [Google Scholar]
- 41.Izzotti A, Balansky RM, Cartiglia C, Camoirano A, Longobardi M, De Flora S. Genomic and transcriptional alterations in mouse fetus liver after transplacental exposure to cigarette smoke. FASEB J 17: 1127–1129, 2003. [DOI] [PubMed] [Google Scholar]
- 42.Jarrett E, Hall E. Selective suppression of IgE antibody responsiveness by maternal influence. Nature 280: 145–147, 1979. [DOI] [PubMed] [Google Scholar]
- 43.Jiang Q, Christen S, Shigenaga MK, Ames BN. gamma-Tocopherol, the major form of vitamin E in the US diet, deserves more attention. Am J Clin Nutr 74: 714–722, 2001. [DOI] [PubMed] [Google Scholar]
- 44.Jishage K, Tachibe T, Ito T, Shibata N, Suzuki S, Mori T, Hani T, Arai H, Suzuki H. Vitamin E is essential for mouse placentation but not for embryonic development itself. Biol Reprod 73: 983–987, 2005. [DOI] [PubMed] [Google Scholar]
- 45.Kalayci O, Besler T, Kilinc K, Sekerel BE, Saraclar Y. Serum levels of antioxidant vitamins (alpha tocopherol, beta carotene, and ascorbic acid) in children with bronchial asthma. Turk J Peds 42: 17–21, 2000. [PubMed] [Google Scholar]
- 46.Kelly FJ, Mudway I, Blomberg A, Frew A, Sandstrom T. Altered lung antioxidant status in patients with mild asthma. Lancet 354: 482–483, 1999. [DOI] [PubMed] [Google Scholar]
- 47.Klebanoff CA, Spencer SP, Torabi-Parizi P, Grainger JR, Roychoudhuri R, Ji Y, Sukumar M, Muranski P, Scott CD, Hall JA, Ferreyra GA, Leonardi AJ, Borman ZA, Wang J, Palmer DC, Wilhelm C, Cai R, Sun J, Napoli JL, Danner RL, Gattinoni L, Belkaid Y, Restifo NP. Retinoic acid controls the homeostasis of pre-cDC-derived splenic and intestinal dendritic cells. J Exp Med 210: 1961–1976, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Lass A, Forster MJ, Sohal RS. Effects of coenzyme Q10 and alpha-tocopherol administration on their tissue levels in the mouse: elevation of mitochondrial alpha-tocopherol by coenzyme Q10. Free Radic Biol Med 26: 1375–1382, 1999. [DOI] [PubMed] [Google Scholar]
- 49.Leonard SW, Terasawa Y, Farese RV Jr, Traber MG. Incorporation of deuterated RRR- or all-rac-alpha-tocopherol in plasma and tissues of alpha-tocopherol transfer protein–null mice. Am J Clin Nutr 75: 555–560, 2002. [DOI] [PubMed] [Google Scholar]
- 50.Lim RH, Kobzik L. Maternal transmission of asthma risk. Am J Reprod Immunol 61: 1–10, 2009. [DOI] [PubMed] [Google Scholar]
- 51.Lim RH, Kobzik L, Dahl M. Risk for asthma in offspring of asthmatic mothers versus fathers: a meta-analysis. PLoS One 5: e10134, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mabalirajan U, Aich J, Leishangthem GD, Sharma SK, Dinda AK, Ghosh B. Effects of vitamin E on mitochondrial dysfunction and asthma features in an experimental allergic murine model. J Appl Physiol 107: 1285–1292, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Marchese ME, Kumar R, Colangelo LA, Avila PC, Jacobs DR Jr, Gross M, Sood A, Liu K, Cook-Mills JM. The vitamin E isoforms alpha-tocopherol and gamma-tocopherol have opposite associations with spirometric parameters: the CARDIA study. Respir Res 15: 31, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Martinez FD. Genes, environments, development and asthma: a reappraisal. Eur Respir J 29: 179–184, 2007. [DOI] [PubMed] [Google Scholar]
- 55.McCary CA, Abdala-Valencia H, Berdnikovs S, Cook-Mills JM. Supplemental and highly elevated tocopherol doses differentially regulate allergic inflammation: reversibility of alpha-tocopherol and gamma-tocopherol's effects. J Immunol 186: 3674–3685, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.McCary CA, Yoon Y, Panagabko C, Cho W, Atkinson J, Cook-Mills JM. Vitamin E isoforms directly bind PKCalpha and differentially regulate activation of PKCalpha. Biochem J 441: 189–198, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Meydani SN, Shapiro AC, Meydani M, Macauley JB, Blumberg JB. Effect of age and dietary fat (fish, corn and coconut oils) on tocopherol status of C57BL/6Nia mice. Lipids 22: 345–350, 1987. [DOI] [PubMed] [Google Scholar]
- 58.Meydani SN, Yogeeswaran G, Liu S, Baskar S, Meydani M. Fish oil and tocopherol-induced changes in natural killer cell-mediated cytotoxicity and PGE2 synthesis in young and old mice. J Nutr 118: 1245–1252, 1988. [DOI] [PubMed] [Google Scholar]
