

Abstract
Both human and insect societies depend on complex and highly coordinated infrastructure systems, such as communication networks, supply chains and transportation networks. Like human-designed infrastructure systems, those of social insects are regularly subject to disruptions such as natural disasters, blockages or breaks in the transportation network, fluctuations in supply and/or demand, outbreaks of disease and loss of individuals. Unlike human-designed systems, there is no deliberate planning or centralized control system; rather, individual insects make simple decisions based on local information. How do these highly decentralized, leaderless systems deal with disruption? What factors make a social insect system resilient, and which factors lead to its collapse? In this review, we bring together literature on resilience in three key social insect infrastructure systems: transportation networks, supply chains and communication networks. We describe how systems differentially invest in three pathways to resilience: resistance, redirection or reconstruction. We suggest that investment in particular resistance pathways is related to the severity and frequency of disturbance. In the final section, we lay out a prospectus for future research. Human infrastructure networks are rapidly becoming decentralized and interconnected; indeed, more like social insect infrastructures. Human infrastructure management might therefore learn from social insect researchers, who can in turn make use of the mature analytical and simulation tools developed for the study of human infrastructure resilience.
Keywords: resilience, social insects, infrastructure, transportation networks, communication networks
1. Introduction
As human societies continue to develop and expand, critical infrastructure systems such as telecommunication and food supply networks, power grids and transportation networks are becoming increasingly complex and interdependent (for examples in electrical infrastructure, see [1,2]). These complex infrastructure systems are not, however, restricted to human societies; social insects such as ants, bees, wasps and termites also use a variety of interconnected infrastructure systems (see the electronic supplementary material, table A). Colony infrastructure systems allow for transportation of resources, communication between colony members, and efficient storage and distribution of resources in response to colony needs [3]. Insect infrastructures are built and maintained in the absence of any centralized control. There is no leader and no master plan; rather, the emergent properties of the infrastructure network are dependent on local interactions between colony members (for a social wasp example, see [4,5]; for honeybee examples, see [6]). Like human-built infrastructure systems, insect systems must function in the face of a slew of disruptions including natural disasters, predator attacks, blockages or breaks in the transportation network, and outbreaks of disease. Unlike human-built infrastructure systems, however, insect infrastructures have been shaped by millions of years of natural selection and so may have evolved novel strategies for maintaining infrastructure function in the face of nature's assaults.
In this review, we bring together and examine literature that sheds light on resilience (or lack thereof) in insect infrastructure networks. Our review aims to answer three main questions: (i) What is resilience? (ii) Can we find examples of resilient infrastructures within the social insects? (iii) Which factors influence resilience in social insect systems? We will delve into these questions by focusing on the three the most well-studied social insect infrastructure systems: transportation networks, communication networks and supply chains. We will conclude by highlighting areas that require more research, proposing a framework for future study, and by examining the potential implications and lessons that can be learnt from the study of resilience in insect infrastructure systems.
2. Defining resilience
Interest in infrastructure resilience has surged in recent years (figure 1). Despite the focus on resilient infrastructure, there seems to be no consistent definition of ‘resilience’. Indeed, several reviews have attempted to define resilience [7,8]. For example, Sandia National Laboratories used the following definition of resilience when developing a framework for assessing the resilience of critical infrastructure systems in the USA: ‘Given the occurrence of a particular ‘disruptive event’ (or set of events), the resilience of a system to that event (or events) is the ability to reduce ‘efficiently’ both the magnitude and duration of the deviation from targeted ‘system performance’ levels' [9, p. 281]. Francis & Bekera [10, p. 93] list 25 slightly different definitions of resilience including the following: ‘Infrastructure resilience is the ability to reduce the magnitude and/or duration of disruptive events. The effectiveness of a resilient infrastructure or enterprise depends upon its ability to anticipate, absorb, adapt to, and/or rapidly recover from a potentially disruptive event’. Despite differences in the wording of each definition, it is clear that infrastructure resilience relates to the ability of an infrastructure system to maintain normal, pre-disturbance levels of performance when experiencing potential activity-disrupting disturbances. In ecology, a more dynamic definition of resilience is often used, such as ‘the capacity of a system to absorb disturbance and reorganize while undergoing change so as to still retain essentially the same function, structure, identity and feedback’ [11, p. 1]. In this review, we wish to combine these slightly different definitions into one that applies in an ecological setting, as well as a technological one. As such, we define resilience, and a resilient system, as one that returns to or exceeds its pre-disturbance level of performance following a perturbation. To achieve this, the system may change state, but the key is that the system remains functional (figure 2). Our definition focuses on the functionality of a system, rather than the specific topology or state of the system. ‘Functionality’ refers to the ability of a system to perform some task and will be experiment-specific; for example, we might investigate the ability of a colony to collect food (harvesting rate), the ability of a transportation system to maintain a high average rate of flow, or the ability of a colony to produce new colony members. Importantly, a system need not maintain its original state in order to be resilient so long as system functionality is preserved. For example, a colony which has been disturbed via flooding might move to a new nest site; the original colony is gone, but the colony's functionality has been restored. To be ‘resilient’ the system must return to a performance level that is equal to or exceeds performance pre-disturbance. Our definition allows for resilience metrics including the amount of time it takes to return to pre-disturbance functionality, the time until recovery begins and the magnitude of disturbance the system can recover from before ceasing to function (collapsing).
Figure 1.

Web of Science citation report for ‘infrastructure resilience’ over the past 20 years. (a) Citations in each year; (b) published items in each year.
Figure 2.
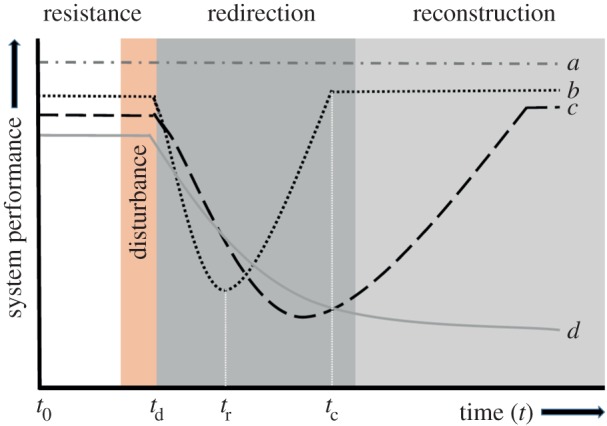
Potential responses of a system to a disturbance. System performance is an experiment-specific measure of system functionality (harvesting rate, brood production rate, traffic flow). Time t0 is the start of the experiment, pre-disturbance, td indicates the start of the disturbance and tr indicates the beginning of the recovery phase for one system and tc indicates the point at which recovery is complete since system performance has returned to pre-disturbance levels. (a) Shows a system that has invested in resistance and as a result does not experience a decrease in functionality after the disturbance. (b) Shows a system that is using redirection. Although there is an initial decrease in performance, this is rapidly mitigated by rerouting flows using existing infrastructure. (c) Shows a system that uses primarily reconstruction-based resilience strategies. Since reconstruction requires the construction of new infrastructure, it takes longer to recover pre-disturbance performance. (d) Shows a non-resilient system which does not recover pre-disturbance performance. (Online version in colour.)
Systems can achieve resilience in different ways. Here, we focus our discussion on three key pathways to resilience (figure 2). Systems may have an inherent capacity to ‘resist’ any perturbation or disturbance that alters the infrastructure from its functional state (resistance). Alternatively, a damaged system may have the capacity to redirect information, energy or resources around a perturbation to ensure continuing function (redirection). Third, a system may have the ability to repair and recover full or even improved functionality after the perturbation (reconstruction). These three forms of resilience are not mutually exclusive, and a system may display one or all responses to a disturbance. In the face of pathogens, for example, colonies can have multiple resilience strategies. They might resist infection through specialized behaviours such as using guards to prevent the entrance of infected individuals, or can simply avoid areas linked to high infection rates. Infected individuals might be socially excluded or killed, effectively redirecting work away from the disturbance (the infected individual). If infection rates become too high, the colony may resort to colony abandonment and reconstruction elsewhere [12].
While colonies may use multiple resistance strategies, we expect that species will differ in their relative investment to resistance, redirection and reconstruction. Consider, for example, the ways in which ant species deal with severe damage to their nests. Some species, like the meat ant (Iridomyrmex purpureus), invest heavily in building strong nest mounds with complex underground nest architecture. The nest itself is made from hardened soil which makes it very difficult to breach [13]. Colonies are therefore highly ‘resistant’ to attack or damage. By contrast, colonies of argentine ants, Linepithima humile, are opportunistic nesters and invest very little into nest construction. In the event of attack, ants rely on a coordinated evacuation response to rapidly relocate nest inhabitants to one of the many other nests that make up their nest network [14–16]; this is an example of redirection. In both species, colony functionality is restored following an attack; in the meat ant, the ability to resist an attack is key, whereas in argentine ants it is the ability to redirect resources around the disruption that leads to colony resilience.
