

ABSTRACT
Growth hormone (GH) is a pleiotropic hormone that triggers STATs, ERK1/2 and Akt signaling, related to cell growth and proliferation. Transgenic mice overexpressing GH present increased body size, with a disproportionate liver enlargement due to hypertrophy and hyperplasia of the hepatocytes. We had described enhanced mitogenic signaling in liver of young adult transgenic mice. We now evaluate the activation of these signaling cascades during the growth period and relate them to the morphological alterations found. Signaling mediators, cell cycle regulators and transcription factors involved in cellular growth in the liver of GH-overexpressing growing mice were assessed by immunoblotting, RT-qPCR and immunohistochemistry. Hepatocyte enlargement can be seen as early as 2-weeks of age in GH-overexpressing animals, although it is more pronounced in young adults. Levels of cell cycle mediators PCNA and cyclin D1, and transcription factor c-Jun increase with age in transgenic mice with no changes in normal mice, whereas c-Myc levels are higher in 2-week-old transgenic animals and cyclin E levels decline with age for both genotypes. STAT3, Akt and GSK3 present higher activation in the adult transgenic mice than in the growing animals, while for c-Src and mTOR, phosphorylation in GH-overexpressing mice is higher than in control siblings at 4 and 9 weeks of age. No significant changes are observed for ERK1/2, neither by age or genotype. Thus, the majority of the mitogenic signaling pathways are gradually up-regulated in the liver of GH-transgenic mice, giving rise to the hepatic morphological changes these mice exhibit.
KEYWORDS: Growth hormone, GH-overexpressing mice, liver, hepatomegaly, PCNA, cell cycle regulators, c-Myc, c-Jun, STAT3, ERK1/2, Akt/mTOR/GSK3β
Introduction
Growth hormone (GH) is implicated in the regulation of body growth but also in lipid, carbohydrate and protein metabolism. In fact, it has recently been defined as essential for integrating metabolism and growth.1 Its actions are either direct or mediated by the insulin-like growth factor 1 (IGF1). Upon GH binding to its receptor (GHR) on the cell surface, several signaling pathways are activated, including the signal transducers and activators of transcription (STATs), the MAPK (mitogen activated protein kinase) kinase (MEK)/extracellularly regulated kinase (ERK) and the phosphatidylinositol 3 kinase (PI3K)/Akt pathways.2-6 STAT5b, regarded as the main GH signaling mediator, is involved in GH-induced IGF1 gene transcription6-9 and it is critical for determining body stature.5 MEK/ERK and PI3K/Akt pathways are engaged in protein synthesis required for cellular growth and promote cellular proliferation and differentiation.10
Transgenic mice overexpressing growth hormone present high circulating GH levels since birth, despite of which, they exhibit accelerated growth only after the third week of postnatal life11-13 and achieve greater adult body size than control mice.14-17 We have recently reported that STAT5 signaling is enhanced in the liver of growing GH-transgenic mice but not in adult animals, supporting a central role of this pathway in the promotion of body growth during the postnatal growth period.13,17,18
Enhanced body growth of these GH-overexpressing mice is concomitant with organomegaly, which is particularly important in the liver.16,17,19-23 Hepatomegaly emerges as a consequence of the hypertrophy and hyperplasia of the hepatocytes, with the frequent progression to liver tumors at advanced ages.24-26 The molecular mechanisms implicated in this process are being studied; we have reported aberrant activation of several signaling cascades in young adult GH-overexpressing mice, including STAT3, MEK/ERK and PI3K/Akt. Indeed, these animals exhibit up-regulation of several signaling mediators of cell proliferation and survival in the liver, including Akt, its downstream targets mTOR and GSK3, and ERK1/2.16,17 They also exhibit higher activation of STAT316,17 which is involved in liver inflammation and cancer development and progression.27,28 Therefore, the activity of these GH-signaling cascades during the growth period was assessed in order to evaluate their contribution to the enlargement of the hepatic tissue and their relation to the morphological alterations found in these animals. The current study presents an in vivo analysis of Src, STAT3, MEK/ERK and PI3K/Akt signaling pathways and the variation in the expression of cell cycle regulators implicated in the promotion of cellular growth in the liver of growing GH-overexpressing mice.
Materials and methods
Animals
Transgenic PEPCK-bGH mice containing the bGH gene fused to control sequences of the rat phosphoenolpyruvate carboxykinase (PEPCK) gene were derived from animals kindly provided by Dr. Thomas E. Wagner and Jeung S. Yun (Ohio University, Athens, OH). Hemizygous transgenic mice were produced by mating transgenic males with normal C57BL/6 X C3H F1 hybrid females purchased from the Jackson Laboratory (Bar Harbor, ME).
Animals of representative ages were chosen: 2 weeks (2w), when animals of both genotypes exhibit similar body size; 4 weeks (4w), when faster growing transgenic mice start to exhibit different body proportions than normal siblings; and nine weeks (9w), young adults used as reference, which have a clear enlargement of body size.13 Non-transgenic siblings were used as controls. The mice were housed 3 to 5 per cage in a room with controlled light (12 h light per day) and temperature (22 ± 2 C); they had free access to food (Rodent Laboratory Chow 5001; not autoclaved; 23.4% protein, 4.5% fat, 5.8% crude fiber; LabDiet, PMI Feeds, Inc.) and tap water.
The appropriateness of the experimental procedure, the number of animals used, and the method of acquisition were in compliance with federal and local laws and with institutional regulations. The experiments were approved by the Southern Illinois University Laboratory Animal Care Committee.
Tissue collection and solubilization
Mice were sacrificed by cervical dislocation under isoflurane anesthesia. Livers were removed and stored at −80°C until use. Samples were prepared for the different assays according to Martinez et al.13
Immunoblotting
Samples were separated by SDS-PAGE and transferred to PVDF membranes as already described.13,29 The membranes were then incubated overnight at 4°C with antibodies against AKT 1:1000 (no. 835), pAKT (Ser473) 1:1000 (no. 1529), ERK1/2 1:1000 (no. 9022), pERK1/2 (Thr202/Tyr204) 1:1000 (no. 9023), GSK3β 1:1000 (no. 9315), pGSK3β (Ser9) 1:1000 (no. 9323), Src 1:3000 (no. 2109), pSrc (Tyr416) 1:1000 (no. 6943), pSTAT3 (Tyr705) 1:2000 (no. 9131), mTOR 1:1000 (H-205, no. 8986), pmTOR (Ser2448) 1:1000 (no. 610622) purchased from Cell Signaling Technology Inc. Antibody against STAT3 1:2000 (no. S21320) was purchased from Transduction Laboratories (BD Biosciences). Immunoreactive proteins were visualized by enhanced chemiluminescence (Pierce ECL-Plus, Thermo Scientific) using hyperfilm ECL (GE Healthcare LifeSciences). Band intensities were quantified using Gel-Pro Analyzer software (Media Cybernetics).
Regular proteins used for loading control of immunoblotting exhibit variation in the liver of PEPCK-bGH mice, as already described.13,16, 17,29 In addition, actin and tubulin abundance is increased in preneoplastic cells.30,31 Therefore, different determinations were routinely performed to assure goodness of assays; protein loading control was performed by Coomassie blue staining of the PVDF membranes after blotting experiments.13
Histology and immunohistochemical staining
Sections of paraffinized livers were stained with hematoxylin and eosin (H&E) following standard procedures. Morphology and size of hepatocytes were evaluated under light microscopy. Relative cell size was estimated by counting the number of cells per field.
Immunostaining of liver paraffinized sections was performed as previously described;29 primary antibodies anti-PCNA 1:100 (no. 7907) and anti-STAT3 1:150 (no. S21320) were purchased from Santa Cruz Biotechnology Laboratories and Transduction Laboratories, respectively. Quantification of PCNA was estimated as percentage of positive (brown stained) nuclei in 500 cells approximately.
