

Abstract
Background. The malaria-specific T-cell response is believed to be important for protective immunity. Antimalarial chemoprevention may affect this response by altering exposure to malaria antigens.
Methods. We performed interferon γ (IFNγ) ELISpot assays to assess the cellular immune response to blood-stage and pre-erythrocytic antigens longitudinally from 1 to 3 years of age in 196 children enrolled in a randomized trial of antimalarial chemoprevention in Tororo, Uganda, an area of high transmission intensity.
Results. IFNγ responses to blood-stage antigens, particularly MSP1, were frequently detected, strongly associated with recent malaria exposure, and lower in those adherent to chemoprevention compared to nonadherent children and those randomized to no chemoprevention. IFNγ responses to pre-erythrocytic antigens were infrequent and similar between children randomized to chemoprevention or no chemoprevention. Responses to blood-stage antigens were not associated with subsequent protection from malaria (aHR 0.96, P = .83), but responses to pre-erythrocytic antigens were associated with protection after adjusting for prior malaria exposure (aHR 0.52, P = .009).
Conclusions. In this high transmission setting, IFNγ responses to blood-stage antigens were common and associated with recent exposure to malaria but not protection from subsequent malaria. Responses to pre-erythrocytic antigens were uncommon, not associated with exposure but were associated with protection from subsequent malaria.
Keywords: malaria, falciparum, IFNγ, T cell, immunity, antimalarial chemoprevention
Plasmodium falciparum malaria continues to be a leading cause of morbidity and mortality in Sub-Saharan Africa [1, 2]. Although children living in endemic settings eventually develop clinical immunity to malaria, most individuals experience numerous infections before clinical protection is achieved. A vaccine to prevent malaria is urgently needed, but progress has been limited by our lack of understanding of both the critical P. falciparum antigenic targets and the immune effector mechanisms needed to confer protective immunity.
Growing evidence suggests that malaria-specific T cells induced by natural infection or by vaccination may protect against clinical disease [3–8]. T-cell responses to blood-stage antigens, including merozoite surface antigen 1 (MSP1), are frequently observed among children living in endemic settings, and a few studies have found them to be associated with protection from future malaria [4, 8]. However, thus far blood-stage vaccines have not proven efficacious [9]. Several lines of evidence have prompted a growing interest in pre-erythrocytic stage malaria antigens as potential vaccine targets. T-cell responses to the pre-erythrocytic circumsporozoite (CSP) antigen have been shown to correlate with protection from future parasitemia [3, 6], and a subunit vaccine (RTS,S) incorporating CSP has modestly reduced clinical P. falciparum malaria in African infants in phase 2 and 3 trials [10–12]. T-cell responses to other pre-erythrocytic proteins including TRAP and LSA-1 have also been associated with protection in humans [5, 7, 13, 14]. Moreover, it has long been known that vaccination with irradiated sporozoites, which arrest development during the liver stage, confers sterile protective immunity in humans [15–18], suggesting an important role for the T-cell response to pre-erythrocytic antigens in mediating vaccine-induced immune protection.
The use of chemoprevention, either seasonal or year-round, has recently been explored as a public health strategy to prevent mortality and morbidity due to childhood malaria in endemic settings [19, 20]. Although it has been shown to be effective in reducing malaria, concerns have been raised that a “rebound” increase in malaria incidence may be observed once chemoprevention is stopped, due to delayed development of protective immune responses [21, 22]. However, recent studies suggest that provision of antimalarial drugs that target blood-stage malaria may actually enhance the development of cellular immune responses directed at pre-erythrocytic antigens and, somewhat paradoxically, foster the development of protective immunity, a strategy termed “infection-treatment vaccination” [23–27]. In these studies, individuals experimentally infected by sporozoites while receiving chloroquine, which prevents blood-stage malaria but allows the clinically silent liver stage infection to develop, consistently exhibited sterile protection upon rechallenge [25–27]. These data suggest that limiting exposure to blood-stage infection may actually enhance the development of immune responses to pre-erythrocytic stages, perhaps due to enhanced exposure to liver stage antigens [28] or avoidance of immunoregulatory mechanisms induced by parasitemia [29]. By analogy, provision of chemoprevention to heavily exposed children might actually encourage pre-erythrocytic responses and foster the development of protective immunity.
In this study, we performed a longitudinal evaluation of malaria-specific T-cell responses generated in response to natural infection and compared the responses of children receiving monthly chemoprevention with dihydroartemisinin-piperaquine (DP) to those receiving no chemoprevention as part of a randomized clinical trial. We hypothesized that interferon γ (IFNγ) responses to pre-erythrocytic antigens would be associated with protection from malaria, and that selective suppression of blood-stage malaria by chemoprevention given to children living in a high endemicity setting may limit the development of T-cell responses to blood-stage antigens and enhance the development of responses to pre-erythrocytic antigens.
METHODS
Study Participants and Design
Samples were obtained from children enrolled in a randomized, controlled, open-label trial comparing the efficacy and safety of 3 regimens vs no therapy for the prevention of malaria in Tororo, a district in eastern Uganda with intense year-round malaria transmission and an entomological inoculation rate estimated at 125 [30]. Details of this trial have been described elsewhere [31], and written informed consent was obtained from the parent or guardian of all study participants. Briefly, 400 infants were enrolled and 393 randomized at 6 months of age to no chemoprevention, monthly sulfadoxine-pyrimethamine, daily trimethoprim-sulfamethoxazole, or monthly dihydroartemisinin-piperaquine (DP). The substudy described in this report includes only samples from infants randomized to DP (n = 98) and no chemoprevention (n = 98). Study drugs were administered at home without supervision. Chemoprevention was given from 6 months through 24 months of age, and study participants were followed for 1 additional year until they reached 36 months of age. Monthly assessments were performed to ensure compliance with study protocols and perform routine blood smears. Children who presented with a fever (tympanic temperature ≥38.0°C) or history of fever in the previous 24 hours had blood obtained by finger prick for a thick smear. If the thick smear was positive for malaria parasites, the patient was diagnosed with malaria regardless of parasite density and given artemether-lumefantrine. Incident episodes of malaria were defined as all febrile episodes accompanied by any parasitemia requiring treatment but not preceded by another treatment in the prior 14 days [2]. The incidence of malaria was calculated as the number of episodes per person years (ppy) at risk.
