

Abstract
For this study, we cultured chondrocyte pellets in Dulbecco’s modified Eagle’s medium plus a 2 % fetal bovine serum medium, and treated them with 2- to 8-mer oligosaccharides of chondroitin sulfate A to examine the effects of these oligosaccharides on the differentiation and protection of chondrocytes. We found low-molecular-weight CSAs to increase the ratio of the gene expression levels of collagen II/collagen I of chondrocytes from the first day up to 14 days after culture compared with those under a CSA-free medium. Moreover, low-molecular-weight CSAs inhibited the expression of matrix metalloproteinases and peptidases, and stimulated an endogenous tissue inhibitor of metalloproteinases. The dp-8 (8-mer) CSA yielded the most effective response among promoting collagen type II protein secretions compared with other groups.
Electronic supplementary material
The online version of this article (doi:10.1007/s10616-014-9788-y) contains supplementary material, which is available to authorized users.
Keywords: Chondroitin-4-sulfate, Chondrocyte, Collagen, Metalloproteinase, Gene
Introduction
Articular cartilage (AC) is a specialized tissue, the majority of which contains chondrocytes. The extracellular matrix (ECM) of cartilage contains proteoglycans (PGs), collagen, and non-collagen proteins, forming a dense layer of tissue that protects against mechanical stress.
Glycosaminoglycans (GAGs), the constituents of PGs, play a critical role in several biological functions including proliferation, cells cycle, cells adhesion, homeostasis, and lipid metabolism (Campo et al. 2009). Chondroitin sulfate (CS), which is a major class of GAGs in ECM cartilage, appears as a long, unbranched polysaccharide chain with a repetitive disaccharide structure of N-acetylgalactosamine and glucuronic acid. Depending on the sulfate position in a disaccharide, CS is classified as chondroitin-4-sulfate (CSA; Fig. 1), chondroitin-6-sulfate (CSC), or dermatan sulfate (DS). Previous studies have shown that CS is a therapeutic agent for treating osteoarthritis (OA), and relieves pain, enhances the functional activity of chondrocytes, and slows disease progression (Uebelhart 2008; Tiraloche et al. 2005; Fischer 1996; Michel et al. 2005). However, several studies have presented inconsistent results regarding the therapeutic effects of CS in treating OA (Fox and Stephens 2009). CS is still commonly used as a chondroprotective drug, and is recognized in Europe as a symptomatic, slow-acting drug for OA treatment. OA is characterized by the loss of AC in focal areas, with varying degrees of osteophyte formation, subchondral bone changes, and synovitis (Young et al. 2005; Sokoloff 1987; Aigner et al. 2006). The three CS mechanisms that potentially influence osteoarthritis are as follows: (1) the anti-inflammatory effect on cellular inflammation; (2) the promotion of cartilage-producing PGs; and (3) the reduced degradation of the ECM (Iovu et al. 2008).
Fig. 1.
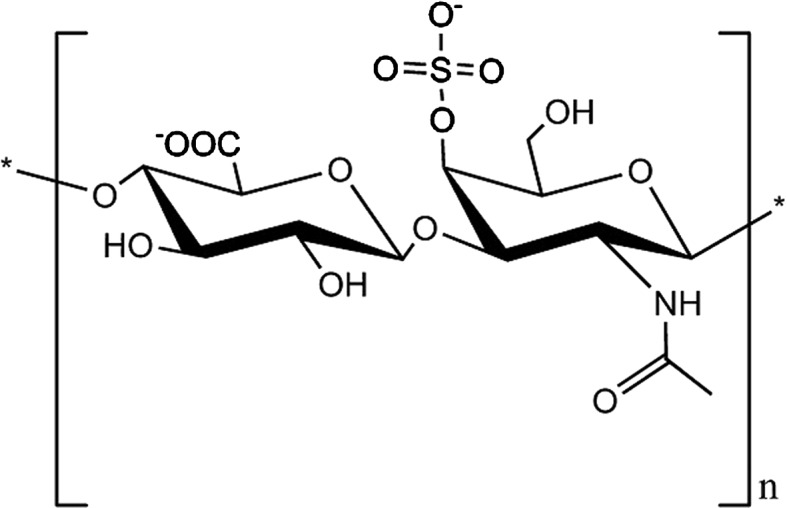
Structure of chondroitin-4-sulfate (CSA)
During the in vitro expansion of chondrocytes, the cells are prone to dedifferentiate when grown on 2D surfaces, and attenuate their original phenotype, whereas cultivation on a 3D scaffold can restore chondrocyte properties (Kafienah et al. 2002). In a pellet culture, which has the form of a 3D culture, the chondrocyte phenotype is maintained, and ECM synthesis is promoted (Holtzer et al. 1960).