Reverse transcriptase quantitative PCR (RT-qPCR)
This method was used to evaluate gene expression and also to assess the genotype of the animals, since growing mice are not readily distinguishable by their body size. For this purpose, the bGH transgene was determined. The protocol used was already described.13 The sequences of primer sets (5′→3′), obtained from Invitrogen, are: bGH (F:GGGCAGATCCTCAAGCAGAC; R: GAAGCAGGAGAGCAGACCG), Cyclin D1 (F:GCGAAGTGGAGACCATCCG; R:GGTCTCCTCCGTCTTGAGC), Cyclin E (F:GGCGGACACAGCTTCGGGTC; R:TGGGTCTTGCAAAAACACGGCCA), Cyclophilin A (F:GCGTCTCCTTCGAGCTGTT; R:AAGTCACCACCCTGGCAC), c-Jun (F:CATTGCCGCCTCCAAGTG; R:CCAGCTCGGAGTTTTGCG), c-Myc (F:TCACCAGCACAACTACGCCG; R:TGCTTCAGGACCCTGCCACT). Cyclophilin A was used as normalizing control.
Statistical analysis
Statistical analysis was performed using the GraphPad Prism statistical program by GraphPad Software, Inc. (San Diego). Data were first analyzed by one-way ANOVA followed by Bonferroni to evaluate sex differences. Subsequently, data were analyzed by 2-way ANOVA and Bonferroni post-test to assess differences by age and genotype. Data were considered statistically significant if P < 0.05.
Results and discussion
Hepatocyte, liver and body growth
Somatometric determinations
Body growth is attained by tissue growth, a process regulated through precise mechanisms that limit the size organs can achieve.32,33 Mice overexpressing GH, apart from increased body size, exhibit hepatomegaly –disproportionate liver growth- at adulthood, due to hypertrophy and hyperplasia of hepatocytes, which frequently progress to hepatoma at older ages.16,20,23,24,26,34,35
Liver and body weight were analyzed in order to detect when liver size becomes disproportionate to body size. Animals of representative ages were chosen in line with our previous report in these GH-overexpressing animals.13 Two-week-old normal and GH-transgenic mice (2w) cannot be distinguished by body weight (Table 1). Four-week-old GH-overexpressing mice (4w) exhibit greater body size than their normal siblings. Nine-week-old mice (9w) are young adults used as reference; GH-transgenic giant animals already present hepatocellular alterations and dysregulation of the Akt and ERK signaling pathways.16,17
Table 1.
Body and liver weight in growing GH-overexpressing mice and normal littermates.
| 2w N | 2w T | 4w N | 4w T | 9w N | 9w T | |
|---|---|---|---|---|---|---|
| BW (g) | 7.5 ± 0.2(a) | 7.5 ± 0.4(A) | 15.3 ± 0.7(b) | 21.9 ± 0.9(B*) | 23.7 ± 1.2(c) | 39.4 ± 1.7(C*) |
| BW ratio for T/N | 0.98 ± 0.05(A) | 1.45 ± 0.09(B) | 1.56 ± 0.09(B) | |||
| LW (g) | 0.22 ± 0.01(a) | 0.27 ± 0.03(A) | 0.94 ± 0.07(b) | 1.62 ± 0.06(B*) | 1.23 ± 0.09(b) | 3.15 ± 0.14(C*) |
| LW ratio for T/N | 1.23 ± 0.07(A) | 1.75 ± 0.20(B) | 2.57 ± 0.14(C) | |||
| LW/BW(%) | 2.9 ± 0.1(a) | 3.6 ± 0.2(A) | 6.1 ± 0.2(b) | 7.4 ± 0.2(B*) | 5.4 ± 0.2(b) | 8.1 ± 0.1(C*) |
| LW/BW ratio for T/N | – | 1.16 ± 0.05(A) | – | 1.23 ± 0.14(A) | – | 1.57 ± 0.68(B) |
Body (BW) and liver weight (LW) of GH-transgenic animals (T) and their non-transgenic littermates (N) was assessed for 2-week-old (2w), 4-week-old (4w) and 9-week-old (9w) mice. Data are presented as mean value ± SEM (n = 8). Different letters denote significant difference by age; small letters correspond to normal mice and capital letters to transgenic animals. Asterisks indicate significant difference between GH-overexpressing animals and their corresponding non-transgenic age controls. Ratios are calculated from individual values and not from the averages in the table, which may account for discrepancies observed.
Both liver and body weight increased with age in normal and transgenic mice, the gain was larger in GH-overexpressing mice. When liver weight was referred to body weight, greater values were obtained for transgenic mice of 4 and 9 weeks of age compared to controls (Table 1). Moreover, an increase in liver weight and in its proportion relative to body size can be apparent as early as 2 weeks of age, although significance is not achieved by that age.
Hepatocellular size
The size and morphology of hepatocytes were evaluated by histological analysis on tissue sections stained with hematoxylin and eosin. To estimate cellular size, cells were counted per optical field; the number of cells per field is inversely proportional to cell size. An increment in hepatocellular size with age in normal and transgenic mice was observed, despite of which, hepatocytes in transgenic mice were bigger at all ages studied. In liver sections of 4-week-old animals, some cells were markedly larger than the average size. In 9-week-old transgenic mice, extended areas of large dysplastic cells were observed (Fig. 1A-C), as previously reported23,25,36; moreover, increased nuclear size can also be detected (Fig. 1A).16 Photomicrographs corresponding to male mice liver are shown. These alterations may be a direct consequence of GH action, since they are not found in transgenic mice overexpressing IGF1.16,34-36
Figure 1.
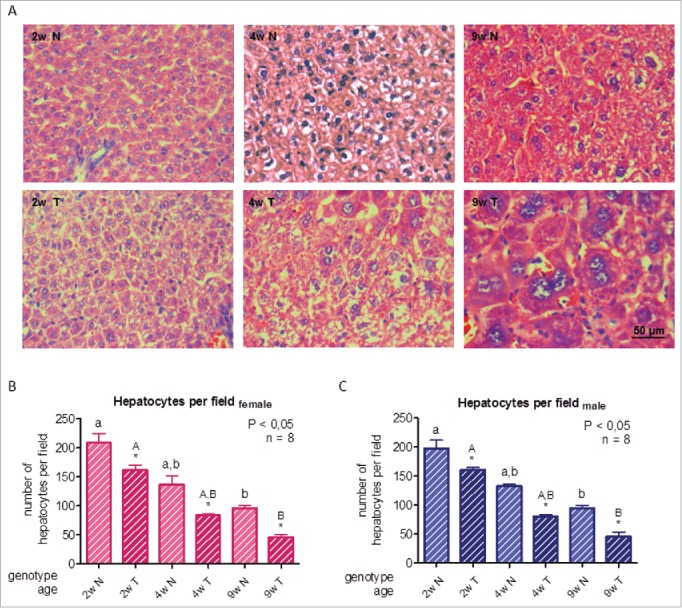
Histological analysis of livers from growing GH-overexpressing transgenic mice and normal littermates. A: Representative photomicrographs of hematoxylin and eosin staining of liver sections from normal and GH-transgenic male mice. B: Hepatocyte quantification in female mice. C: Hepatocyte quantification in male mice. Hepatocyte size of GH-transgenic animals (T) and their non-transgenic littermates (N) was assessed for 2-week-old (2w), 4-week-old (4w) and 9-week-old (9w) mice. Hepatocyte size was estimated counting the number of cells per visual field; original magnification of 400x. Data are the mean ± SEM of 8 sets of different individuals per group. Different letters denote significant difference by age within a genotype; small letters correspond to normal mice and capital letters to transgenic animals. Asterisks indicate significant difference between GH-overexpressing animals and their corresponding non-transgenic age controls.