Sample Collection and Processing
Approximately 6–10 mL blood was collected from each subject at routine visits 3 times during the chemoprevention intervention (12–16, 20, and 24 months of age) and 2 times after the intervention (28 and 32–36 months of age). Peripheral blood mononuclear cells (PBMC) were isolated by density gradient centrifugation (Ficoll-Histopaque; GE Life Sciences). PBMC were used fresh for ELISpot assays, and remaining PBMC were cryopreserved for use in flow cytometric studies.
Ex-Vivo IFNγ ELISpots
Overlapping peptides (18mers overlapping by 11 amino acids) spanning the 3D7 sequence of 7 pre-erythrocytic (CSP [6], TRAP [7], LSA1[5], SIAP1 [32], SIAP2 [32], CelTOS [32], P52 [33]) and 3 blood-stage P. falciparum proteins (AMA1[34], MSP1 [8], HGXPRT[35]) were synthesized (Mimotopes, Australia) and pooled into 16 protein-specific pools (25–54 peptides per pool, Supplementary Table 1). Fresh PBMC were plated at 2 × 105 cells/well in 96-well plates precoated with 2 μg/mL anti-IFNγ monoclonal antibody. Cells were stimulated with malaria peptide pools (10 µg/mL/peptide), phytohemagluttinin (1 µg/mL), or media alone. Plates were incubated overnight at 37°C and developed using standard methods [36]. Individual IFNγ-secreting cells were visualized as purple spots and counted by a plate reader. The magnitude of IFNγ ELISpot responses was calculated as the number of spot-forming cells per million input cells (SFC/M) after subtraction of the background response (mean SFC/10^6 cells of 3 negative control wells). For proteins with >1 peptide pools, responses to the overall protein were summed. ELISpot assays with mean background responses >30 SFC/M were not considered in the analysis. A response was considered positive if >3× the average negative control wells and >20 SFCs/10^6 PBMC [36]. Because response frequencies for most individual antigens were low, for the multivariate analyses we grouped responses into stage-specific categories (ie, responses to any pre-erythrocytic or any blood-stage antigen).
Intracellular Cytokine Staining
ELISpot responses with magnitudes >250 SFC/M were further characterized by flow cytometry. Thawed PBMC were rested overnight, then stimulated with media, peptide pools (5 µg/mL/peptide), or phorbol miristate acetate/calcium ionophore at 1 × 106 cells/condition. Anti-CD28 and –CD49d were added for costimulation (0.5 µg/mL, BD Pharmingen). APC-conjugated CD107a (Biolegend), Brefeldin-A (10 µg/mL) and Monensin (10 µg/mL, BD Pharmingen) were added at the time of incubation. After incubation, cells were washed, fixed, and permeabilized [37], and stained using the following antibodies: Brilliant violet (BV) 650-conjugated CD3, PerCP–conjugated anti-CD4, BV 511-conjugated CD14 and CD19, BV 605-conjugated CD45RA, FITC-conjugated CCR7, APC-H7-conjugated CD8, PE-Cy7-conjugated IFNγ (Biolegend) PE-conjugated anti-IL-10 (BD Pharmingen), and live/dead aqua amine (Invitrogen).
Piperaquine Levels
Piperaquine (PQ) drug levels were measured at 4 random time points during the intervention in subjects randomized to DP, using dried blood spot samples as previously described [31]. The PQ assay demonstrated a lower limit of quantitation of 10 ng/mL with a calibration range of 10–100 ng/mL. Subjects were characterized as “adherent” if ≥3 of 4 PQ measurements were ≥10 ng/mL, and “nonadherent” if <3 PQ measurements were ≥10 ng/mL.
Statistical Analysis
Data analysis was performed using Stata version 12 (Stata Corp, College Station, Texas) and PRISM version 6 (Graph Pad). Associations between current/prior malaria infection (malaria or asymptomatic parasitemia at the time of the assay, or duration since last episode of malaria if the bloodsmear at the time of the assay was negative), age, and the presence or absence of ELISpot responses were measured using generalized estimating equations with robust standard errors to account for repeated measures in the same child. Associations between DP chemoprevention during the intervention and ELISpot responses at each timepoint were assessed using a χ2 test of proportions.
The cumulative risk of developing malaria following ELISpot assays was estimated using the Kaplan–Meier product limit formula. For the primary analysis, associations between 24 month ELISpot responses were made using Cox proportional hazards models adjusting for prior or current malaria exposure and chemoprevention randomization assignment. As a secondary analysis, associations between ELISpot responses at all timepoints and time to next episode of malaria were made using a within-subject variance-corrected cox-proportional hazards model, adjusting for prior/current malaria exposure, age, chemoprevention randomization, and their interaction. Adjusted predicted survival curves were generated using predictions from the final Cox model. Negative binomial regression was used to estimate associations between ELISpot responses at 24 months of age and the incidence of malaria in the following year (incidence rate ratios [IRR]), adjusting for prior malaria and chemoprevention assignment as above. In all analyses, a 2-tailed P value < .05 was considered to be statistically significant.
