

Abstract
Neutrophils are the most abundant (40% to 75%) type of white blood cells and among the first inflammatory cells to migrate towards the site of inflammation. They are key players in the innate immune system and play major roles in cancer biology. Neutrophils have been proposed as key mediators of malignant transformation, tumor progression, angiogenesis and in the modulation of the antitumor immunity; through their release of soluble factors or their interaction with tumor cells. To characterize the specific functions of neutrophils, a fast and reliable method is coveted for in vitro isolation of neutrophils from human blood. Here, a density gradient separation method is demonstrated to isolate neutrophils as well as mononuclear cells from the blood. The procedure consists of layering the density gradient solution such as Ficoll carefully above the diluted blood obtained from patients diagnosed with chronic lymphocytic leukemia (CLL), followed by centrifugation, isolation of mononuclear layer, separation of neutrophils from RBCsby dextran then lysis of residual erythrocytes. This method has been shown to isolate neutrophils ≥ 90 % pure. To mimic the tumor microenvironment, 3-dimensional (3D) experiments were performed using basement membrane matrix such as Matrigel. Given the short half-life of neutrophils in vitro, 3D experiments with fresh human neutrophils cannot be performed. For this reason promyelocytic HL60 cells are differentiated along the granulocytic pathway using the differentiation inducers dimethyl sulfoxide (DMSO) and retinoic acid (RA). The aim of our experiments is to study the role of neutrophils on the sensitivity of lymphoma cells to anti-lymphoma agents. However these methods can be generalized to study the interactions of neutrophils or neutrophil-like cells with a large range of cell types in different situations.
Keywords: Immunology, Issue 109, Neutrophils, HL60diff, Lymphoma, 3D culture, Anti-lymphoma agents, Flow cytometry
Introduction
Innate immune cells constitute an essential proportion of the cells within the tumor microenvironment and have been associated with tumor malignancy in patients and animal models of cancer1. Recently, it has become more widely appreciated that chronic immune responses play critical roles in promoting tumor progression, metastasis and resistance to chemotherapies2. Macrophages are important innate immune cells that have been shown to directly regulate tumor cell response to chemotherapy 3,4. However, the role of neutrophils, key players in the innate immune system, in regulating tumor response to anti-cancer treatment is not known. The aims of these protocols are to use a fast and credible method to separate neutrophils from CLL patient's blood samples and to differentiate HL60 cells along the granulocytic pathway in order to study their role in regulating the sensitivity of lymphoma cells to anti-lymphoma agents.
Neutrophils are the most abundant cellular component of the innate immune system in blood 5 and act as a first line of defense against invading microorganisms 6. Neutrophils have an essential role in rising effective innate immune responses in addition to variable effector functions in several pathological conditions 7. Therefore, a fast and credible method to isolate neutrophils from other blood cells, such as density gradient separation method, is required for in vitro studies. Using this method for neutrophil isolation will facilitate further research on neutrophil-mediated immunological functions in vivo and ex vivo.
The ability to obtain pure populations of neutrophils is an important first step for the investigation of patients with immunological diseases 8. Density gradient separation method is an ideal technique in which a high yield of cells is obtained. The method involves the addition of density gradient solution in the bottom of a tube containing diluted human blood followed by centrifugation at 300 g for 35 min without break. The ring of the mononuclear cells appears at the interface and the neutrophils reside below the former. This method have significant advantages with respect to other available methods such as neutrophil isolation kits which are much more expensive9. In addition, isolating neutrophils from human blood by commercial kits using antibodies directed to a surface marker specific for human neutrophils, increase the risk of cell activation or differentiation. Density gradient separation method allows the isolation of neutrophils within a short period of time. Within the same step, mononuclear cells are also separated and recovered. It's a fundamental technique in which a high yield of pure cells is obtained in order to achieve functional integrity.
In order to mimic the tumor microenvironment, 3D experiments were performed. Given the short half-life of neutrophils in vitro, the 3D experiments with fresh human neutrophils are not conclusive. For this reason, promyelocytic (HL60) cells are induced to differentiate into neutrophil-like cells using the differentiation inducers dimethyl sulfoxide (DMSO) and retinoic acid (RA). Using differentiated HL60 cells (HL60diff) will prevent having different responses of neutrophils due to isolation from different donors.
In vitro 3D culture models represent an intermediate stage between in vitro 2D models and in vivo models. In 2D culture, the cells spread on plastic surface forming unnatural cell attachments to deposited proteins that are denatured on this synthetic surface. Conversely, the cells in 3D culture form natural cell-cell attachments since the cells and the extracellular matrix they synthesize are the natural material to which they are attached. For this reason, 3D co-culture models, especially between cancer cells and other cell types, have been very useful for indicating their contribution to tumor growth, angiogenesis, and metastasis. As a result, 3D cultures make the cell culture mimic the physiological conditions that exist in vivo 10.
Protocol
1. Neutrophil Isolation and Co-culture with Primary Leukemic Cells
NOTE: Procedures were conducted under the approval of Lyon Hospital Ethics committee with all patients signing informed consent.
- Isolation of Primary Leukemic Cells and Neutrophils
- Collect tubes of peripheral blood on EDTA (1.8 mg EDTA per milliliter of blood) from patients diagnosed with chronic lymphocytic leukemia (CLL).
- Add each 15 ml of blood to a sterile 50 ml tube and dilute with 15 ml RPMI (dilution 1:1), then carefully and slowly add 15 ml of density gradient solution to the bottom of the tube without mixing the phases. Ensure that the density gradient solution is at RT for the preparation.