Cell cycle regulators
To evaluate hepatocellular proliferation, immunohistochemistry of the S-phase-related proliferating cell nuclear antigen (PCNA) was performed. In normal mice, the proportion of hepatocytes positive for PCNA in nucleus did not significantly vary at any age studied, although values were numerically higher for 4-week-old animals than in the young adult. For 2-week-old transgenic mice, levels were similar to those of controls, while GH-overexpressing animals had higher PCNA nuclear staining than their normal littermates since the fourth week onwards, with a broader difference between normal and transgenic mice in 9-week-old animals (Fig. 2A-C), suggestive of increased cell mitotic activity in GH-overexpressing animals.
Figure 2.
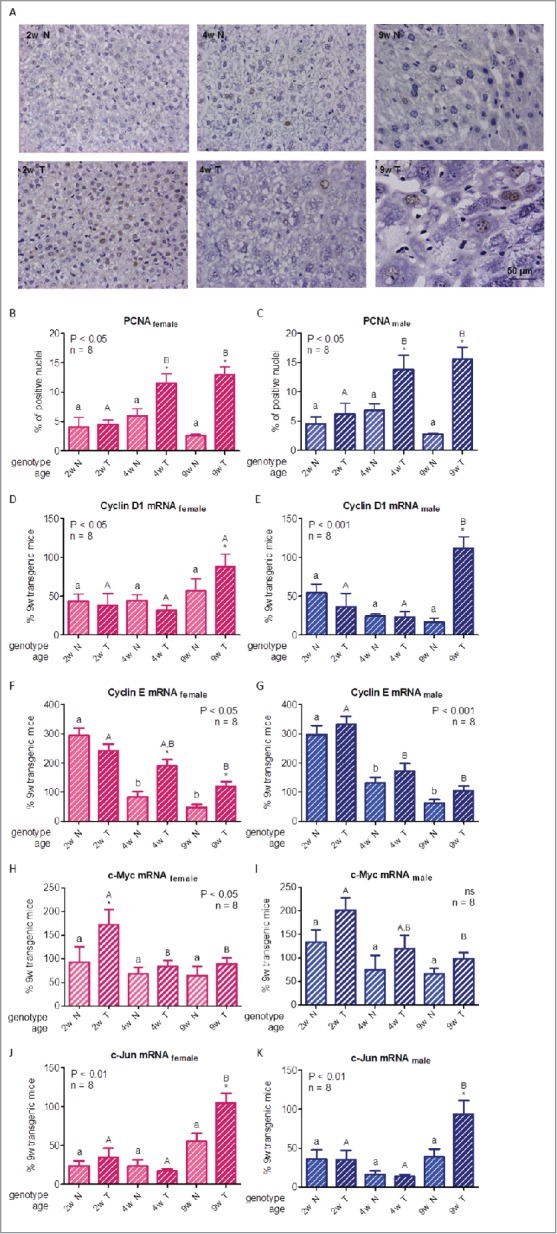
For figure legend, see next page. Hepatocellular proliferation markers in growing GH-overexpressing transgenic mice and normal littermates. A: Representative photomicrographs of immunohistochemical staining of liver sections from normal and GH-transgenic male mice with anti-PCNA antibody; original magnification of 400x. B,C: PCNA immunostaining quantification in female and male mice liver respectively. D,E: Cyclin D1 mRNA hepatic expression in female and male mice. F,G: Cyclin E mRNA hepatic expression in female and male mice. H,I: c-Myc mRNA hepatic expression in female and male mice. J,K: c-Jun mRNA hepatic expression in female and male mice. Different cell cycle regulators and transcription factors involved in cellular proliferation were assessed in liver of GH-transgenic animals (T) and their non-transgenic littermates (N) for 2-week-old (2w), 4-week-old (4w) and 9-week-old (9w) mice. PCNA content was calculated as percentage of positive (brown stained) nuclei in 500 cells approximately. For simplification, only pictures from males are shown (A). To determine gene expression, mRNA was assessed by RT-qPCR from total RNA extracts. Values were related to cyclophilin A levels, referred to the average for 9-week-old transgenic female and male mice and expressed as percentage. Data are the mean ± SEM of the indicated n number of samples per group, each one representing a different animal. Different letters denote significant difference by age within genotype; small letters correspond to normal mice and capital letters to transgenic animals. Asterisks indicate significant difference between GH-overexpressing animals and their corresponding non-transgenic age controls.
The cell cycle comprises different successive phases; the transition between them is tightly controlled by a complex of cyclin-dependent kinases (CDKs) and their activating cyclins. Expression of the cyclins varies throughout the cycle; cyclins D and E are the main activators of the G1 to S phase transition; moreover, their dysregulation has been associated with cancer.37,38 Therefore, we evaluated cyclins D1 and E gene expression by RT-qPCR. Cyclin D1 mRNA content variations were not statistically significant for normal mice at any age studied or for 2- and 4-week-old GH-overexpressing mice, while an increase in gene expression was observed for 9-week-old transgenic mice (Fig. 2D-E). Cyclin E gene expression was elevated in 2-week-old mice, with similar values for both genotypes, suggesting a physiological role of the activator at this age, after which, its abundance declined. Transgenic animals of 4 and 9 weeks of age exhibited higher levels than age-matched normal littermates, although the differences were significant only for female mice (Fig. 2F-G). We had already reported that GH-transgenic adult mice display higher expression of these cyclins.16 Acting together in the control of the initial steps of the cell cycle, higher expression of cyclin D1 in 9-week-old, and of cyclin E in 4- and 9-week-old GH-overexpressing mice liver, would be favoring cell cycle progression and, in consequence, an elevated proliferation of the hepatocytes observed in these mice.
The proto-oncogenes c-Myc and c-Jun are involved in cell cycle progression, and their expression is induced through the ERK and Akt mitogenic signaling pathways28,39 that are enhanced in the liver of adult GH-transgenic mice.17 Moreover, they are among the few transcription factors related to cell cycle regulators.40 Therefore, their expression was evaluated in the liver of normal and transgenic growing mice. c-Myc mRNA content exhibited a declining tendency with age, which was significant for GH-overexpressing animals only. Although transgenic mice had slightly higher values than controls, difference between genotypes was significant for 2-week-old females only (Fig. 2H-I), despite we had previously described a marked difference between normal and transgenic animals in young adults both at the mRNA and protein level.16 It is well known that c-Myc mRNA and protein expression levels correlate with cell proliferation; moreover, this upregulation is noteworthy in the transition from quiescence into the proliferative state, but not in continuously dividing cells.41,42 It is therefore not surprising to find higher levels of this transcription factor in 2-week-old GH-overexpressing mice and lower levels at older ages. However, as c-Myc induces the expression of the positive cell-cycle regulator cyclin D1, a closer relationship between these 2 modulators would have been expected.42
When c-Jun mRNA content was analyzed, the differences found for normal animals with age did not achieve statistical significance, whereas, transgenic mice presented similar levels to normal controls in 2- and 4 week-old animals, but exhibited higher gene expression at the age of 9-weeks. When results were assessed by genotype, 9-week-old mice had higher levels than their normal controls (Fig. 2J-K). The mRNA expression pattern of c-Jun and cyclin D1 are parallel, in accordance with c-Jun influence over cyclin D1 activity.37,40
To recapitulate, body growth implies an accompanying growth of tissues, yet GH-overexpressing transgenic mice display a disproportionate liver size increase. In 4- and 9-week-old transgenic mice liver, higher cellular proliferation is evidenced by the higher expression of different cell cycle regulators implicated in this process. Also, cell size is larger in GH-overexpressing mice than in their normal controls at every age studied. At adulthood, dysplastic cells predominate, indicative of a premalignant condition.
Mitogenic signaling pathways
The STAT3, Ras/MEK/ERK and PI3K/Akt pathways, increased in the liver of GH-transgenic adult mice,17 are signaling cascades controlling the expression of the cell cycle regulators described.37,41,42 These mitogenic signaling pathways, as well as Src-kinase activation which was also enhanced in young adult GH-overexpressing mice,17 were analyzed during the growth period, in order to evaluate their association with the morphological and biological changes described in the previous section.
