

Abstract
Ethanolamine (Etn) is a naturally occurring aminoalcohol necessary for synthesis of the phospholipid phosphatidylethanolamine (PE), a major component of biological membranes. We recently reported that Etn treatment increases cellular PE levels, thereby inducing cytoprotective autophagy and protecting against aging across species.
Keywords: aging, autophagy, ethanolamine, phosphatidylethanolamine, phosphatidylserine decarboxylase
Phosphatidylethanolamine (PE) is a central intermediate of lipid metabolism and a major component of biological membranes.1,2 Within cellular membranes, PE not only serves as a structural phospholipid but also regulates the tethering of proteins and fusion processes.3 PE orchestrates the interaction with membrane-associated proteins, both by directly ingraining proteins into membranes and as a precursor for glycosylphosphatidylinositol (GPI) anchors.3,4 Importantly, PE is also directly involved in the process of macroautophagy (hereafter termed autophagy), a lysosome-dependent cellular recycling mechanism5 that protects cells against lethal stress and extends longevity in model organisms.6 During autophagy, double-membraned structures that are highly abundant in PE engulf superfluous, supernumerary, or dysfunctional macromolecules or organelles contained in the cytoplasm, forming vesicles (autophagosomes). These autophagosomes then fuse with lysosomes to generate autophagolysosomes in which the luminal cargo is degraded. Importantly, PE serves as a lipid anchor for a protein essential for several steps of the autophagic process that is called autophagy related protein 8 (Atg8) in yeast or microtubule-associated protein 1A/1B-light chain 3 (LC3) in mammalian cells.7 The importance of LC3 (or Atg8) in the autophagic process is reflected by the fact that many assays designed to measure autophagy rely on quantitation of the conjugation of LC3 to PE. This lipidation step increases the electrophoretic mobility of LC3 (which can be measured by immunoblot analysis) and causes its redistribution from a diffuse cytosolic pattern to so-called autophagic puncta (which can be visualized by immunofluorescence or by tagging LC3 with green fluorescent protein).
Given the widespread functions of PE as a precursor of several biosynthetic pathways, there is high demand for this metabolite (Fig. 1). A common PE pool feeds into all major cellular PE-consuming pathways, thus resulting in competition for PE between pathways.4 As we have recently shown, this limitation can be overcome by genetic or pharmacological interventions. Overexpression of yeast phosphatidylserine decarboxylase 1 (Psd1), which catalyzes the step from phosphatidylserine to PE, increased the levels of intracellular PE. Similarly, external administration of ethanolamine (Etn), a precursor of PE (Fig. 1) can increase the abundance of intracellular PE.8 Supporting a crucial regulatory role for PE in autophagy, we observed that both external supply of PE and an increase in its internal generation similarly increased autophagic flux.8 Stimulation of autophagy by PE could efficiently protect cells from necrotic or apoptotic cell death.6,9 This is an important finding because unwarranted cell death may contribute to several age-related diseases including stroke, atherosclerosis, and cardiovascular disease, or neurodegenerative disorders such as Alzheimer's, Parkinson's, or Huntington's disease. Importantly, pharmacological Etn treatment extended the lifespan of yeast and fruit flies, as well as cultured mammalian cells,8 underlining the potential of Etn as a potent autophagy and longevity drug.
Figure 1.
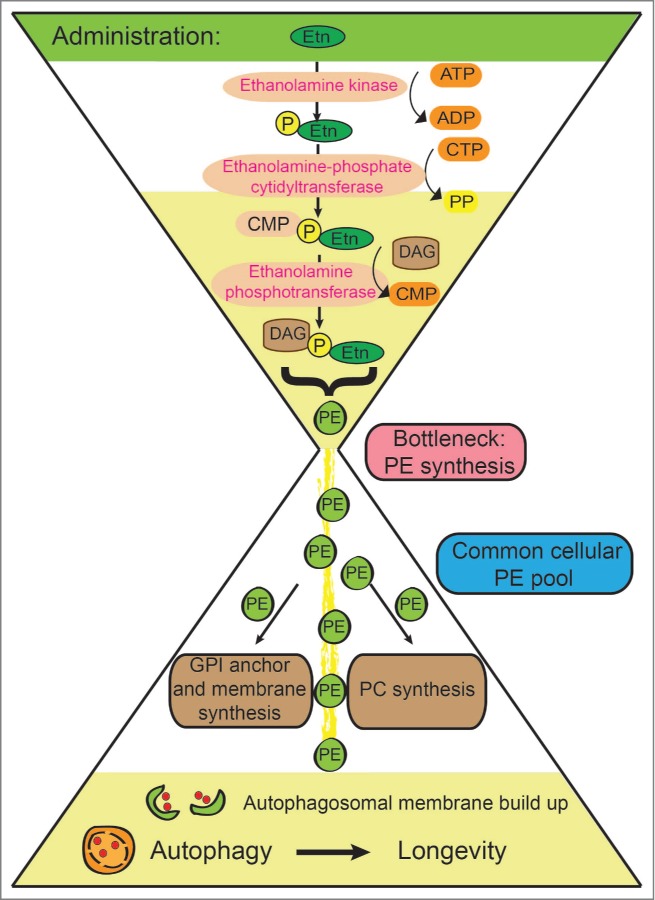
The ‘PE hourglass’. Simplified schematic of the conversion of ethanolamine (Etn) into phosphatidylethanolamine (PE) and its downstream products. Alternative pathways for PE production were omitted for clarity. PE synthesis is rate limiting because newly synthesized PE feeds into a common cellular PE pool, which is used for diverse cellular processes including glycosylphosphatidylinositol (GPI) anchor, membrane, and phosphatidylcholine (PC) synthesis as well as the generation of autophagic membranes. ADP, adenosine diphosphate; ATP, adenosine triphosphate; CMP, cytosine monophosphate; Etn, ethanolamine; CTP, cytosine triphosphate; DAG, diacylglycerol; P, phosphate; PE, phosphatidylethanolamine; PP, pyrophosphate.