Chen et al. (2011a) discussed the effect of oligosaccharide CSCs on chondrocytes with a collagen type II scaffold. Their results revealed that oligosaccharide CSCs can enhance the chondrogenic gene expression of chondrocytes when grown on a collagen type II scaffold. The purpose of the present study is to investigate the effects of CSAs in vitro on chondrocyte pellet cultures. CSC and CSA are structurally different, and an investigation to determine whether their effects on chondrocytes differ as well is warranted.
Materials and methods
Preparing CSAs of different molecular weights
Low-molecular-weight CSAs were prepared by degrading high-molecular-weight CSAs (Sigma, St. Louis, MO, USA), by executing a reaction of chondroitinase ABC (Sigma) for 6 h (Radhakrishnamurthy et al. 1986). In brief, 240 mg of CSAs was dissolved in 1 mL of a 0.05 M Tris–HCl (Sigma) buffer containing 100 µg of bovine serum albumin (Sigma). Afterward, 1.7 U of chondroitinase ABC was added to 1 mL of the CSA buffer, which was incubated with shaking at 37 °C for 6 h. The digestion reaction was initiated at 100 °C for 5 min, and then cooled to room temperature. The impure and insoluble matter was removed using a 0.22-µm filter. Different molecular weights of CSA were separated through fast-protein liquid chromatography (FPLC, GE Healthcare, Pittsburgh, PA, USA) with a Superdex 26/60 column (GE Healthcare). The molecular weight of each fraction was measured using gel permeation chromatography (GPC, JASCO, Portland, OR, USA), and was expressed as the mean ± standard deviation, as detailed in Table 1 in the Supplementary data.
Chondrocyte pellet culture
Normal human articular chondrocytes were obtained from Camber Bio-Science (Walkersville, MD, USA). Cells in frozen state were thawed and suspended in a growth medium (CGM) composed of Dulbecco’s modified Eagle’s medium (DMEM): Nutrient Mixture F-12 Medium (1:1), 10 % fetal bovine serum, and 1 % penicillin/streptomycin. The chondrocytes were seeded in a culture flask with 104 cells/cm2 and enrichment-cultured for 7 day. The cells were trypsinized and collected with a density of 3 × 105 cells/mL of CGM in each 15-mL tube, and centrifuged at 500g for 10 min at 4 °C. Afterward, the cells in the tube were cultured at 37 °C in a 5 % CO2 atmosphere, and the pellets formed within 3 days of culturing. The pellets were cultured in DMEM with 2 % fetal bovine serum, 0.5 % insulin-transferrin-selenium (ITS), 1 % penicillin, and 50 µM/mL of CSAs with varying molecular weights. All medium components were purchased from Gibco-Life Technologies (Grand Island, NY, USA).
Histological analysis
The cell pellets that were cultured in the medium with or without CSAs with varying molecular weights for 14 days and were fixed in 10 % (v/v) phosphate buffered formaldehyde. The samples were then dehydrated using an ethanol gradient of 50 % (v/v), 70 % (v/v), 95 % (v/v), and 100 % (v/v) ethanol, and were embedded in paraffin in 8-μm-thick sections. Finally, these sections were subjected to Hematoxylin and Eosin staining (H&E) (Sigma), Alcian blue staining (Sigma), and immunohistochemistry staining. The histological characteristics were observed under a TS100 optical microscope (Nikon, Melville, NY, USA).
Immunohistochemistry staining
The sample sections were washed with xylene, 100, 95, 70, and 50 % ethanol, and PBS. They were treated with Dako cytomation Target Retrieval Solution (Dako Cytomation, Carpinteria, CA, USA) for 20 min at 100 °C with a pH level of 6.0. They were then blocked using 5 % goat serum (Gibco) for 1 h at 37 °C. A collagen type II (COL II) (Abcam, Cambridge, MA, USA) antibody (1:100) was applied to these slides, which were left overnight at 4 °C. The slides were then treated with anti-rabbit-FITC (1:150; The Jackson Laboratory, Bar Harbor, ME, USA) for 1.5 h at 37 °C, and were counterstained with propidium iodide for 10 min (Sigma). The histological characteristics were observed using an inverted confocal laser-scanning microscope (CLSM, TCS SL, Leica, Wetzlar, Germany), with excitations at 488, 543, and 633 nm. Superimposed imaging was performed using the LCS Lite software (version 2.0, Leica; Meech et al. 2005; Vinardell et al. 2009).