3. Resilience in insect infrastructures
3.1. Transportation networks
In human systems, transportation networks are designed to allow the flow of resources between locations through physical structures, such as roads, railways, shipping lanes and flight paths. Modes of transport vary from animal powered (walking, cycling and carts) to machines (automobiles, ships and planes). Many social insects are also dependent upon transportation networks for the movement and distribution of resources such as food, water and foragers. Familiar examples include the trail networks of ants and the tunnel networks of termites. Insect transportation networks vary extensively in the cost of construction, from the relatively ‘cheap’ pheromone trail systems of trail-laying ants, to the cleared and maintained physical trails and tunnel networks of some ant and termite species (e.g. wood ants, Formica rufa, and meat ants, Iridomyrmex purpureus).
3.1.1. Topological resilience
In transportation networks, the capacity of a network to maintain connectivity as an increasing fraction of trails/tracks/roads are removed is characteristic of a resilient network [17]. The more connected a network is, the greater the resistance to damage; this is known as ‘robustness'. In graph theory, robustness is defined as ‘the capacity of a network to preserve connectivity as an increasing fraction of edges are removed’ [14]. For example, ‘complete networks', in which every point is connected to every other point (figure 3a), have a high degree of robustness as it would take significant damage to multiple trails before any single point was disconnected from the network. Returning to our definition of resilience, this high level of connectivity allows the network to remain functional through both resistance, as many connections must be damaged to isolate any one node, and, once damage has occurred, through redirection of traffic along undamaged trails. Such connectivity, however, results in networks which are expensive to build and maintain. Conversely, networks can minimize building costs by connecting points to one another using the least amount of trail (minimal spanning trees and Steiner minimum trees, herein MST and SMT, respectively; figure 3b). However, MSTs and SMTs are also minimally robust (figure 3). Thus, transportation networks face a key tradeoff between robustness and cost; this is known as the ‘transportation problem’ the formulation of which is attributed to eighteenth century mathematician Gaspard Monge [17].
Figure 3.

Examples of network connectivity modelled from meat ant nests: (a) complete networks are highly resilient and very costly to build; (b) a minimum spanning tree network, where the minimal number of connections are made between each node. Minimum spanning trees are cheap to build and maintain, but are vulnerable to disturbances; (c) real meat ant network. Red circles indicate nests, or nodes, and are connected by black lines, indicating trails or edges. Adapted from Cabanes et al. [18]. (Online version in colour.)
In laboratory settings, multi-nest (polydomous) colonies of argentine ants form network connections that are close to MST [19]. The topologies of such networks have very low robustness. They are unable to resist the initial damaging event, and since damage to any section of the trail results in the disconnection of at least one nest, they are unable to redirect resources to that nest by another route. However, the trail-laying behaviour of individual ants likely reconstructs the network cheaply and efficiently. In L. humile, individuals moving through the environment will continually deposit pheromones, even if they have not found food [20]. When a disturbance occurs and disrupts a trail, individual ants are capable of navigating their way to the nest location, laying a pheromone trail as they go. This allows the network to repair and recover quickly from disturbances [19,21]. Although the topology of the network lacks robustness in terms of the resistance to disturbances, the essentially ‘self-healing’ nature of the trail system makes it highly resilient to damage through reconstruction.
While argentine ants build topologically minimal networks which lack robustness, some ant species appear to invest more into robustness by increasing the connectivity of nests. Cook et al. [22] examined the topology of six different polydomous ant species using previously published colony maps. Colonies did not build minimal networks, as the number of connections between nests was higher than expected from MST. However, the number of connections was still relatively low, closer to an MST than to a maximally connected network [22]. Similar results were observed in polydomous meat ant colonies [18]. In these networks, we see increased investment in resistance and redirection. Interestingly, both wood ants and meat ants build physical trail networks which imply increased construction costs compared with the relatively cheap networks built by argentine ants. We suggest that the different resistance strategies employed by trail-laying and physical trail building ants might be due to differences in the cost of network construction. When the network is costly to build and maintain, it might be better to build topological robustness into the system, thus allowing for resistance and redirection. When, however, a network is relatively cheap to build, perhaps colonies would do well to invest primarily in reconstruction.
3.1.2. Traffic congestion
Following a boom in resources, transportation networks may suffer from congestion as more foragers, particularly among trail-laying ant species, use the same trails to gather food. As anyone who has ever been stuck in a traffic jam knows, traffic congestion can cripple a transportation network and severely decrease its efficiency. A resilient transportation network is one that can maintain its efficiency in the face of increased traffic. Many species of eusocial insect will self-organize lanes to optimize traffic flow, a behaviour aimed at resisting disturbances to the transport network. These insects include termites [23], some leaf cutter ants [24] and army ants [25]. The army ant, Eciton burchelli, forms up to three lanes on their trails during massive swarm raids which can involve up to 20 000 individuals. Lane formation is a self-organized process based on the way in which ants react to collisions. Collisions between outbound and inbound nest-mates causes outbound individuals to turn away from the contact point, while still moving forward. This leads outbound individuals to give a wider berth to inbound ants [25], ultimately resulting in a decreased collision rate and an increase in the rate of flow of ants to and from the resource.
Much of the work on insect transportation networks has focused either on network topology or on the management of traffic congestion in simplified environments. Combining these areas of research will lead to advances in our understanding of resilient transportation network design. Traffic, after all, is strongly influenced by the topology of the transportation network with certain configurations leading to an increased likelihood of congestion. Similarly, the topology of the network can provide alternative routes around congested sections of the network. There is also a paucity of data on the reconstruction of damaged networks, particularly for species that build physical trails. For example, how are networks repaired, and how is traffic redirected during the reconstruction process? Answering these questions in multiple species will give us a much better understanding of how resilience in insect transportation networks is maintained.
3.2. Communication networks
Communication networks are an integral part of social insect colonies. The communication network conveys information about the location and quality of food resources, ongoing enemy attacks and the general health of the colony. The communication mechanisms used by social insects are diverse, including physical interaction, pheromone use, auditory calling, vibrational signals and trophallaxis; indeed, a comprehensive review of social insect communication mechanisms is well beyond the scope of this review. In this section, we will focus on the characteristics of social insect communication systems which result in (or prevent) resilience.
3.2.1. Direct versus indirect communication
In communication systems, information transfer can be either direct (individual to individual) or indirect. In pre-literate human societies, for example, information transfer between a speaker and her audience was limited to those within hearing range (direct communication). The dawn of writing allowed for the use of large-scale externalized communication, where the information could persist long after the original writer had moved on (indirect communication). Social insect systems also use both direct and indirect information transfer. The dance language of honeybees, for example, involves successful foragers transmitting information directly to dance followers. Tandem running, where one ant (leader) leads other ants (recruits) to a food source, is another example of direct information transfer [26]. By contrast, trail-laying ant species leave chemical ‘messages' that can last long after the trail-layer is gone. The individual trail-laying ant is not required to physically interact with another individual for the information to be shared. The ant Euprenolepis procera, for example, uses stable, long-lasting pheromone trails to harvest mushrooms which tend to regrow in the same locations. The long-lasting pheromone ‘memory’ allows colonies to scout for mushrooms in areas where mushrooms had previously been found [27]. Volatile pheromones such as alarm pheromones are another example of indirect communication. Alarm pheromones alert nest-mates to a threat, in much the same way as a fire alarm alerts people in a building to a fire without all individuals having to observe the fire, or be told individually about the fire.
Indirect communication systems are resistant against the loss of individuals; even if the original releaser of an alarm pheromone is killed, the ‘message’ can still propagate to nearby individuals. Similarly, once an ant has laid a trail to a newly discovered food source, its particular presence is no longer required as all colony members now have access to the information about resource location. While indirect communication mechanisms are resistant to the loss of particular individuals, they may also be inflexible to changes in the environment, thus leading to the propagation of outdated information. It has been suggested that trail-laying ants have difficulty redirecting their workforce following a change in food quality, such as when a higher quality food source is discovered after a trail has already been established to a lower quality food source [28–30]. This inflexibility occurs because the new trail cannot compete with the older, continually re-enforced trail [28–30]. By contrast, direct communication allows individuals to be led to higher quality food sources in spite of established recruitment to the lower quality food source [28]. In this view, trail-laying species tend to have high resistance to individual loss, but suffer from an inability to redirect foragers following disruption; the converse would be true for species that use direct communication mechanisms. Things are not so simple, however; several recent studies have shown that trail-laying ants are nowhere near as inflexible as previously thought, and that some level of modularity of pheromone signals exists [27,31–34]. In at least one species, the ability to redirect foragers following a change in quality appears to be due to the use of two distinct pheromones: one short acting pheromone which elicits a very strong recruitment response and a long acting pheromone which elicits a much weaker response [32]. In another species, Paratrechina longicornis, it was demonstrated that the specificity of some pheromones was reduced, and four distinct glands could be used for communication. This allows flexibility in the signal message, as it can be graded in terms of strength, persistence of the pheromone, and may be applied in different situations, such as for defence, attack or a positive signal for food [34]. Thus, colonies using indirect communication may have mechanisms that allow them to mitigate inflexibility while still being relatively resistant to the loss of individuals.
Within a colony, the use of direct or indirect communication is often context-dependent, and most species use a mixture of both communication types. For example, if a honeybee colony is under attack, bees will rush to defend the hive [35,36]. An alarm pheromone is released by alerted and attacking workers, guiding other bees to the site of danger. In this instance, there is no direct communication between workers; the pheromone indirectly communicates the message of danger to other workers. The loss of one responder does not diminish the signal and thus allows continued response until the threat has been neutralized. When communicating the location of a new food source, however, honeybees engage in direct communication through the waggle dance [37]. During the dance, they convey directions to only those bees following the dance. In this instance, the loss of a successful forager could mean the loss of a lucrative food source, a relatively less resilient approach to resource gathering than indirect communication through pheromone trails.
