

Abstract
In neuronal recording studies on anesthetized animals, reliable measures for the transitional moment of consciousness are frequently required. Previous findings suggest that pupil fluctuations reflect the neuronal states during quiet wakefulness, whose correlation was unknown for the anesthetized condition. Here, we investigated the pupillary changes under isoflurane anesthesia simultaneously with the electroencephalogram (EEG) and electromyogram (EMG). The pupil was tracked by using a region-based active contour model. The dose was given to the animal in a stepwise increasing mode (simulating induction of anesthesia) or in a stepwise decreasing mode (simulating emergence of anesthesia). We found that the quickly widening pupil action (mydriasis) characterizes the transitional state in anesthesia. Mydriasis occurred only in the light dose in the emergence phase, and the events were accompanied by an increase of burst activity in the EEG followed by EMG activity in 47% of the mydriasis events. Our findings suggest that recording such pupil changes may offer a noncontact monitoring tool for indexing the transitional state of the brain, particularly when a lower threshold dose is applied.
Keywords: eye tracker, EEG, EMG, mydriasis, anesthesia
INTRODUCTION
General anesthesia is a state of unconsciousness with varying depths. Anesthetics modulate the spontaneous network of the brain [1], inducing spatial and temporal change of the electrophysiological state of the cortical area. As the anesthetic state changes over time, monitoring patients during anesthesia is important for the safety of the surgery. For this reason, there are demands for continuous and noninvasive monitoring of the brain state during anesthesia.
Pupillometry is a method for tracking neurological responses by calculating the changes in the pupil diameter. During 50 years of history, after several similar studies [2,3,4,5] have shown that sensory response and mental effort correlate with pupil dilation, many studies have reported the correlation between pupil size and mental contents [6,7,8,9,10]. Recently, pupillometric researches have widened the field of neuroscience. McGinley, et al. [11] discovered that locomotion and arousal modulate the pupil diameter, hippocampal ripple rate, and auditory cortex membrane potential simultaneously. Vinck, et al. [12] found that the pupil diameter also reflects the difference of visual cortex activation between quiescence and locomotion. Reimer, et al. [13] showed that pupil dilation and constriction track the cortical membrane potential during "quiet wakefulness"—the moments between exploratory behaviors. The interesting part of the covariation of the pupil and neurological activity is that the pupil diameter also modulates the neural state during the absence of consciousness. The pupil size of rats under urethane [14] and isoflurane [15] anesthesia correlated with slow fluctuations of the cortical state measured with the electrocorticogram. This study indicates that the pupil diameter change during anesthesia may be an indicator of the electrophysiological state transition of the brain.
In this study, we investigated the correlation of pupil size, electroencephalogram (EEG) signals, and electromyogram (EMG) signals during isoflurane anesthesia. Isoflurane is an inhalation drug that safely and reversibly induces the surgical level of anesthesia [16] with its own electrophysiological characteristics. The depth of isoflurane anesthesia was characterized by periods of high-voltage activity that is interspersed quasi-periodically with long (approximately 10~20 sec) periods of isoelectric activity [1,17]. This so-called burst-suppression pattern, which, when it appears on EEG, means that <5% of the cortical neurons and <40% of the thalamic neurons are active [18], is used as an index of the depth of anesthesia. The importance of this suppression pattern is that it can be universally observed in various species including human, birds, goats, and rodents [1,19,20,21,22,23]. This feature allows conducting an experiment with mouse EEG for predicting the cortical state of the human brain during isoflurane anesthesia. Although which mental state the "burst" of this suppression pattern represents is unknown, the burst signals have been observed to occur frequently at a low dose of isoflurane anesthesia [21,23,24]. Therefore, in this study, we correlated the occurrence of the burst-suppression pattern with pupillary change. As there is some relation between cortical burst and the level of consciousness while there is also some connection between the pupil diameter and arousal level [25,26], some concurrent feature may be expected to exist between isoflurane-induced burst and changes in the pupil diameter with the level of arousal. By recording pupil fluctuations with the EEG and EMG simultaneously with this assumption, we observed that pupillary change predicts the transition of the burst-suppression pattern during the change in the level of anesthesia from the deep to the light stage.
MATERIALS AND METHODS
All surgical and experimental procedures were performed in accordance with the institutional guidelines for the Institutional Animal Care and Use Committee, which follows Act 1992 of the Korea Lab animal Care Regulations and associated guidelines. All efforts were made to minimize the number of animals and the animal's pain and suffering.