Src signaling
GH signaling initiates through activation of JAK2, the GHR-associated tyrosine kinase. However, some signaling mediators can be activated independently of JAK2, by members of the Src family of kinases, particularly by Src. In this way, STATs like STAT3, or the MEK/ERK pathway can also be recruited by Src.43,44 At a physiological level, Src protein-tyrosine kinase is a key player that participates in cell growth, division, migration and survival.45
The tyrosine kinase activity of Src is regulated through a balance between phosphorylation and dephosphorylation of positive and negative regulatory residues.46-48 We have previously shown that Src is upregulated in the liver of adult PEPCK-bGH mice, and that increased phosphorylation status and subsequent kinase activity are mainly a consequence of the higher Src protein abundance in these animals.17 In the current study, Src phosphorylation at Tyr416, a positive regulatory autophosphorylation site, and its protein content were evaluated by Western blotting. When phosphorylated Src levels were analyzed, no significant differences were detected in normal mice throughout the period studied, whereas transgenic animals displayed values similar to those of controls for 2-week-old mice, but exhibited increased phosphorylation levels in 4- and 9-week-old animals (Fig. 3A-B). Moreover, 9-week-old transgenic females displayed higher levels than males. Src protein hepatic abundance presented an age-dependent decrease in both genotypes, with higher levels in transgenic mice at all ages studied (Fig. 3C-D). Hence, while phosphorylated Src levels increase with age in transgenic mice, Src protein abundance decreases, suggesting that the hyperactivation of this kinase has a participation in the aberrant transformation of the liver of GH-overexpressing mice.
Figure 3.
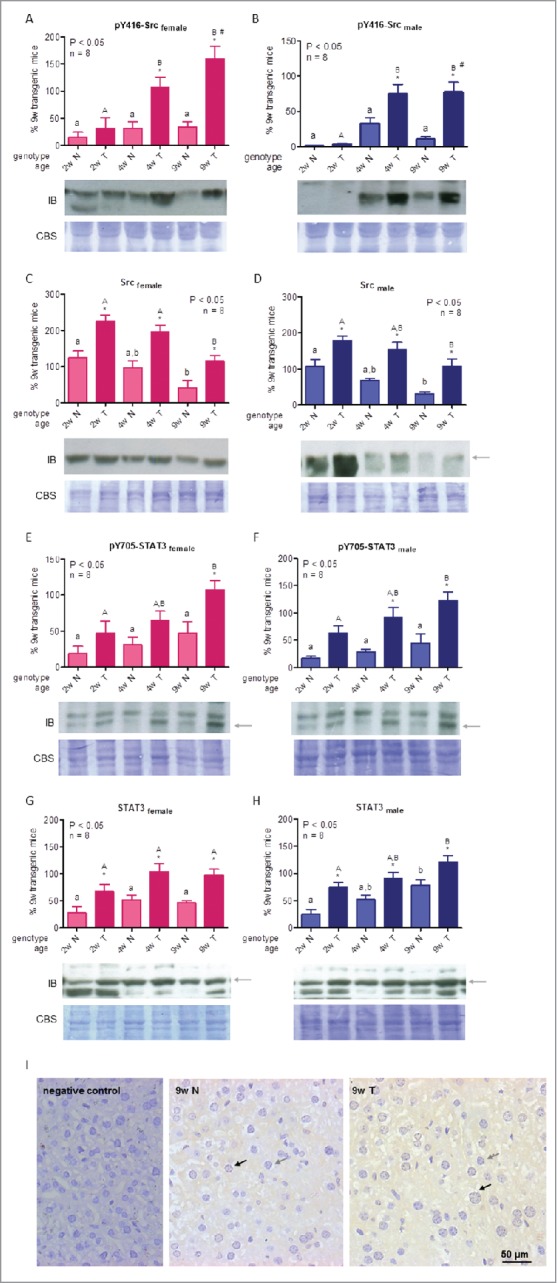
For figure legend, see next page. Src and STAT3 signaling in liver of growing GH-overexpressing mice and normal littermates. A,B: Src phosphorylation at tyrosine 416 in female and male mice. C,D: Src protein abundance in female and male mice. E,F: STAT3 phosphorylation at tyrosine 705 in female and male mice. G,H: STAT3 protein abundance in female and male mice. I: STAT3 liver immunohistochemical staining in male adult mice. Signaling mediators were assessed in liver of GH-transgenic animals (T) and their non-transgenic littermates (N) for 2-week-old (2w), 4-week-old (4w) and 9-week-old (9w) mice. To determine phosphorylation and protein abundance, equal amounts of solubilized liver protein were assessed by immunoblotting. Representative immunoblots (IB) as well as Coomassie blue staining (CBS) of PVDF membranes are shown. Bands were quantified by scanning densitometry. Results were expressed as percentage, referred to the average for 9-week-old transgenic female and male mice. Data are the mean ± SEM of the indicated n number of samples per group, each one representing a different animal. Different letters denote significant difference by age within genotype; small letters correspond to normal mice and capital letters to transgenic animals. Asterisks indicate significant difference between GH-overexpressing animals and their corresponding non-transgenic age controls. Number sign indicates significant difference between sexes. Arrows show the quantified band. For IHC analysis (Panel I), representative photomicrographs of immunohistochemical staining with anti-STAT3 antibody of liver sections from normal and GH-transgenic male adult mice is shown. Positive nuclei are shown with black arrows and negative ones with gray arrows.
STAT3 signaling
STATs, particularly, STAT3, are transcription factors involved in the promotion of cell proliferation, differentiation and survival. In normal cells, ligand-mediated activation of the STATs is a temporary event, whereas in numerous types of cancer, usually associated to dysregulation of growth factors, the oncogenic factor STAT3 is constitutively active.49
STAT3 may be activated by different tyrosine kinases, including JAK2 and Src.49 STAT3 phosphorylation at its activating residue, Tyr705, as well as its protein content, were evaluated in mice at 2, 4 and 9 weeks of age by Western blotting. STAT3 phosphorylation increased with age in both genotypes, although statistical significance was achieved in transgenic mice only. GH-overexpressing animals exhibited numerically higher levels than normal siblings at all ages; however, statistical differences were reached for 9-week-old females and for 4- and 9-week old males only (Fig. 3E-F). STAT3 protein content was higher in transgenic mice than in controls at all ages. There was a tendency toward increasing levels of STAT3 protein with increasing age in males of both genotypes, which was not as pronounced for female animals (Fig. 3G-H). Results found for GH-overexpressing mice suggest that this transcription factor could be related to the transforming processes that take place in the liver of these animals.17 STAT3 modifies the expression of genes that participate in cell cycle progression, like c-fos, c-myc and cyclin D1.37,49 However, there is no clear correspondence between the activated transcription factor and the cell cycle regulators determined.
STAT3 is activated in hepatocytes by GH, but also by the pro-inflammatory cytokine IL6, which is released by the Kupffer cells of the liver.27,50 In addition, elevated levels of activated STAT3 were observed in close relation with high hepatic expression of IL6 in GH-transgenic mice.51 However, STAT3 can also be activated in the Kupffer cells by other factors, like IL10, where it has the opposite effect, suppressing liver inflammation.52,53 Therefore, it was important to assess STAT3 cellular localization. Immunohistochemical analysis was carried out in liver sections of 9-week-old normal and transgenic animals. STAT3 staining was detected mainly in hepatocytes, both in the cytoplasm as well as at the nuclear level (Fig. 3I), with a greater proportion in transgenic mice, thus suggesting this protein may have a role in the promotion of liver inflammation in GH-overexpressing animals.