Given its properties as a detergent and emulsifier, Etn is widely used in cosmetic, lifestyle, and household products such as hair colors, bleaches, washing powders, disinfectants, kitchen cleaners, shampoos, and soaps. Thus, its toxicologic profile as an external agent that may irritate the skin and mucosa is well studied. However, it remains to be determined whether Etn can be used internally within the scope of increasing organismal PE levels, thereby stimulating autophagy and possibly conferring health benefits.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Funding
This work has been supported by Fonds zur Förderung der Wissenschaftlichen Forschung in Österreich (FWF), projects SFB Lipotox F3005-B12, P23490-B12 and P24381-B20 to F.M. The authors gratefully acknowledge support from NAWI Graz and BioTechMed. GK is supported by the Ligue contre le Cancer (équipe labelisée); Agence National de la Recherche (ANR); Association pour la recherche sur le cancer (ARC); Cancéropôle Ile-de-France; Institut National du Cancer (INCa); Fondation Bettencourt-Schueller; Fondation de France; Fondation pour la Recherche Médicale (FRM); the European Commission (ArtForce); the European Research Council (ERC); the LabEx Immuno-Oncology; the SIRIC Stratified Oncology Cell DNA Repair and Tumor Immune Elimination (SOCRATE); the SIRIC Cancer Research and Personalized Medicine (CARPEM); and the Paris Alliance of Cancer Research Institutes (PACRI).
References
- 1.Bürgermeister M, Birner-Grünberger R, Nebauer R, Daum G. Contribution of different pathways to the supply of phosphatidylethanolamine and phosphatidylcholine to mitochondrial membranes of the yeast Saccharomyces cerevisiae. Biochim Biophys Acta 2004; 1686:161-8; PMID:15522832; http://dx.doi.org/ 10.1016/j.bbalip.2004.09.007 [DOI] [PubMed] [Google Scholar]
- 2.Vance JE. Thematic Review Series: Glycerolipids. Phosphatidylserine and phosphatidylethanolamine in mammalian cells: two metabolically related aminophospholipids. J Lipid Res 2008; 49:1377-87; PMID:18204094; http://dx.doi.org/ 10.1194/jlr.R700020-JLR200 [DOI] [PubMed] [Google Scholar]
- 3.Van den Brink-van der Laan E, Antoinette Killian J, de Kruijff B. Nonbilayer lipids affect peripheral and integral membrane proteins via changes in the lateral pressure profile. Biochim Biophys Acta BBA - Biomembr 2004; 1666:275-88; PMID:15519321; http://dx.doi.org/ 10.1016/j.bbamem.2004.06.010 [DOI] [PubMed] [Google Scholar]
- 4.Wilson-Zbinden C, Santos AX da S dos, Stoffel-Studer I, van der Vaart A, Hofmann K, Reggiori F, Riezman H, Kraft C, Peter M. Autophagy Competes for a Common Phosphatidylethanolamine Pool with Major Cellular PE-Consuming Pathways in Saccharomyces cerevisiae. Genetics 2014; genetics114169797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Feng Y, He D, Yao Z, Klionsky DJ. The machinery of macroautophagy. Cell Res 2014; 24:24-41; PMID:24366339; http://dx.doi.org/ 10.1038/cr.2013.168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Madeo F, Tavernarakis N, Kroemer G. Can autophagy promote longevity? Nat Cell Biol 2010; 12:842-6; PMID:20811357; http://dx.doi.org/ 10.1038/ncb0910-842 [DOI] [PubMed] [Google Scholar]
- 7.Ichimura Y, Kirisako T, Takao T, Satomi Y, Shimonishi Y, Ishihara N, Mizushima N, Tanida I, Kominami E, Ohsumi M, et al.. A ubiquitin-like system mediates protein lipidation. Nature 2000; 408:488-92; PMID:11100732; http://dx.doi.org/ 10.1038/35044114 [DOI] [PubMed] [Google Scholar]
- 8.Rockenfeller P, Koska M, Pietrocola F, Minois N, Knittelfelder O, Sica V, Franz J, Carmona-Gutierrez D, Kroemer G, Madeo F. Phosphatidylethanolamine positively regulates autophagy and longevity. Cell Death Differ [Internet] 2015; 22:499-508; [cited 2015 Jan 21]; Available from: http://www.nature.com/cdd/journal/vaop/ncurrent/full/cdd2014219a.html [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Eisenberg T, Knauer H, Schauer A, Büttner S, Ruckenstuhl C, Carmona-Gutierrez D, Ring J, Schroeder S, Magnes C, Antonacci L, et al.. Induction of autophagy by spermidine promotes longevity. Nat Cell Biol 2009; 11:1305-14; PMID:19801973; http://dx.doi.org/ 10.1038/ncb1975 [DOI] [PubMed] [Google Scholar]