

Abstract
Background
The parent-child relationship may be an important factor in the development of adolescent depressive and anxious symptoms. In adults, depressive symptoms relate to increased amygdala and attenuated prefrontal activation to maternal criticism. The current pilot study examined how depressive and anxiety symptoms in a high-risk adolescent population relate to neural responses to maternal feedback. Given previous research relating oxytocin to maternal behavior, we conducted exploratory analyses using oxytocin receptor (OXTR) genotype.
Methods
Eighteen females (ages 12–16) listened to maternal praise, neutral, and critical statements during functional magnetic resonance imaging. Participants completed the Mood and Feelings Questionnaire and the Screen for Child Anxiety Related Emotional Disorders. The OXTR single nucleotide polymorphism, rs53576, was genotyped. Linear mixed models were used to identify symptom or allele (GG, AA/AG) by condition (critical, neutral, praise) interaction effects on brain activation.
Results
Greater symptoms related to greater right amygdala activation for criticism and reduced activation to praise. For left amygdala, greater symptoms related to reduced activation to both conditions. Anxiety symptoms related to differences in superior medial PFC activation patterns. Parental OXTR AA/AG allele related to reduced activation to criticism and greater activation to praise within the right amygdala.
Conclusions
Results support a relationship between anxiety and depressive symptoms and prefrontal-amygdala responses to maternal feedback. The lateralization of amygdala findings suggests separate neural targets for interventions reducing reactivity to negative feedback or increasing salience of positive feedback. Exploratory analyses suggest that parents' OXTR genetic profile influences parent-child interactions and related adolescent brain responses.
Keywords: Prefrontal cortex, Amygdala, Oxytocin, Maternal behavior, Parent-child relations
Highlights
-
•
Adolescent brain responses to maternal praise and criticism were examined.
-
•
Anxiety related to prefrontal-amygdala response to both criticism and praise.
-
•
Exploratory results support a role for the OXTR gene in parent-child interactions.
1. Introduction
Findings from the National Comorbidity Study indicate that anxiety disorders are the most commonly diagnosed mental health conditions in adolescence, with almost 32% lifetime prevalence (Merikangas et al., 2010). An estimated 14% of adolescents will experience mood disorders such as depression and 20% will experience comorbidity between at least two classes of mental health disorders. Several risk factors have been identified for the development of adolescent mood and anxiety symptoms or disorders, including poverty, parental education, maternal mental health, distressed or separated parental relationships, and being female (Merikangas et al., 2010, Spence et al., 2002).
The quality of social relationships, and the parent-child relationship in particular, may be important contributors to mental health outcomes for those at high risk (Salzinger et al., 2002). For example, low parental communication or warmth, and parent-child hostility have been found to relate to greater depression and anxiety, lower self-esteem, and more suicide attempts (Low and Stocker, 2005, Moore et al., 2004). However, it must also be recognized that the relationship between parent-child interactions and child mental health may be bidirectional (Nelemans et al., 2014). Interestingly, maternal criticism was found to predict the onset of depression, independent of the effects of maternal depression (Burkhouse et al., 2012).
Neuroimaging research related to anxiety and depression has focused on a circuitry involving amygdala, striatal, and medial prefrontal cortex (PFC) regions, which are thought to be involved in emotion generation and threat reactivity, reward processing, and the monitoring and regulation of emotional responses, respectively (Casey et al., 2010, Kerestes et al., 2014). Most of the neuroimaging literature on depression and anxiety symptoms and disorders has focused on the processing or anticipation of threat-related or distressing stimuli (i.e., emotional face images). Considering the potential importance of the parent-child relationship, there has been an initial cadre of studies aiming to understand neural response patterns when processing parental criticism and its potential role in psychopathology. One healthy adolescent study found parental criticism to be associated with decreased activity in cognitive control (i.e., dlPFC, ACC) and social cognitive networks (temporoparietal junction, posterior cingulate cortex) as well as increased activity in affective networks, including lentiform nucleus and posterior insula (Lee et al., 2015). Studies investigating neural responses to peer rejection have reported depression or anxiety symptoms to relate to differences in amygdala, ACC, and PFC recruitment (Guyer et al., 2008a, Masten et al., 2011, Silk et al., 2014). One interesting study examined neural responses to videos of maternal affective behavior (aggressive, positive, and neutral) (Whittle et al., 2012). This study reported that adolescent depressive symptoms related to reduced rostral ACC response to positive affective behavior of their mother, as well as reduced striatal activity to positive affective behavior of either their mother or an unfamiliar mother. Studies with adults indicate that a history of depression may relate to reduced ACC and dlPFC activation to either parental criticism or praise and greater amygdala activation to criticism (Hooley et al., 2009, Hooley et al., 2005). However, there have been no studies investigating (a) how neural responses during parental feedback may relate to adolescent mood and anxiety symptoms or (b) anticipation of parental feedback, which may be particularly relevant for understanding anxiety (Grupe and Nitschke, 2013).
Parent and child genotype is an obvious contributing factor to parent-child interactions, child mental health, and associated neural responses. The single nucleotide polymorphism (SNP) rs53376, in the third intron of the oxytocin receptor gene (OXTR; A/G), may be particularly important for social and caregiving behavior, as well as mental health. Individuals with GG alleles (compared to AA or AG alleles) perform better at emotional and empathy recognition tasks, have lower stress reactivity in many contexts, and report higher levels of optimism and self-esteem and less depression (Rodrigues et al., 2009, Saphire-Bernstein et al., 2011). Further, mothers with GG genotypes have been found to display more sensitive parenting (Bakermans-Kranenburg and van Ijzendoorn, 2008) and greater heart rate responses to infant cries (Riem et al., 2011). However, there have been inconsistencies in the directionality of findings and some indication that these effects may be moderated by context, such as experience of child abuse, interparental conflict, or mother's mental state (Bradley et al., 2011, Michalska et al., 2014). One potential mechanism through which the OXTR gene may exert its effects is through the influence on amygdala structure and function. The GG allele has been associated with differences in amygdala volume, as well as increased amygdala activation and decreased amygdala-hypothalamus connectivity during emotional face processing (Furman et al., 2011, Tost et al., 2010). The OXTR genotype of an individual may contribute most directly to that individual's brain development and anatomy. However, it is also likely that parental genotype, by influencing maternal behavior, would in turn influence the emotional and brain development of the child.