MEK/ERK signaling
GH, as well as several other hormones and cytokines, activates the MEK/ERK signaling cascade. This activation begins with the recruitment of Shc to an activated receptor complex and its subsequent phosphorylation, Shc then interacts with the adapter protein Grb2. Pre-associated with Grb2 is SOS, a guanine nucleotide exchange factor which once located at the cell membrane activates Ras, triggering a sequence of serine/threonine kinase phosphorylations: Raf-1, MEK1/2 and ERK1/2 (hereafter referred to as ERK). In turn, phosphorylated ERKs translocate to the nucleus, where they activate transcription factors like Elk-1 and c-Jun, or induce the expression of c-Fos and c-Myc, that regulate cell proliferation and survival.5,42
ERK1 and ERK2 (44 and 42 kDa respectively), 2 overlapping kinases, are activated by concomitant phosphorylation at Thr202 and Tyr204. When ERK activation was evaluated in liver of growing GH-overexpressing mice, no significant differences were observed by genotype or age (Fig. 4A-B). The lack of variation of phosphorylated ERK levels between genotypes is in line with a previous report of our group in adult mice liver.54 However, Miquet et al.17 observed a variation in Tyr204 phosphorylation between genotypes but similar levels in Thr202 activation. When ERK hepatic abundance was assessed, it presented the highest levels in 2-week-old mice and a subsequent decrease in 4- and 9-week-old animals. Four-week-old animals presented similar abundance, whereas 9-week-old transgenic animals had higher levels than controls, in line with previous reports.17 Moreover, higher content at 2 weeks of age was more pronounced in females for both genotypes (Fig. 4C-D). In light of these results, MEK/ERK pathway does not seem to be participating in the hepatic alterations observed in GH-transgenic adult mice, despite the up-regulation of c-Jun, and of cyclin D1, 2 major targets of the MAPK pathway.55
Figure 4.

ERK and Akt signaling in liver of growing GH-overexpressing mice and normal littermates. A,B: ERK- phosphorylation at threonine 202 and tyrosine 204 in female and male mice. C,D: ERK protein abundance in female and male mice. E,F: Akt phosphorylation at serine 473 in female and male mice. G,H: Akt protein abundance in female and male mice. I,J: GSK3 phosphorylation at serine 9 in female and male mice. K,L: GSK3 protein abundance in female and male mice. M,N: mTOR phosphorylation at serine 2448 in female and male mice. O,P: mTOR protein abundance in female and male mice. Signaling mediators were assessed in liver of GH-transgenic animals (T) and their non-transgenic littermates (N) for 2-week-old (2w), 4-week-old (4w) and 9-week-old (9w) mice. Equivalent amounts of solubilized tissue were analyzed by immunoblotting. Representative immunoblots (IB) as well as Coomassie blue staining (CBS) of PVDF membranes are shown. Bands were quantified by scanning densitometry. Values were expressed as percentage, referred to the average for 9-week-old transgenic female and male mice. Data are the mean ± SEM of the indicated n number of samples per group, each one representing a different animal. Different letters denote significant difference by age within genotype; small letters correspond to normal mice and capital letters to transgenic animals. Asterisks indicate significant differences between GH-overexpressing animals and their corresponding non-transgenic age controls; ns stands for non-significant. Number sign indicates significant difference between sexes. Arrows indicate the quantified bands.
Akt signaling
The PI3K/Akt cascade initiates with the interaction of the phosphatidylinositol-3´-kinase (PI3K) to a ligand-activated receptor complex.56 Phosphatidylinositol (3,4,5)-triphosphate (PIP3) generated at the plasma membrane recruits PDK1 (pyruvate deshydrogenase lipoamide kinase isozyme 1) and Akt, where PDK1 phosphorylates Akt on T308, partially activating it.56 Akt is fully activated by Ser473 phosphorylation by the mTOR complex 2.57 Although Akt is regarded as the main PI3K effector, they may act independently of each other as well.56
The glycogen synthase kinase 3 (GSK3) is a pro-apoptotic factor; Akt phosphorylates the GSK3β isoform at Ser9, inhibiting its activity, and thus promoting cell survival.58 Another Akt substrate is the mammalian target of rapamycin (mTOR), a protein-kinase that controls the mammalian translation machinery, promoting protein synthesis; mTOR participates in 2 catalytic complexes, mTORC 1 and 2, regulating different cellular functions.10 Phosphorylation of mTOR residues Thr2446 and Ser2448 correlates with its activity, with serine phosphorylation being the predominant modification.59
When assessed by age, Akt phosphorylation at Ser473 displayed no significant changes in normal mice, although there was a trend for its levels to be higher in 9-week-old male mice compared to younger animals, in agreement with previous reports of our group.16 In GH-overexpressing mice, maximal phosphorylation levels were observed for 9-week-old animals in both sexes. When genotypes were compared, values of p-Akt appeared to be higher in transgenic male mice than in their normal siblings at all ages, although levels were statistically higher than controls for 9-week-old animals only (Fig. 4E-F).
Akt protein content did not vary with age in normal mice. In GH-overexpressing mice, Akt abundance was similar to non-transgenic controls at 2 weeks, and seemed to increase thereafter, although this age difference did not achieve statistical significance. However, this increase in Akt content in transgenic mice accounts for the difference found for genotype in 4- and 9-week-old animals (Fig. 4G-H). Taken together, these results suggest that elevated levels of phosphorylated Akt observed in transgenic mice can be attributed to higher protein content in these animals.
GSK3 phosphorylation levels at Ser9 were similar to phospho-Akt ones, age differences did not achieve statistical significance for normal mice while maximal levels were found for 9-week-old transgenic animals. When genotypes were compared, significant differences were observed in 9-week-old animals for both sexes (Fig. 4I-J). GSK3 hepatic abundance displayed a decreasing tendency with age in normal mice, whereas transgenic animals presented similar levels to controls at 2 weeks of age and higher levels at 4 weeks, which declined thereafter. Even though these changes were not significant, when differences were assessed by genotype, GH-transgenic mice showed higher GSK3 content than normal siblings in 4- and 9-week-old animals (Fig. 4 K-L). Moreover, the high levels found for GH-overexpressing 4-week-old animals revealed sexual dimorphism. GSK3 was reported to phosphorylate cyclin D1 to promote its degradation,37,58 thus, phosphorylation levels at inhibitory residue of the kinase could have been expected to correlate with cell cycle regulator expression.
mTOR phosphorylation at Ser2448 displayed a declining tendency in normal male mice and an increasing trend in transgenic female mice, although when variations associated with age were analyzed within each genotype, no statistical differences were observed (Fig. 4M-N). However, due to the divergent age patterns, higher levels were found in transgenic mice compared to controls at 4- and 9-weeks of age. Differences between sexes were observed in 2-week-old transgenic mice, where male mice had a higher degree of modification at its activating residue. Similarly to phosphorylated mTOR values, no statistically significant differences were observed in mTOR protein content with age for both genotypes but 4- and 9-week-old GH-overexpressing mice exhibited higher levels than normal controls of the same age (Fig. 4O-P). mTOR has a major role in the regulation of cellular growth and metabolism, mainly inducing protein synthesis, and it was also linked to the upregulation of c-Myc, thus reinforcing its oncogenic potential.60
In summary, the hepatic content of Akt and its substrates GSK3 and mTOR is higher in transgenic mice than in normal siblings in 4- and 9-week-old animals, and it is accompanied by a concomitant increase in their phosphorylation levels. Although growing transgenic mice exhibit greater values than normal siblings, the differences are more pronounced in young adults.
The PI3K/Akt and mTOR signaling pathway has been found altered in different tumors, including those of the liver.42,58,60-62 PI3K/Akt has a crucial role in hepatocyte growth, mainly regulating cell size;63 in consequence, mice overexpressing Akt exhibit hepatomegaly.64 If current results on hepatomegaly are compared to the Akt signaling described, a correlation between this cascade and liver enlargement in transgenic mice can be proposed, which is evident at the fourth week and becoming much more pronounced for 9-week-old animals.