- Centrifuge at 300 x g for 35 min at RT and without brake. The blood should separate into four distinct phases as shown in Figure 1A, top to bottom: platelet and plasma, mononuclear cells (white ring), density gradient solution, granulocytes and erythrocytes.
- Collect the white ring which represents the primary leukemic cells with a plastic Pasteur pipette and transfer to a new 50 ml tube.
- Fill the tube with PBS (contains calcium and magnesium) up to 50 ml in total and centrifuge at 300 g for 10 min at RT.
- Resuspend the pellet with 5 ml PBS then add PBS up to 50 ml in total and centrifuge at 300 g for 10 min at RT.
- Resuspend the pellet with complete RPMI medium (RPMI 1640 supplemented with 10% fetal bovine serum (FBS), 2 mM glutamine, 100 U/ml penicillin and 100 mg/ml streptomycin) for counting using cell viability counter.
- Aspirate the upper phases leaving the granulocytes and erythrocytes phase and add PBS up to 25 ml then add 3% dextran in 0.9% NaClup to 50 ml in total. Mix the tubes 10 times and keep them for 30 min at RT without mixing.
- Collect the upper RBC-poor neutrophil layer and place them into a clean sterile 50 ml tube and centrifuge the tubes at 300 g for 10 min at RT. Resuspend each pellet with 5 ml PBS then add red cell lysis buffer up to 50 ml in total.
- Keep the tubes in dark for 15 min at RT then centrifuge at 500 g for 10 min at RT. Wash the pellets with PBS (add PBS up to 50 ml in total) then centrifuge at 500 g for 10 min at RT.
- Resuspend the pellets with complete RPMI medium for counting using cell viability counter. Keep 3 x 105 of each cell type in 50 µl PBS-FBS (4%) in 5 ml plastic tubes to determine the purity of their isolation using flow cytometry.
- Analyze the Morphological Appearances of the Isolated Cell Populations
- To do so, resuspend each 3 x 104 neutrophils and 3 x 104 mononuclear cells in 150 µl complete RPMI medium. Assemble the glass slides, filter cards, and sample chambers in the cytospin centrifuge, ensuring that the centrifuge is well balanced.
- Place each cell type into sample chamber and cyto-centrifuge at 750 x g for 10 min at RT Remove the slides, filter cards, and sample chambers from the centrifuge. Disassemble carefully, so as not to damage the cells on the slide. Discard filter cards and sample chambers.
- Stain the cells using Giemsa staining kit and keep the slides for 1 hr to dry. Examine the cells by microscope with 100X magnification.
- Test the Purity of Isolated Cells by FACS
- Label the isolated primary leukemic cells with anti-human CD19 antibody conjugated to APC (5 µl/106 cells). Label the purified neutrophils with mixture of anti-human antibodies listed in Table 1 (5 µl/106 cells). Incubate the cells in dark for 30 min at 4 oC.
- Wash the cells with 500 µl PBS-FBS (4%) the centrifuge the tubes at 300 g for 5 min at RT.
- Resuspend each pellet in 200 µl PBS-FBS (4%).
- Analyze on a flow cytometer using the following optical configuration: 488 nm laser: FSC-A (488 nM), SSC-A (488/10BP), FITC (530/30 BP), PerCP-Cy5.5 (695/40 BP), PE (575/26 BP); 633 nm laser: APC (660/20 BP), APC-Cy7 (780/60 BP). Ensure the color compensation.
- Coculture of Primary Leukemic Cells with Autologous Neutrophils
- Seed 2 x 105 cells/ml of primary leukemic cells alone or with autologous neutrophils at ratio 1:10 in complete RPMI medium. To do so, adjust cell numbers by diluting cell suspensions and add 2 x 105 cells/ml of primary leukemic cells to 2 x 106 cells/ml neutrophils. Mix carefully.
- Add 25 µM of Bruton's tyrosine kinase (Btk) inhibitor (ibrutinib) to the cells. Incubate for 24 hr at 37 °C with 5% CO2.
- Cell Harvest and FACS Analysis
- Collect the cells in 5 ml plastic tubes for FACS analysis and centrifuge the tubes at 300 g for 5 min at RT. Wash the pellets with 1 ml PBS-FBS (4%) then centrifuge at 300 g for 5 min at RT.
- Resuspend pellets in Annexin V and PI using commercial kit and incubate the cells in dark for 10 min at RT. Analyze on a flow cytometer using the gating strategy of Figure 2A and the following optical configuration: 488 nm laser: FSC-A (488 nM), SSC-A (488/10BP), FITC (530/30 BP), PI (610/20 BP). Ensure the color compensation.
2. Differentiation of HL60 Cells along the Granulocytic Pathway and Their Coculture with RL Lymphoma B Cells in 3D Model
- Differentiation of Human Promyelocytic (HL60) Cells into Neutrophil-like Cells
- Adjust HL60 cell number to 3 x 105cells/ml then add 1 µM retinoic acid (RA) and 1.25% DMSO to induce their differentiation. Seed each 3 x 105 HL60 cells per well in 48-well plate then incubate for 48 hr at 37 °C with 5% CO2.
- Later, collect the cells and centrifuge at 300 g for 5 min at RT. Resuspend the cells with complete RPMI medium for counting using cell viability counter.
- Then dilute cell suspension in complete RPMI medium to adjust HL60 cell number to 3 x 105 cells/ml and induce again their differentiation by adding 1 µM retinoic acid (RA) and 1.25 % DMSO. Seed each 3 x 105 HL60 cells per well in 48-well plate and incubate for another 48 hr at 37 °C with 5% CO2.