- 59.Muller-Schmehl K, Beninde J, Finckh B, Florian S, Dudenhausen JW, Brigelius-Flohe R, Schuelke M. Localization of alpha-tocopherol transfer protein in trophoblast, fetal capillaries' endothelium and amnion epithelium of human term placenta. Free Radic Res 38: 413–420, 2004. [DOI] [PubMed] [Google Scholar]
- 60.Nelson SE, Rogers RR, Frantz JA, Ziegler EE. Palm olein in infant formula: absorption of fat and minerals by normal infants. Am J Clin Nutr 64: 291–296, 1996. [DOI] [PubMed] [Google Scholar]
- 61.Odaka Y, Nakano M, Tanaka T, Kaburagi T, Yoshino H, Sato-Mito N, Sato K. The influence of a high-fat dietary environment in the fetal period on postnatal metabolic and immune function. Obesity (Silver Spring) 18: 1688–1694, 2010. [DOI] [PubMed] [Google Scholar]
- 62.Okamoto N, Murata T, Tamai H, Tanaka H, Nagai H. Effects of alpha tocopherol and probucol supplements on allergen-induced airway inflammation and hyperresponsiveness in a mouse model of allergic asthma. Int Arch Allergy Immunol 141: 172–180, 2006. [DOI] [PubMed] [Google Scholar]
- 63.Patel A, Liebner F, Netscher T, Mereiter K, Rosenau T. Vitamin E chemistry. Nitration of non-alpha-tocopherols: products and mechanistic considerations. J Org Chem 72: 6504–6512, 2007. [DOI] [PubMed] [Google Scholar]
- 64.Plantinga M, Hammad H, Lambrecht BN. Origin and functional specializations of DC subsets in the lung. Eur J Immunol 40: 2112–2118, 2010. [DOI] [PubMed] [Google Scholar]
- 65.Podda M, Weber C, Traber MG, Packer L. Simultaneous determination of tissue tocopherols, tocotrienols, ubiquinols, and ubiquinones. J Lipid Res 37: 893–901, 1996. [PubMed] [Google Scholar]
- 66.Rebholz SL, Jones T, Burke KT, Jaeschke A, Tso P, D'Alessio DA, Woollett LA. Multiparity leads to obesity and inflammation in mothers and obesity in male offspring. Am J Physiol Endocrinol Metab 302: E449–E457, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Reboul E, Klein A, Bietrix F, Gleize B, Malezet-Desmoulins C, Schneider M, Margotat A, Lagrost L, Collet X, Borel P. Scavenger receptor class B type I (SR-BI) is involved in vitamin E transport across the enterocyte. J Biol Chem 281: 4739–4745, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Schunemann HJ, Grant BJ, Freudenheim JL, Muti P, Browne RW, Drake JA, Klocke RA, Trevisan M. The relation of serum levels of antioxidant vitamins C and E, retinol and carotenoids with pulmonary function in the general population. Am J Respir Crit Care Med 163: 1246–1255, 2001. [DOI] [PubMed] [Google Scholar]
- 69.Shvedova AA, Kisin ER, Kagan VE, Karol MH. Increased lipid peroxidation and decreased antioxidants in lungs of guinea pigs following an allergic pulmonary response. Toxicol Appl Pharmacol 132: 72–81, 1995. [DOI] [PubMed] [Google Scholar]
- 70.Smit HA, Grievink L, Tabak C. Dietary influences on chronic obstructive lung disease and asthma: a review of the epidemiological evidence. Proc Nutr Soc 58: 309–319, 1999. [DOI] [PubMed] [Google Scholar]
- 71.Stock AT, Bedoui S. Vitamin A notches up CD11b hi DC development. Eur J Immunol 43: 1441–1444, 2013. [DOI] [PubMed] [Google Scholar]
- 72.Suchankova J, Voprsalova M, Kottova M, Semecky V, Visnovsky P. Effects of oral alpha-tocopherol on lung response in rat model of allergic asthma. Respirology 11: 414–421, 2006. [DOI] [PubMed] [Google Scholar]
- 73.Tabak C, Smit HA, Rasanen L, Fidanza F, Menotti A, Nissinen A, Feskens EJ, Heederik D, Kromhout D. Dietary factors and pulmonary function: a cross sectional study in middle aged men from three European countries. Thorax 54: 1021–1026, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Tang H, Chen F, Tan Q, Tan S, Liu L, Zhang F. Regulation of CD11b transcription by decreasing PRC2 and increased acH4 level during ATRA-induced HL-60 differentiation. Acta Biochim Biophys Sin 41: 588–593, 2009. [DOI] [PubMed] [Google Scholar]
- 75.Traber MG, Kayden HJ. Preferential incorporation of alpha-tocopherol vs gamma-tocopherol in human lipoproteins. Am J Clin Nutr 49: 517–526, 1989. [DOI] [PubMed] [Google Scholar]