RESULTS
Clinical Characteristics of Study Cohort
The study cohort consisted of 196 infants followed from 6 months through 3 years of age. Infants were randomized to receive monthly DP (n = 98) or no chemoprevention (n = 98) from 6 months through 24 months of age, then followed for an additional year after stopping chemoprevention. Self-reported insecticide-treated bednet usage was 98.5% throughout the study. During the intervention, the incidence of malaria in the no chemoprevention arm was 6.95 episodes ppy; monthly DP had a protective efficacy of 58% (95% confidence interval [CI], 45%–67%, P < .001) which waned over the period of the intervention [31]. Notably, there was significant nonadherence to DP; only 31% of subjects had ≥3 of 4 measured piperaquine levels ≥10 ng/mL. In adherent children, monthly DP had a protective efficacy of 87% (95% CI, 8%–22%, P < .001). In the year after the intervention was stopped, the incidence of malaria in the no chemoprevention arm was 10.85 episodes ppy, with no significant difference in the group that had received DP chemoprevention that were adherent (9.26 episodes ppy) or nonadherent (11.53 episodes ppy). A total of 790 ELISpots were performed from these 196 children (mean 4 ELISpots/child, Supplementary Figure 1). Of these, 162 ELISpot assays (21%) were performed on samples obtained while the subject had parasitemia (positive blood smear), of which 80 were performed on samples obtained while the subject had symptomatic malaria (parasitemia with a fever requiring treatment).
Frequent Recognition of MSP1 but not of Pre-erythrocytic Antigens
To define the malaria antigens targeted by T cells in these children, we analyzed the production of IFNγ after stimulation with peptides spanning blood-stage and pre-erythrocytic antigens longitudinally using ELISpot assays. In general, IFNγ responses were of low magnitude, with a majority of responses <200 SFCs/M (Figure 1A). MSP1, a blood-stage antigen, was most frequently recognized, with 36% of subjects recognizing this antigen at 24 months of age (Figure 1B). In contrast, responses to pre-erythrocytic antigens were infrequent, with recognition frequencies <10% to most, including the leading vaccine candidate antigen CSP (Figure 1B). Of responders to blood-stage antigens (n = 231), 92% only had a response to a single antigen, and 8% had a response to 2 antigens. Of responders to pre-erythrocytic antigens (n = 86), 85% had a response to a single antigen, 11% had a response to 2 antigens, and 3% had a response to ≥3 antigens.
Figure 1.
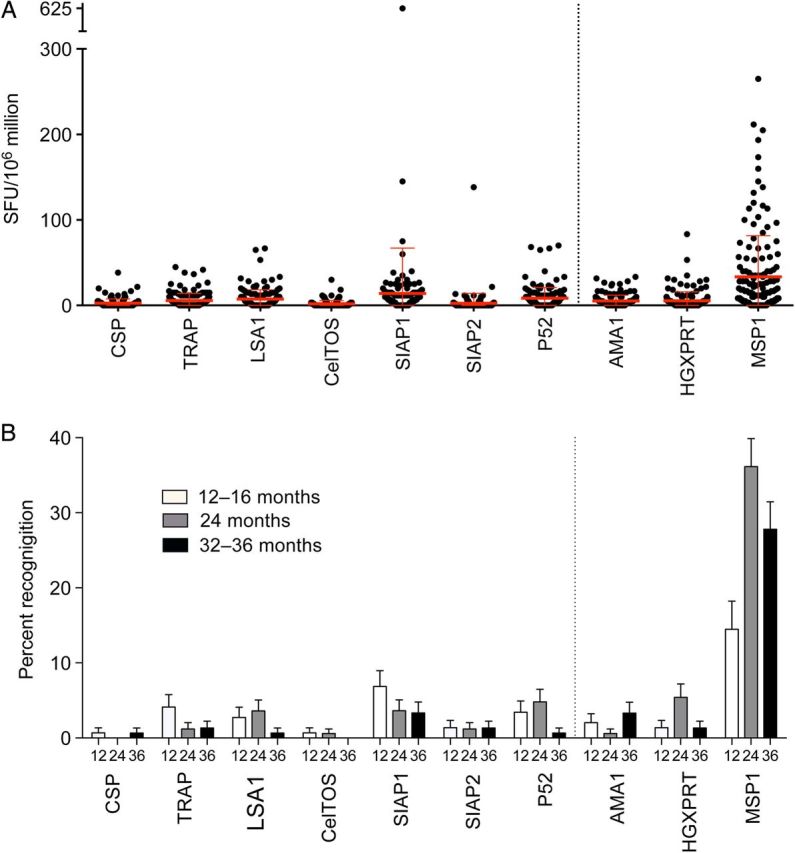
IFNγ ELISpot responses to pre-erythrocytic and blood-stage antigens. A, Magnitude of IFNγ ELISpot responses to 7 pre-erythrocytic (antigens on left of dotted line) and 3 blood-stage malaria proteins (antigens on right of dotted line) at 24 months of age, expressed as spot-forming cells (SFC) per million PBMC. B, Percent of subjects recognizing each protein at 12–16, 24, and 32–36 months of age. Abbreviations: IFNγ, interferon γ; PBMC, peripheral blood mononuclear cells.
Functional and phenotypic analysis of MSP1-responding cells in a representative child revealed both CD4+ and CD8+ IFNγ-producing cells, with a significant proportion of CD4+ T cells that co-produced IL-10 (Figure 2A and B). While most responses to pre-erythrocytic antigens were small in magnitude, a notable exception was one subject who had a response of >500 SFCs/M to SIAP1 at all measured timepoints. A more detailed functional and phenotypic analysis revealed that the majority of SIAP1-specific cells in this child were CD8+ T cells that co-produced IFNγ and TNFα and degranulated in responses to stimulation (Figure 2C).
Figure 2.