- Analysis of HL60 differentiation (HL60diff)
- Collect the cells and centrifuge the tubes at 300 g for 5 min at RT. Resuspend the pellet with complete RPMI medium for counting using cell viability counter.
- To analyze the changes in cell-surface markers expression by flow cytometry. Place each 3 x 105 undifferentiated HL60 cells (from cell culture) and 3 x 105 HL60diff cells in 5 ml plastic tubes for FACS analysis and centrifuge the tubes at 300 g for 5 min at RT.
- Resuspend the pellets in 50 µl PBS-FBS (4%). Label the cells with anti-human CD11b conjugated to AF700 or anti-human CD38 conjugated to APC (5 µl/106 cells). Incubate the cells in dark for 30 min at 4 oC.
- Wash with 500 µl PBS-FBS (4%) then centrifuge at 300 g for 5 min at RT. Resuspend the pellets in 200 µl PBS-FBS (4%).
- Analyze on flow cytometer using the gating strategy of Figure 3A and the following optical configuration: 488 nm laser: FSC-A (488 nM), SSC-A (488/10BP); 633 nm laser: APC (660/20 BP), Alexa Fluor 700 (730/45 BP).
- To test the morphological changes in HL60diff, immobilize the cells onto glass slides for microscopic examination.
- To do so, resuspend each 3 x 104 undifferentiated HL60 cells (from cell culture) and 3 x 104 HL60diff in 150 µl complete RPMI medium. Assemble the glass slides, filter cards, and sample chambers in the cytospin centrifuge, ensuring that the centrifuge is balanced.
- Place each cell type into sample chamber and cyto-centrifuge at 1,500 rpm for 10 min at RT. Remove the slides, filter cards, and sample chambers from the centrifuge. Disassemble carefully, so as not to damage the cells on the slide. Discard filter cards and sample chambers.
- Stain the cells using Giemsa staining kit and keep the slides for 1 hr to dry. Examine the cells by microscope with 100X magnification.
- 3-dimensional (3D) Culture NOTE: Ensure that the materials used in this experiment are cold and the experiment is taking place on ice.
- Resuspend each 5 x 104 RL cells, either alone or mixed with HL60diff at ratio RL:HL60diff 1:10, with 300 µl basement membrane matrix using 1 ml pipette tip cut with a sterile scissor in order to widen the opening to about 2 to 3 mm. Avoid bubbles during this step.
- Seed each 300 µl of cell suspension/well in 24-well plate. Avoid bubbles during this step. Incubate the plate for 30 min at 37 °C with 5% CO2then add 1 ml complete RPMI medium for each well and incubate for 7 days at 37 °C with 5% CO2. Change the medium every two days. Add 10 nM vincristine at day 5.
- FACS Analysis
- After 7 days of culture, aspirate the medium and wash each well with 1 ml ice-cold PBS twice. Add 3 ml/well of ice-cold PBS-EDTA (5 mM). Detach the gel from the bottom of the well by scraping using the bottom of 200 µl pipette tip. Shake the plate gently on ice for 30 min.
- Transfer cell suspension into sterile 15 ml tube and shake the tubes gently on ice for another 30 min. Check for the appearance of homogeneous cell suspension. (If it's not the case, then shake the cells for longer time or add more PBS-EDTA).
- Centrifuge the tubes at 300 x g for 10 min at RT. Wash the pellets with PBS then centrifuge at 300 x g for 10 min at RT.
- Resuspend with PBS-FBS (4%) and label with anti-human CD19 antibody conjugated to PE-Cy7 and anti-human CD38 antibody conjugated to APC (5 µl/106 cells). Incubate in dark for 30 min at 4 oC.
- Wash with 500 µl PBS-FBS (4%) then centrifuge tubes at 300 g for 5 min at RT. Resuspend the cells with Annexin V and PI using commercial kit.
- Analyze on flow cytometer using the gating strategy of Figure 4A and the following optical configuration: 488 nm laser: FSC-A (488 nM), SSC-A (488/10BP), FITC (530/30 BP), PI (610/20 BP), PE-Cy7 (780/60 BP); 633 nm laser: APC (660/20 BP). Ensure the color compensation.
Representative Results
Density gradient separation method described here provides primary leukemic cells and unstimulated neutrophils isolated from the blood of CLL patients. Figure 1A represents the different blood layers obtained after density gradient centrifugation (from top to bottom: platelets and plasma, white ring represents the mononuclear cells, density gradient solution, granulocytes and erythrocytes). Figure 1B and 1C show the differences in the morphological appearances between neutrophils (multi-lobed nuclei cells) and mononuclear cells respectively.
Results in Figure 1D represent the forward-scatter (FSC) vs side-scatter (SSC) plot of primary leukemic cells (mononuclear cells). Labeling this population with anti-human CD19 conjugated to APC shows a complete right shift of the histogram (Figure 1E) with only one peak which indicates that this population is positive for CD19 and pure. Neutrophils are also ≥90% pure as verified by flow cytometry after labeling the isolated neutrophils with a mixture of fluorochrome-conjugated monoclonal antibodies listed in Table 1. Figure 1F represents FSC vs SSC scatter plot of the isolated neutrophils which are positive for CD45 (Figure 1G), positive for CD15 and negative for CD14 (Figure 1H), positive for both CD15 and CD16 (Figure 1I), positive for CD16 and negative for CD56 (Figure 1J).