3.2.2. Topology of communication networks
The topology of a communication network has strong effects on how information flows through the system. Communication networks can be homogeneous, where each individual in the network is equally connected to other individuals in the network, or they can be heterogeneous, with a few highly connected individuals being responsible for the majority of information dissemination. Homogeneous networks may be more resilient to the loss of individuals, as all individuals are equally privy to all information, whereas heterogeneous networks may be more vulnerable to disturbance with the loss of a key individual potentially leading to the loss of important information. Honeybee colonies, for example, have a heterogeneous communication network with regards to foraging information, since a few individuals (dancing foragers) are responsible for the majority of information transmission [5]. The loss of a dancing forager results in the loss of information regarding pollen or nectar locations.
Eusocial wasp colonies, Ropalidia marginata, on the other hand, are characterized by network homogeneity in small colonies [4]. Such a homogeneous network makes small colony social networks resilient [4]. As the colony grows (up to 40 individuals), it becomes less likely that individuals will communicate directly with all other members, and the topology becomes more heterogeneous. Information transfer becomes dependent on a few key individuals, leading to the same vulnerability within the network as described for honeybees if those individuals are removed [3,4]. The loss of key individuals responsible for information transfer and memory of specific resource locations can lead to a drop in resource supply, thus illustrating the potential for localized disruptions to cascade into related infrastructure systems. Interestingly, loss of individuals leads to increased connectivity with remaining individuals, and the ability to quickly establish new connections. This suggests that the colony has mechanisms to quickly reconstruct the communication network following the loss of individuals.
3.3. Supply chains
In humans, the term ‘supply chain’ includes all the steps and processes necessary to move a product from a supplier to a consumer. Social insects face the daunting task of collecting enough food to meet the nutritional demands of the colony; this results in surprisingly complex supply chains which involve not only food collection, but also sourcing, transportation, processing and storage steps. In leaf cutter ants, for example, leaves are sourced from a variety of plants, transported back to the nest along a transportation network, and converted into a nutrient-rich food for fungal gardens which are themselves tended by workers [38–40]. The production of honey by honeybees involves hundreds of foragers sourcing nectar from a variety of different plants, nectar processors who make the honey through a process of regurgitation and evaporation, and cell builders who construct storage space for the honey [41,42].
As resources are typically variable throughout time and space, colonies need to have mechanisms that allow them to maintain a steady supply of resources during boom and bust periods [43,44]. Resources can appear, disappear or change quality. Competition with other colonies or species can change the relative risks associated with particular resources, and natural disasters such as flooding can cut off access to resources.
3.3.1. Warehousing resources
One way to remain resilient in the face of a variable resource supply is to store excess resources in warehouses (‘warehousing’). Honeybees exhibit a form of warehousing, where extra pollen and nectar are stored within wax combs. Honeybee colonies increase storage of resources in late summer, both within the nest and within their own bodies [45,46]. In some areas, honeybees decrease or cease breeding altogether over the winter, preventing an increase in demand through an increase in brood. The remaining bees rely on warehoused pollen reserves to sustain themselves until the spring [45,47].
While building warehouses is a good way to store food against periods of scarcity, the construction of storage space can be costly. Thus, insects must balance the costs of constructing new storage space against the benefit of increased stores and the projected need for storage space in the future. In honeybees, construction of new storage space (comb) occurs only when two conditions are met: (i) the colony is experiencing a high level of incoming nectar and (ii) when a threshold number of existing nectar combs have already been filled [48]. This threshold is typically low, with enough empty comb often present to accommodate several kilograms of honey. By using two criteria, honeybees minimize the chance that excess honeycomb will be built. By setting a low threshold, the bees further ensure that storage space will be available in the event of a sudden increase in supply [48].
There are other examples of eusocial insects warehousing resources. For example, Mexican honey wasps, Brachygastra mellifica, store excess nectar in comb as honey [49]. Some ant species have developed specialized workers, called repletes, with extended abdomens in which they store resources, such as nectar, honey dew and/or water [50–52]. The presence of repletes has been posited as a contributing factor to the success of ant species in arid and semi-arid environments, which without adequate storage facilities would not survive a harsh environment characterized by periods of scarcity and erratic resource supply [52]. Repletes are often spread across a number of galleries, a potentially resilient approach to protect resource supply from disturbances that damage galleries, or from the spread of toxins or disease [53].
Another interesting example of warehousing in eusocial insects is seen in Crematogaster torosa which has nests that house only workers and no brood, but are connected to the polydomous nest network [54]. These ‘outstations' are highly defensible and serve as a storehouse for specialized foods such as ant jerky (dried insect tissue). Ant jerky is only suitable for larvae and not adult workers, yet only workers are present in the outstations. Outstations apparently act as warehouses for the colony, allowing food to be spread over a wide spatial area, thus reducing the risk of wide scale food loss should one or a few nests be damaged. Further, defensible outstations may act as back up nests, as seen in Pheidole desertorum, where the colony can relocate to outstations if attacked by another species. Lastly, outstations are often located close to resources that may vary in quality throughout the year. By keeping these stations manned, workers can quickly and efficiently gather resources when the resource increases in quality. Resources can then be moved to other areas of the polydomous network as required [54].
3.3.2. Supply chain topology
In human supply chains, it has long been recognized that the ability to source from multiple suppliers results in supply chain resilience [55]. In 2000, a fire at an electronics plant destroyed millions of microchips used in the design of mobile phones. The electronics plant supplied microchips to two companies: Nokia and Ericsson. Following the fire, Ericsson, which only sourced microchips from the single factory, lost 400 million dollars' worth of sales as production of their phone was severely impacted by an inability to source microchips. Nokia, on the other hand, sourced microchips from several geographically distributed suppliers and so simply switched to a different factory. Nokia's strategy achieved resilience not simply by having multiple warehouses, but also by ensuring that they were spread over multiple geographical locations [55,56]. Ant colonies, like human businesses, can also make use of multiple, geographically spread warehouses. The outstations described in the previous section are an excellent example of multiple warehousing. In ants, supply chain resilience may partially explain why some colonies build and maintain multiple nests (polydomy) despite the additional building and maintenance costs. By spreading colonies over a wider area, polydomy might allow colonies to take advantage of more spread out resources [57]. These storage locations may then be treated as food sources for foragers inhabiting one of the other multiple nests, as seen in Iridomyrmex sanguineus, facilitating resource relocation between nests [58].
Distribution networks within ant colonies also display resilience in response to famine events and the spread of toxins. After a famine event, Temnothorax albipennis ants are capable of distributing resources among nest-mates much faster than under normal conditions, resulting in 95% of the colony being fed within 30 min of the first forager returning with food [53]. Rapid food distribution is facilitated by individual behaviour such that individuals involved in within nest tasks, such as brood care, move towards the entrance of the nest to meet incoming foragers laden with food [53]. Further, food laden returning foragers will move further into the nest to distribute food after a famine event.
3.3.3. Changes in resource supply and demand
To function efficiently, colonies must ensure they are able to deal with fluctuations in both supply and demand. Ant colonies are able to track fluctuations in resource quality by rapidly redistributing foragers. Honeybees track changes in resource supply and demand by monitoring storage rates within the wax comb of their hives. As the proportion of empty comb increases, foraging activity for nectar and pollen collection also increases [59,60]. Pollen foragers directly assess the amount of pollen stored in the comb [61–64], whereas nectar foragers assess storage levels through interactions with nest-mates [61]. This is an intriguing behavioural difference: why do nectar foragers rely on indirect information about colony needs, while pollen foragers use direct sources of information? How does this affect the bee's ability to meet changes in supply and demand?
In social insects, adults and larvae have different nutritional requirements such that adults depend primarily on carbohydrates while the developing larvae require large amounts of protein. This sets up the interesting situation where the individuals involved in resource sourcing, collection and transportation (adults) must consider the requirements of individuals (the larvae) with nutritional needs that differ strongly from their own. This adds an additional layer of complexity when it comes to meeting colony demand for resources. In honeybees, protein foraging is stimulated by pheromones released by the larvae; in combination with assessments of the storage comb, brood pheromones allow pollen foragers to accurately assess the colony-level demand for protein [62].
3.4. Features that contribute to colony-level resilience
3.4.1. Task switching and behavioural flexibility
Division of labour, where individuals specialize in particular tasks, is a common feature of social insect societies. While division of labour has some clear benefits—namely, specialized individuals are much more efficient than un-specialized workers—it comes with a potential cost should specialized individuals be lost. The solution to this potential issue is to have flexible workers who are capable of switching tasks when needed. For example, when the demand for pollen increases, honeybees previously employed in nectar foraging switch to pollen foraging until the demand is met and pollen stores have been refilled [61]. Task switching is possible even when colonies have castes physically specialized for particular tasks. However, castes can differ in their ability or propensity to switch tasks. In many Pheidole ant species, individuals are separated into two distinct castes: majors, who are larger and have disproportionately large heads, and the much smaller minors who are primarily responsible for resource collection. When minors are removed from the colony, major workers expand their behavioural repertoire to act as substitutes for the missing minors, attaining 75% or more of the minor workers' activity. The change in behavioural repertoire occurs very rapidly and is thus unlikely to be dependent on hormonal change or learning experience [65]. Minors, however, do not expand their repertoires when majors are removed, suggesting that they are behaviourally inflexible. The ability to compensate for the loss of specialized colony members makes colonies resilient against unexpected losses; indeed, in the Pheidole, majors were capable of ‘rescuing’ de-populated colonies by taking on all the roles normally performed by minors [65]. The relative inflexibility of minors is interesting, as it suggest that there are constraints which prevent some castes from taking on new roles.