The aims of the current pilot study were to extend previous research by (1) examining how depression and anxiety symptoms in high-risk, female adolescents relate to neural responses to maternal feedback and (2) probing both anticipation and processing of maternal feedback, as anticipation of distressing stimuli has been considered particularly important for understanding anxiety (Grupe and Nitschke, 2013). Further understanding of how depression and anxiety relate to neural responses to maternal feedback could provide guidance and potential neural targets for parent-child interventions. Finally, we explored whether parent or child OXTR allele impacts amygdala response or depressive and anxiety symptoms. Although underpowered to make definitive conclusions, these exploratory results provide direction for future larger studies investigating genotypic contributions to the relationship between maternal feedback and depression and anxiety.
2. Materials and methods
2.1. Participants
Eighteen 12–16 year old adolescent females were recruited from a larger study (N = 171) focused on resilience factors for adolescents sampled from urban areas of Tulsa, OK with high rates for teen pregnancy, violence exposure, and school dropout. See Table 1 for sample characteristics.
Table 1.
Sample descriptive statistics.
| Variable | M or % | SD |
|---|---|---|
| Age | 14.39 | 1.42 |
| Mother education level | ||
| Grade 10–11 | 5.6% | |
| High school graduate/GED | 27.8% | |
| Some college or trade school | 38.9% | |
| Four years of college | 27.8% | |
| Below poverty linea | 44.4% | |
| Ethnicity | ||
| European American | 22.2% | |
| African American | 33.3% | |
| Native American | 22.2% | |
| Other | 22.2% | |
| SCARED | 26.33 | 16.50 |
| MFQ | 12.39 | 8.69 |
| BAI | 12.28 | 14.44 |
| CES-D | 13.44 | 12.20 |
Median annual income for household.
All participants attended two sessions. Session 1 included self-report measures and recordings of mothers' feedback statements. Session 2, completed an average of one week after Session 1, included fMRI scanning concurrently with the maternal feedback paradigm. All participants provided assent, and mothers provided informed consent. Study procedures were approved by the Oklahoma State University Center for Health Sciences Institutional Review Board. Exclusion criteria included MRI contraindications (i.e., metal in body), pregnancy, left-handedness, psychiatric or other medications altering brain function, and significant medical conditions. Saliva samples for OXTR genotyping were obtained during a separate session, as part of the larger study.
2.2. Self-report measures
Youth symptoms of depression and anxiety were acquired using the child-report Child Mood and Feelings Questionnaire (MFQ, Angold et al., 1995) and the Screen for Child Anxiety Related Emotional Disorders (SCARED, Birmaher et al., 1997), respectively. Parent symptoms were assessed using the Beck Anxiety Inventory (BAI, Beck et al., 1988) and Center for Epidemiological Studies – Depression scale (CES-D, Radloff, 1977).
2.3. Maternal feedback challenge
The maternal feedback challenge (Fig. 1) was based on previous studies (Hooley et al., 2005, Hooley et al., 2009, Lee et al., 2015), but modified to an event-related design to allow modeling of anticipation. During Session 1, mothers were guided in generating 6 each of praise, neutral, and critical statements, to be based on topics previously discussed by the dyad. Mothers were recorded saying each statement. In Session 2, youth participants listened to these statements during six, 280 s fMRI runs. Nine recorded statements (standardized to 7 s) were presented within each run (3 each, praise, neutral, critical) in pseudorandom order (total 18 presentations for each statement type). In each trial, a 2 second cue (cyan, magenta, or yellow dots) signaled comment type, followed by a 4–8 second interstimulus interval (white dot), 7-second recorded comment, and 10–21 second intertrial interval (white dot). To encourage attention, 3–18 s into the interval (counterbalanced across trials) the white dot turned gray, to which participants were instructed to press a button. This cue remained gray until button press or the end of the intertrial interval. A bivariate 5 × 5 rating grid (Larsen et al., 2009) appeared pseudo-randomly during 1/3 of the intertrial intervals (for 7 s) to acquire emotional ratings for the recorded comments. The horizontal dimension assessed negative ratings, and the vertical assessed positive ratings (range of 0, “Not at all” to 5, “Extremely”). This same grid was used to obtain overall emotional ratings for each cue type post-scan. The cue colors signaling each comment type were counterbalanced across participants, who were informed about the association of each cue prior to the task.
Fig. 1.

Schematic of task. Participants viewed one of three colored dots signaling a subsequent praise, neutral, or critical comment, followed by a 4–8 second interstimulus interval (white dot), the 7 second comment recording, and a 10–21 second intertrial interval (white dot). A vigilance cue (gray dot) would be shown at some point during the intertrial, which would remain on the screen until a button press or the end of the intertrial interval. Ratings grids were presented during 1/3 of the intertrial intervals.
2.4. MRI acquisition and preprocessing
Imaging was performed using a Discovery MR750 3 Tesla MRI scanner (GE Healthcare, Milwaukee, Wisconsin) with an 8-channel receive-only head coil. Functional scans were acquired using gradient-recalled echo-planar imaging (EPI) sequences with sensitivity encoding (SENSE acceleration factor: R = 2 in the phase encoding direction; matrix size: 96 × 96, FOV/slice/gap = 240/3.5/0.5 mm, in-plane resolution: 2.5 × 2.5 mm2, sagittal plane: 35 slices, TR/TE = 2000/25 ms, flip angle: 90°, sampling bandwidth: 250 kHz, 140 volumes). EPI images were reconstructed into a 128 × 128 matrix and voxel size of 1.875 × 1.875 × 3.5 mm3. Structural MRI scans employed a T1-weighted Magnetization Prepared Rapid Gradient Echo (MPRAGE) imaging sequence with SENSE (acceleration factor: R = 2, scan time = 5 min 5 s; matrix size: 256 × 256, FOV/slice = 240/1.1 mm, in-plane resolution: 0.938 × 0.938 mm2, TR/TE = 5/1.95 ms, inversion and delayed times: TI/TD = 725/1400 ms, flip angle: 8°, sampling bandwidth: 31.25 kHz, 134 axial slices per volume).