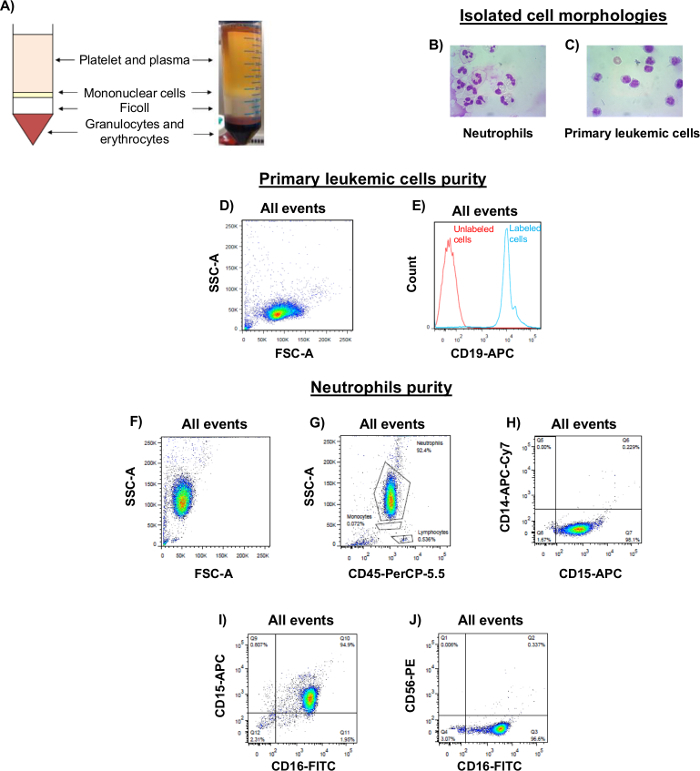 Figure 1. Analysis of Morphological Appearances and Purity of Primary Leukemic Cells and Neutrophils Isolated from Patients' Blood. (A) Schematic view represents different blood layers after density gradient centrifuging. (B-C) Giemsa staining represents the morphological differences between (B) neutrophils and (C) primary leukemic cells (mononuclear cells). Photos were taken by microscope with 100X magnification. (D) Forward-scatter (FSC) and side-scatter (SSC) plot represents the isolated primary leukemic cell population. (E) Isolated primary leukemic cells were labeled with anti-CD19-APC and analyzed for the expression of CD19. The red line indicates the unlabeled cells and the blue line indicates cells labeled with anti-CD19-APC. (F) FSC vs SSC scatter plot represents the isolated neutrophil population. (G-J) Isolated neutrophils were labeled with a mixture of antibodies listed in Table 1 and analyzed for the expression of (G) CD45, (H) CD14 and CD15, (I) CD15 and CD16, (J) CD16 and CD56. Please click here to view a larger version of this figure.
Figure 1. Analysis of Morphological Appearances and Purity of Primary Leukemic Cells and Neutrophils Isolated from Patients' Blood. (A) Schematic view represents different blood layers after density gradient centrifuging. (B-C) Giemsa staining represents the morphological differences between (B) neutrophils and (C) primary leukemic cells (mononuclear cells). Photos were taken by microscope with 100X magnification. (D) Forward-scatter (FSC) and side-scatter (SSC) plot represents the isolated primary leukemic cell population. (E) Isolated primary leukemic cells were labeled with anti-CD19-APC and analyzed for the expression of CD19. The red line indicates the unlabeled cells and the blue line indicates cells labeled with anti-CD19-APC. (F) FSC vs SSC scatter plot represents the isolated neutrophil population. (G-J) Isolated neutrophils were labeled with a mixture of antibodies listed in Table 1 and analyzed for the expression of (G) CD45, (H) CD14 and CD15, (I) CD15 and CD16, (J) CD16 and CD56. Please click here to view a larger version of this figure.
| Antigens | Fluorochrome | Clone |
| CD14 | APC-Vio770 (Cy-7) | TÜK4 |
| CD15 | APC | VIMC6 |
| CD16 | FITC | VEP13 |
| CD45 | PerCP-Vio700 (Cy5.5) | 5B1 |
| CD56 | PE | AF12-7H3 |
Table 1. Fluorochrome-conjugated Purified Monoclonal Antibodies Used to Test the Purity of Isolated Neutrophils. All antibodies were obtained from Miltenyi biotech.
To investigate the effect of neutrophils on sensitivity of primary leukemic cells (CLL) to ibrutinib, the latter were cultured alone or with autologous neutrophils (N) for 24 hr at ratio N:CLL 10:1, in the presence or absence of ibrutinib at 25 µM. The percentage of alive primary leukemic cells (Annexin V negative/PI negative) was measured by double staining with annexin V-FITC and PI, followed by flow cytometric analysis. FSC vs SSC scatterplot in Figure 2A represents the gates of neutrophils and primary leukemic cells. Neutrophils are much more granular than the primary leukemic cells so they appear with higher SSC. Gating on the primary leukemic cells co-cultured with neutrophils in the presence of ibrutinib, results show higher percentage of alive cells (Figure 2C, 84.8%) compared to the cells cultured alone (Figure 2B, 53.1%). The box graph in Figure 2D shows that the decrease of cell viability induced by ibrutinib was significantly inhibited by the presence of neutrophils (40.1 ± 3.1 vs 60.1 ± 3.5, p <0.001, 19 patients).