The ability of some workers to switch tasks without requiring a change in development or learning experience means that colonies can respond quickly to changes in the work force, potentially allowing them to prevent a minor disturbance from having a major impact. A loss of pollen foragers can lead to a loss of information regarding resource locations as well as a decrease in pollen collection and storage [66]. However, the ability of all individual bees to switch tasks rapidly (for example, from a nurse bee to a forager) prevents a potential failure cascade where the decrease in food resources causes a slowdown and eventual cessation of reproduction, which in turn means fewer foragers and even less resource collection. Task switching, however, has its limits. Hastening behavioural development in honeybees can lead to colony collapse because precocious foragers lack experience and are inefficient [67]. The presence of younger foragers leads to a feedback loop, with the foraging force becoming progressively younger over time as the current foragers die prematurely due to their inefficiency and lack of experience. This leads to a decrease in food levels, which increases the demand for early foragers, increasing colony death rate and resulting in the eventual collapse of the colony [67]. Thus, the resilience of the system is dependent on the scale of the disturbance and the type of response. Small losses in the forager force can be compensated for through the switching of tasks among other foragers and younger bees. However, in the face of large forager losses, colonies face a failure cascade ultimately leading to the death of the colony.
3.4.2. Redundant individuals
Humans have long anthropomorphized social insects as being hardworking; hence such phrases as ‘busy as a bee’. In reality, however, social insect colonies often maintain a large number of seemingly ‘lazy’ individuals that do not work. In a range of social insects, ‘inactivity’ occupies as much as 50% of colony task time [68]. The existence of lazy workers has prompted a lot of discussion among social insect scientists since producing workers who apparently serve no useful function is a tremendous energy cost to the colony. It has been suggested, although rarely tested, that redundant individuals may act as a reserve work force allowing the colony to respond rapidly to disturbance [69–71]. In this view, disturbance events, such as predator attack or the discovery of a high-quality resource, trigger idle workers into becoming active [71]. However, the reserve workforce hypothesis has not been experimentally validated, so at present the explanation for lazy workers remains unclear. In particular, we do not know much about the economics of laziness: at what point does it become adaptive to maintain a reserve workforce? We might expect that colonies living in highly variable environments might have higher numbers of backup workers than those living in relatively stable environments; this idea remains to be tested. Interestingly, large ant colonies maintain a larger proportion and absolute number of redundant workers, suggesting that they are more resilient to disturbance than are smaller colonies [69,70].
3.4.3. Memory
When systems have memory, they can return to their original state quickly. The potential impact of memory on resilience seems clear; systems that have more memory should be more resilient than those that lack memory. In a series of field experiments, Granovskiy et al. [72] found that honeybee colonies with communication disabled (i.e. information gathered through the waggle dance was halted) were still able to track changes in resource quality by consistently allocating a greater number of foragers to rewarding resources. Using a simulation model, they showed that this resilience was due to the memory of individual bees which remember and monitor previously rewarding food sources. The fact that the memory of rewarding foods is preserved within the memories of individual bees allows for colony-level resilience even after the communication systems are rendered non-functional.
Memory need not be encoded in individual brains. Pheromone trails are an example of an externalized memory system. By externalizing the colony memory, no one individual is key for access to resources. All individuals are equipped with sensors in order to detect pheromone trails, and all are capable of following them, collecting a resource, and returning without any requirement for their own memories and without any direction from other nest-mates. Both systems of encoding memory—externalized and internalized—provide resilience, albeit in different ways. If pheromone trails disappear or if worker numbers fall below the threshold needed to maintain the trails, then information about the location of foods is irrevocably lost. For example, when a threshold number of pharaoh ants are removed from the population, trails are no longer maintained and foraging becomes disorganized [73]. Conversely, individuals that have internalized memories can continue foraging even if all other foragers have been removed, since each individual is fully capable of independently remembering food sources. However, these systems are more vulnerable to the loss of key individuals who have within them the memory of particular food resources. In house hunting honeybees, for example, less than 5% of bees know the location of their new nest site; all other bees rely on these 5% to guide them safely to the new nest. The loss of only a few of these 5% would be catastrophic to the swarm which would lose knowledge of the new nest site [74,75].
In Temnothorax ants, memory is encoded in the spatial arrangement of individuals within the nest [76,77]. Particular tasks within the colony are spatially segregated, and individual ants have ‘spatial fidelity zones', task-associated spatial regions of the nest to which they will always return. If the colony is forced to re-locate due to the destruction of the nest, individuals will return to the same relative positions within the new nest. As a consequence, division of labour is maintained, since each spatial zone is also associated with a particular task. This unique form of memory allows colonies to rapidly reconstruct their infrastructure systems following a massive disturbance (the destruction of the nest).
Perhaps most interesting are those systems that use both externalized and individual-based memory. Some ant species are capable of overriding pheromone information with their own memories. The ant Lasius niger, for example, will not follow a pheromone trail if the individual ant remembers better foraging locations. Such a system is inherently resilient because information is stored at two levels. The loss of knowledgeable individuals is less damaging because the information they have is externalized in the trail. At the same time, individuals can remember and navigate to food sources in the event the trail network is compromised [78].
The length of memory—whether internal or external—could have an impact on system resilience. In some ants, pheromones range from short-lived chemicals that evaporate within hours if not reinforced, to long-term trail pheromones that remain active days or weeks after being laid [79–81]. In honeybees, the memory of rewarding food sources diminishes over a period of 10 days, after which the food source is forgotten. Long-term memory has the advantage of preserving a ‘blueprint’ which can be relied upon in the event of environmental change. On the other hand, long-term memory can also mean that systems become ‘stuck’ on suboptimal solutions, similar to the situation discussed for species using indirect communication mechanisms. Given the limitations of both short- and long-term memory retention, it is perhaps not surprising to note that many, if not most, species use several layers of memory, each with a different retention period. Lasius niger, for example, retains memory at the individual level (of an unknown duration) as well as memory encoded in trail pheromones; Pheidole megacephala uses both a short and a long acting trail pheromone.
3.4.4. Decentralized control
Decentralization is arguably one of the greatest sources of resilience in social insect colonies. In social insect societies, there is no leader and no blueprint; each individual makes decisions based primarily on local information. Even the queen, who is responsible for reproduction, is replaceable. In the event of queen death, honeybees, for example, can rear emergency queens, and do so very quickly [82]. Decentralization contributes to resilience by ensuring that no individual is irreplaceable; it therefore allows colonies to be resistant to the loss of individuals. Moreover, damage to the system can be dealt with as it happens, without need for time-consuming communications with managers. We suggest that decentralization also allows for faster redirection, since no control centre need monitor damage and select alternative routes; rather, these processes are an emergent property of the system. While decentralization is commonly cited as a source of resilience in social insect colonies, no studies have empirically tested this assumption. Although all social insect systems are to some extent decentralized, they do vary in their degree of decentralization and it would be interesting to determine the relationship, if any, between centralization and resilience.
3.5. Resilience in social insects: a framework and prospectus
In this review, we have pulled together literature on insect infrastructure resilience from a variety of sources. Very few of these studies set out to study resilience per se, and there is currently no general framework for studying resilience in social insects. We suggest that focusing on the three pathways to resilience (resistance, redirection and reconstruction) provides a common way of describing responses in a variety of systems. We also advocate an economic approach that expressly considers the costs and benefits of different resilience strategies.
3.5.1. Comparisons between systems
Given the incredible variety of social insect systems, it is likely that different systems have evolved different solutions to disruption. This diversity of solutions makes social insects a potentially powerful testbed for examining theoretical ideas about resilience in infrastructure systems. Conducting large-scale manipulations on human infrastructure networks is intractable and often unethical; as a consequence, we must rely on case studies or simulations in order to test our understanding of how complex infrastructure systems function under disruption. Social insects, however, are amenable to direct experimentation. We do not suggest that social insect systems are proxies for human infrastructures systems; there are clear ways in which systems differ on a fundamental level. However, by comparing numerous systems, we can begin to understand the general features that confer resilience to particular kinds and magnitudes of disruptions. Moreover, we can begin to understand the environmental factors that lead to the evolution of particular kinds of resilience. Some work has already occurred in this direction. For example, Linksvayer & Janssen [83] reviewed the response of ants to disturbances ranging from predation and pathogens to forest fire and climate change. They identify colony size as a key factor allowing resilient responses in ant colonies.
We also suggest that more research should be aimed at field studies of wild colonies. While field research presents many logistical challenges, we suspect that the relatively simplistic laboratory environment does not give us sufficient insight into the response of colonies to disturbances in the wild. We do not know much about the regular and irregular disturbance schedules that colonies experience, nor do we fully understand the mechanisms species have evolved to deal with different kinds of disturbance. If studies on field colonies are infeasible (as is the case when we need to observe in-nest interactions), then care should be taken to design a laboratory environment that mimics at least some of the complexity a colony would experience in the wild. Hybrid approaches, where observation nests are placed in the field, may provide a compromise between the control of laboratory studies and the environmental complexity of field studies [68].