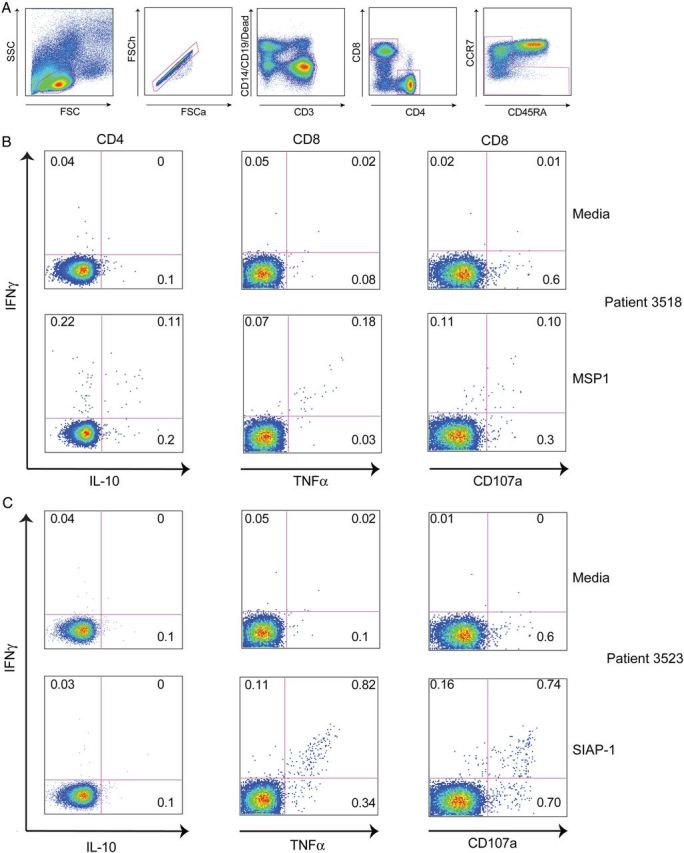
T-cell responses to malaria antigens characterized by multiparameter flow cytometry. A, Gating strategy to identify live, CD4+ and CD8+ memory T-cell populations. B, Intracellular cytokine assay demonstrating the T-cell response of one representative malaria-exposed child to MSP1 (bottom row), with media control shown in row above. C, Intracellular cytokine assay demonstrating the T-cell response of one malaria-exposed child to SIAP1 (bottom row), with media control shown in row above. Shown are CD4 (first column) and CD8 (right 2 columns) production of IFNγ (y-axis), IL-10 (x-axis, column 1), TNFa (x-axis, columns 2) and mobilization of the degranulation marker CD107a (y-axis, column 3). Abbreviations: IFNγ, interferon γ; IL, interleukin; TNF, tumor necrosis factor.
Recent Malaria, Parasitemia, and age Associated With Blood-stage but not Pre-erythrocytic Responses
The frequency of IFNγ responses to pre-erythrocytic and blood-stage antigens differed markedly based on a child's clinical malaria history. Recent malaria was strongly associated with a higher likelihood of response to blood-stage antigens (Figure 3; Table 1). However, individuals who were parasitemic at the time of the assay were significantly less likely to have a blood-stage response than children who were not parasitemic (RR 0.47, 95% CI, .32–.70, P < .001). These data are consistent with previous studies that have reported suppression of T-cell responses in the setting of current infection [38, 39]. In addition, blood-stage response frequencies increased gradually with age, with a slight decline at the final time point (Table 1). Similar findings were obtained when only analyzing responses to MSP1. In contrast, responses to pre-erythrocytic antigens were not associated with recent malaria, concurrent parasitemia, or age (Table 1). Stratification by chemoprevention randomization assignment did not significantly influence any of these results. Together, these findings suggest that the factors influencing the presence of IFNγ responses to blood-stage antigens differ from those for pre-erythrocytic malaria antigens, with recent infection correlating with higher blood-stage responses, and concurrent parasitemia suppressing blood-stage responses but not pre-erythrocytic responses.
Figure 3.

Recent malaria associated with IFNγ responses to blood-stage but not pre-erythrocytic antigens. A, Recognition frequencies to any blood-stage antigen in children stratified by prior or current malaria exposure. B, Recognition frequencies to any pre-erythrocytic antigen in children stratified by prior or current malaria exposure. Abbreviation: IFNγ, interferon γ.
Table 1.
Relationship of IFNγ Response Frequencies to Current and Prior Malaria Exposure
| Predictors of Malaria Exposure | Any Blood-stage Response |
Any Pre-erythrocytic |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Proportion Response (%) | Unadjusted RR (95% CI) |
P Value | Adjusted RR (95% CI) |
P Value | Proportion Response (%) | Unadjusted RR (95% CI) |
P Value | Adjusted RR (95% CI) |
P Value | |
| Prior or current malaria exposure | ||||||||||
| Malaria >240 da | 8/121 (6.6) | Ref | Ref | Ref | Ref | 14/121 (11.6) | Ref | Ref | Ref | Ref |
| Malaria 181–240 da | 2/18 (11.1) | 2.07 (.52–8.28) | .304 | 1.99 (.49–8.10) | .334 | 3/18 (16.7) | 1.53 (.49–4.79) | .463 | 1.57 (.49–4.98) | .447 |
| Malaria 121–180 da | 8/36 (22.2) | 3.55 (1.52–8.27) | .003 | 3.30 (1.42–7.68) | .006 | 6/36 (16.7) | 1.46 (.62–3.46) | .387 | 1.47 (.61–3.55) | .386 |
| Malaria 61–120 da | 27/78 (34.6) | 5.92 (2.76–12.7) | <.001 | 5.39 (2.51–11.57) | <.001 | 8/78 (10.3) | 0.89 (.43–1.86) | .764 | 0.95 (.45–2.01) | .888 |
| Malaria 1–60 da | 158/362 (43.7) | 7.28 (3.65–14.5) | <.001 | 6.60 (3.24–13.42) | <.001 | 35/362 (9.7) | 0.86 (.47–1.56) | .614 | 0.95 (.49–1.85) | .888 |
| Current AP | 17/82 (20.7) | 3.33 (1.46–7.57) | .004 | 3.04 (1.29–7.16) | .011 | 14/82 (17.1) | 1.50 (.74–3.03) | .26 | 1.79 (.82–3.91) | .147 |
| Current Malaria | 11/80 (13.8) | 2.20 (.89–5.42) | .088 | 2.07 (.84–5.13) | .116 | 6/80 (7.5) | 0.65 (.25–1.66) | .37 | 0.69 (.27–1.82) | .458 |
| Age at ELISpot in months | ||||||||||
| 12–16 | 25/146 (17.1) | Ref | Ref | Ref | Ref | 18/146 (12.3) | Ref | Ref | Ref | Ref |
| 20 | 38/155 (24.5) | 1.40 (.90–2.16) | .136 | 1.33 (.88–2.00) | .172 | 16/155 (10.3) | 0.85 (.45–1.58) | .602 | 0.87 (.46–1.64) | .664 |
| 24 | 61/166 (36.8) | 2.12 (1.45–3.11) | <.001 | 1.68 (1.17–2.41) | .005 | 22/166 (13.3) | 1.08 (.66–1.77) | .766 | 1.04 (.62–1.75) | .891 |
| 28 | 60/159 (37.7) | 2.19 (1.50–3.19) | <.001 | 1.62 (1.13–2.33) | .009 | 18/159 (11.3) | 0.92 (.49–1.74) | .797 | 0.92 (.48–1.76) | .794 |
| 32–36 | 47/151 (31.1) | 1.80 (1.17–2.76) | .007 | 1.25 (.81–1.92) | .318 | 12/151 (8.0) | 0.65 (.32–1.33) | .236 | 0.59 (.26–1.35) | .213 |
Abbreviations: AP, asymptomatic parasitemia; CI, confidence interval; RR, relative risk using generalized estimating equations adjusted for repeated measures in the same patient.