Data were preprocessed and analyzed using Analysis of Functional NeuroImages (AFNI) software package (Cox, 1996) and R statistical package (www.r-project.org). Advanced Normalization Tools (ANTS; http://stnava.github.io/ANTs/) was used to optimize alignment of functional data to the TT_N27 T1-weighted template. Each scan was corrected for slice acquisition timing and co-registered to the first volume. The structural image was aligned to the first EPI image, resampled to EPI scan resolution, and warped to the TT_N27 template (resampled to 1.875 mm3) using ANTS. The EPI images were normalized to the template using the same warping parameters. EPI data were spatially smoothed with a 1.875 mm FWHM Gaussian kernel and scaled by the mean to estimate percent signal change (PSC). Time points with > 0.5 mm motion were censored from analyses. Individual time-series data were analyzed using a multiple regression model using regressors to quantify PSC for both cue (Cue-crit, Cue-neut, Cue-praise) and recording presentation periods for each statement type (Rec-crit, Rec-neut, Rec-praise). The following regressors of no interest were included: (1) motion parameters (roll, pitch, yaw, superior, left, and posterior; demeaned and derivative values), (2) linear trend to eliminate slow signal drifts, (3) interim periods between cues and recordings, and (4) periods that involved valence ratings.
2.5. OXTR genotyping
Saliva samples were taken from 16 youth and parent dyads. A Salimetrics modified PureLink Genomic DNA extraction method was used to isolate DNA from passive drool, following Nemoda et al. (2011). Samples were aliquoted and stored at − 20 °C. Taqman® SNP Genotyping Assays (Applied Biosystems/LifeTech) were employed to amplify and detect the two alleles (A/G) for the rs53576 SNP of the oxytocin receptor gene (OXTR). Only one youth and one parent (not of the same dyad) were homozygous for the A allele. Therefore, those homozygous for the A allele were combined into one group with those that were heterozygous (AG). This resulted in 10 GG youth, 8 GG parents and 6 AA/AG youth and 8 AA/AG parents.
2.6. Data analyses
2.6.1. Behavioral analyses
Correlations between self-report measures were examined using Spearman's rho (rs). Linear mixed effects (LME) analyses were conducted to examine main and interaction effects for symptom severity (SCARED or MFQ) and condition (crit, neut, praise) on both negative and positive valence ratings during the maternal feedback challenge. Results meeting p < 0.05 are described, but only those meeting Bonferroni correction are interpreted in the Discussion. With two LME analyses (positive and negative ratings) conducted for each measure (SCARED, MFQ), the adjusted p-threshold was 0.013 (0.05/4).
2.6.2. Neuroimaging analyses
fMRI PSC for both cues and recordings, across all condition types (neutral, praise, critical), were extracted from predefined probability maps of left and right basolateral amygdala (Amunts et al., 2005) and subjected to LME analysis. ROI analyses focused on the basolateral amygdala because it receives highly processed sensory information that is hypothesized to support the amygdala's role in social-emotional behavior, such as the encoding of threat (Davis and Whalen, 2001). Main and interaction effects for symptom severity (SCARED or MFQ) and condition (neutral, praise, critical) were examined using separate LMEs for cues and recordings (subject as random effects). Amygdala ROI results meeting p < 0.05 are described; only those meeting Bonferroni correction are interpreted. Bonferroni correction was applied for the analyses of cues and recordings separately, with each involving two analyses (SCARED, MFQ) for each amygdala ROI (left, right), resulting in an adjusted p-threshold of 0.013 (0.05/4).
Identical LMEs were conducted voxel-wise across the whole brain. Recent evidence demonstrates that traditional methods of implementing fMRI cluster size multiple comparison corrections fail to account for spatial autocorrelations, and thereby render cluster thresholds that do not adequately control false-positive inferences (Eklund et al., 2015). We therefore employed the spatial autocorrelation function (acf) option when using the AFNI programs 3dFWHMx to estimate intrinsic smoothness in the images and 3dClustSim to estimate the probability of false positive clusters. This option allows for non-Gaussian models and spatial autocorrelation functions, and calculates moments of differences to a larger radius. Clusters were considered significant at a voxel-wise p < 0.005, corrected for multiple comparisons using the spatial autocorrelation function procedures noted above, resulting in a minimum cluster threshold of 48 voxels. Results from whole-brain analyses are summarized in the main text and detailed in the Appendix.
2.6.3. Exploratory OXTR analyses
LME analyses were conducted to examine main effects of youth and parent OXTR allele on adolescent symptom severity (SCARED, MFQ) and allele by condition interaction effects on valence ratings and amygdala PSC. Bonferroni correction was applied for the analyses of valence ratings (positive, negative) and amygdala PSC (left, right) separately, with each involving two analyses (youth, parent allele) for each of the identified measures, resulting in an adjusted p-threshold of 0.013 (0.05/4).
Including age as a covariate did not influence any of the amygdala ROI, OXTR, or behavioral results and thus, results are reported without age in the models.
3. Results
3.1. Behavioral analyses
Youth SCARED and MFQ scores (rs = 0.85, p < 0.001) were highly correlated as were parent BAI and CES-D scores (rs = 0.92, p < 0.001), indicating the questionnaires related to depression and anxiety were indexing highly overlapping constructs. There was one outlier in MFQ score, with a score of 40 (> 3 SDs above the mean). Thus, all analyses involving MFQ were conducted with this participant removed (for a total N = 17) and SCARED score was used as the primary self-report variable of interest (and the only one examined in whole-brain analyses). SCARED score correlated with parent BAI (rs = 0.49, p = 0.037), though this finding did not withstand Bonferroni correction (p < 0.013). There were no correlations between SCARED (rs = 0.30, p = 0.219) or MFQ (rs = 0.31, p = 0.213) scores and parent CES-D.