3.5.2. The economics of resilience
We suggest an economics-based approach to comparing the use of resilience strategies in social insect colonies. The value of a resistance strategy probably depends on the severity and frequency of disruption, as well as the amount of functionality lost while the system recovers. Importantly, the different resilience pathways involve investments at different time points (figure 2). Resistance, for example, typically requires an investment before a perturbation occurs. This will result in wasted effort if the disruption does not occur; we therefore predict that resistance is best suited to situations in which disruptions are common and/or predictable. Resistance also prevents a disturbance from having an impact on system performance, again making it well suited for frequent or predictable disturbances. In contrast to resistance, the costs of reconstruction are paid only after the disturbance has occurred; there are few, if any, up-front costs. We expect that in most situations reconstruction will take the longest of the three pathways to restore system functionality because new infrastructure needs to be created. Reconstruction is therefore best suited for catastrophic, low probability disturbances such as nest or trail destruction. Redirection is intermediate between resistance and reconstruction as it occurs once a disruption has started and system performance has already been impacted; there is therefore a cost in the form of lost performance. However, recovery time is faster than would be expected from reconstruction as redirection uses existing infrastructure rather than creating new infrastructure. Box 1 provides a case study examining the economics of resilience in ant transportation networks.
Box 1. The economics of resilience in insect transportation networks: case study.
Understanding how evolution has shaped resistance strategies requires a comparative approach of many different species and different types of disruption (e.g. trail obstruction, nest destruction, change in traffic conditions, loss of foragers, flooding). Below is a case study which describes how thinking of the economics of resilience can improve our understanding of insect behaviour.
Building resilient transportation networks. Many species of ant build transportation networks to move food, brood and other resources from one place to another. Trails include pheromone-based trails (e.g. Linepithima humile), underground tunnel systems (e.g. Solenopsis invicta), walled trenches (Dorylus wilverthi and D. nigricans) and physical trails cleared of vegetation and other obstructions (Iridomyrmex purpureus and Atta spp.). When damaged or obstructed, pheromone-based trail systems are easily repaired: individuals need only deposit new pheromone. In contrast, physical trail networks or tunnels need to be rebuilt, a process that is likely to be time consuming and energetically expensive. The delay in repair also means greater loss of functionality since the trail may be unusable during the repair process. Assuming that physical trails and tunnels cost more to build and take longer to fix, we predict that damage to physical trail systems or tunnels will have a greater impact on functionality than damage to pheromone-based trail systems. Functionality in most cases will mean the rate at which food is returned to the nest from a food source (harvesting rate), but could also be quantified in terms of traffic volumes and traffic speeds. The prediction that the loss of functionality will be more severe for pheromone-based trail systems leads to our second, more interesting prediction: species using physical trail networks will invest more into resistance strategies by building topologically robust trail systems, while colonies using pheromone-based trail systems will invest more into reconstruction strategies. For easy-to-fix pheromone-based trail systems, there is little need for topological robustness since reconstruction is a quick, cheap process. In contrast, tunnels or physical trails take considerable time and energy to reconstruct; thus, we expect greater investments in resisting damage.
Testing our hypothesis requires a comparison of the topologies of tunnel systems, physical trails and pheromone trails. Cook et al. [22] used published maps of inter-nest networks of six ant species as the basis for calculating network robustness. While the dataset is an important step in the right direction, the small sample size for pheromone trail layers (one species) makes it difficult to draw conclusions about investments in resilience strategies. As more researchers become interested in insect transportation networks we will hopefully see an increase in the number of published network maps. This will allow for a better understanding of how different species deal with resilience in their transportation systems.
3.5.3. Learning from human infrastructures
In contrast to the study of resilience in biological systems, supply chain management (e.g. [84]), engineering (e.g. [85]), ecology (e.g. [7]) and socioeconomics (e.g. [86,87]) all have a detailed literature on resilience including a suite of well-developed modelling and analytical tools [88–91]. Our understanding of resilience in social insect systems could be greatly advanced by borrowing some of the ideas, tools and concepts from resilience work in other disciplines. For example, phase transitions, characterized by a sudden change from one state to another, are an important concept from the study of resilience in complex systems [92]. Phase transitions have been observed in the foraging behaviour of ants, where systems transition abruptly from functional to non-functional [73]. Colonies of leaf cutter ants also undergo a phase transition from unstable colony growth when colonies are small, to stable growth after they reach a threshold number of individuals [93]. Are phase transitions a common feature of insect infrastructures? Or do some infrastructures experience a more gradual decline in functionality? For example, one might predict that in systems where memory resides predominantly in individuals, the removal of these individuals would lead to a gradual decline in colony functionality, rather than the abrupt phase transition observed in pheromone-laying species. Future research should focus on understanding the factors that lead to phase transitions rather than gradual declines.
In socioeconomic systems, the idea that systems can adapt to changes is key to understanding resilience [87]. Following the devastating Hurricane Katrina, the City of New Orleans undertook a major plan to strengthen and increase levees, and to design better evacuation and shelter plans [94]. As a result, New Orleans is likely more resilient against similar events than it was before. We assume that social insects can evolve to be resilient against perturbations over many generations, but can an individual colony adapt its infrastructure systems following a perturbation? There is some evidence that ant colonies can adapt their nest architecture following disturbance. Post flooding, workers of five South American ant species built earthen levees around their nest entrances; these levees prevent experimental flood waters from entering the nest [95]. During an extremely hot summer, Zakharov & Zakharov [96] observed Formica ants modifying their nests by building dense covers over aphid colonies and trails; these covers were thought to serve as thermal insulation [96]. These studies raise the intriguing possibility that ants may have techniques for adapting infrastructure systems following disturbances. Future work would do well to investigate adaptation in other insect infrastructure systems. For example, we know that some ant colonies build networks which balance robustness and cost; could this balance be shifted towards increased robustness (at the expense of cost) if a colony was exposed to frequent disruption?
Another important concept from human infrastructures is the concept of cascading failures. A cascading failure occurs when the failure of a single component causes connected components to fail. Cascading failures have been implicated in several large-scale blackouts, where a point failure in the grid (i.e. damaged power transmission line) causes a rapidly propagating wave of failure, ultimately blacking out large areas [97]. Failure cascades have also been described in metabolic networks, where the removal of a single enzyme can disrupt all subsequent reactions [98]. Cascading failures have not explicitly been reported in the social insect literature; however, several examples appear to bear the hallmarks of failure cascades. Recent work on colony collapse disorder in honeybees seems to suggest that these colonies are destroyed by a cascading failure event. The loss of experienced foragers results in the production of inexperienced and inefficient precocial foragers who become ‘over loaded’, stimulating the production of even more precocial foragers [67,99]. Do other social insect systems similarly experience cascading failures? Do some systems have in-built mechanisms to arrest failure cascades?
The increasing interdependency of human infrastructure networks has been cited as a serious threat. Interdependency means that a single failure in one network can quickly propagate across multiple networks, greatly magnifying the extent of the damage. The Internet, for example, relies on the power grid for power while the power grid is itself controlled via the Internet. Damage to either network can result in the failure of its co-dependent networks. Network interdependency is also a potential problem for social insects. In ants, the transportation network used to distribute workers and brood between nests is also used to collect and distribute food; damage to the transportation network thus impacts the supply chain. Very few insect studies consider multiple infrastructure systems; researchers tend to focus on a single infrastructure. Future research would do well to investigate how perturbations in one infrastructure influence the performance of interconnected infrastructures. Interestingly, honeybees can continue to forage effectively despite disruption of their communication infrastructure [72]; this decoupling of normally dependent infrastructures may provide clues as to how humans can build resilient interconnected infrastructure systems.
3.5.4. Learning from nature
In this review, we have highlighted the many similarities between human and insect infrastructures. However, human and insect infrastructure networks differ in fundamental ways. Human infrastructure networks have traditionally been managed in a centralized, hierarchical manner. In times of infrastructure failure, the usual human response is ‘crisis management’, where a relatively small group of people work together to organize an emergency response. Similarly, the ‘resistance’ phase of infrastructure resilience usually involves pre-disaster planning, again largely developed by a small, centralized group of people. Not only are the people involved in planning disaster responses possessed of a large and sophisticated cognitive system but they also make use of computers, simulation modelling and extensive data from previous crises. The centralized, ‘top-down’ approach to managing resilience differs dramatically from the decentralized approach seen in social insects, where resilience emerges as a result of the collective behaviours of individuals which lack the cognitive sophistication of humans. Given these significant differences, can we learn anything about infrastructure resilience from social insects, or is the analogy we have presented too superficial to allow meaningful cross over between disciplines? This question is difficult to answer at present because there have not been enough studies focused on the mechanisms of resilience in social insects; indeed, a major goal of this review is to stimulate research in this direction. Nevertheless, there are reasons to believe that social insects might provide insight into new ways of managing infrastructure networks.