Conclusion
Liver weight and hepatocyte size were evaluated in growing GH-transgenic mice and in their normal littermates. Disproportional hepatic growth could be detected as early as 2 weeks of age in GH-overexpressing mice; which, however, was more pronounced in young adults. One of the possible causes of greater tissue size is the increased cell size. Hypertrophy was detected in transgenic mice at all ages studied, with some large dysplastic hepatocytes in the growing mice and a greater extent of hepatic dysplasia in adults. Another cause of liver enlargement is the increase of cell number. Indeed, expression of different cell cycle regulators implicated in proliferation is increased in hepatocytes of transgenic adult mice, although some of them are up-regulated since the fourth week.
GH-transgenic mice have been extensively studied in order to elucidate the mechanisms through which GH exerts its action. Here, we show that the ERK pathway did not exhibit increased signaling activity in liver of GH-overexpressing mice during the growth period. On the other hand, the PI3K/Akt cascade was enhanced in transgenic mice liver during growth, with maximal values in the young adult. These signaling routes, related to cellular growth and proliferation, are induced by GH. Therefore, it is reasonable to expect they may be increased in GH-overexpressing mice. Cell cycle regulators and transcription factors determined are involved in cellular growth and they are augmented in different carcinomas as well, hence it is not surprising to find that most of them are enhanced in the dysplastic hepatocyte found in the young adult GH-overexpressing mice.
In light of all the evidence, hepatomegaly found in GH-transgenic adult mice starts developing at early ages, during the growth period. Akt signaling pathway may be implicated in the promotion of hepatocyte hypertrophy and hyperplasia.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
DT, AIS, JGM and LG are Career Investigators of CONICET; VGP is supported by a Fellowship from CONICET. Support for these studies was provided by UBA (20020130100412BA; 20020100100631), CONICET (PIP 427 and 320), and ANPCYT (PICT 2010-0398 and PICT 2013-2953) (Argentina) to DT and AIS, UBA (20020120200122) and ANPCyT (PICT 2008-0397) to JGM, and NIH (AG019899) to AB.
References
- [1].Sperling MA. Traditional and novel aspects of the metabolic actions of growth hormone. Growth Horm IGF Res 2015; PMID:26194064; http://dx.doi.org/ 10.1016/j.ghir.2015.06.005 [DOI] [PubMed] [Google Scholar]
- [2].Brooks AJ, Waters MJ. The growth hormone receptor: mechanism of activation and clinical implications. Nat Rev Endocrinol 2010; 6:515–25; PMID:20664532; http://dx.doi.org/ 10.1038/nrendo.2010.123 [DOI] [PubMed] [Google Scholar]
- [3].Xu J, Messina JL. Crosstalk between growth hormone and insulin signaling. Vitam Horm 2009; 80:125–53; PMID:19251037; http://dx.doi.org/ 10.1016/S0083-6729(08)00606-7 [DOI] [PubMed] [Google Scholar]
- [4].Lichanska AM, Waters MJ. New insights into growth hormone receptor function and clinical implications. Horm Res 2008; 69:138–45; PMID:18219216; http://dx.doi.org/ 10.1159/000112586 [DOI] [PubMed] [Google Scholar]
- [5].Carter-Su C, Schwartz J, Argetsinger LS. Growth hormone signaling pathways. Growth Horm IGF Res 2015; http://dx.doi.org/ 10.1016/j.ghir.2015.09.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Chia DJ. Minireview: mechanisms of growth hormone-mediated gene regulation. Mol Endocrinol 2014; 28:1012–25; PMID:24825400; http://dx.doi.org/ 10.1210/me.2014-1099 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Barclay JL, Kerr LM, Arthur L, Rowland JE, Nelson CN, Ishikawa M, d'Aniello EM, White M, Noakes PG, Waters MJ. In vivo targeting of the growth hormone receptor (GHR) Box1 sequence demonstrates that the GHR does not signal exclusively through JAK2. Mol Endocrinol 2010; 24:204–17; PMID:19884384; http://dx.doi.org/ 10.1210/me.2009-0233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Rotwein P. Mapping the growth hormone–Stat5b–IGF-I transcriptional circuit. Trends Endocrinol Metab 2012; 23:186–93; PMID:22361342; http://dx.doi.org/ 10.1016/j.tem.2012.01.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Woelfle J, Rotwein P. In vivo regulation of growth hormone-stimulated gene transcription by STAT5b. Am J Physiol Endocrinol Metab 2004; 286:E393–401; PMID:14761873; http://dx.doi.org/ 10.1152/ajpendo.00389.2003 [DOI] [PubMed] [Google Scholar]
- [10].Gao B, Roux PP. Translational control by oncogenic signaling pathways. Biochim Biophys Acta 2015; 1849:753–65; PMID:25477072; http://dx.doi.org/3383777 10.1016/j.bbagrm.2014.11.006 [DOI] [PubMed] [Google Scholar]
- [11].Mathews LS, Hammer RE, Brinster RL, Palmiter RD. Expression of insulin-like growth factor I in transgenic mice with elevated levels of growth hormone is correlated with growth. Endocrinology 1988; 123:433–7; PMID:3383777; http://dx.doi.org/ 10.1210/endo-123-1-433 [DOI] [PubMed] [Google Scholar]
- [12].McGrane MM, Yun JS, Moorman AF, Lamers WH, Hendrick GK, Arafah BM, Park EA, Wagner TE, Hanson RW. Metabolic effects of developmental, tissue-, and cell-specific expression of a chimeric phosphoenolpyruvate carboxykinase (GTP)/bovine growth hormone gene in transgenic mice. J Biol Chem 1990; 265:22371–9; PMID:1702419 [PubMed] [Google Scholar]
- [13].Martinez CS, Piazza VG, Diaz ME, Boparai RK, Arum O, Ramirez MC, Gonzalez L, Becu-Villalobos D, Bartke A, Turyn D, et al.. Growth hormone (GH)/STAT5 signaling during the growth period in liver of mice overexpressing GH. J Mol Endocrinol 2015; 54(2):171–84; PMID:25691498 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Palmiter RD, Brinster RL, Hammer RE, Trumbauer ME, Rosenfeld MG, Birnberg NC, Evans RM. Dramatic growth of mice that develop from eggs microinjected with metallothionein-growth hormone fusion genes. Nature 1982; 300:611–5; PMID:6958982; http://dx.doi.org/ 10.1038/300611a0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Searle TW, Murray JD, Baker PJ. Effect of increased production of growth hormone on body composition in mice: transgenic versus control. J Endocrinol 1992; 132:285–91; PMID:1541927; http://dx.doi.org/ 10.1677/joe.0.1320285 [DOI] [PubMed] [Google Scholar]
- [16].Miquet JG, Freund T, Martinez CS, Gonzalez L, Diaz ME, Micucci GP, Zotta E, Boparai RK, Bartke A, Turyn D, et al.. Hepatocellular alterations and dysregulation of oncogenic pathways in the liver of transgenic mice overexpressing growth hormone. Cell Cycle 2013; 12:1042–57; PMID:23428905; http://dx.doi.org/ 10.4161/cc.24026 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Miquet JG, Gonzalez L, Matos MN, Hansen CE, Louis A, Bartke A, Turyn D, Sotelo AI. Transgenic mice overexpressing GH exhibit hepatic upregulation of GH-signaling mediators involved in cell proliferation. J Endocrinol 2008; 198:317–30; PMID:18480380; http://dx.doi.org/ 10.1677/JOE-08-0002 [DOI] [PubMed] [Google Scholar]