Surgical preparation
Experiments were conducted on adult male B6/129 (hybrid of C57/Bl6X129) F1 mice, weighing 20~35 g, aged 8~12 weeks (n=10). For surgery, animals were maintained in deep anesthesia induced by an intraperitoneal injection of a mixture of ketamine (120 mg/kg) and xylazine (6 mg/kg). A stereotaxic apparatus (Kopf instrument, USA) was used to fix the target area, and the animal's head was restrained by ear bars during surgery. For EEG/EMG electrode implantation, the skin was opened carefully and the surface of the skull was cleaned and dried for implantation of screws. The microscrew electrodes were implanted on the frontal area (anterioposterior (AP) 1.2 mm; mediolateral (ML) 1.5 mm; dorsoventral (DV) 0 mm), parietal area (AP -2 mm; ML -2 mm; DV 0 mm), and both the left and right side of the cerebellum for ground and reference electrodes. For EMG, a tungsten wire (0.003 mm, A-M Systems, USA) electrode was implanted into the neck muscle beyond the skull. All electrode coordinates follow the mouse brain atlas [27]. Next, the electrodes were fixed with dental cement (Vertex Self Curing, Dentimex, Germany). Scalp skin was sutured after cement hardened. All animals had at least one week of recovery period from surgery before their first recording session started.
Pupil and electrophysiological recordings
The recording was performed in a Faraday cage with dim light. Prior to recording, mice were anesthetized in an induction chamber (4% concentration of isoflurane combined with pure oxygen) for 4 to 5 min. After anesthesia on a surgical level was assured by toe and tail pinching, the mouse was placed on a stereotaxic apparatus, and then the nose was connected to a respiration tube. Gel-type eye drops (Liposic ophthalmic gel, Bausch+Lomb, Germany) were spread on the surface of the animal's eye to prevent drying.
Two different paradigms of applying the dose were tested. One paradigm mimicked the induction of anesthesia (referred to as the induction phase), and the dose given to the animal started from 0.5%, with 0.5% step-ups to the highest level. The other paradigm mimicked the emergence from anesthesia (referred to as the induction phase), and the dose given to the animal started from the highest level, with 0.5% step-downs to the lowest level, which was 0.5% or 0.3%. The highest level was determined prior to recording in each mouse, which was set to the maximal dose with no sign of breathing difficulty for several minutes. The highest level was 2.0% in six mice, 1.5% in three mice, and 1.0% in one mouse. During recording, each dose was tested for 15 min. In two mice, the dose was down to 0.3% in the emergence phase, when the animal showed no mydriatic pupil responses at 0.5%, and no response to tail/toe pinch. The induction and emergence phases were tested 16 times in 10 mice.
The pupil was recorded with an infrared LED webcam (DABOA-DA500, DABOA, Korea). In order to avoid the image reflection, the infrared LED was removed from the camera. The camera was fixed on a stereotaxic arm to maintain the focusing distance, which was 0.7~1.3 cm from the mouse's left eye. A custom-built infrared illuminator (850 nm, LED) was fixed above the eyelid. The light from the illuminator was detected by a photodetector and was converted to a sync signal. The video was recorded with 640×480 pixels at 30 fps. To avoid the dark adaptation effect, recording started 15~30 minutes after the elimination of the visible ray.
EEG and EMG were recorded with Intan RHD2000 data acquisition system with RHD2000 Interface software (Intan technologies, USA, setting 1), or SynAmps2 amplifiers with Neuroscan Scan 4.3 software (Compudemics NeuroScan, USA, setting 2), both at a sampling frequency of 2 kHz. EMG signal was re-referenced by subtracting the averaged value of all the electrodes to eliminate the ground and reference.