a Days since malaria if blood smear negative at time of the assay.
Impact of Chemoprevention on Malaria-specific ELISpot Responses
We next compared IFNγ ELISpot responses between children randomized to DP vs no chemoprevention to test the hypothesis that selective suppression of blood-stage malaria by chemoprevention may alter the balance of responses to blood-stage and pre-erythrocytic antigens. Given the high levels of nonadherence to DP observed in this study, children were stratified as being on no chemoprevention, DP nonadherent, and DP adherent (Figure 4). During the intervention, children who were DP nonadherent had similar blood-stage recognition frequencies to children who were randomized to no chemoprevention (Figure 4A). However, DP adherent children were consistently less likely to have IFNγ responses to blood-stage malaria antigens in comparison to children randomized to no chemoprevention and DP nonadherent children at 12–16 months (4% vs 18%, P = .076), 20 months (8% vs 19%, P = .11), and 24 months of age (3% vs 22%, P = .001). Post-intervention, children in all 3 groups had similar recognition of blood-stage antigens (Figure 4A). Similar results were obtained when only analyzing responses to MSP1, and when comparing responses between adherent children and either nonadherent or children assigned no chemoprevention (data not shown). In contrast, DP chemoprevention was not consistently or significantly associated with a difference in the frequency of IFNγ responses to pre-erythrocytic malaria antigens (Figure 4B). Together, these results suggest that taking DP chemoprevention does not significantly enhance ELISpot responses to this wide panel of pre-erythrocytic antigens, and is associated with lower T-cell recognition of blood-stage malaria antigens compared to children not taking chemoprevention.
Figure 4.

Impact of chemoprevention on ELISpot responses to blood-stage and pre-erythrocytic antigens. A, Recognition frequencies to any blood-stage antigen at 12–16, 20, 24, 28, and 32–36 months of age in children stratified by antimalarial chemoprevention group (no chemoprevention, DP: Non-adherent, and DP: Adherent. Solid lines represent ELISpot responses during the intervention; dotted lines represent ELISpot responses post-intervention. B, Recognition frequencies to any pre-erythrocytic antigens in children stratified by antimalarial chemoprevention group. Abbreviation: DP, dihydroartemisinin-piperaquine.
Pre-erythrocytic and Blood-stage Responses are Differentially Associated With Protection From Malaria
Finally, we sought to determine whether IFNγ responses to blood-stage and pre-erythrocytic antigens were associated with subsequent protection from malaria. Our primary analysis focused on the relationship of ELISpot responses at 24 months of age (when chemoprevention was stopped) to the time to the next episode of malaria, and to the incidence of malaria in the year following the intervention. Given the challenge of distinguishing protection from a lack of exposure to malaria-infected mosquitos [40], we adjusted all analyses for indices of prior or current malaria as surrogate measures of exposure intensity. On univariate analysis, IFNγ responses to blood-stage antigens at 24 months of age were associated with a 50% increased hazard of developing malaria and a nonsignificant increased risk of malaria in the year following the assay (Table 2). After controlling for prior or current malaria exposure, there was no significant association between blood-stage responses and subsequent protection. Results were similar when stratifying by chemoprevention randomization assignment (data not shown), and when analyzing responses to MSP1 alone (Table 2). In contrast, IFNγ responses to pre-erythrocytic antigens were associated with a 48% reduced hazard of developing malaria after adjustment for prior or current malaria exposure (P = .009, Table 2, Figure 5), and a nonsignificant decreased risk of malaria (IRR 0.83, P = .12) in the year following the assay. As a secondary analysis, we examined the relationship of ELISpot responses at all timepoints and time to next episode of malaria, adjusting for age, current/prior malaria exposure, and chemoprevention randomization assignment. This analysis yielded nearly identical results (blood-stage aHR 0.97, 95% CI, .77–1.2, P = .809; pre-erythrocytic aHR 0.71, 95% CI, .50–.99, P = .041). Together, these findings suggest that IFNγ responses to pre-erythrocytic antigens may be associated with protection from future malaria, but responses to blood-stage antigens are not.
Table 2.