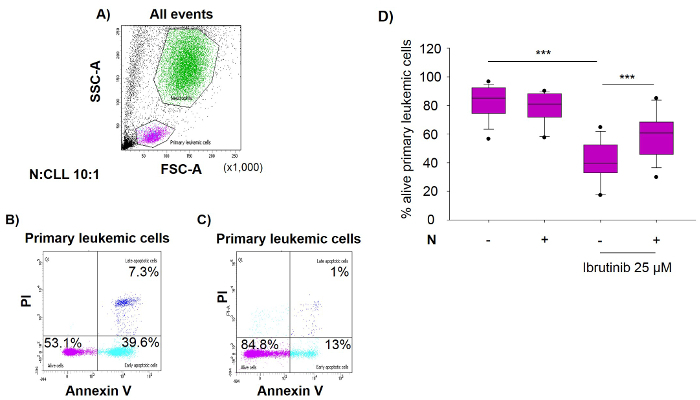 Figure 2. Autologous Neutrophils Protect Primary Leukemic Cells against Ibrutinib. Blood was collected from patients diagnosed with chronic lymphocytic leukemia (CLL). Primary leukemic cells were isolated and cultured alone or together with autologous neutrophils (N) at primary leukemic cells:Nratio 1:10 for 24 hr, in the presence or absence of 25 µM ibrutinib. The percentage of live primary leukemic cells (Annexiv V negative/PI negative) was measured by double staining with annexin V-FITC and PI, followed by flow cytometric analysis. (A) Forward-scatter (FSC) and side-scatter (SSC) plot represents the gates of neutrophils and primary leukemic cells. (B) Bi-dimensional dot-blot shows the percentages of alive and apoptotic primary leukemic cells cultured alone and treated with 25 µM ibrutinib. (C) Bi-dimensional dot-blot shows the percentages of alive and apoptotic primary leukemic cells co-cultured with neutrophils and treated with 25 µM ibrutinib. (D) Box plot represents the percentage of alive primary leukemic cells of 19 patients. Data are expressed as mean ± SD. ***p <0.001 Please click here to view a larger version of this figure.
Figure 2. Autologous Neutrophils Protect Primary Leukemic Cells against Ibrutinib. Blood was collected from patients diagnosed with chronic lymphocytic leukemia (CLL). Primary leukemic cells were isolated and cultured alone or together with autologous neutrophils (N) at primary leukemic cells:Nratio 1:10 for 24 hr, in the presence or absence of 25 µM ibrutinib. The percentage of live primary leukemic cells (Annexiv V negative/PI negative) was measured by double staining with annexin V-FITC and PI, followed by flow cytometric analysis. (A) Forward-scatter (FSC) and side-scatter (SSC) plot represents the gates of neutrophils and primary leukemic cells. (B) Bi-dimensional dot-blot shows the percentages of alive and apoptotic primary leukemic cells cultured alone and treated with 25 µM ibrutinib. (C) Bi-dimensional dot-blot shows the percentages of alive and apoptotic primary leukemic cells co-cultured with neutrophils and treated with 25 µM ibrutinib. (D) Box plot represents the percentage of alive primary leukemic cells of 19 patients. Data are expressed as mean ± SD. ***p <0.001 Please click here to view a larger version of this figure.
The short half-life of neutrophils in vitro makes their use in 3D culture ineffectual. Differentiation of HL60 cells along the granulocytic pathway was induced by differentiation inducers. Two different parameters (cell-surface markers expression and morphological changes) were used to measure the differentiation of HL60. FSC vs SSC scatter plot in Figure 3A show that HL60 cells are larger in size compared to HL60diffcells with higher FSC. Labeling the cells (HL60 or HL60diff)with anti-human CD11b antibody conjugated to AF700 and anti-human CD38antibody conjugated to APC shows an increase in CD11b and CD38 expression (Figure 3B) upon differentiation which is an indicative of granulocytic differentiation. HL60diffcells also show morphological changes detected by the appearance of multi-lobed nuclei (Figure 3C).
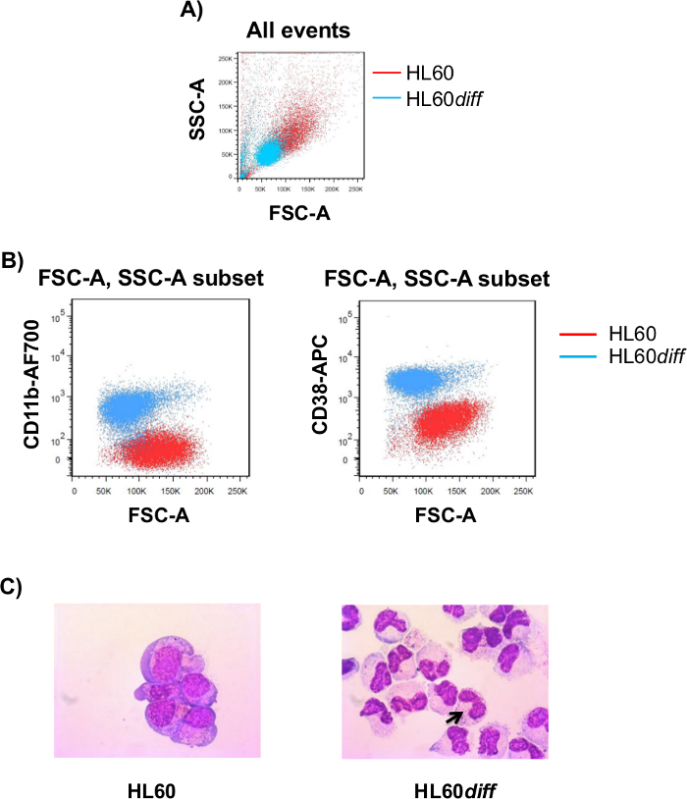 Figure 3. Structural Parameters of Differentiated HL60 Cells. (A) Forward-scatter (FSC) and side-scatter (SSC) plots represent HL60 and differentiated HL60 cells (HL60diff). (B) HL60 and HL60diff cells were labeled with anti-CD11b-AF700 or anti-CD38-APC followed by flow cytometry analysis. After gating on each cell population in the FSC-A vs. SSC-A scatter plot (A), the cells were analyzed for the expressions of CD11b or CD38. (C) HL60 cells show morphological changes consistent with differentiation toward granulocytes. Photos were taken by microscope with 100X magnification. Bold black arrow point the multi-lobed nucleus. Graphs are representative of five independent experiments. Please click here to view a larger version of this figure.