While early infrastructure systems were largely centralized, modern infrastructures such as power grids are becoming increasingly decentralized and self-organized (e.g. [100,101]). For example, power grids are decentralizing due to the advent of ‘smart’ power grids, where individual houses equipped with solar panels can now contribute energy back to the grid [102]. The rise of the ‘Internet of things', where electronics and appliances contain sensors and can collect and exchange data with one another, is another example of how our infrastructure systems are becoming more self-organized and less centralized. While autonomous ‘smart systems' have the potential to revolutionize sustainable energy, they also pose unique challenges. How do you build a semi-autonomous grid that can deal with rapid, unexpected changes in both supply and demand? How do you build self-healing systems which can respond quickly to damage? How do you prevent the propagation of incorrect sensor data from causing cascading errors? Infrastructure failures can now happen far too quickly (milliseconds) and spread far too fast for any individual human to mitigate; we are therefore in need of systems that can implement mitigation measures rapidly and autonomously. We argue that social insects provide a rich toolbox of potential techniques for managing the increasingly decentralized and interconnected infrastructures of modern society. By comparing multiple systems, we can also develop a clearer understanding of how system-level features enhance or detract from resilience to different kinds of disturbances.
4. Conclusion
Thinking of insect systems as a set of interrelated ‘infrastructures', as we have done here, is a good way to advance our understanding of resilience as it allows us to borrow some of the conceptual tools currently used to discuss resilience in human infrastructure systems. In the electronic supplementary material, table B, we provide a list of outstanding questions we feel would be excellent targets for research; we hope these will serve as a jumping off point for researchers interested in investigating resilience in social insects. As demonstrated in our case study, there is room for significant targeted research to examine resilience in eusocial insect networks. We also suggest that considering the different ways systems will respond (resistance, redistribution and reconstruction) provides a framework for comparisons between species, ultimately allowing us to investigate the ecological and physical forces that may shape the evolution of particular resilience strategies. Studying insect infrastructures from the perspective of system resilience has the potential to lead us to insights that may one day help design better human-built infrastructures.
Supplementary Material
Supplementary Material
Authors' contributions
T.L. conceived of the review and revised the manuscript; E.J.T.M. drafted the manuscript. Both authors gave final approval for publication.
Competing interests
We declare we have no competing interests.
Funding
The Branco Weiss Society in Science Fellowship (T.L.) and an Australian Research Council Discovery grant no. DP140103643 to T.L.
References
- 1.Holmgren Å, Molin S. 2006. Using disturbance data to assess vulnerability of electric power delivery systems. J. Infrastruct. Syst. 12, 243–251. ( 10.1061/(ASCE)1076-0342(2006)12:4(243)) [DOI] [Google Scholar]
- 2.van der Vleuten E, Lagendijk V. 2010. Interpreting transnational infrastructure vulnerability: European blackout and the historical dynamics of transnational electricity governance. Energy Policy 38, 2053–2062. ( 10.1016/j.enpol.2009.11.030) [DOI] [Google Scholar]
- 3.Fewell JH. 2003. Social insect networks. Science 301, 1867–1870. ( 10.1126/science.1088945) [DOI] [PubMed] [Google Scholar]
- 4.Naug D. 2009. Structure and resilience of the social network in an insect colony as a function of colony size. Behav. Ecol. Sociobiol. 63, 1023–1028. ( 10.1007/s00265-009-0721-x) [DOI] [Google Scholar]
- 5.Naug D. 2008. Structure of the social network and its influence on transmission dynamics in a honeybee colony. Behav. Ecol. Sociobiol. 62, 1719–1725. ( 10.1007/s00265-008-0600-x) [DOI] [Google Scholar]
- 6.Breed M, Williams D, Queral A. 2002. Demand for task performance and workforce replacement: undertakers in honeybee, Apis mellifera, colonies. J. Insect Behav. 15, 319–329. ( 10.1023/A:1016261008322) [DOI] [Google Scholar]
- 7.Gunderson LH. 2000. Ecological resilience—in theory and application. Annu. Rev. Ecol. Syst. 31, 425–439. ( 10.1146/annurev.ecolsys.31.1.425) [DOI] [Google Scholar]
- 8.Holling CS. 1996. Engineering resilience versus ecological resilience. Washington, DC: The National Academies Press. [Google Scholar]
- 9.Vugrin ED, Warren DE, Ehlen MA. 2011. A resilience assessment framework for infrastructure and economic systems: quantitative and qualitative resilience analysis of petrochemical supply chains to a hurricane. Process Safety Progress 30, 280–290. ( 10.1002/prs.10437) [DOI] [Google Scholar]
- 10.Francis R, Bekera B. 2014. A metric and frameworks for resilience analysis of engineered and infrastructure systems. Reliab. Eng. Syst. Safety 121, 90–103. ( 10.1016/j.ress.2013.07.004) [DOI] [Google Scholar]
- 11.Walker B, Holling CS, Carpenter SR, Kinzig A. 2004. Resilience, adaptability and transformability in social–ecological systems. Ecol. Soc. 9, 5. [Google Scholar]
- 12.Cremer S, Armitage SA, Schmid-Hempel P. 2007. Social immunity. Curr. Biol. 17, R693–R702. ( 10.1016/j.cub.2007.06.008) [DOI] [PubMed] [Google Scholar]
- 13.Cowan J, Humphreys G, Mitchell P, Murphy C. 1985. An assessment of pedoturbation by two species of mound-building ants, Camponotus intrepidus (Kirby) and Iridomyrmex purpureus (F. Smith). Soil Res. 23, 95–107. ( 10.1071/SR9850095) [DOI] [Google Scholar]
- 14.Holway DA. 1995. Distribution of the Argentine ant (Linepithema humile) in northern California. Conserv. Biol. 9, 1634–1637. ( 10.1046/j.1523-1739.1995.09061634.x) [DOI] [Google Scholar]
- 15.Heller NE, Gordon DM. 2006. Seasonal spatial dynamics and causes of nest movement in colonies of the invasive Argentine ant (Linepithema humile). Ecol. Entomol. 31, 499–510. ( 10.1111/j.1365-2311.2006.00806.x) [DOI] [Google Scholar]
- 16.McGlynn TP. 2012. The ecology of nest movement in social insects. Annu. Rev. Entomol. 57, 291–308. ( 10.1146/annurev-ento-120710-100708) [DOI] [PubMed] [Google Scholar]
- 17.Perna A, Latty T. 2014. Animal transportation networks. J. R. Soc. Interface. 11, 20140334 ( 10.1098/rsif.2014.0334) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cabanes G, van Wilgenburg E, Beekman M, Latty T. 2014. Ants build transportation networks that optimize cost and efficiency at the expense of robustness. Behav. Ecol. 26, 223–231. ( 10.1093/beheco/aru175) [DOI] [Google Scholar]
- 19.Latty T, Ramsch K, Ito K, Nakagaki T, Sumpter DJ, Middendorf M, Beekman M. 2011. Structure and formation of ant transportation networks. J. R. Soc. Interface 8, 1298–1306. ( 10.1098/rsif.2010.0612) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Aron S, Pasteels JM, Deneubourg JL. 1989. Trail-laying behaviour during exploratory recruitment in the Argentine ant, Iridomyrmex humilis (Mayr). Biol. Behav. 14, 207–217. [Google Scholar]