Pupil detection
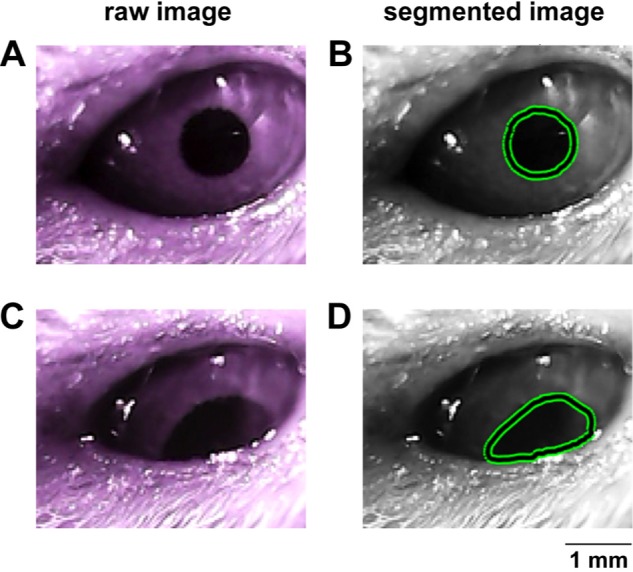
Pupil dynamics were analyzed frame by frame based on a region-based active contour model, by using a localized_seg.m function presented by Lankton and Tannenbaum [28]. The code is downloadable from http://www.shawnlankton.com/2008/04/active-contour-matlab-code-demo/. The model is based on the Snake algorithm, but finds the segment from the local image statistics rather than global image statistics. Compared to the Hough circle detection algorithm [28,29,30], a widely used shape detection technique, the region-based active contour model finds the mouse pupil more accurately and robustly. Briefly, the algorithm starts segmentation with a rough 'initial contour' drawn manually on the image, then optimizes the region contour by deforming it iteratively while comparing foreground and background image contrast statistics. The contour of the pupil from each frame was used as an initial contour in the subsequent frame. The contour of a full or partially occluded pupil is shown in Fig. 1. Additional post-hoc processing included error correction when necessary. The pixel coordinate was converted to mm. The constriction/dilation responses of the pupil were estimated according to the area of pupil.
Fig. 1. Detection of pupillary aperture. Captured images of NIR camera for (A) the pupil with a full shot and (B) the pupil occluded by the eyelid. Segmented images for (C) the pupil with a full shot and (B) the pupil occluded by the eyelid. The green lines indicate the detected boundary of the pupil. The area and the center of the contour are calculated to access the pupil size and location. The occluded pupil was identified by the ratio of vertical and horizontal length, and was removed in the analysis.
Data analysis
After converting the image data into pupil data, the zero time in pupil and EEG/EMG signals were synchronized using the synch signal. As the events of interest, the mydriasis (quickly widening of pupil) in the pupil responses and the burst of EEG were the focus in this study. Typically, mydriasis is referred to as pupil dilation under sympathetic stimulation; here, we refer to it as quickly widening pupillary dilation.
Detection of mydriasis
The criteria of mydriasis are as follows: (i) the pupillary aperture starts from a constricted pupil to full dilation, and, (ii) the pupillary dilation occurs within 1 min. The threshold for the constricted pupil was 1/8 in terms of the ratio of the pupil area to eye surface (the area between two eyelids), whereas the threshold for the full dilation was 1/3. The thresholds were determined after a visual inspection of the images. The instantaneous dilation speed was also investigated by calculating the temporal change in the pupil size (mm2/sec).
Detection of EEG burst
The burst in EEG was defined by a large and fast fluctuation of EEG for a short period of time, and the burst ratio was calculated according to Land et al. [24]. First, the threshold was determined as a median value of the EEG amplitude during the anesthetic state, acquired for 30 min at a 1.0% concentration dose. The periods of absolute EEG higher than the threshold were counted as the burst period, and the burst ratio was defined by the ratio of the burst period to the observation time period.
Results
Mydriasis occurring in the recovery phase of anesthesia
Different doses of anesthesia were tested to mimic the induction of anesthesia (induction phase, stepwise increase in anesthetic dose), or emergence of anesthesia (emergence phase, stepwise decrease in anesthetic dose). Changes in the pupillary size were observed in both the induction and recovery modes, including the highest dose with typical patterns of fluctuations, such as slowly widening or shifting motions. The pupil fluctuations occurred irregularly in various patterns. The mydriatic patterns (full dilation of the pupil in a short period of time) were observed in all the recorded mice, but they occurred only in the emergence phase (43 events in total). The maximal dose showing the mydriatic patterns was referred as the critical dose for mydriasis occurrence. The critical dose was 1% in two mice, 0.5% in six mice, and 0.3% in two mice. The occurrence rate of mydriasis in 0.3% was 10±8.4 times per hour; in 0.5% it was 9±1.8 per hour, whereas the value was 1.6±0.54 per hour in 1.0% dose.
