

The developmental genetics of biological robustness (Viewpoint)
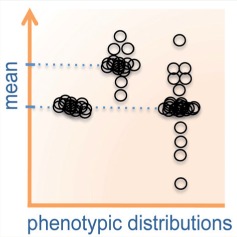
Biological systems face constant perturbations, yet often manage to produce invariable developmental phenotypes. Developmental robustness is a property observed across phyla and recent work in yeast, animals and plants has just started revealing its genetic basis. Mestek Boukhibar and Barkoulas (pp. 699–707) highlight aspects related to the definition of developmental robustness and discuss some underlying mechanisms. They piece together examples from different systems to advocate the importance of studying the phenotypic variance, rather than averages of developmental traits, which is commonplace in biology.
Obdiplostemony: the occurrence of a transitional stage linking robust flower configurations (Viewpoint)

Obdiplostemony, the two-whorled stamen configuration with outer stamens in alternisepalous position, has been a controversial condition for a long time. Ronse De Craene and Bull-Hereñu (pp. 709–724) review the definition and distribution of obdiplostemony, and examine its contribution to floral diversification and species evolution. Primary and secondary obdiplostemony, with at least five developmental pathways, are recognized. Obdiplostemony is instrumental in the transition to haplostemony by a weakening of the petal sector in the flower. However, in several cases this trend is reversed by a weakening of the antesepalous stamens leading to obhaplostemony. A broad definition of obdiplostemony is preferred, comprising flowers with petal loss, androecia where one stamen whorl is staminodial or incomplete, and anisomerous gynoecia. Obdiplostemony illustrates how developmental flexibility may lead to a high divergence of floral morphologies.
The significance of developmental robustness for species diversity (Viewpoint)

Why some taxa are more species-rich than others still represents a major conundrum in biology. Ecological opportunities and evolutionary innovations are considered main factors that drive biodiversity, but are sometimes insufficient to explain current patterns of species richness. In their viewpoint article, Melzer and Theißen (pp. 725–732) argue that developmental robustness might be an underappreciated factor promoting biodiversity. They hypothesize that only the robust integration of evolutionary innovations enables organisms to seize new ecological opportunities, which may in turn foster the origin of new innovations. Developmental robustness may thus be an important factor for an increase in morphological complexity and biodiversity.
Developmental mechanisms underlying variable, invariant and plastic phenotypes (Review)

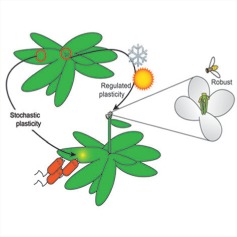
Discussions of phenotypic robustness often consider scenarios where invariant phenotypes are optimal and assume that developmental mechanisms have evolved to buffer phenotypes against stochastic and environmental perturbations. However, plastic plant phenotypes that vary between environments or variable phenotypes that vary stochastically within an environment are also likely to be advantageous in some scenarios. Abley et al. (pp. 733–748) review work from microbes, animals and plants that uncovers mechanisms underlying invariant, variable and plastic phenotypes. They discuss evidence that specific types of genetic networks underlie each class of phenotype and can confer a selective advantage in particular environments.
Development and evolution of extremely complex flowers (Review)

The two very species-rich families Apocynaceae and Orchidaceae contain the most complex flowers in the angiosperms. Although they are not related they show a number of amazing convergent similarities in flower structure. Endress (pp. 749–767) reviews and compares development and evolution of this complexity in both families. It has arisen by progressive synorganization of the basic floral organs, especially androecium and gynoecium, and partly also by the emergence of novel floral parts. As a result the flowers look puzzling and the basic organs can no longer be easily recognized at first sight.
Genetic assimilation and the evolution of novelty (Review)

Most, if not all, organisms possess the ability to alter their phenotype in direct response to changes in their environment, a phenomenon known as phenotypic plasticity. Selection can break this environmental sensitivity, however, and cause a formerly environmentally induced trait to evolve to become fixed through a process called genetic assimilation. Characterizing the proximate mechanisms that underlie genetic assimilation may advance our basic understanding of how novel traits and species evolve. Here, Ehrenreich and Pfennig (pp. 769–779) discuss the potential role of genetic assimilation in evolution, and propose genetic and molecular mechanisms that might facilitate the process.
An evo-devo perspective on diversity and disparity (Review)

Diversity (number of included species) and disparity (extent of morphospace occupied) are indicators of a clade’s success. Many large genera show high diversity with low disparity; others like Euphorbia are highly diverse but also exhibit high disparity. Minelli (pp. 781–794) analyses large plant and animal clades from the point of view of evolutionary developmental biology, focusing on three main concepts. First, evolvability, in terms of genetic or developmental conditions favouring multiple parallel evolutionary transitions. Second, phenotypic plasticity, as facilitator of speciation. Third, modularity and heterochrony. His analysis suggests priorities for future research, including the possible role of saltational evolution in fostering diversity and/or disparity.
Molecular mechanisms of noise and robustness (Review)
Robustness, the inverse of noise, is a molecularly programmed feature of biological systems. The molecular networks of some organismal phenotypes like development are designed to maximize robustness, while other phenotypes like disease resistance have their pathways structured to allow for decreased robustness/increased noise to counteract evolution in the attacking organism. Lachowiec et al. (pp. 795–809) describe the specific molecular components that are utilized in networks to finely tune the proper level of robustness/noise for an array of phenotypes.
Evolution of unusual morphologies in Lentibulariaceae and Podostemaceae (Review)