LME analyses revealed condition (crit, neut, praise) main effects on both positive (Cue: F(2,30) = 34.41, p < 0.001; Rec: F(2,30) = 56.54, p < 0.001) and negative ratings (Cue: F(2,30) = 32.71, p < 0.001; Rec: F(2,30) = 32.54, p < 0.001) during the maternal feedback challenge, which were in the expected direction (see Inline Supplementary Table S1 and supplemental results). There was a main effect of MFQ (F(1,15) = 8.01, p = 0.013) but not SCARED (F(1,15) = 3.87, p = 0.07) on positive ratings to the recordings, with greater symptom severity relating to less positive ratings. These main effects were qualified by MFQ by condition (F(2,30) = 5.61, p = 0.009) and SCARED by condition (F(2,30) = 5.27, p = 0.010) interaction effects on positive ratings to the cue, with greater symptom severity relating to less positive ratings of neutral cues. These findings survived Bonferroni correction (p < 0.013). There were no main or interaction effects of symptom severity measures on negative ratings (all ps > 0.10).
Inline Supplementary Table S1.
Table S1.
Descriptive statistics for positive and negative ratings of cues and recordings.
| Variable | M | SD |
|---|---|---|
| Positive Ratings – Cue | ||
| Neutral | 1.78 | 1.35 |
| Praise | 3.33 | 0.59 |
| Critical | 1.00 | 0.84 |
| Positive Ratings – Recording | ||
| Neutral | 2.28 | 1.08 |
| Praise | 3.22 | 0.65 |
| Critical | 1.27 | 0.93 |
| Negative Ratings – Cue | ||
| Neutral | .089 | 1.02 |
| Praise | 0.28 | 0.57 |
| Critical | 2.33 | 0.76 |
| Negative Ratings – Recording | ||
| Neutral | 0.57 | 0.57 |
| Praise | 0.57 | 0.63 |
| Critical | 1.97 | 0.71 |
Note: M = mean, SD = standard deviation.
3.2. Amygdala ROI analyses
For the cue, there was a main effect of condition on right amygdala activation (F(2,32) = 4.49, p = 0.019), with PSC greatest for critical and least for praise conditions (see Inline Supplementary Fig. S1). This finding was qualified by interactions (SCARED: F(2,32) = 9.80, p < 0.001; MFQ: F(2,32) = 9.94, p < 0.001), with greater anxiety or depressive symptoms relating to greater activation to critical and less to praise conditions (see Fig. 2 for SCARED results). The interaction effects survived Bonferroni correction (p < 0.013). The described results remained significant when either age or parental symptom severity (BAI or MFQ) were includes as covariates.
Fig. S1.

Average percent signal change within the basolateral amygdala regions of interest to each task condition. Statistical results for amygdala percent signal change are described in the main text, including both condition main effects as well as condition by symptom (anxiety, depression) interaction effects. Error bars represent standard error.
Fig. 2.

Relationships between anxiety symptoms and brain activation to cues and recordings. (A) Percent signal change (PSC) extracted from basolateral amygdala regions of interest. A1 demonstrates the Screen for Child Anxiety Related Emotional Disorders (SCARED) by condition interaction effect for the right amygdala response to cues. A2 and A3 demonstrate the SCARED by condition interaction effect for the right and left amygdala response to recordings. (B) Prefrontal cortex (PFC) and amygdala clusters identified via whole-brain analyses. B1 and B2 demonstrate interaction effects for medial PFC clusters to cues and recordings, respectively. B3 demonstrates interaction effects for left amygdala to recordings.
For the recording, there was a main effect of condition on left amygdala activation (F(2,32) = 3.35, p = 0.050), with PSC greatest for critical and least for praise conditions. There was also an interaction for the left amygdala (SCARED: F(2,32) = 9.74, p < 0.001; MFQ: F(2,32) = 8.10, p = 0.001), with greater anxiety or depressive symptoms relating to greater activation to neutral and less activation to praise and critical conditions. There was an interaction effect for anxiety (SCARED: F(2,32) = 6.01, p < 0.001) but not depression (MFQ: F(2,32) = 2.91, p = 0.070) on right amygdala activation during the recording. Similar to the cue, greater anxiety symptoms related to greater activation to critical and less to praise conditions. All interaction effect results survived Bonferroni correction (p < 0.013). The described results remained significant when either age or parental symptom severity (BAI or MFQ) were includes as covariates.
Inline Supplementary Figure S1.
3.3. Whole-brain analyses
For the cue, SCARED by condition interactions were identified within the superior medial PFC (see Fig. 2; BA 10, 79 voxels; F(2,32) = 11.51, x,y,z = 12,62,2; greater symptoms related to greater activation to praise and relatively less activation to neutral and critical) and postcentral gyrus (BA 3, 55 voxels, F(2,32) = 8.36, x,y,z = − 57,− 10,22; greater symptoms related to reduced activation to praise). For the recordings, interactions were also identified within the superior medial PFC (BA 10; 103 voxels, F(2,32) = 7.80, x,y,z = − 5,54,0; greater symptoms related to greater activation to neutral and critical and less activation to praise) and the left amygdala (see Fig. 2; 77 voxels; F(2,32) = 8.08, x,y,z = − 24,− 7,− 16; greater symptoms related to greater activation to critical and less activation to praise), as well as the middle temporal gyrus (BA 21, 89 voxels, F(2,32) = 8.78, x,y,z = − 61,− 16,− 7; greater symptoms related to greater activation to neutral and less activation to praise) and culmen of the cerebellum (59 voxels, F(2,32) = 9.66, x,y,z = 0,− 46,0; greater symptoms relating to greater activation to critical and neutral). There were no clusters identified for condition main effects that survived the specified threshold used for whole-brain analyses.