EEG activation increased during mydriasis
Fig. 2 shows the representative traces of pupillary responses and parietal EEG at two different doses (2% versus 0.3%) collected from the emergence phase. The dilation occurred slowly in 2%; hence, it was not counted as a mydriasis event. Although the maximum size of pupil was similar, the EEG traces and spectrum showed a distinctive feature at low and high doses. The parietal EEG showed a slow oscillation under deep anesthesia, whereas the EEG was fast and irregular under light anesthesia.
Fig. 2. Pupil size and EEG. Time traces of pupil size and parietal EEG activation in (A) deep anesthesia (2% isoflurane) and (B) light anesthesia (0.5%) in emergence phase. The quickly widening dilation of pupil exemplified in (B) was counted as mydriasis. The spectrograms of the corresponding EEG traces were shown accordingly.

Mydriasis precedes the changes in EEG and EMG
Fig. 3 shows representative traces of pupil size, frontal and parietal EEG, and EMG. The earliest phase of pupil dilation precedes the responses in EEG and EMG. The typical patterns of EEG under anesthesia are characterized by a slow oscillation with infrequent burst activity (exemplified in Fig. 2B), changing to fast and complex EEG rhythms as the pupil rapidly dilated (exemplified in Fig. 2C). The mydriasis and burst events were logged according to the definition depicted in the Methods. In ten mice, 43 mydriasis events were logged. The burst of EEG occurred intermittently in an episodic way at the critical dose. The episodic EEG burst became continual after mydriasis was monitored (exemplified in Fig. 4). All the mydriasis events were followed by a continual burst of EEG, although the patterns were not unitary (i.e., frequently a occurring burst or a continuous burst). None of the continual burst episodes preceded the mydriasis in the recorded mice. EMG with a high amplitude was also logged, and 47% of the mydriasis were accompanied by EMG responses.
Fig. 3. Pupil dilation precedes burst of EEG. (A) A representative trace of pupil size, frontal and parietal EEG data and EMG during mydriasis. EEG and EMG (B) before and (C) after onset of mydriasis.
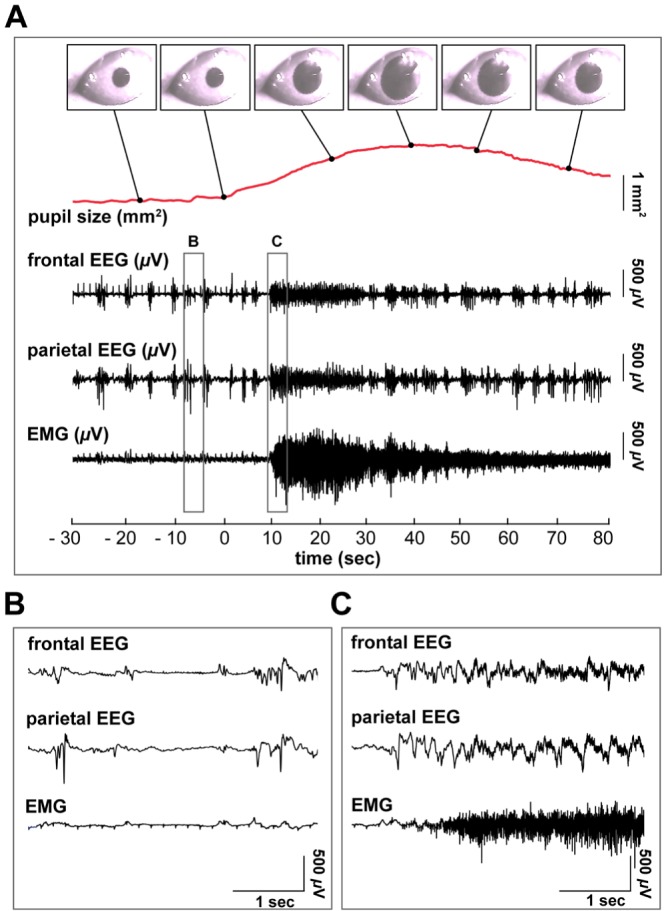
Fig. 4. Pupil dilation speed during mydriasis coincides with burst ratio of EEG. (A) Raw pupil size and (B) dilation rate of a mydriasis period with (C) the burst ratio and (D) the spectrogram. The burst ratio maximized at the point of the dilation rate of pupil size reaches at the peak (approximately 10 sec after the mydriasis onset).