Figure 3. Structural Parameters of Differentiated HL60 Cells. (A) Forward-scatter (FSC) and side-scatter (SSC) plots represent HL60 and differentiated HL60 cells (HL60diff). (B) HL60 and HL60diff cells were labeled with anti-CD11b-AF700 or anti-CD38-APC followed by flow cytometry analysis. After gating on each cell population in the FSC-A vs. SSC-A scatter plot (A), the cells were analyzed for the expressions of CD11b or CD38. (C) HL60 cells show morphological changes consistent with differentiation toward granulocytes. Photos were taken by microscope with 100X magnification. Bold black arrow point the multi-lobed nucleus. Graphs are representative of five independent experiments. Please click here to view a larger version of this figure.
To test the effect of HL60diff cells on regulating RL cell response to vincristine in 3D model, RL cells were cultured alone or with HL60diff in basement membrane matrix in the presence or absence of vincristine at 10 nM. Using the gating strategy shown in Figure 4A, both populations are discriminated based on CD19 and CD38 expression; RL cells positive for CD19 and HL60diff cells positive for CD38. Gating on RL cells co-cultured with HL60diff cells in the presence of vincristine, results show higher percentage of alive cells (Figure 4C, 35.9%) compared to RL cells cultured alone (Figure 4B, 19.7%). The bar graph in Figure 4D shows a significant increase in the percentage of alive RL cells (CD19 positive Annexin V negative/PI negative) in the presence of HL60diff cells (19.5 ± 0.2 vs 33.1 ± 1.0, p <0.001).
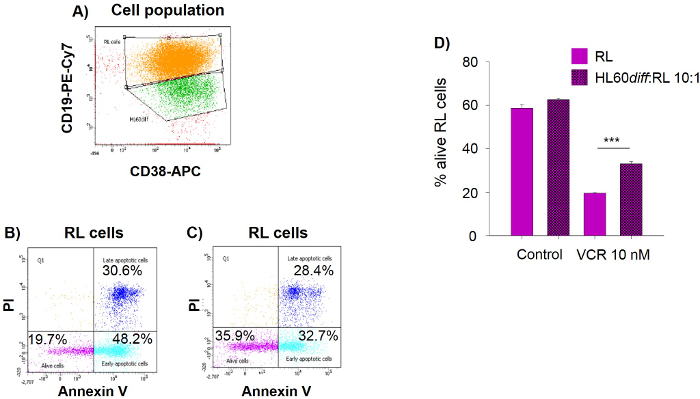 Figure 4. Neutrophil-like HL60diff Cells Protect RL Lymphoma Cells against Vincristine in 3D Culture. RL cells were cultured alone or together with HL60diff cells at RL:HL60diff ratio 1:10 for 7 days in basement membrane matrix. On day 5, vincristine (VCR) was added at a concentration of 10 nM. Spheroids were dissociated on day 7 and cells were labeled with anti-CD19-PECy7 and anti-CD38-APC then resuspended with annexin V-FITC and PI followed by flow cytometric analysis. (A) Forward-scatter (FSC) and side-scatter (SSC) plot represents the gates of RL cells and HL60diff cells. (B) Bi-dimensional dot-blot shows the percentages of alive and apoptotic RL cells cultured alone and treated with 10 nM vincristine. (C) Bi-dimensional dot-blot shows the percentages of alive and apoptotic RL cells co-cultured with HL60diff and treated with 10 nM vincristine. (D) The bar graph represents the percentage of alive RL cells. Data are expressed as mean ± SD. ***p ≤0.001. Please click here to view a larger version of this figure.
Figure 4. Neutrophil-like HL60diff Cells Protect RL Lymphoma Cells against Vincristine in 3D Culture. RL cells were cultured alone or together with HL60diff cells at RL:HL60diff ratio 1:10 for 7 days in basement membrane matrix. On day 5, vincristine (VCR) was added at a concentration of 10 nM. Spheroids were dissociated on day 7 and cells were labeled with anti-CD19-PECy7 and anti-CD38-APC then resuspended with annexin V-FITC and PI followed by flow cytometric analysis. (A) Forward-scatter (FSC) and side-scatter (SSC) plot represents the gates of RL cells and HL60diff cells. (B) Bi-dimensional dot-blot shows the percentages of alive and apoptotic RL cells cultured alone and treated with 10 nM vincristine. (C) Bi-dimensional dot-blot shows the percentages of alive and apoptotic RL cells co-cultured with HL60diff and treated with 10 nM vincristine. (D) The bar graph represents the percentage of alive RL cells. Data are expressed as mean ± SD. ***p ≤0.001. Please click here to view a larger version of this figure.
Discussion
Described here, an effective, simple, fast and inexpensive protocol for the isolation of neutrophils from human blood with high purity using density gradient centrifugation approach and within the same step mononuclear cells are also separated and recovered. The isolated cell populations are ≥90% pure.