- [18].Miquet JG, Sotelo AI, Bartke A, Turyn D. Suppression of growth hormone (GH) Janus tyrosine kinase 2/signal transducer and activator of transcription 5 signaling pathway in transgenic mice overexpressing bovine GH. Endocrinology 2004; 145:2824–32; PMID:15016718; http://dx.doi.org/ 10.1210/en.2003-1498 [DOI] [PubMed] [Google Scholar]
- [19].Brem G, Wanke R, Wolf E, Buchmuller T, Muller M, Brenig B, Hermanns W. Multiple consequences of human growth hormone expression in transgenic mice. Mol Biol Med 1989; 6:531–47; PMID:263481311316754 [PubMed] [Google Scholar]
- [20].Hoeflich A, Nedbal S, Blum WF, Erhard M, Lahm H, Brem G, Kolb HJ, Wanke R, Wolf E. Growth inhibition in giant growth hormone transgenic mice by overexpression of insulin-like growth factor-binding protein-2. Endocrinology 2001; 142:1889–98; PMID:11316754; http://dx.doi.org/ 10.1210/endo.142.5.8149 [DOI] [PubMed] [Google Scholar]
- [21].Wanke R, Hermanns W, Folger S, Wolf E, Brem G. Accelerated growth and visceral lesions in transgenic mice expressing foreign genes of the growth hormone family: an overview. Pediatr Nephrol 1991; 5:513–21; PMID:1911131; http://dx.doi.org/ 10.1007/BF01453693 [DOI] [PubMed] [Google Scholar]
- [22].Shea BT, Hammer RE, Brinster RL. Growth allometry of the organs in giant transgenic mice. Endocrinology 1987; 121:1924–30; PMID:3678132; http://dx.doi.org/ 10.1210/endo-121-6-1924 [DOI] [PubMed] [Google Scholar]
- [23].Orian JM, Lee CS, Weiss LM, Brandon MR. The expression of a metallothionein-ovine growth hormone fusion gene in transgenic mice does not impair fertility but results in pathological lesions in the liver. Endocrinology 1989; 124:455–63; PMID:2642419; http://dx.doi.org/ 10.1210/endo-124-1-455 [DOI] [PubMed] [Google Scholar]
- [24].Snibson KJ. Hepatocellular kinetics and the expression of growth hormone (GH) in the livers and liver tumours of GH-transgenic mice. Tissue Cell 2002; 34:88–97; PMID:12165243; http://dx.doi.org/ 10.1016/S0040-8166(02)00012-5 [DOI] [PubMed] [Google Scholar]
- [25].Snibson KJ, Bhathal PS, Hardy CL, Brandon MR, Adams TE. High, persistent hepatocellular proliferation and apoptosis precede hepatocarcinogenesis in growth hormone transgenic mice. Liver 1999; 19:242–52; PMID:10395045; http://dx.doi.org/ 10.1111/j.1478-3231.1999.tb00042.x [DOI] [PubMed] [Google Scholar]
- [26].Orian JM, Tamakoshi K, Mackay IR, Brandon MR. New murine model for hepatocellular carcinoma: transgenic mice expressing metallothionein-ovine growth hormone fusion gene. J Natl Cancer Inst 1990; 82:393–8; PMID:2154583; http://dx.doi.org/ 10.1093/jnci/82.5.393 [DOI] [PubMed] [Google Scholar]
- [27].He G, Karin M. NF-kappaB and STAT3 - key players in liver inflammation and cancer. Cell Res 2011; 21:159–68; PMID:21187858; http://dx.doi.org/ 10.1038/cr.2010.183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Llovet JM, Bruix J. Molecular targeted therapies in hepatocellular carcinoma. Hepatology 2008; 48:1312–27; PMID:18821591; http://dx.doi.org/ 10.1002/hep.22506 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Martinez CS, Piazza VG, Ratner LD, Matos MN, Gonzalez L, Rulli SB, Miquet JG, Sotelo AI. Growth hormone STAT5-mediated signaling and its modulation in mice liver during the growth period. Growth Horm IGF Res 2013; 23:19–28; PMID:23245546; http://dx.doi.org/ 10.1016/j.ghir.2012.11.002 [DOI] [PubMed] [Google Scholar]
- [30].Popow A, Nowak D, Malicka-Blaszkiewicz M. Actin cytoskeleton and beta-actin expression in correlation with higher invasiveness of selected hepatoma Morris 5123 cells. J Physiol Pharmacol 2006; 57(Suppl 7):111–23; PMID:17228099 [PubMed] [Google Scholar]
- [31].Miller LM, Menthena A, Chatterjee C, Verdier-Pinard P, Novikoff PM, Horwitz SB, Angeletti RH. Increased levels of a unique post-translationally modified betaIVb-tubulin isotype in liver cancer. Biochemistry 2008; 47:7572–82; PMID:18570381; http://dx.doi.org/ 10.1021/bi8005225 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Lui JC, Baron J. Mechanisms limiting body growth in mammals. Endocr Rev 2011; 32:422–40; PMID:21441345; http://dx.doi.org/ 10.1210/er.2011-0001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Stanger BZ. The biology of organ size determination. Diabetes 2008; 10(Suppl 4):16–22; http://dx.doi.org/ 10.1111/j.1463-1326.2008.00938.x [DOI] [PubMed] [Google Scholar]
- [34].Quaife CJ, Mathews LS, Pinkert CA, Hammer RE, Brinster RL, Palmiter RD. Histopathology associated with elevated levels of growth hormone and insulin-like growth factor I in transgenic mice. Endocrinology 1989; 124:40–8; PMID:2642417; http://dx.doi.org/ 10.1210/endo-124-1-40 [DOI] [PubMed] [Google Scholar]
- [35].Bartke A. Can growth hormone (GH) accelerate aging? Evidence from GH-transgenic mice. Neuroendocrinology 2003; 78:210–6; PMID:14583653; http://dx.doi.org/ 10.1159/000073704 [DOI] [PubMed] [Google Scholar]
- [36].Vergani G, Mayerhofer A, Bartke A. Acute effects of human growth hormone on liver cells in vitro: a comparison with livers of mice transgenic for human growth hormone. Tissue Cell 1991; 23:607–12; PMID:1776152; http://dx.doi.org/ 10.1016/0040-8166(91)90017-N [DOI] [PubMed] [Google Scholar]
- [37].Takahashi-Yanaga F, Sasaguri T. GSK-3beta regulates cyclin D1 expression: a new target for chemotherapy. Cell Signal 2008; 20:581–9; PMID:18023328; http://dx.doi.org/ 10.1016/j.cellsig.2007.10.018 [DOI] [PubMed] [Google Scholar]
- [38].Stamatakos M, Palla V, Karaiskos I, Xiromeritis K, Alexiou I, Pateras I, Kontzoglou K. Cell cyclins: triggering elements of cancer or not? World J Surg Oncol 2010; 8:111; PMID:21176227; http://dx.doi.org/ 10.1186/1477-7819-8-111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Feitelson MA, Sun B, Satiroglu Tufan NL, Liu J, Pan J, Lian Z. Genetic mechanisms of hepatocarcinogenesis. Oncogene 2002; 21:2593–604; PMID:11971194; http://dx.doi.org/ 10.1038/sj.onc.1205434 [DOI] [PubMed] [Google Scholar]
- [40].Bakiri L, Lallemand D, Bossy-Wetzel E, Yaniv M. Cell cycle-dependent variations in c-Jun and JunB phosphorylation: a role in the control of cyclin D1 expression. EMBO J 2000; 19:2056–68; PMID:10790372; http://dx.doi.org/ 10.1093/emboj/19.9.2056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Bretones G, Delgado MD, Leon J. Myc and cell cycle control. Biochim Biophy Acta 2015; 1849:506–16; PMID:24704206; http://dx.doi.org/ 10.1016/j.bbagrm.2014.03.013 [DOI] [PubMed] [Google Scholar]