3.4. OXTR gene analyses
There was an interaction between parent OXTR allele and condition for negative ratings of the recordings (F(2,28) = 4.05, p = 0.029; see Fig. 3). Post-hoc t-tests (see supplementary results) indicated that this effect was driven by youth with AA/AG parents providing more negative ratings of critical statements. There was no interaction effect between parent OXTR allele and condition for positive ratings of the recordings (F(2,28) = 1.524, p = 0.235). There was an interaction effect for both right (F(2,28) = 7.23, p = 0.003) and left amygdala (F(2,28) = 3.36, p = 0.049) activation to the recordings (see Fig. 3). Only the right amygdala activation to the recording survived Bonferroni correction (p < 0.013). Post-hoc t-tests (see Supplementary results) indicated that, for the right amygdala, this effect was most likely driven by youth with AA/AG parents exhibiting greater activation to praise and reduced activation for critical statements. Parent OXTR allele did not relate to amygdala activation to the cue (left: F(2,28) = 0.157, p = 0.855; right: F(2,28) = 0.961, p = 0.395).
Fig. 3.
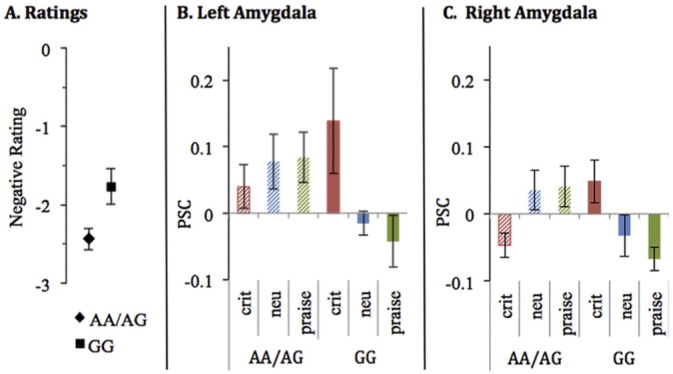
Relationships between OXTR alleles and (A) negative ratings of maternal criticism. (B) left amygdala, and (C) right amygdala.
There were no significant findings for relationships between youth OXTR allele and ratings of the recordings (negative ratings: F(2,28) = 0.85, p = 0.434; positive ratings: F(2,28) = 0.437, p = 0.650), amygdala activity to the cues (right: interaction, F(2,28) = 0.191, p = 0.827; left: interaction, F(2,28) = 0.332, p = 0.720), or amygdala activity to the recordings (right: F(2,28) = 2.215, p = 0.128; left, interaction: F(2,28) = 0.297, p = 0.746). Neither parent nor youth OXTR genetic profile was found to relate to self-reported adolescent anxiety (youth OXTR: F(1,14) = 0.037, p = 0.850; parent OXTR: F(1,14) = 3.032, p = 0.104) or depression ratings (youth OXTR: F(1,14) = 1.437, p = 0.251; parent OXTR: F(1,14) = 0.868, p = 0.367).
4. Discussion
The results of this pilot study indicate that anxiety and depressive symptoms were associated with differences in activation within amygdala and superior medial PFC regions to maternal feedback. In addition, parent OXTR AA/AG allele was associated with differences in right amygdala activation to maternal feedback.
Within the right amygdala, greater depression or anxiety symptoms related to greater activation to critical and reduced activation to praise (for cues and recordings). This finding is consistent with previous research showing depression and/or anxiety relates to increased limbic responsivity during both anticipation and processing of negatively valenced or threatening stimuli (Beesdo et al., 2009, Guyer et al., 2008b, Monk et al., 2008). However, within the left amygdala, increased depression or anxiety symptoms related to greater activation to neutral recordings and less activation to both critical and praise recordings. The left amygdala has been implicated most consistently in youth studies of depression (Kerestes et al., 2014), though often it is reported to exhibit increased activation to negative stimuli (i.e., emotional faces). Previous research supports lateralization of function, in which the right amygdala is more associated with processing negative emotion while the left is associated with processing both pleasant and unpleasant emotions (Baas et al., 2004, Lanteaume et al., 2007). With increased mood symptoms, adolescents in the current study seem to have a more valence-specific response in the right amygdala but a blunted response to processing general salience in the left amygdala. This suggests that (a) dampening right amygdala response to maternal criticism or other distressing social interactions and/or (b) enhancing left amygdala activation to praise or other positive emotional stimuli could be potential targets for interventions. The first may be served via interventions that reduce negative parent-child interactions or via cognitive and exposure-based protocols that reduce negative affective responses to criticism. The second may be targeted by increasing positive parent-child interactions or via interventions targeting positive affect (Sin and Lyubomirsky, 2009). In the current study, the differences in amygdala activity were most likely not due to differences in how adolescents with more severe symptoms were perceiving their mother's feedback (as symptoms did not relate to negative or positive ratings of the recordings). Perhaps one treatment target could be increased awareness of underlying neural processes. Notably, there has been recent research targeting adult depression using fMRI neurofeedback to enhance left amygdala response to positive memories (Young et al., 2014).