Several methods are available for neutrophil isolation from human blood. These include similar methods using discontinuous gradients11,12, or using commercial kits for neutrophil isolation by positive immuno-magnetic selection during which neutrophils are immuno-magnetic labeled with specific antibody such as anti-human CD16 then neutrophils enriched by binding of the CD16-positive cells on a magnetic column 13. Commercial kits with negative immuno-magnetic selection are also available during which the whole blood or granulocyte suspension are labeled with a cocktail of antibodies that bind on cells other than neutrophils 9( i.e., antibody complexes that label RBCs, platelets and unwanted cells such as CD2, CD3, CD9, CD19, CD36, CD56, glycophorin A and dextran-coated magnetic particles), then neutrophils are enriched by elution as the antibody-negative fraction of cells that does not bind on the magnetic column. In addition, neutrophils can be isolated by Fluorescence Activated Cell Sorting 14.
Some techniques have been shown to provide high yields of pure neutrophils, such as positive immuno-magnetic selection. However, this method has disadvantages compared to density gradient centrifugation methods due to the labeling agents that bind on the surface of neutrophils and this could potentially alter their function by inducing their activation or differentiation. Also using the positive immuno-magnetic selection method, the antibody-labeled neutrophils bind on a magnetic column for separation and this may also affect their function. Fluorescence Activated Cell Sorting has another limitation insofar as this method requires longer time to collect the cells which may negatively affect neutrophil survival due to their short half-life.
Density gradient centrifugation method are very comparable and the neutrophil purity can exceed 90 % using both procedures, but the former is a two-layer gradient which is technically less challenging compared to layering gradient solution which consist of 40%, 60% and 80% vol/vol in PBS. It was mentioned by Swamydas et al. 15that intermixing of the gradient solution interfaces often takes place due to the small density differences between the three layers. With respect to the negative immuno-magnetic selection approaches, they have also been reported to be effective for neutrophil isolation with high purity and viability9. Their advantage over positive immuno-magnetic selection and Fluorescence Activated Cell Sorting is that neutrophils are not labeled with any labeling agents and do not bind to magnetic column, thus avoiding cell activation but this method is much more expensive and more time consuming compared to density gradient method.
Neutrophils normally undergo rapid spontaneous apoptosis both in vitro and in vivo16,17. HL60promyelocyticcells retain many characteristics of human leukocyte progenitors, such as the potential to undergo differentiation into neutrophils 18. Unlike neutrophils, HL60diff cells do not rapidly undergo apoptosis and using these cells solves the trouble of having different responses of neutrophils due to their isolation from different donors.
Recently, 3D cultures become more mature and pertinent to human and animal physiology, and an attractive model for different biological research especially in the field of cancer. The pattern shift from 2D to 3D culture is progressing rapidly since the latter is more conspicuous during studying various pathological conditions such as cancer 19. There is an increasing awareness of the drawbacks of 2D culture where adding a third dimension to a cell's environment is important in order to take a closer look at the importance of cellular interactions 20 and to study the differences in cellular behavior and characteristics. 3D culture creates an environment similar to conditions in a living organism (e.g., human being) leading to more relevant research. However, 3D culture involves the complex interplay between different partners such as cells, extracellular matrices and interstitial fluids which do not present in 2D culture. Thus, efforts will be required in order to establish method calibration and to count on good laboratory practices as well as effective supplies especially the commercial brands that are extensively tested and monitored.
The tumor microenvironment consists of multiple cell types, including many immune cell populations that participate in and regulate tumorigenesis, metastasis and response to anticancer agents 21,22. Several studies have shown that an increase of neutrophil infiltration in tumors is significantly correlated with acquired resistance to several anti-cancer agents such as anti-VEGF therapy23,24. Furthermore, elevation in the pretreatment neutrophil count in patients with metastatic renal cell carcinoma 25, as well as the presence of a high level of intra-tumoral neutrophils in patients with different solid tumors,26 have been proposed as prognostic factors for poor survival. Our study suggests that neutrophils may play a role in protecting lymphoma B cells from anticancer agent-induced apoptosis.
In summary, the reliable and reproducible method described here can be employed by any laboratory to collect neutrophils and mononuclear cells from human blood. In addition, the differentiation of HL60 cells along the granulocytic pathway is presented in order to avoid some neurophil problems such as the rapid spontaneous apoptosis. These approaches were used to study the role of neutrophils on sensitivity of lymphoma cells to anti-lymphoma agents where neutrophils show a protective effect on lymphoma cells against these agents using 2D and 3D culture systems. These approaches can be used for a variety of neutrophil functional studies that should deepen our understanding of neutrophil role in cancer biology. In particular this method can be used to better understand the cross-talk between neutrophils and tumor cells or between neutrophils and other components of the microenvironment.
Disclosures
The authors have nothing to disclose.
Acknowledgments
This work was supported by the Institute National du Cancer (INCa-DGOS-4664).