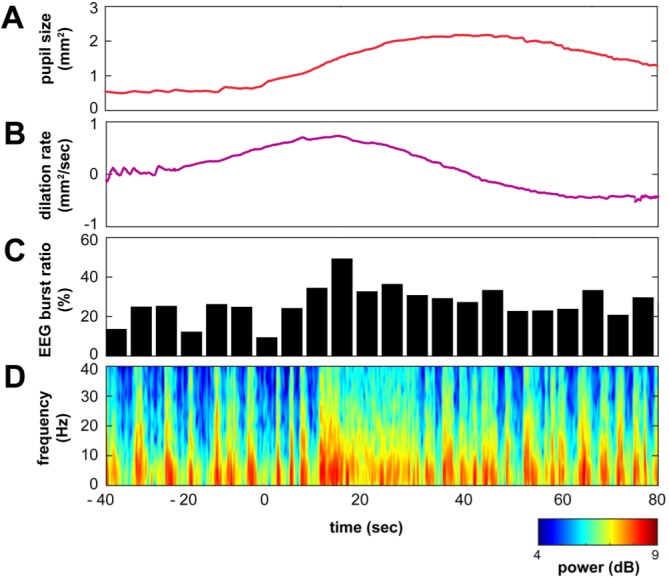
The speed of pupil dilation coincides with a burst ratiopattern in EEG
The burst ratio of EEG was calculated every 5 sec and compared to the pupil dynamics. As shown in Fig. 4, the temporal pattern of the burst ratio coincides with the dilation speed rather than the pupil size. The peak latencies of the EEG burst ratio were calculated with respect to the pupil size and the dilation. Within 5 sec of tolerance, no delay was observed with respect to the dilation speed, whereas the latency between the peaks of EEG burst ratio and pupil size was 14±3 sec. Considering the inverse of the burst ratio has been considered as anesthesia depth, pupil dilation indicates the transition of the brain state, which is consistent with previously reported observations by Reimer, et al. [13].
Discussion
Path-dependency of mydriasis
In the present study, mydriasis occurred on the step-wise emergence from anesthetic state, while it did not occur during induction of anesthesia. This result is in line with previous observation that there is a path-dependent feature of brain state during transition of (the) anesthetic state in ketamine-xylazine cocktail induced anesthesia [31], and the observation of Friedman, et al. [32] that there is the path-dependency of behavior response between induction and emergence period of halothane and isoflurane anesthesia. Both observations suppose that this asymmetry between induction and emergence of anesthesia generates from neural inertia, which means the inherent resistance to changes in arousal state. Our observation implies that mydriasis can be a marker for neural inertia.
Mydriasis as a guidance of anesthetic level transition on recovery phase
The simultaneous observation of EEG and EMG shows that the mydriasis implies a mixed state of anesthetized and conscious states. All the mydriasis events were accompanied by the increase of burst activity in the EEG, and half of the events were followed by EMG activity. Considering the recovery of motion requires the transition from anesthetized state to awaken state, the mydriasis preceding the recovery of motion implies a sign of anesthetic level transition. While we claimed mydriasis is a fast dilation compared to other periods during anesthesia, the dilation speed is slower than awaken state shown in the previous studies (Reimer et al, 2014). We observed strong slow rhythm (Fig. 3C) on both frontal and parietal area during mydriasis, which did not occur on awaken animal. Other observation is that some of mydriasis events did not came with EMG signal, a feature of wakefulness. For these reasons, mydriasis is a post-stage of awakening, not the awaken state itself. For these reasons, we suggest that mydriasis is an indicator of transitional state before recovering consciousness.
Monitoring of anesthetized animal is essential on animal research, especially on modern neuroscience such as using fMRI equipment or two-photon microscope. There are studies that dilation and constriction of pupil is correlated with brain state on anesthetized mice and rats [14,15]. Our observation shows that pupil diameter precedes electrophysiological change of brain during transitional state of anesthesia. Observing pupil diameter enables watching out transitional state of anesthesia without recording electrophysiological activation during surgery and experiment of animal research.
Acknowledgements
Study conception and design: JHC, Acquisition of data: JEK, Analysis and interpretation of data: JEK, HBH, JHC, Programming: HBH, Writing the manuscript: JHC, JEK. This work was National Research Council of Science and Technology on project "Development of Solution for Diagnosis, Treatment and Care system of Dementia".