Whole-brain analyses indicated that anxiety symptoms related to differences in activation within superior medial PFC (to both the cue and recordings). Previous studies have reported that adults with a history of depression exhibit decreased prefrontal activation during maternal criticism, though these reports were of ACC or dlPFC regions (Hooley et al., 2005, Hooley et al., 2009). The recording (as opposed to anticipation) phase in the current paradigm was most similar to the maternal feedback paradigms used in previous studies. During this phase, anxiety symptoms related to less superior medial PFC during the praise condition. This is somewhat consistent with the previous study by Whittle and colleagues (Whittle et al., 2012) that identified relationships between adolescent depressive symptoms and neural responses (rostral ACC; putamen) to positive (but not aggressive) maternal affective behavior. This suggests that, at least for adolescents, mood/anxiety difficulties may relate to how they are monitoring and regulating responses to positive information and feedback (Forbes et al., 2006, Forbes et al., 2010, Silk et al., 2012). The findings of decreased superior medial PFC responses could reflect either reduced cognitive appraisal or self-relevant processing of these praise comments (Qin and Northoff, 2011, Seeley et al., 2007). Interestingly, the directionality of findings within the superior medial PFC was opposite during the cue phase of the paradigm, suggesting that with greater adolescent anxiety may come greater appraisal or self-relevant processing when anticipating praise but then less during the praise feedback itself. This provides further evidence that the superior medial PFC may be important to consider in the relationship between parent-adolescent interactions and adolescent mental health.
OXTR genotype analyses revealed that parental AA/AG, compared to GG, genotypes were associated with greater activation to praise and reduced activation to criticism within the right amygdala (as well as differences in left amygdala activation and negative ratings of critical statements, which did not survive Bonferroni correction). Given the sample size, this should be considered an exploratory, preliminary finding. However, this suggests that the OXTR genotype of the mother may contribute to her maternal behavior, which in turn influences the parent-child relationship and the child's neural response to feedback. The AA genotype has been associated with less sensitive parenting in some studies (Bakermans-Kranenburg and van Ijzendoorn, 2008). Thus, it is plausible that reduced sensitivity of the parent to the child's distress may result in an adaptive change in salience processing, for example within the amygdala. This is somewhat consistent with previous findings that the AA/AG allele is associated with decreased amygdala activity during emotional face processing (Tost et al., 2010); however, we found that parental allele, rather than the allele of the adolescent being scanned, related to amygdala activity. In addition to the direct passage of genes between parent and offspring, this represents another plausible way in which parental DNA may impact child mental health.
Study limitations included a relatively small sample size, limiting our ability to interpret genetic findings and assess more complex mediation or moderation relationships between variables. Although we purposely included participants from various ethnic backgrounds, the small sample size limits generalizability to populations with different ethnic or cultural composition than our current sample. The current study focused on female adolescents due to their increased risk of experiencing anxiety and depression, limiting generalizability to male populations. While validated self-report measures were used to assess depression and anxiety symptoms, no clinical diagnostic assessment was completed. Lastly, the current study was cross-sectional and thus precluded determination of causal relationships or determination of whether identified differences represent risk or resiliency factors.
5. Conclusions
The current pilot investigation represents the first study to examine how anxiety and depressive symptoms in adolescents may relate to neural responses to maternal feedback. Greater anxiety and depressive symptoms related to reduced left amygdala activation to all salient feedback (criticism and praise) and increased right amygdala activation specifically to criticism. These results may be useful in considering potential neural targets for behavioral interventions targeting either negative or positive affect and parent-child interactions. Exploratory findings provide rationale for future, larger studies to investigate the contribution of parent and child OXTR genetic profiles in the development of maladaptive parent-child interactions.
Acknowledgments
This research was supported by the Oklahoma Center for the Advancement of Science and Technology (OCAST; HR11-130) and the William K. Warren Foundation.
Footnotes
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.nicl.2016.03.009.
Appendix A. Supplementary data
Supplementary material
References
- Amunts K., Kedo O., Kindler M., Pieperhoff P., Mohlberg H., Shah N.J., Habel U., Schneider F., Zilles K. Cytoarchitectonic mapping of the human amygdala, hippocampal region and entorhinal cortex: intersubject variability and probability maps. Anat. Embryol. (Berl) 2005;210:343–352. doi: 10.1007/s00429-005-0025-5. [DOI] [PubMed] [Google Scholar]
- Angold A., Costello E.J., Messer S.C., Pickles A., Winder F., Silver D. Development of a short questionnaire for use in epidemiological studies of depression in children and adolescents. Int. J. Methods Psychiatr. Res. 1995:237–249. [Google Scholar]
- Baas D., Aleman A., Kahn R.S. Lateralization of amygdala activation: a systematic review of functional neuroimaging studies. Brain Res. Brain Res. Rev. 2004;45:96–103. doi: 10.1016/j.brainresrev.2004.02.004. [DOI] [PubMed] [Google Scholar]
- Bakermans-Kranenburg M.J., van Ijzendoorn M.H. Oxytocin receptor (OXTR) and serotonin transporter (5-HTT) genes associated with observed parenting. Soc. Cogn. Affect. Neurosci. 2008;3:128–134. doi: 10.1093/scan/nsn004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck A.T., Epstein N., Brown G., Steer R.A. An inventory for measuring clinical anxiety: psychometric properties. J. Consult. Clin. Psychol. 1988;56:893–897. doi: 10.1037//0022-006x.56.6.893. [DOI] [PubMed] [Google Scholar]
- Beesdo K., Lau J.Y., Guyer A.E., McClure-Tone E.B., Monk C.S., Nelson E.E., Fromm S.J., Goldwin M.A., Wittchen H.U., Leibenluft E., Ernst M., Pine D.S. Common and distinct amygdala-function perturbations in depressed vs anxious adolescents. Arch. Gen. Psychiatry. 2009;66:275–285. doi: 10.1001/archgenpsychiatry.2008.545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Birmaher B., Khetarpal S., Brent D., Cully M., Balach L., Kaufman J., Neer S.M. The Screen for Child Anxiety Related Emotional Disorders (SCARED): scale construction and psychometric characteristics. J. Am. Acad. Child Adolesc. Psychiatry. 1997;36:545–553. doi: 10.1097/00004583-199704000-00018. [DOI] [PubMed] [Google Scholar]
- Bradley B., Westen D., Mercer K.B., Binder E.B., Jovanovic T., Crain D., Wingo A., Heim C. Association between childhood maltreatment and adult emotional dysregulation in a low-income, urban, African American sample: moderation by oxytocin receptor gene. Dev. Psychopathol. 2011;23:439–452. doi: 10.1017/S0954579411000162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burkhouse K.L., Uhrlass D.J., Stone L.B., Knopik V.S., Gibb B.E. Expressed emotion-criticism and risk of depression onset in children. J. Clin. Child Adolesc. Psychol. 2012;41:771–777. doi: 10.1080/15374416.2012.703122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey B.J., Jones R.M., Levita L., Libby V., Pattwell S.S., Ruberry E.J., Soliman F., Somerville L.H. The storm and stress of adolescence: insights from human imaging and mouse genetics. Dev. Psychobiol. 2010;52:225–235. doi: 10.1002/dev.20447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox R.W. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Comput. Biomed. Res. 1996;29:162–173. doi: 10.1006/cbmr.1996.0014. [DOI] [PubMed] [Google Scholar]
- Davis M., Whalen P.J. The amygdala: vigilance and emotion. Mol. Psychiatry. 2001;6:13–34. doi: 10.1038/sj.mp.4000812. [DOI] [PubMed] [Google Scholar]
- Eklund A., Nichols T., Knutsson H. Can parametric statistical methods be trusted for fMRI based group studies? 2015. (arXiv preprint arXiv:1511.01863.)