- 21.Goss S, Aron S, Deneubourg JL, Pasteels JM. 1989. Self-organized shortcuts in the Argentine ant. Naturwissenschaften 76, 579–581. ( 10.1007/BF00462870) [DOI] [Google Scholar]
- 22.Cook Z, Franks D, Robinson EH. 2014. Efficiency and robustness of ant colony transportation networks. Behav. Ecol. Sociobiol. 68, 509–517. ( 10.1007/s00265-013-1665-8) [DOI] [Google Scholar]
- 23.Jander R, Daumer K. 1974. Guide-line and gravity orientation of blind termites foraging in the open (Termitidae: Macrotermes, Hospitalitermes). Insectes Sociaux 21, 45–69. ( 10.1007/BF02222979) [DOI] [Google Scholar]
- 24.Dussutour A, Beshers S, Deneubourg JL, Fourcassié V. 2009. Priority rules govern the organization of traffic on foraging trails under crowding conditions in the leaf-cutting ant Atta colombica. J. Exp. Biol. 212, 499–505. ( 10.1242/jeb.022988) [DOI] [PubMed] [Google Scholar]
- 25.Couzin ID, Franks NR. 2003. Self-organized lane formation and optimized traffic flow in army ants. Proc. R. Soc. Lond. B 270, 139–146. ( 10.1098/rspb.2002.2210) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Hölldobler B, Möglich M, Maschwitz U. 1974. Communication by tandem running in the ant Camponotus sericeus. J. Comp. Physiol. 90, 105–127. ( 10.1007/BF00694481) [DOI] [Google Scholar]
- 27.von Beeren C, Lizon à l'Allemand S, Hashim R, Witte V. 2014. Collective exploitation of a temporally unpredictable food source: mushroom harvesting by the ant Euprenolepis procera. Anim. Behav. 89, 71–77. ( 10.1016/j.anbehav.2013.12.006) [DOI] [Google Scholar]
- 28.Beckers R, Deneubourg JL, Goss S, Pasteels JM. 1990. Collective decision making through food recruitment. Insectes Sociaux 37, 258–267. ( 10.1007/BF02224053) [DOI] [Google Scholar]
- 29.Nicolis SC, Deneubourg JL. 1999. Emerging patterns and food recruitment in ants: an analytical study. J. Theoret. Biol. 198, 575–592. ( 10.1006/jtbi.1999.0934) [DOI] [PubMed] [Google Scholar]
- 30.Sumpter DJT, Beekman M. 2003. From nonlinearity to optimality: pheromone trail foraging by ants. Anim. Behav. 66, 273–280. ( 10.1006/anbe.2003.2224) [DOI] [Google Scholar]
- 31.Reid CR, Sumpter DJT, Beekman M. 2011. Optimisation in a natural system: Argentine ants solve the Towers of Hanoi. J. Exp. Biol. 214, 50–58. ( 10.1242/jeb.048173) [DOI] [PubMed] [Google Scholar]
- 32.Dussutour A, Nicolis SC, Shephard G, Beekman M, Sumpter DJT. 2009. The role of multiple pheromones in food recruitment by ants. J. Exp. Biol. 212, 2337–2348. ( 10.1242/jeb.029827) [DOI] [PubMed] [Google Scholar]
- 33.Latty T, Beekman M. 2013. Keeping track of changes: the performance of ant colonies in dynamic environments. Anim. Behav. 85, 637–643. ( 10.1016/j.anbehav.2012.12.027) [DOI] [Google Scholar]
- 34.Witte V, Attygalle AB, Meinwald J. 2007. Complex chemical communication in the crazy ant Paratrechina longicornis Latreille (Hymenoptera: Formicidae). Chemoecology 17, 57–62. ( 10.1007/s00049-006-0364-6) [DOI] [Google Scholar]
- 35.Boch R, Shearer DA, Stone BC. 1962. Identification of iso-amyl acetate as an active component in the sting pheromone of the honey bee. Nature 195, 1018–1020. ( 10.1038/1951018b0) [DOI] [PubMed] [Google Scholar]
- 36.Blum MS. 1969. Alarm pheromones. Annu. Rev. Entomol. 14, 57–80. ( 10.1146/annurev.en.14.010169.000421) [DOI] [Google Scholar]
- 37.Von Frisch K. 1955. The dancing bees: an account of the life and senses of the honey bee. New York, NY: Harcourt, Brace. [Google Scholar]
- 38.Currie CR, Scott JA, Summerbell RC, Malloch D. 1999. Fungus-growing ants use antibiotic-producing bacteria to control garden parasites. Nature 398, 701–704. ( 10.1038/19519) [DOI] [Google Scholar]
- 39.Pinto-Tomás AA, Anderson MA, Suen G, Stevenson DM, Chu FST, Cleland WW, Weimer PJ, Currie CR. 2009. Symbiotic nitrogen fixation in the fungus gardens of leaf-cutter ants. Science 326, 1120–1123. ( 10.1126/science.1173036) [DOI] [PubMed] [Google Scholar]
- 40.Wirth R. 2003. Herbivory of leaf-cutting ants: a case study on Atta colombica in the tropical rainforest of Panama. Berlin, Germany: Springer Science & Business Media. ( 10.1007/978-3-662-05259-4) [DOI] [Google Scholar]
- 41.Seeley TD. 1989. The honey bee colony as a superorganism. Am. Sci. 77, 546–553. ( 10.2307/27856005) [DOI] [Google Scholar]
- 42.Rinderer T, Collins A, Tucker K. 1985. Honey production and underlying nectar harvesting activities of Africanized and European honeybees. J. Apic. Res. 24, 161–167. ( 10.1080/00218839.1985.11100666) [DOI] [Google Scholar]
- 43.Cook SC, Eubanks MD, Gold RE, Behmer ST. 2010. Colony-level macronutrient regulation in ants: mechanisms, hoarding and associated costs. Anim. Behav. 79, 429–437. ( 10.1016/j.anbehav.2009.11.022) [DOI] [Google Scholar]
- 44.Fewell J, Winston M. 1992. Colony state and regulation of pollen foraging in the honey bee, Apis mellifera L. Behav. Ecol. Sociobiol. 30, 387–393. ( 10.1007/BF00176173) [DOI] [Google Scholar]
- 45.Moritz B, Crailsheim K. 1987. Physiology of protein digestion in the midgut of the honeybee (Apis mellifera L.). J. Insect Physiol. 33, 923–931. ( 10.1016/0022-1910(87)90004-7) [DOI] [Google Scholar]
- 46.Simpson J. 1955. The significance of the presence of pollen in the food of worker larvae of the honey-bee. Q. J. Microsc. Sci. 3, 117–120. [Google Scholar]
- 47.Crailsheim K, Hrassnigg N, Gmeinbauer R, Szolderits MJ, Schneider LHW, Brosch U. 1993. Pollen utilization in non-breeding honeybees in winter. J. Insect Physiol. 39, 369–373. ( 10.1016/0022-1910(93)90024-L) [DOI] [Google Scholar]
- 48.Pratt SC. 1999. Optimal timing of comb construction by honeybee (Apis mellifera) colonies: a dynamic programming model and experimental tests. Behav. Ecol. Sociobiol. 46, 30–42. ( 10.1007/s002650050589) [DOI] [Google Scholar]
- 49.Sugden EA, McAllen RL. 1994. Observations on foraging, population and nest biology of the Mexican honey wasp, Brachygastra mellifica (Say) in Texas [Vespidae: Polybiinae]. J. Kansas Entomol. Soc. 67, 141–155. ( 10.2307/25085503) [DOI] [Google Scholar]
- 50.Burgett DM, Young RG. 1974. Lipid storage by honey ant repletes. Ann. Entomol. Soc. Am. 67, 743–744. ( 10.1093/aesa/67.5.743) [DOI] [Google Scholar]
- 51.Conway JR. 1977. Analysis of clear and dark amber repletes of the honey ant, Myrmecocystus mexicanus hortideorum. Ann. Entomol. Soc. Am. 70, 367–369. ( 10.1093/aesa/70.3.367) [DOI] [Google Scholar]
- 52.Rissing SW. 1984. Replete caste production and allometry of workers in the honey ant, Myrmecocystus mexicanus Wesmael (Hymenoptera: Formicidae). J. Kansas Entomol. Soc. 57, 347–350. ( 10.2307/25084522) [DOI] [Google Scholar]
- 53.Sendova-Franks AB, Hayward RK, Wulf B, Klimek T, James R, Planqué R, Britton NF, Franks NR. 2010. Emergency networking: famine relief in ant colonies. Anim. Behav. 79, 473–485. ( 10.1016/j.anbehav.2009.11.035) [DOI] [Google Scholar]
- 54.Lanan M, Dornhaus A, Bronstein J. 2011. The function of polydomy: the ant Crematogaster torosa preferentially forms new nests near food sources and fortifies outstations. Behav. Ecol. Sociobiol. 65, 959–968. ( 10.1007/s00265-010-1096-8) [DOI] [Google Scholar]
- 55.Chopra S, Sodhi MS. 2012. Managing risk to avoid supply-chain breakdown. MIT Sloan Management Review, Fall 2004.
- 56.Sheffi Y, Rice JB Jr. 2005. A supply chain view of the resilient enterprise. MIT Sloan Management Review, vol. 47.