Association of ELISPot Responses at 24 Months of age With Protection From Symptomatic Malaria in Following Year
| Predictor | Time Until Next Episode of Malaria |
Future Incidence of Malaria |
||||||
|---|---|---|---|---|---|---|---|---|
| Univariate |
Multivariatea |
Univariate |
Multivariatea |
|||||
| HR (95% CI) | P Value | HR (95% CI) | P Value | IRR (95% CI) | P Value | IRR (95% CI) | P Value | |
| Any Blood-stage response | 1.50 (1.09–2.07) | .014 | 0.96 (.67–1.37) | .827 | 1.16 (.97–1.39) | .096 | 1.00 (.84–1.18) | .987 |
| MSP1 Response | 1.54 (1.11–2.13) | .009 | 0.99 (.70–1.41) | .961 | 1.18 (.99–1.41) | .067 | 1.02 (.86–1.20) | .840 |
| Any Pre-erythrocytic Response | 0.65 (.41–1.03) | .067 | 0.52 (.32–.85) | .009 | 0.83 (.64–1.07) | .150 | 0.83 (.66–1.05) | .123 |
Abbreviations: CI, confidence interval; HR, hazard ratio; IRR, incidence rate ratio.
a Controlling for prior or current malaria exposure strata (Table 1) and chemoprevention randomization assignment.
Figure 5.

ELISpot responses and protection from symptomatic malaria. A, Survival curves for IFNγ ELISpot responses to blood-stage antigens at 24 months of age after adjusting for prior or current malaria exposure (with categories as defined in Table 1) and chemoprevention randomization assignment. The solid black line represents responders to any blood-stage antigen and the dashed line represents nonresponders. B, Survival curves for ELISpot responses to pre-erythrocytic antigens at 24 months of age after adjusting for prior or current malaria. Abbreviation: IFNγ, interferon γ.
DISCUSSION
In this cohort of young children living in a high endemicity setting in Uganda, we evaluated malaria-specific IFNγ ELISpot responses to a large panel of pre-erythrocytic and blood-stage antigens, with MSP1-specific responses most frequently detected. Although responses to blood-stage antigens—specifically MSP1—were associated with recent malaria exposure, they were not associated with protection. IFNγ responses to pre-erythrocytic antigens were less frequently observed, but in contrast to blood-stage responses, were associated with subsequent protection, as measured by a reduced hazard of developing malaria. Finally, although chemoprevention was effective at reducing the incidence of malaria, it did not significantly enhance responses to pre-erythrocytic antigens but was rather associated with a lower frequency of T-cell responses to blood-stage antigens in adherent children.
MSP1 was considered a leading vaccine candidate following studies showing protection in vaccinated old world monkeys [41, 42], but a phase 2 trial of the C-terminal 42-kDa region of the MSP1 3D7 allele showed no efficacy against malaria in vaccinated children [9]. More recently, MSP1-specific IFNγ ELISpot responses were reported to correlate with protection from malaria in a treatment reinfection study of naturally exposed Kenyan children 1–14 years of age [8]. In contrast to that study, we found that IFNγ responses to MSP1 were not associated with protection but instead associated with recent exposure to malaria. These diverging results may be explained by the young age of children enrolled in our cohort, as T-cell responses are known to be less efficacious in early childhood, and responses to MSP1 may be qualitatively different in young children in comparison to older children and adults [43]. Furthermore, children enrolled in our cohort were exposed to intense, year-round transmission, and this continual antigen exposure could induce immunoregulatory mechanisms that dampen the inflammatory response. We have recently shown in a separate cohort of children that CD4+ T cells co-producing interleukin 10 (IL-10) and IFNγ dominate the malaria-specific response in heavily exposed young children, and this population was also not associated with protection [29]. This IL-10 producing Th1 population may play an important role in preventing immunopathology but may interfere with the development of antimalarial immunity, as has been shown with other parasitic infections [44].
We observed that responses to pre-erythrocytic antigens were associated with protection from malaria, consistent with observations from prior studies [3, 6, 7] and with studies of the modestly protective RTS/S vaccine, which is based on the pre-erythrocytic antigen CSP [10–12]. Liver stage-specific CD8+ T cells have been demonstrated to be important in protective antimalarial immunity in animal models [45–48]. However, a notable finding of our study was the low recognition frequency of responses to pre-erythrocytic antigens, including the leading vaccine candidates CSP, TRAP, and LSA1. This is in agreement with prior studies reporting infrequent responses to pre-erythrocytic antigens in young children, including children vaccinated with RTS/S [49]. Homing of T cells to the liver and other tissues may partially explain this low recognition frequency and has been demonstrated in non-human primate studies [50], but sampling of tissues was not feasible in our study.
Finally, we observed that monthly chemoprevention with DP did not significantly enhance responses to pre-erythrocytic antigens, although it did appear to reduce responses to blood-stage antigens, particularly in adherent children. Children randomized to DP did not have significantly more or less malaria in the year following the intervention than children randomized to no chemoprevention, although the high levels of nonadherence to the study intervention make it difficult to draw definitive conclusions from either of these observations. Studies employing directly observed therapy would enable a better assessment of the impact of antimalarial chemoprevention on the development of malaria-specific T-cell responses.
In conclusion, among naturally exposed children living in a high endemicity setting, IFNγ ELISpot responses to blood-stage antigens were frequently observed in young children but were not associated with protection. Responses to pre-erythrocytic antigens were associated with prospective protection from malaria, but were not enhanced in children receiving antimalarial chemoprevention. A better understanding of correlates of protective immunity, as well as factors that impede the development of immunity to pre-erythrocytic antigens, will help inform the design of future strategies for antimalarial chemoprevention and vaccines.
Supplementary Data
Supplementary materials are available at The Journal of Infectious Diseases online (http://jid.oxfordjournals.org). Supplementary materials consist of data provided by the author that are published to benefit the reader. The posted materials are not copyedited. The contents of all supplementary data are the sole responsibility of the authors. Questions or messages regarding errors should be addressed to the author.
Notes
Acknowledgments. We are grateful to all the parents and guardians for kindly giving their consent and to the study participants for their cooperation. We thank all the members of the study team in Uganda for their tireless effort and excellent work.
Financial support. This study received financial support from the National Institutes of Health (HD059454, RO1AI093615, K23AI100949, K24AI113002, KL2TR00014) and the Burroughs Wellcome Fund/American Society of Tropical Medicine and Hygiene.
Potential conflicts of interest. All authors: No potential conflicts of interest.