- Forbes E.E., Christopher May J., Siegle G.J., Ladouceur C.D., Ryan N.D., Carter C.S., Birmaher B., Axelson D.A., Dahl R.E. Reward-related decision-making in pediatric major depressive disorder: an fMRI study. J. Child Psychol. Psychiatry. 2006;47:1031–1040. doi: 10.1111/j.1469-7610.2006.01673.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forbes E.E., Olino T.M., Ryan N.D., Birmaher B., Axelson D., Moyles D.L., Dahl R.E. Reward-related brain function as a predictor of treatment response in adolescents with major depressive disorder. Cogn. Affect. Behav. Neurosci. 2010;10:107–118. doi: 10.3758/CABN.10.1.107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furman D.J., Chen M.C., Gotlib I.H. Variant in oxytocin receptor gene is associated with amygdala volume. Psychoneuroendocrinology. 2011;36:891–897. doi: 10.1016/j.psyneuen.2010.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grupe D.W., Nitschke J.B. Uncertainty and anticipation in anxiety: an integrated neurobiological and psychological perspective. Nat. Rev. Neurosci. 2013;14:488–501. doi: 10.1038/nrn3524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guyer A.E., Lau J.Y., McClure-Tone E.B., Parrish J., Shiffrin N.D., Reynolds R.C., Chen G., Blair R.J., Leibenluft E., Fox N.A., Ernst M., Pine D.S., Nelson E.E. Amygdala and ventrolateral prefrontal cortex function during anticipated peer evaluation in pediatric social anxiety. Arch. Gen. Psychiatry. 2008;65:1303–1312. doi: 10.1001/archpsyc.65.11.1303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guyer A.E., Monk C.S., McClure-Tone E.B., Nelson E.E., Roberson-Nay R., Adler A.D., Fromm S.J., Leibenluft E., Pine D.S., Ernst M. A developmental examination of amygdala response to facial expressions. J. Cogn. Neurosci. 2008;20:1565–1582. doi: 10.1162/jocn.2008.20114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hooley J.M., Gruber S.A., Scott L.A., Hiller J.B., Yurgelun-Todd D.A. Activation in dorsolateral prefrontal cortex in response to maternal criticism and praise in recovered depressed and healthy control participants. Biol. Psychiatry. 2005;57:809–812. doi: 10.1016/j.biopsych.2005.01.012. [DOI] [PubMed] [Google Scholar]
- Hooley J.M., Gruber S.A., Parker H.A., Guillaumot J., Rogowska J., Yurgelun-Todd D.A. Cortico-limbic response to personally challenging emotional stimuli after complete recovery from depression. Psychiatry Res. 2009;172:83–91. doi: 10.1016/j.pscychresns.2009.02.001. [DOI] [PubMed] [Google Scholar]
- Kerestes R., Davey C.G., Stephanou K., Whittle S., Harrison B.J. Functional brain imaging studies of youth depression: a systematic review. Neuroimage Clin. 2014;4:209–231. doi: 10.1016/j.nicl.2013.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lanteaume L., Khalfa S., Regis J., Marquis P., Chauvel P., Bartolomei F. Emotion induction after direct intracerebral stimulations of human amygdala. Cereb. Cortex. 2007;17:1307–1313. doi: 10.1093/cercor/bhl041. [DOI] [PubMed] [Google Scholar]
- Larsen J.T., Norris C.J., McGraw A.P., Hawkley L.C., Cacioppo J.T. The evaluative space grid: a single-item measure of positivity and negativity. Cognition and Emotion. 2009;23:453–480. [Google Scholar]
- Lee K.H., Siegle G.J., Dahl R.E., Hooley J.M., Silk J.S. Neural responses to maternal criticism in healthy youth. Soc. Cogn. Affect. Neurosci. 2015;10:902–912. doi: 10.1093/scan/nsu133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Low S.M., Stocker C. Family functioning and children's adjustment: associations among parents' depressed mood, marital hostility, parent-child hostility, and children's adjustment. J. Fam. Psychol. 2005;19:394–403. doi: 10.1037/0893-3200.19.3.394. [DOI] [PubMed] [Google Scholar]
- Masten C.L., Eisenberger N.I., Borofsky L.A., McNealy K., Pfeifer J.H., Dapretto M. Subgenual anterior cingulate responses to peer rejection: a marker of adolescents' risk for depression. Dev. Psychopathol. 2011;23:283–292. doi: 10.1017/S0954579410000799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merikangas K.R., He J.P., Burstein M., Swanson S.A., Avenevoli S., Cui L., Benjet C., Georgiades K., Swendsen J. Lifetime prevalence of mental disorders in U.S. adolescents: results from the National Comorbidity Survey Replication–Adolescent Supplement (NCS-A) J. Am. Acad. Child Adolesc. Psychiatry. 2010;49:980–989. doi: 10.1016/j.jaac.2010.05.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michalska K.J., Decety J., Liu C., Chen Q., Martz M.E., Jacob S., Hipwell A.E., Lee S.S., Chronis-Tuscano A., Waldman I.D., Lahey B.B. Genetic imaging of the association of oxytocin receptor gene (OXTR) polymorphisms with positive maternal parenting. Front. Behav. Neurosci. 2014;8:21. doi: 10.3389/fnbeh.2014.00021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Monk C.S., Telzer E.H., Mogg K., Bradley B.P., Mai X., Louro H.M., Chen G., McClure-Tone E.B., Ernst M., Pine D.S. Amygdala and ventrolateral prefrontal cortex activation to masked angry faces in children and adolescents with generalized anxiety disorder. Arch. Gen. Psychiatry. 2008;65:568–576. doi: 10.1001/archpsyc.65.5.568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore P.S., Whaley S.E., Sigman M. Interactions between mothers and children: impacts of maternal and child anxiety. J. Abnorm. Psychol. 2004;113:471–476. doi: 10.1037/0021-843X.113.3.471. [DOI] [PubMed] [Google Scholar]