- 57.Schmolke A. 2009. Benefits of dispersed central-place foraging: an individual-based model of a polydomous ant colony. Am. Nat. 173, 772–778. ( 10.1086/598493) [DOI] [PubMed] [Google Scholar]
- 58.McIver JD. 1991. Dispersed central place foraging in Australian meat ants. Insectes Sociaux 38, 129–137. ( 10.1007/BF01240963) [DOI] [Google Scholar]
- 59.Rinderer TE, Baxter JR. 1978. Effect of empty comb on hoarding behavior and honey production of the honey bee. J. Econ. Entomol. 71, 757–759. ( 10.1093/jee/71.5.757) [DOI] [Google Scholar]
- 60.Dreller C, Page RE Jr, Fondrk MK. 1999. Regulation of pollen foraging in honeybee colonies: effects of young brood, stored pollen, and empty space. Behav. Ecol. Sociobiol. 45, 227–233. ( 10.1007/s002650050557) [DOI] [Google Scholar]
- 61.Camazine S. 1993. The regulation of pollen foraging by honey bees: how foragers assess the colony's need for pollen. Behav. Ecol. Sociobiol. 32, 265–272. ( 10.1007/BF00166516) [DOI] [Google Scholar]
- 62.Pankiw T, Page RE Jr, Kim Fondrk M. 1998. Brood pheromone stimulates pollen foraging in honey bees (Apis mellifera). Behav. Ecol. Sociobiol. 44, 193–198. ( 10.1007/s002650050531) [DOI] [Google Scholar]
- 63.Dreller C, Tarpy DR. 2000. Perception of the pollen need by foragers in a honeybee colony. Anim. Behav. 59, 91–96. ( 10.1006/anbe.1999.1303) [DOI] [PubMed] [Google Scholar]
- 64.Calderone NW, Johnson BR. 2002. The within-nest behaviour of honeybee pollen foragers in colonies with a high or low need for pollen. Anim. Behav. 63, 749–758. ( 10.1006/anbe.2001.1957) [DOI] [Google Scholar]
- 65.Wilson E. 1984. The relation between caste ratios and division of labor in the ant genus Pheidole (Hymenoptera: Formicidae). Behav. Ecol. Sociobiol. 16, 89–98. ( 10.1007/BF00293108) [DOI] [Google Scholar]
- 66.Gill RJ, Ramos-Rodriguez O, Raine NE. 2012. Combined pesticide exposure severely affects individual- and colony-level traits in bees. Nature 491, 105–108. ( 10.1038/nature11585) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Perry CJ, Søvik E, Myerscough MR, Barron AB. 2015. Rapid behavioral maturation accelerates failure of stressed honey bee colonies. Proc. Natl Acad. Sci. USA 112, 3427–3432. ( 10.1073/pnas.1422089112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Charbonneau D, Hillis N, Dornhaus A. 2015. ‘Lazy’ in nature: ant colony time budgets show high ‘inactivity’ in the field as well as in the lab. Insectes Sociaux 62, 31–35. ( 10.1007/s00040-014-0370-6) [DOI] [Google Scholar]
- 69.Karsai I, Wenzel JW. 1998. Productivity, individual-level and colony-level flexibility, and organization of work as consequences of colony size. Proc. Natl Acad. Sci. USA 95, 8665–8669. ( 10.1073/pnas.95.15.8665) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Schmid-Hempel P. 1990. Reproductive competition and the evolution of work load in social insects. Am. Nat. 135, 501–526. ( 10.2307/2462050) [DOI] [Google Scholar]
- 71.Winston ML, Fergusson LA. 1985. The effect of worker loss on temporal caste structure in colonies of the honeybee (Apis mellifera L.). Can. J. Zool. 63, 777–780. ( 10.1139/z85-113) [DOI] [Google Scholar]
- 72.Granovskiy B, Latty T, Duncan M, Sumpter DJT, Beekman M. 2012. How dancing honey bees keep track of changes: the role of inspector bees. Behav. Ecol. 23, 588–596. ( 10.1093/beheco/ars002) [DOI] [Google Scholar]
- 73.Beekman M, Sumpter DJT, Ratnieks FLW. 2001. Phase transition between disordered and ordered foraging in Pharaoh's ants. Proc. Natl Acad. Sci. USA 98, 9703–9706. ( 10.1073/pnas.161285298) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Seeley TD, Morse RA, Visscher PK. 1979. The natural history of the flight of honey bee swarms. Psyche: J. Entomol. 86, 103–113. ( 10.1155/1979/80869) [DOI] [Google Scholar]
- 75.Seeley TD, Buhrman SC. 1999. Group decision making in swarms of honey bees. Behav. Ecol. Sociobiol. 45, 19–31. ( 10.1007/s002650050536) [DOI] [Google Scholar]
- 76.Backen SJ, Sendova-Franks AB, Franks NR. 2000. Testing the limits of social resilience in ant colonies. Behav. Ecol. Sociobiol. 48, 125–131. ( 10.1007/s002650000219) [DOI] [Google Scholar]
- 77.Sendova-Franks AB, Franks NR. 1994. Social resilience in individual worker ants and its role in division of labour. Proc. R. Soc. Lond. B 256, 305–309. ( 10.1098/rspb.1994.0085) [DOI] [Google Scholar]
- 78.Grüter C, Czaczkes T, Ratnieks FW. 2011. Decision making in ant foragers (Lasius niger) facing conflicting private and social information. Behav. Ecol. Sociobiol. 65, 141–148. ( 10.1007/s00265-010-1020-2) [DOI] [Google Scholar]
- 79.Attygalle AB, Morgan ED. 1985. Ant trail pheromones. In Advances in insect physiology (eds Berridge JETMJ, Wigglesworth VB), pp. 1–30. New York, NY: Academic Press. [Google Scholar]
- 80.Jeanson R, Ratnieks FLW, Deneubourg JL. 2003. Pheromone trail decay rates on different substrates in the Pharaoh's ant, Monomorium pharaonis. Physiol. Entomol. 28, 192–198. ( 10.1046/j.1365-3032.2003.00332.x) [DOI] [Google Scholar]
- 81.Jackson DE, Martin SJ, Holcombe M, Ratnieks FLW. 2006. Longevity and detection of persistent foraging trails in Pharaoh's ants, Monomorium pharaonis (L.). Anim. Behav. 71, 351–359. ( 10.1016/j.anbehav.2005.04.018) [DOI] [Google Scholar]
- 82.Holmes MJ, Oldroyd BP, Allsopp MH, Lim J, Wossler TC, Beekman M. 2010. Maternity of emergency queens in the Cape honey bee, Apis mellifera capensis. Mol. Ecol. 19, 2792–2799. ( 10.1111/j.1365-294X.2010.04683.x) [DOI] [PubMed] [Google Scholar]
- 83.Linksvayer TA, Janssen MA. 2009. Traits underlying the capacity of ant colonies to adapt to disturbance and stress regimes. Syst. Res. Behav. Sci. 26, 315–329. ( 10.1002/sres.928) [DOI] [Google Scholar]
- 84.Ponomarov SY, Holcomb MC. 2009. Understanding the concept of supply chain resilience. Int. J. Log. Manag. 20, 124–143. ( 10.1108/09574090910954873) [DOI] [Google Scholar]
- 85.Bhamra R, Dani S, Burnard K. 2011. Resilience: the concept, a literature review and future directions. Int. J. Prod. Res. 49, 5375–5393. ( 10.1080/00207543.2011.563826) [DOI] [Google Scholar]
- 86.Levin SA, et al. 1998. Resilience in natural and socioeconomic systems. Environ. Dev. Econ. 3, 221–262. ( 10.1017/S1355770X98240125) [DOI] [Google Scholar]
- 87.Engle NL. 2011. Adaptive capacity and its assessment. Glob. Environ. Change 21, 647–656. ( 10.1016/j.gloenvcha.2011.01.019) [DOI] [Google Scholar]
- 88.Haimes YY, Jiang P. 2001. Leontief-based model of risk in complex interconnected infrastructures. J. Infrastruct. Syst. 7, 1–12. ( 10.1061/(ASCE)1076-0342(2001)7:1(1)) [DOI] [Google Scholar]
- 89.Ouyang M. 2014. Review on modeling and simulation of interdependent critical infrastructure systems. Reliab. Eng. Syst. Safety 121, 43–60. ( 10.1016/j.ress.2013.06.040) [DOI] [Google Scholar]
- 90.Dudenhoeffer DD, Permann MR, Manic M. 2006. CIMS: a framework for infrastructure interdependency modeling and analysis. In Proc. 38th Conf. on Winter Simulation, Monterey, CA, 3–6 December 2006, pp. 478–485.
- 91.Rinaldi SM.2004. Modeling and simulating critical infrastructures and their interdependencies. In Proc. 37th Annual Hawaii Int. Conf. on System Sciences, Big Island, HI, 5–8 January 2004 . ( ) [DOI]
- 92.Carpenter S, Walker B, Anderies JM, Abel N. 2001. From metaphor to measurement: resilience of what to what? Ecosystems 4, 765–781. ( 10.1007/s10021-001-0045-9) [DOI] [Google Scholar]
- 93.Clark R, Fewell J. 2014. Transitioning from unstable to stable colony growth in the desert leafcutter ant Acromyrmex versicolor. Behav. Ecol. Sociobiol. 68, 163–171. ( 10.1007/s00265-013-1632-4) [DOI] [Google Scholar]
- 94.Kates RW, Colten CE, Laska S, Leatherman SP. 2006. Reconstruction of New Orleans after Hurricane Katrina: a research perspective. Proc. Natl Acad. Sci. USA 103, 14 653–14 660. ( 10.1073/pnas.0605726103) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.LeBrun EG, Moffett M, Holway DA. 2011. Convergent evolution of levee building behavior among distantly related ant species in a floodplain ant assemblage. Insectes Sociaux 58, 263–269. ( 10.1007/s00040-011-0151-4) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Zakharov AA, Zakharov RA. 2014. Ants under the conditions of an extremely hot summer. Entmol. Rev. 94, 526–540. ( 10.1134/S0013873814040071) [DOI] [Google Scholar]
- 97.Dobson I, Carreras BA, Lynch VE, Newman DE. 2007. Complex systems analysis of series of blackouts: cascading failure, critical points, and self-organization. Chaos Interdiscip. J. Nonlinear Sci. 17, 026103 ( 10.1063/1.2737822) [DOI] [PubMed] [Google Scholar]
- 98.Smart AG, Amaral LAN, Ottino JM. 2008. Cascading failure and robustness in metabolic networks. Proc. Natl Acad. Sci. USA 105, 13 223–13 228. ( 10.1073/pnas.0803571105) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Barron AB. 2015. Death of the bee hive: understanding the failure of an insect society. Curr. Opin. Insect Sci. 10, 45–50. ( 10.1016/j.cois.2015.04.004) [DOI] [PubMed] [Google Scholar]
- 100.Choi TY, Dooley KJ, Rungtusanatham M. 2001. Supply networks and complex adaptive systems: control versus emergence. J. Operat. Manag. 19, 351–366. ( 10.1016/S0272-6963(00)00068-1) [DOI] [Google Scholar]
- 101.Vespignani A. 2010. Complex networks: the fragility of interdependency. Nature 464, 984–985. ( 10.1038/464984a) [DOI] [PubMed] [Google Scholar]
- 102.Kim Y-J, Thottan M, Kolesnikov V, Lee W. 2010. A secure decentralized data-centric information infrastructure for smart grid. IEEE Commun. Mag. 48, 58–65. ( 10.1109/MCOM.2010.5621968) [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.