References
- 1.Sellers KK, Bennett DV, Hutt A, Fröhlich F. Anesthesia differentially modulates spontaneous network dynamics by cortical area and layer. J Neurophysiol. 2013;110:2739–2751. doi: 10.1152/jn.00404.2013. [DOI] [PubMed] [Google Scholar]
- 2.Hess EH, Polt JM. Pupil size as related to interest value of visual stimuli. Science. 1960;132:349–350. doi: 10.1126/science.132.3423.349. [DOI] [PubMed] [Google Scholar]
- 3.Hess EH, Polt JM. Pupil size in relation to mental activity during simple problem-solving. Science. 1964;143:1190–1192. doi: 10.1126/science.143.3611.1190. [DOI] [PubMed] [Google Scholar]
- 4.Hess EH, Polt JM. Changes in pupil size as a measure of taste difference. Percept Mot Skills. 1966;23:451–455. [Google Scholar]
- 5.Kahneman D, Beatty J. Pupil diameter and load on memory. Science. 1966;154:1583–1585. doi: 10.1126/science.154.3756.1583. [DOI] [PubMed] [Google Scholar]
- 6.Koss MC. Pupillary dilation as an index of central nervous system alpha 2-adrenoceptor activation. J Pharmacol Methods. 1986;15:1–19. doi: 10.1016/0160-5402(86)90002-1. [DOI] [PubMed] [Google Scholar]
- 7.Peavler WS. Pupil size, information overload, and performance differences. Psychophysiology. 1974;11:559–566. doi: 10.1111/j.1469-8986.1974.tb01114.x. [DOI] [PubMed] [Google Scholar]
- 8.Aston-Jones G, Rajkowski J, Kubiak P, Alexinsky T. Locus coeruleus neurons in monkey are selectively activated by attended cues in a vigilance task. J Neurosci. 1994;14:4467–4480. doi: 10.1523/JNEUROSCI.14-07-04467.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Laeng B, Sirois S, Gredebäck G. Pupillometry: a window to the preconscious? Perspect Psychol Sci. 2012;7:18–27. doi: 10.1177/1745691611427305. [DOI] [PubMed] [Google Scholar]
- 10.Kloosterman NA, Meindertsma T, van Loon AM, Lamme VA, Bonneh YS, Donner TH. Pupil size tracks perceptual content and surprise. Eur J Neurosci. 2015;41:1068–1078. doi: 10.1111/ejn.12859. [DOI] [PubMed] [Google Scholar]
- 11.McGinley MJ, David SV, McCormick DA. Cortical Membrane Potential Signature of Optimal States for Sensory Signal Detection. Neuron. 2015;87:179–192. doi: 10.1016/j.neuron.2015.05.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Vinck M, Batista-Brito R, Knoblich U, Cardin JA. Arousal and locomotion make distinct contributions to cortical activity patterns and visual encoding. Neuron. 2015;86:740–754. doi: 10.1016/j.neuron.2015.03.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Reimer J, Froudarakis E, Cadwell CR, Yatsenko D, Denfield GH, Tolias AS. Pupil fluctuations track fast switching of cortical states during quiet wakefulness. Neuron. 2014;84:355–362. doi: 10.1016/j.neuron.2014.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Blasiak T, Zawadzki A, Lewandowski MH. Infra-slow oscillation (ISO) of the pupil size of urethane-anaesthetised rats. PLoS One. 2013;8:e62430. doi: 10.1371/journal.pone.0062430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Takahashi H, Tokushige H, Shiramatsu TI, Noda T, Kanzaki R. Covariation of pupillary and auditory cortical activity in rats under isoflurane anesthesia. Neuroscience. 2015;300:29–38. doi: 10.1016/j.neuroscience.2015.05.004. [DOI] [PubMed] [Google Scholar]
- 16.Ferron JF, Kroeger D, Chever O, Amzica F. Cortical inhibition during burst suppression induced with isoflurane anesthesia. J Neurosci. 2009;29:9850–9860. doi: 10.1523/JNEUROSCI.5176-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ching S, Purdon PL, Vijayan S, Kopell NJ, Brown EN. A neurophysiological-metabolic model for burst suppression. Proc Natl Acad Sci U S A. 2012;109:3095–3100. doi: 10.1073/pnas.1121461109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Steriade M, Amzica F, Contreras D. Cortical and thalamic cellular correlates of electroencephalographic burst-suppression. Electroencephalogr Clin Neurophysiol. 1994;90:1–16. doi: 10.1016/0013-4694(94)90108-2. [DOI] [PubMed] [Google Scholar]