- [42].Hsieh AL, Walton ZE, Altman BJ, Stine ZE, Dang CV. MYC and metabolism on the path to cancer. Semin Cell Dev Biol 2015; 43:11–21; PMID:26277543 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Zhu T, Ling L, Lobie PE. Identification of a JAK2-independent pathway regulating growth hormone (GH)-stimulated p44/42 mitogen-activated protein kinase activity. GH activation of Ral and phospholipase D is Src-dependent. J Biol Chem 2002; 277:45592–603; PMID:12218045; http://dx.doi.org/ 10.1074/jbc.M201385200 [DOI] [PubMed] [Google Scholar]
- [44].Rowlinson SW, Yoshizato H, Barclay JL, Brooks AJ, Behncken SN, Kerr LM, Millard K, Palethorpe K, Nielsen K, Clyde-Smith J, et al.. An agonist-induced conformational change in the growth hormone receptor determines the choice of signalling pathway. Nat Cell Biol 2008; 10:740–7; PMID:18488018; http://dx.doi.org/ 10.1038/ncb1737 [DOI] [PubMed] [Google Scholar]
- [45].Roskoski R., Jr Src protein-tyrosine kinase structure, mechanism, and small molecule inhibitors. Pharmacol Res 2015; 94:9–25; http://dx.doi.org/ 10.1016/j.phrs.2015.01.003 [DOI] [PubMed] [Google Scholar]
- [46].Bjorge JD, Jakymiw A, Fujita DJ. Selected glimpses into the activation and function of Src kinase. Oncogene 2000; 19:5620–35; PMID:11114743; http://dx.doi.org/ 10.1038/sj.onc.1203923 [DOI] [PubMed] [Google Scholar]
- [47].Boggon TJ, Eck MJ. Structure and regulation of Src family kinases. Oncogene 2004; 23:7918–27; PMID:15489910; http://dx.doi.org/ 10.1038/sj.onc.1208081 [DOI] [PubMed] [Google Scholar]
- [48].Roskoski R, Jr. Src kinase regulation by phosphorylation and dephosphorylation. Biochem Biophys Res Commun 2005; 331:1–14; PMID:15845350; http://dx.doi.org/ 10.1016/j.bbrc.2005.03.012 [DOI] [PubMed] [Google Scholar]
- [49].Aggarwal BB, Kunnumakkara AB, Harikumar KB, Gupta SR, Tharakan ST, Koca C, Dey S, Sung B. Signal transducer and activator of transcription-3, inflammation, and cancer: how intimate is the relationship? Ann N Y Acad Sci 2009; 1171:59–76; PMID:19723038; http://dx.doi.org/ 10.1111/j.1749-6632.2009.04911.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Berasain C, Castillo J, Perugorria MJ, Latasa MU, Prieto J, Avila MA. Inflammation and liver cancer: new molecular links. Ann N Y Acad Sci 2009; 1155:206–21; PMID:19250206; http://dx.doi.org/ 10.1111/j.1749-6632.2009.03704.x [DOI] [PubMed] [Google Scholar]
- [51].Friedbichler K, Themanns M, Mueller KM, Schlederer M, Kornfeld JW, Terracciano LM, Kozlov AV, Haindl S, Kenner L, Kolbe T, et al.. Growth-hormone-induced signal transducer and activator of transcription 5 signaling causes gigantism, inflammation, and premature death but protects mice from aggressive liver cancer. Hepatology 2012; 55:941–52; PMID:22031092; http://dx.doi.org/ 10.1002/hep.24765 [DOI] [PubMed] [Google Scholar]
- [52].Horiguchi N, Wang L, Mukhopadhyay P, Park O, Jeong WI, Lafdil F, Osei-Hyiaman D, Moh A, Fu XY, Pacher P, et al.. Cell type-dependent pro- and anti-inflammatory role of signal transducer and activator of transcription 3 in alcoholic liver injury. Gastroenterology 2008; 134:1148–58; PMID:18395093; http://dx.doi.org/ 10.1053/j.gastro.2008.01.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Sakamori R, Takehara T, Ohnishi C, Tatsumi T, Ohkawa K, Takeda K, Akira S, Hayashi N. Signal transducer and activator of transcription 3 signaling within hepatocytes attenuates systemic inflammatory response and lethality in septic mice. Hepatology 2007; 46:1564–73; PMID:17705264; http://dx.doi.org/ 10.1002/hep.21837 [DOI] [PubMed] [Google Scholar]
- [54].Gonzalez L, Diaz ME, Miquet JG, Sotelo AI, Fernandez D, Dominici FP, Bartke A, Turyn D. GH modulates hepatic epidermal growth factor signaling in the mouse. J Endocrinol 2010; 204:299–309; PMID:20032199; http://dx.doi.org/ 10.1677/JOE-09-0372 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Sturgill TW. MAP kinase: it's been longer than fifteen minutes. Biochem Biophys Res Commun 2008; 371:1–4; PMID:18406346; http://dx.doi.org/ 10.1016/j.bbrc.2008.04.002 [DOI] [PubMed] [Google Scholar]
- [56].Faes S, Dormond O. PI3K and AKT: Unfaithful partners in cancer. Int J Mol Sci 2015; 16:21138–52; PMID:26404259; http://dx.doi.org/ 10.3390/ijms160921138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Regad T. Targeting RTK Signaling Pathways in Cancer. Cancers 2015; 7:1758–84; PMID:26404379; http://dx.doi.org/ 10.3390/cancers7030860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Jacobs KM, Bhave SR, Ferraro DJ, Jaboin JJ, Hallahan DE, Thotala D. GSK-3beta: A Bifunctional Role in Cell Death Pathways. Int J Cell Biol 2012; 2012:930710; PMID:22675363; http://dx.doi.org/ 10.1155/2012/930710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Sekulic A, Hudson CC, Homme JL, Yin P, Otterness DM, Karnitz LM, Abraham RT. A direct linkage between the phosphoinositide 3-kinase-AKT signaling pathway and the mammalian target of rapamycin in mitogen-stimulated and transformed cells. Cancer Res 2000; 60:3504–13; PMID:10910062 [PubMed] [Google Scholar]
- [60].Liko D, Hall MN. mTOR in health and in sickness. J Mol Med (Berl) 2015; 93:1061–73; PMID:26391637; http://dx.doi.org/ 10.1007/s00109-015-1326-7 [DOI] [PubMed] [Google Scholar]
- [61].Song G, Ouyang G, Bao S. The activation of Akt/PKB signaling pathway and cell survival. J Cell Mol Med 2005; 9:59–71; PMID:15784165; http://dx.doi.org/ 10.1111/j.1582-4934.2005.tb00337.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Villanueva A, Chiang DY, Newell P, Peix J, Thung S, Alsinet C, Tovar V, Roayaie S, Minguez B, Sole M, et al.. Pivotal role of mTOR signaling in hepatocellular carcinoma. Gastroenterology 2008; 135:1972–83, 83 e1-11; PMID:18929564; http://dx.doi.org/ 10.1053/j.gastro.2008.08.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Haga S, Ozaki M, Inoue H, Okamoto Y, Ogawa W, Takeda K, Akira S, Todo S. The survival pathways phosphatidylinositol-3 kinase (PI3-K)/phosphoinositide-dependent protein kinase 1 (PDK1)/Akt modulate liver regeneration through hepatocyte size rather than proliferation. Hepatology 2009; 49:204–14; PMID:19065678; http://dx.doi.org/ 10.1002/hep.22583 [DOI] [PubMed] [Google Scholar]
- [64].Ono H, Shimano H, Katagiri H, Yahagi N, Sakoda H, Onishi Y, Anai M, Ogihara T, Fujishiro M, Viana AY, et al.. Hepatic Akt activation induces marked hypoglycemia, hepatomegaly, and hypertriglyceridemia with sterol regulatory element binding protein involvement. Diabetes 2003; 52:2905–13; PMID:14633850; http://dx.doi.org/ 10.2337/diabetes.52.12.2905 [DOI] [PubMed] [Google Scholar]