All authors have submitted the ICMJE Form for Disclosure of Potential Conflicts of Interest. Conflicts that the editors consider relevant to the content of the manuscript have been disclosed.
References
- 1.WHO. World Malaria Report 2013. World Malaria Report 2013 2013:1–284.
- 2.Jagannathan P, Muhindo MK, Kakuru A, et al. Increasing incidence of malaria in children despite insecticide-treated bed nets and prompt anti-malarial therapy in Tororo, Uganda. Malar J 2012; 11:435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hoffman SL, Oster CN, Mason C, et al. Human lymphocyte proliferative response to a sporozoite T cell epitope correlates with resistance to falciparum malaria. J Immunol 1989; 142:1299–303. [PubMed] [Google Scholar]
- 4.Luty AJ, Lell B, Schmidt-Ott R, et al. Interferon-gamma responses are associated with resistance to reinfection with Plasmodium falciparum in young African children. J Infect Dis 1999; 179:980–8. [DOI] [PubMed] [Google Scholar]
- 5.Kurtis JD, Hollingdale MR, Luty AJ, Lanar DE, Krzych U, Duffy PE. Pre-erythrocytic immunity to Plasmodium falciparum: the case for an LSA-1 vaccine. Trends Parasitol 2001; 17:219–23. [DOI] [PubMed] [Google Scholar]
- 6.Reece WH, Pinder M, Gothard PK, et al. A CD4(+) T-cell immune response to a conserved epitope in the circumsporozoite protein correlates with protection from natural Plasmodium falciparum infection and disease. Nat Med 2004; 10:406–10. [DOI] [PubMed] [Google Scholar]
- 7.Todryk SM, Bejon P, Mwangi T, et al. Correlation of memory T cell responses against TRAP with protection from clinical malaria, and CD4 CD25 high T cells with susceptibility in Kenyans. PLoS One 2008; 3:e2027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Moormann AM, Sumba PO, Chelimo K, et al. Humoral and Cellular Immunity to Plasmodium falciparum Merozoite Surface Protein 1 and Protection From Infection With Blood-Stage Parasites. J Infect Dis 2013; 208:149–58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ogutu BR, Apollo OJ, McKinney D, et al. Blood-stage malaria vaccine eliciting high antigen-specific antibody concentrations confers no protection to young children in Western Kenya. PLoS One 2009; 4:e4708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bejon P, Lusingu J, Olotu A, et al. Efficacy of RTS,S/AS01E vaccine against malaria in children 5 to 17 months of age. N Engl J Med 2008; 359:2521–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Agnandji ST, Lell B, Fernandes JF, et al. A phase 3 trial of RTS,S/AS01 malaria vaccine in African infants. N Engl J Med 2012; 367:2284–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rts SCTP. Efficacy and safety of the RTS,S/AS01 malaria vaccine during 18 months after vaccination: a phase 3 randomized, controlled trial in children and young infants at 11 African sites. PLoS Med 2014; 11:e1001685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.John CC, Moormann AM, Sumba PO, Ofulla AV, Pregibon DC, Kazura JW. Gamma interferon responses to Plasmodium falciparum liver-stage antigen 1 and thrombospondin-related adhesive protein and their relationship to age, transmission intensity, and protection against malaria. Infect Immun 2004; 72:5135–42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Hill AV. Pre-erythrocytic malaria vaccines: towards greater efficacy. Nat Rev Immunol 2006; 6:21–32. [DOI] [PubMed] [Google Scholar]
- 15.Nussenzweig RS, Vanderberg J, Most H, Orton C. Protective immunity produced by the injection of x-irradiated sporozoites of Plasmodium berghei. Nature 1967; 216:160–2. [DOI] [PubMed] [Google Scholar]
- 16.Clyde DF, Most H, McCarthy VC, Vanderberg JP. Immunization of man against sporozite-induced falciparum malaria. Am J Med Sci 1973; 266:169–77. [DOI] [PubMed] [Google Scholar]
- 17.Hoffman SL, Goh LM, Luke TC, et al. Protection of humans against malaria by immunization with radiation-attenuated Plasmodium falciparum sporozoites. J Infect Dis 2002; 185:1155–64. [DOI] [PubMed] [Google Scholar]
- 18.Seder RA, Chang LJ, Enama ME, et al. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science 2013; 341:1359–65. [DOI] [PubMed] [Google Scholar]
- 19.Aponte JJ, Schellenberg D, Egan A, et al. Efficacy and safety of intermittent preventive treatment with sulfadoxine-pyrimethamine for malaria in African infants: a pooled analysis of six randomised, placebo-controlled trials. Lancet 2009; 374:1533–42. [DOI] [PubMed] [Google Scholar]
- 20.Wilson AL, Taskforce IP. A systematic review and meta-analysis of the efficacy and safety of intermittent preventive treatment of malaria in children (IPTc). PLoS One 2011; 6:e16976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Greenwood BM, David PH, Otoo-Forbes LN, et al. Mortality and morbidity from malaria after stopping malaria chemoprophylaxis. Trans R Soc Trop Med Hyg 1995; 89:629–33. [DOI] [PubMed] [Google Scholar]
- 22.Aponte JJ, Menendez C, Schellenberg D, et al. Age interactions in the development of naturally acquired immunity to Plasmodium falciparum and its clinical presentation. PLoS Med 2007; 4:e242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Belnoue E, Costa FT, Frankenberg T, et al. Protective T cell immunity against malaria liver stage after vaccination with live sporozoites under chloroquine treatment. J Immunol 2004; 172:2487–95. [DOI] [PubMed] [Google Scholar]
- 24.Putrianti ED, Silvie O, Kordes M, Borrmann S, Matuschewski K. Vaccine-like immunity against malaria by repeated causal-prophylactic treatment of liver-stage Plasmodium parasites. J Infect Dis 2009; 199:899–903. [DOI] [PubMed] [Google Scholar]
- 25.Roestenberg M, McCall M, Hopman J, et al. Protection against a malaria challenge by sporozoite inoculation. N Engl J Med 2009; 361:468–77. [DOI] [PubMed] [Google Scholar]