- 76.Traber MG, Siddens LK, Leonard SW, Schock B, Gohil K, Krueger SK, Cross CE, Williams DE. Alpha-tocopherol modulates Cyp3a expression, increases gamma-CEHC production, and limits tissue gamma-tocopherol accumulation in mice fed high gamma-tocopherol diets. Free Radic Biol Med 38: 773–785, 2005. [DOI] [PubMed] [Google Scholar]
- 77.Troisi RJ, Willett WC, Weiss ST, Trichopoulos D, Rosner B, Speizer FE. A prospective study of diet and adult-onset asthma. Am J Respir Crit Care Med 151: 1401–1408, 1995. [DOI] [PubMed] [Google Scholar]
- 78.Uauy R, Hoffman DR, Birch EE, Birch DG, Jameson DM, Tyson J. Safety and efficacy of omega-3 fatty acids in the nutrition of very low birth weight infants: soy oil and marine oil supplementation of formula. J Pediatr 124: 612–620, 1994. [DOI] [PubMed] [Google Scholar]
- 79.Umegaki K, Uramoto H, Esashi T. Lack of influence of a long-term high or low vitamin E diet on the oxidative DNA damage in the bone marrow of mice. Intl J Vitamin Nutr Res 67: 149–154, 1997. [PubMed] [Google Scholar]
- 80.van Rijt LS, Lambrecht BN. Dendritic cells in asthma: a function beyond sensitization. Clin Exp Allergy 35: 1125–1134, 2005. [DOI] [PubMed] [Google Scholar]
- 81.van Schayck CP, Smit HA. The prevalence of asthma in children: a reversing trend. Eur Respir J 26: 647–650, 2005. [DOI] [PubMed] [Google Scholar]
- 82.Vander Lugt B, Khan AA, Hackney JA, Agrawal S, Lesch J, Zhou M, Lee WP, Park S, Xu M, DeVoss J, Spooner CJ, Chalouni C, Delamarre L, Mellman I, Singh H. Transcriptional programming of dendritic cells for enhanced MHC class II antigen presentation. Nat Immunol 15: 161–167, 2014. [DOI] [PubMed] [Google Scholar]
- 83.Vanhees K, Coort S, Ruijters EJ, Godschalk RW, van Schooten FJ, Barjesteh van Waalwijk van Doorn-Khosrovani S. Epigenetics: prenatal exposure to genistein leaves a permanent signature on the hematopoietic lineage. FASEB J 25: 797–807, 2011. [DOI] [PubMed] [Google Scholar]
- 84.Vollmer WM, Osborne ML, Buist AS. 20-year trends in the prevalence of asthma and chronic airflow obstruction in an HMO. Am J Respir Crit Care Med 157: 1079–1084, 1998. [DOI] [PubMed] [Google Scholar]
- 85.Wagner JG, Harkema JR, Jiang Q, Illek B, Ames BN, Peden DB. Gamma-tocopherol attenuates ozone-induced exacerbation of allergic rhinosinusitis in rats. Toxicol Pathol 37: 481–491, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Wagner JG, Jiang Q, Harkema JR, Ames BN, Illek B, Roubey RA, Peden DB. Gamma-tocopherol prevents airway eosinophilia and mucous cell hyperplasia in experimentally induced allergic rhinitis and asthma. Clin Exp Allergy 38: 501–511, 2008. [DOI] [PubMed] [Google Scholar]
- 87.Wagner KH, Kamal-Eldin A, Elmadfa I. Gamma-tocopherol–an underestimated vitamin? Ann Nutr Metab 48: 169–188, 2004. [DOI] [PubMed] [Google Scholar]
- 88.Wang X, Pesakhov S, Harrison JS, Danilenko M, Studzinski GP. ERK5 pathway regulates transcription factors important for monocytic differentiation of human myeloid leukemia cells. J Cell Physiol 229: 856–867, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Weiss ST. Diet as a risk factor for asthma. Ciba Fndn Symp 206: 244–257, 1997. [PubMed] [Google Scholar]
- 90.Williams JW, Tjota MY, Clay BS, Vander Lugt B, Bandukwala HS, Hrusch CL, Decker DC, Blaine KM, Fixsen BR, Singh H, Sciammas R, Sperling AI. Transcription factor IRF4 drives dendritic cells to promote Th2 differentiation. Nat Commun 4: 2990, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Wiser J, Alexis NE, Jiang Q, Wu W, Robinette C, Roubey R, Peden DB. In vivo gamma-tocopherol supplementation decreases systemic oxidative stress and cytokine responses of human monocytes in normal and asthmatic subjects. Free Radic Biol Med 45: 40–49, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Wolf G. How an increased intake of alpha-tocopherol can suppress the bioavailability of gamma-tocopherol. Nutr Rev 64: 295–299, 2006. [DOI] [PubMed] [Google Scholar]
- 93.Woollett LA. Maternal cholesterol in fetal development: transport of cholesterol from the maternal to the fetal circulation. Am J Clin Nutr 82: 1155–1161, 2005. [DOI] [PubMed] [Google Scholar]
- 94.Yokota A, Takeuchi H, Maeda N, Ohoka Y, Kato C, Song SY, Iwata M. GM-CSF and IL-4 synergistically trigger dendritic cells to acquire retinoic acid-producing capacity. Int Immunol 21: 361–377, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]