References
- Gocheva V, et al. IL-4 induces cathepsin protease activity in tumor-associated macrophages to promote cancer growth and invasion. Genes Dev. 2010;24:241–255. doi: 10.1101/gad.1874010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grivennikov SI, Greten FR, Karin M. Immunity, Inflammation, and Cancer. Cell. 2010;140:883–899. doi: 10.1016/j.cell.2010.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchem JB, et al. Targeting tumor-infiltrating macrophages decreases tumor-initiating cells, relieves immunosuppression, and improves chemotherapeutic responses. Cancer Res. 2013;73:1128–1141. doi: 10.1158/0008-5472.CAN-12-2731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeNardo DG, et al. Leukocyte Complexity Predicts Breast Cancer Survival and Functionally Regulates Response to Chemotherapy. Cancer Discov. 2011;1:54–67. doi: 10.1158/2159-8274.CD-10-0028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Carlo E, et al. The intriguing role of polymorphonuclear neutrophils in antitumor reactions. Blood. 2001;97:339–345. doi: 10.1182/blood.v97.2.339. [DOI] [PubMed] [Google Scholar]
- Kumar V, Sharma A. Neutrophils: Cinderella of innate immune system. Int. Immunopharmacol. 2010;10:1325–1334. doi: 10.1016/j.intimp.2010.08.012. [DOI] [PubMed] [Google Scholar]
- Mantovani A, Cassatella MA, Costantini C, Jaillon S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011;11:519–531. doi: 10.1038/nri3024. [DOI] [PubMed] [Google Scholar]
- Kelbaek H. Sterile isolation of polymorphonuclear leukocytes from large blood volumes. J. Clin. Chem. Clin. Biochem. Z. FürKlin. Chem. Klin. Biochem. 1985;23:17–20. [PubMed] [Google Scholar]
- Aynaud M-M, et al. Human Tribbles 3 protects nuclear DNA from cytidine deamination by APOBEC3A. J. Biol. Chem. 2012;287:39182–39192. doi: 10.1074/jbc.M112.372722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ziòłkowska K, et al. Long-term three-dimensional cell culture and anticancer drug activity evaluation in a microfluidic chip. Biosens.Bioelectron. 2013;40:68–74. doi: 10.1016/j.bios.2012.06.017. [DOI] [PubMed] [Google Scholar]
- Eggleton P, Gargan R, Fisher D. Rapid method for the isolation of neutrophils in high yield without the use of dextran or density gradient polymers. J. Immunol. Methods. 1989;121:105–113. doi: 10.1016/0022-1759(89)90425-0. [DOI] [PubMed] [Google Scholar]
- Behnen M, et al. Immobilized immune complexes induce neutrophil extracellular trap release by human neutrophil granulocytes via FcγRIIIB and Mac-1. J. Immunol. Baltim.Md 1950. 2014;193:1954–1965. doi: 10.4049/jimmunol.1400478. [DOI] [PubMed] [Google Scholar]
- Hirsch G, Lavoie-Lamoureux A, Beauchamp G, Lavoie J-P. Neutrophils are not less sensitive than other blood leukocytes to the genomic effects of glucocorticoids. PloS One. 2012;7:44606. doi: 10.1371/journal.pone.0044606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dorward DA, et al. Technical advance: autofluorescence-based sorting: rapid and nonperturbing isolation of ultrapure neutrophils to determine cytokine production. J. Leukoc. Biol. 2013;94:193–202. doi: 10.1189/jlb.0113040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swamydas M, Lionakis MS. Isolation, Purification and Labeling of Mouse Bone Marrow Neutrophils for Functional Studies and Adoptive Transfer Experiments. J Vis Exp. 2013. p. e50586. [DOI] [PMC free article] [PubMed]
- Scaife H, Woldehiwet Z, Hart CA, Edwards SW. Anaplasmaphagocytophilum reduces neutrophil apoptosis in vivo. Infect. Immun. 2003;71:1995–2001. doi: 10.1128/IAI.71.4.1995-2001.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maianski NA, Maianski AN, Kuijpers TW, Roos D. Apoptosis of neutrophils. ActaHaematol. 2004;111:56–66. doi: 10.1159/000074486. [DOI] [PubMed] [Google Scholar]
- Dalton WT, et al. HL-60 cell line was derived from a patient with FAB-M2 and not FAB-M3. Blood. 1988;71:242–247. [PubMed] [Google Scholar]
- Hogan C, Kajita M, Lawrenson K, Fujita Y. Interactions between normal and transformed epithelial cells: Their contributions to tumourigenesis. Int. J. Biochem. Cell Biol. 2011;43:496–503. doi: 10.1016/j.biocel.2010.12.019. [DOI] [PubMed] [Google Scholar]
- Page H, Flood P, Reynaud EG. Three-dimensional tissue cultures: current trends and beyond. Cell Tissue Res. 2013;352:123–131. doi: 10.1007/s00441-012-1441-5. [DOI] [PubMed] [Google Scholar]
- Mantovani A. Macrophages, Neutrophils, and Cancer: A Double Edged Sword. New J. Sci. 2014. p. 271940.
- Hanahan D, Coussens LM. Accessories to the crime: functions of cells recruited to the tumor microenvironment. Cancer Cell. 2012;3:309–322. doi: 10.1016/j.ccr.2012.02.022. [DOI] [PubMed] [Google Scholar]
- Shojaei F, Ferrara N. Refractoriness to antivascular endothelial growth factor treatment: role of myeloid cells. Cancer Res. 2008;68:5501–5504. doi: 10.1158/0008-5472.CAN-08-0925. [DOI] [PubMed] [Google Scholar]
- Liang J, et al. Neutrophils promote the malignant glioma phenotype through S100A4. Clin Cancer Res Off J Am Assoc Cancer Res. 2014;20:187–198. doi: 10.1158/1078-0432.CCR-13-1279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Teramukai S, et al. Pretreatment neutrophil count as an independent prognostic factor in advanced non-small-cell lung cancer: an analysis of Japan Multinational Trial Organisation LC00-03. Eur. J. Cancer Oxf.Engl 1990. 2009;45:1950–1958. doi: 10.1016/j.ejca.2009.01.023. [DOI] [PubMed] [Google Scholar]
- Zhu Q, et al. The IL-6-STAT3 axis mediates a reciprocal crosstalk between cancer-derived mesenchymal stem cells and neutrophils to synergistically prompt gastric cancer progression. Cell Death Dis. 2014;5:1295. doi: 10.1038/cddis.2014.263. [DOI] [PMC free article] [PubMed] [Google Scholar]