- 26.Roestenberg M, Teirlinck AC, McCall MB, et al. Long-term protection against malaria after experimental sporozoite inoculation: an open-label follow-up study. Lancet 2011; 377:1770–6. [DOI] [PubMed] [Google Scholar]
- 27.Friesen J, Silvie O, Putrianti ED, Hafalla JC, Matuschewski K, Borrmann S. Natural immunization against malaria: causal prophylaxis with antibiotics. Sci Transl Med 2010; 2:40ra9. [DOI] [PubMed] [Google Scholar]
- 28.Cockburn IA, Chen YC, Overstreet MG, et al. Prolonged antigen presentation is required for optimal CD8+ T cell responses against malaria liver stage parasites. PLoS Pathog 2010; 6:e1000877. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Jagannathan P, Eccles-James I, Bowen K, et al. IFNγ/IL-10 Co-producing Cells Dominate the CD4 Response to Malaria in Highly Exposed Children. PLoS Pathog 2014; 10:e1003864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kilama M, Smith DL, Hutchinson R, et al. Estimating the annual entomological inoculation rate for Plasmodium falciparum transmitted by Anopheles gambiae s.l. using three sampling methods in three sites in Uganda. Malar J 2014; 13:111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bigira V, Kapisi J, Clark TD, et al. Protective efficacy and safety of three antimalarial regimens for the prevention of malaria in young Ugandan children: a randomized controlled trial. PLoS Med 2014; 11:e1001689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Doolan DL, Southwood S, Freilich DA, et al. Identification of Plasmodium falciparum antigens by antigenic analysis of genomic and proteomic data. Proc Natl Acad Sci U S A 2003; 100:9952–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.van Schaijk BC, Janse CJ, van Gemert GJ, et al. Gene disruption of Plasmodium falciparum p52 results in attenuation of malaria liver stage development in cultured primary human hepatocytes. PLoS ONE 2008; 3:e3549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Huaman MC, Mullen GE, Long CA, Mahanty S. Plasmodium falciparum apical membrane antigen 1 vaccine elicits multifunctional CD4 cytokine-producing and memory T cells. Vaccine 2009; 27:5239–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Makobongo MO, Riding G, Xu H, et al. The purine salvage enzyme hypoxanthine guanine xanthine phosphoribosyl transferase is a major target antigen for cell-mediated immunity to malaria. Proc Natl Acad Sci U S A 2003; 100:2628–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Streeck H, Frahm N, Walker BD. The role of IFN-gamma Elispot assay in HIV vaccine research. Nat Protoc 2009; 4:461–9. [DOI] [PubMed] [Google Scholar]
- 37.Lamoreaux L, Roederer M, Koup R. Intracellular cytokine optimization and standard operating procedure. Nat Protoc 2006; 1:1507–16. [DOI] [PubMed] [Google Scholar]
- 38.Ho M, Webster HK, Looareesuwan S, et al. Antigen-specific immunosuppression in human malaria due to Plasmodium falciparum. J Infect Dis 1986; 153:763–71. [DOI] [PubMed] [Google Scholar]
- 39.Hviid L, Kurtzhals JA, Goka BQ, Oliver-Commey JO, Nkrumah FK, Theander TG. Rapid reemergence of T cells into peripheral circulation following treatment of severe and uncomplicated Plasmodium falciparum malaria. Infect Immun 1997; 65:4090–3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bejon P, Warimwe G, Mackintosh CL, et al. Analysis of immunity to febrile malaria in children that distinguishes immunity from lack of exposure. Infect Immun 2009; 77:1917–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Stowers AW, Chen Lh LH, Zhang Y, et al. A recombinant vaccine expressed in the milk of transgenic mice protects Aotus monkeys from a lethal challenge with Plasmodium falciparum. Proc Natl Acad Sci U S A 2002; 99:339–44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Stowers AW, Cioce V, Shimp RL, et al. Efficacy of two alternate vaccines based on Plasmodium falciparum merozoite surface protein 1 in an Aotus challenge trial. Infect Immun 2001; 69:1536–46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chelimo K, Embury PB, Sumba PO, et al. Age-related differences in naturally acquired T cell memory to Plasmodium falciparum merozoite surface protein 1. PLoS One 2011; 6:e24852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.O'Garra A, Vieira P. T(H)1 cells control themselves by producing interleukin-10. Nat Rev Immunol 2007; 7:425–8. [DOI] [PubMed] [Google Scholar]
- 45.Schofield L, Villaquiran J, Ferreira A, Schellekens H, Nussenzweig R, Nussenzweig V. Gamma interferon, CD8+ T cells and antibodies required for immunity to malaria sporozoites. Nature 1987; 330:664–6. [DOI] [PubMed] [Google Scholar]
- 46.Romero P, Maryanski JL, Corradin G, Nussenzweig RS, Nussenzweig V, Zavala F. Cloned cytotoxic T cells recognize an epitope in the circumsporozoite protein and protect against malaria. Nature 1989; 341:323–6. [DOI] [PubMed] [Google Scholar]
- 47.Schmidt NW, Podyminogin RL, Butler NS, et al. Memory CD8 T cell responses exceeding a large but definable threshold provide long-term immunity to malaria. Proc Natl Acad Sci U S A 2008; 105:14017–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Overstreet MG, Cockburn IA, Chen YC, Zavala F. Protective CD8 T cells against Plasmodium liver stages: immunobiology of an ‘unnatural’ immune response. Immunol Rev 2008; 225:272–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Olotu A, Moris P, Mwacharo J, et al. Circumsporozoite-Specific T Cell Responses in Children Vaccinated with RTS,S/AS01(E) and Protection against P falciparum Clinical Malaria. PLoS One 2011; 6:e25786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Epstein JE, Tewari K, Lyke KE, et al. Live Attenuated Malaria Vaccine Designed to Protect through Hepatic CD8+ T Cell Immunity. Science 2011; 334:475–80. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.