Rutishauser (pp. 811–832) presents a pictorial report on released bauplans as typical for bladderworts (Utricularia, Lentibulariaceae) and river-weeds (Podostemaceae) within eudicots (angiosperms). Lentibulariaceae are carnivorous, possessing flypaper traps, eel-traps or sucking traps. They live as humid terrestrials, as submerged aquatics (except for their flowers), or even as epiphytes. Most Podostemaceae are restricted to rocks in tropical river-rapids and waterfalls. As submerged haptophytes during the rainy season, they emerge with their flowers afterwards. The recent scientific progress in the developmental biology and evolutionary history of both Lentibulariaceae and Podostemaceae is summarized.
Developmental hourglass in plant and animal embryogenesis (Review)

Embryogenesis has evolved independently in plants and animals. Comparative studies in animals have pointed towards an hourglass pattern of embryo development where early events are less conserved across species but lead to a highly conserved middle period, before diversifying again. Cridge et al. (pp. 833–843) explore comparative morphological studies and more recent molecular work in plants to ask if a similar hourglass pattern of embryogenesis exists. They then discuss what convergent evolution of such a pattern in plants and animals tells us about the developmental constraints imposed upon embryogenesis and the evolution of the underlying gene regulatory networks.
Tinkering with transcription factor networks for developmental robustness of Ranunculales flowers (Review)

Species within the Ranunculales bear flowers of formidable variation; for example, one can find novel floral organs and loss of floral organs, zygomorphic and radially symmetric flowers, and variation in floral organ number. Becker (pp. 845–858) reviews recent developments on the molecular aspects of Ranunculales flower development, focusing especially on aspects not found in Brassicaceae development, such as the ring meristem producing variable numbers of stamens. A new hypothesis for the evolution of the progressively fixed angiosperm floral bauplan is proposed, comparing the genetic interactions observed in Ranunculales with those found in Brassicaceae.
Why should we investigate the morphological disparity of plant clades? (Research in Context)

Macroevolution of major clades is often studied by plotting their taxonomic diversity through time. It is equally informative, but less commonplace, to investigate how clades explore anatomical ‘design’ space by quantifying their morphological disparity through time. Counterintuitively, diversity and disparity are usually decoupled. Metazoan clades often reach their highest disparity relatively early in their evolution, while diversity is still low. Oyston et al. (pp. 859–879) show that this macroevolutionary rule of thumb holds true in a sample of major plant clades, and discuss the types of data and methodological approaches that will facilitate future work on plant disparity.
Stochastic variation in Cardamine hirsuta petal number

Petal number is a robust trait in flowering plants. Crucifer flowers have four petals but Monniaux et al. (pp. 881–887) show that petal number varies from zero to four in Cardamine hirsuta. They mapped quantitative trait loci controlling stochastic variation in petal number and found that both the mean and uniformity of this trait share a common genetic basis. They suggest that this genetic variation, which is available to alter the mean and variance of petal number in C. hirsuta, may be cryptic in species such as Arabidopsis thaliana with robust petal number.
Pollinators and corolla shape robustness in Brassicaceae

How pollinators may influence the integration and developmental robustness of angiosperm flowers is still unknown. Using geometric morphometrics, Gómez et al. (pp. 889–904) studied the evolution of the phenotypic variation, disparity and integration of the corolla in 111 Brassicaceae, and explored the role of pollinators in mediating its evolution. Floral integration and robustness were evolutionary labile and varied among pollination niches. Whereas plants visited by efficient bees and beeflies displayed integrated corollas, those visited by nocturnal moths had corollas with high phenotypic variation and disparity. This suggests that pollinators can influence the evolution of important developmental attributes of the corolla even in plants with generalized pollination systems and conserved floral bauplans.
SUPERMAN allelic series – reframing flower developmental homeostasis

Breuil-Broyer et al. (pp. 905–923) show that SUPERMAN is a flower developmental canalization and robustness factor. First, partial dissociation of sexual boundary and flower meristem termination control in the sup allelic series (superman, superwoman, and supersex) has facilitated characterization of the partial independence of SUP-mediated WUS repression on the AG and CLV pathways. Setting a precise sexual boundary may have facilitated refinement of flower development, including co-option of the meristem patterning system for gynoecium development. Secondly, novel, late SUP functions in carpel patterning show that SUP operates in conjunction with KNUCKLES, a SUP-related placenta boundary gene. Lastly, combined understanding of both early and late SUP functions supports an evo-devo scenario reconciling the Mostly Male and Out of Male/Out of Female theories, and reveals the central role of class B/C homeotic gene balance modulation in the evolutionary change from uni- to bisexuality.
Robustness of floral organ numbers

Despite the contribution of phenotypic variation in floral morphologies to speciation, species diversity has been recognized by modal morphologies where variation is averaged out. Here Kitazawa and Fujimoto (pp. 925–935) show a relationship between the species-representative phenotype and phenotypic variation, by analysing quantitatively intraspecific variation of the organ numbers within flowers of Ranunculaceae and the numbers of flowers within the capitula of Asteraceae. The relationship followed a common mathematical function, showing robustness on the species-representative organ numbers.
Evolution of flower development during an adaptive radiation in Caiophora

The rare occurrence of a reversion to an ancestral state following adaptive radiations of pollination syndromes suggests that changes in the ancestral developmental programme of flowers are common in this scenario. Strelin et al. (pp. 937–947) provide evidence that the latter took place during the recent diversification into different pollination syndromes (bee, hummingbird, and rodent pollination) of Caiophora (Loasaceae subfam. Loasoideae) in the central Andes. They use geometric morphometrics techniques to show that derived (Caiophora) species have divergent ontogenetic patterns of floral development from those observed for basal bee-pollinated species.