- Nelemans S.A., Hale W.W., 3rd, Branje S.J., Hawk S.T., Meeus W.H. Maternal criticism and adolescent depressive and generalized anxiety disorder symptoms: a 6-year longitudinal community study. J. Abnorm. Child Psychol. 2014;42:755–766. doi: 10.1007/s10802-013-9817-x. [DOI] [PubMed] [Google Scholar]
- Nemoda Z., Horvat-Gordon M., Fortunato C.K., Beltzer E.K., Scholl J.L., Granger D.A. Assessing genetic polymorphisms using DNA extracted from cells present in saliva samples. BMC Med. Res. Methodol. 2011;11:170. doi: 10.1186/1471-2288-11-170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin P., Northoff G. How is our self related to midline regions and the default-mode network? NeuroImage. 2011;57:1221–1233. doi: 10.1016/j.neuroimage.2011.05.028. [DOI] [PubMed] [Google Scholar]
- Radloff L.S. The CES-D Scale: a self-report depression scale for research in the general population. Appl. Psychol. Meas. 1977;1:385–401. [Google Scholar]
- Riem M.M., Pieper S., Out D., Bakermans-Kranenburg M.J., van Ijzendoorn M.H. Oxytocin receptor gene and depressive symptoms associated with physiological reactivity to infant crying. Soc. Cogn. Affect. Neurosci. 2011;6:294–300. doi: 10.1093/scan/nsq035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodrigues S.M., Saslow L.R., Garcia N., John O.P., Keltner D. Oxytocin receptor genetic variation relates to empathy and stress reactivity in humans. Proc. Natl. Acad. Sci. U. S. A. 2009;106:21437–21441. doi: 10.1073/pnas.0909579106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salzinger S., Feldman R.S., Stockhammer T., Hood J. An ecological framework for understanding risk for exposure to community violence and the effects of exposure on children and adolescents. Aggress. Violent Behav. 2002;7:423–451. [Google Scholar]
- Saphire-Bernstein S., Way B.M., Kim H.S., Sherman D.K., Taylor S.E. Oxytocin receptor gene (OXTR) is related to psychological resources. Proc. Natl. Acad. Sci. U. S. A. 2011;108:15118–15122. doi: 10.1073/pnas.1113137108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seeley W.W., Menon V., Schatzberg A.F., Keller J., Glover G.H., Kenna H., Reiss A.L., Greicius M.D. Dissociable intrinsic connectivity networks for salience processing and executive control. J. Neurosci. 2007;27:2349–2356. doi: 10.1523/JNEUROSCI.5587-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silk J.S., Davis S., McMakin D.L., Dahl R.E., Forbes E.E. Why do anxious children become depressed teenagers? The role of social evaluative threat and reward processing. Psychol. Med. 2012;42:2095–2107. doi: 10.1017/S0033291712000207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silk J.S., Siegle G.J., Lee K.H., Nelson E.E., Stroud L.R., Dahl R.E. Increased neural response to peer rejection associated with adolescent depression and pubertal development. Soc. Cogn. Affect. Neurosci. 2014;9:1798–1807. doi: 10.1093/scan/nst175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sin N.L., Lyubomirsky S. Enhancing well-being and alleviating depressive symptoms with positive psychology interventions: a practice-friendly meta-analysis. J. Clin. Psychol. 2009;65:467–487. doi: 10.1002/jclp.20593. [DOI] [PubMed] [Google Scholar]
- Spence S.H., Najman J.M., Bor W., O'Callaghan M.J., Williams G.M. Maternal anxiety and depression, poverty and marital relationship factors during early childhood as predictors of anxiety and depressive symptoms in adolescence. J. Child Psychol. Psychiatry. 2002;43:457–469. doi: 10.1111/1469-7610.00037. [DOI] [PubMed] [Google Scholar]
- Tost H., Kolachana B., Hakimi S., Lemaitre H., Verchinski B.A., Mattay V.S., Weinberger D.R., Meyer-Lindenberg A. A common allele in the oxytocin receptor gene (OXTR) impacts prosocial temperament and human hypothalamic-limbic structure and function. Proc. Natl. Acad. Sci. U. S. A. 2010;107:13936–13941. doi: 10.1073/pnas.1003296107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whittle S., Yucel M., Forbes E.E., Davey C.G., Harding I.H., Sheeber L., Yap M.B., Allen N.B. Adolescents' depressive symptoms moderate neural responses to their mothers' positive behavior. Soc. Cogn. Affect. Neurosci. 2012;7:23–34. doi: 10.1093/scan/nsr049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Young K.D., Zotev V., Phillips R., Misaki M., Yuan H., Drevets W.C., Bodurka J. Real-time FMRI neurofeedback training of amygdala activity in patients with major depressive disorder. PLoS One. 2014;9:e88785. doi: 10.1371/journal.pone.0088785. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material