- 19.Li D, Liang Z, Wang Y, Hagihira S, Sleigh JW, Li X. Parameter selection in permutation entropy for an electroencephalographic measure of isoflurane anesthetic drug effect. J Clin Monit Comput. 2013;27:113–123. doi: 10.1007/s10877-012-9419-0. [DOI] [PubMed] [Google Scholar]
- 20.Sellers KK, Bennett DV, Hutt A, Williams JH, Fröhlich F. Awake vs. anesthetized: layer-specific sensory processing in visual cortex and functional connectivity between cortical areas. J Neurophysiol. 2015;113:3798–3815. doi: 10.1152/jn.00923.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Antognini JF, Wang XW, Carstens E. Isoflurane anaesthetic depth in goats monitored using the bispectral index of the electroencephalogram. Vet Res Commun. 2000;24:361–370. doi: 10.1023/a:1006400315247. [DOI] [PubMed] [Google Scholar]
- 22.McIlhone AE, Beausoleil NJ, Johnson CB, Mellor DJ. Effects of isoflurane, sevoflurane and methoxyflurane on the electroencephalogram of the chicken. Vet Anaesth Analg. 2014;41:613–620. doi: 10.1111/vaa.12154. [DOI] [PubMed] [Google Scholar]
- 23.Kortelainen J, Väyrynen E, Jia X, Seppänen T, Thakor N. EEG-based detection of awakening from isoflurane anesthesia in rats. Conf Proc IEEE Eng Med Biol Soc. 2012;2012:4279–4282. doi: 10.1109/EMBC.2012.6346912. [DOI] [PubMed] [Google Scholar]
- 24.Land R, Engler G, Kral A, Engel AK. Auditory evoked bursts in mouse visual cortex during isoflurane anesthesia. PLoS One. 2012;7:e49855. doi: 10.1371/journal.pone.0049855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Honda S, Kohama T, Tanaka T, Yoshida H. Quantitative evaluation of arousal level based on the analyses of microsaccade rates and pupil fluctuations. Conf Proc IEEE Eng Med Biol Soc. 2013;2013:2108–2111. doi: 10.1109/EMBC.2013.6609949. [DOI] [PubMed] [Google Scholar]
- 26.Honda S, Kohama T, Tanaka T, Yoshida H. Quantitative assessments of arousal by analyzing microsaccade rates and pupil fluctuations prior to slow eye movements. Conf Proc IEEE Eng Med Biol Soc. 2014;2014:2229–2232. doi: 10.1109/EMBC.2014.6944062. [DOI] [PubMed] [Google Scholar]
- 27.Franklin KB, Paxinos G. The mouse brain in stereotaxic coordinates. San Diego, CA: Academic Press; 1997. [Google Scholar]
- 28.Lankton S, Tannenbaum A. Localizing region-based active contours. IEEE Trans Image Process. 2008;17:2029–2039. doi: 10.1109/TIP.2008.2004611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Blake A, Isard M. Active contours: the application of techniques from graphics, vision, control theory and statistics to visual tracking of shapes in motion. 1st ed. New York, NY: Springer; 1998. [Google Scholar]
- 30.Wang L, Chen Y, Pan X, Hong X, Xia D. Level set segmentation of brain magnetic resonance images based on local Gaussian distribution fitting energy. J Neurosci Methods. 2010;188:316–325. doi: 10.1016/j.jneumeth.2010.03.004. [DOI] [PubMed] [Google Scholar]
- 31.Hwang E, Kim S, Han K, Choi JH. Characterization of phase transition in the thalamocortical system during anesthesia-induced loss of consciousness. PLoS One. 2012;7:e50580. doi: 10.1371/journal.pone.0050580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Friedman EB, Sun Y, Moore JT, Hung HT, Meng QC, Perera P, Joiner WJ, Thomas SA, Eckenhoff RG, Sehgal A, Kelz MB. A conserved behavioral state barrier impedes transitions between anesthetic-induced unconsciousness and wakefulness: evidence for neural inertia. PLoS One. 2010;5:e11903. doi: 10.1371/journal.pone.0011903. [DOI] [PMC free article] [PubMed] [Google Scholar]