Quantifying total glycosaminoglycans
The chondrocyte pellets that were harvested on days 3, 7, 10, and 14 were digested using a papain (Sigma) digesting solution for 24 h at 60 °C. The total GAG content of the pellets was analyzed using 200 µL of 1,9-dimethyl methylene blue solution and 8-µL aliquots of a papain digesting solution. The optical density was determined at 530 nm.
RNA isolation, cDNA synthesis, and real-time PCR
Total RNA was isolated from the chondrocyte pellets by using the trizol method (Invitrogen, Carlsbad, CA, USA). In brief, the pellets were washed with PBS, and 1 mL of trizol was added to each sample and homogenized using the PowerGen 125 Tissue Homogenizer for 30–60 s. The trizol mixture was centrifuged, and the supernatant was precipitated using a series of solvents before quantification, which was performed with the Quant-iT™ RNA Assay Kit (Molecular Probes, Eugene, OR, USA). The total RNA was then reverse-transcribed to cDNA by using the RevertAid H minus First-Strand cDNA Synthesis Kit (Fermentas International Inc., Burlington, ON, Canada), according to the manufacturer instructions.
Real-time PCR assays were performed using TaqMan® Universal PCR Master Mix (Applied Biosystems, Forster City, CA, USA), according to the manufacturer instructions, on an ABI Prism 7300 Real Time PCR System (Applied Biosystems). Primers and probes for human GAPDH (NM_002046, 70 bp), collagen type I (NM_000088, 66 bp), collagen type II (NM_033150, 124 bp), Aggrecan (NM_013227, 93 bp), a disintegrin and metalloproteinase with thrombospondins-5 (ADAMTS5) (NM_007038, 65 bp), a disintegrin and metalloproteinase with thrombospondins-4 (ADAMTS4) (NM_005099, 63 bp), a tissue inhibitor of matrix metalloproteinase (TIMP3) (NM_000362, 59 bp), metalloproteinase-1 (MMP1) (NM_002412, 64 bp), and metalloproteinase-13 (MMP13) (NM_002427, 91 bp) were obtained from Applied Biosystems. GAPDH was used as the house-keeping gene and the internal control. The mRNA gene expression in CSA- or non-CSA-treated chondrocyte cell pellets was calculated relative to the gene expression in the control chondrocyte cells on a 2D culture, in accordance with the delta–delta Ct method (2−ΔΔCt; Schmittgen and Livak 2008).
Statistical analysis
All data were represented as the mean ± standard deviation. Statistical analysis was performed using one-way analysis of variance (ANOVA), followed by pairwise comparisons of the sample means by using Student’s t test to determine the differences among groups. The results were considered to be significantly different with a P value <0.05. All statistical analyses were performed using SPSS version 20 (IBM, New York, NY, USA). When analyzing the biological gradients, a relative quantitation of CSA oligosaccharides was used as the dummy variable, and the JMP Cochran Armitage Trend Test, version 10 (SAS Inc., Cary, NC, USA), was conducted.
Results
Oligosaccharide CSA preparations
Chondroitinase ABC was used to digest high-molecular-weight CSAs into oligosaccharides in 2-, 4-, or 8-glycan length segments, which were denoted as dp-2, dp-4, and dp-8, respectively. They were separated according to the molecular weight and purified using FPLC. The molecular weight of each fraction was confirmed using GPC, as shown in Table 1 in the Supplementary data.
Morphology and ECM-component analysis
The morphology and ECM staining of the pellet culture of chondrocytes at Day 14 are shown in Fig. 2. The chondrocytes were found to have distributed evenly within the pellets, as revealed by the results of H&E staining. The lacuna and round morphology of typical chondrocytes was more prominent in the CSA-supplemented groups of dp-4 and dp-8, as shown by a comparison of Fig. 2a, b with Fig. 2c, d. Regarding COL II staining (shown in Fig. 2i–l), the dp-8 group exhibited greater COL II formation, and a quantitative comparison is shown in Fig. 2n, from the COL II fluorescence area percentage for the different treatment groups. Therefore, oligo-CSA can clearly help stimulate more COL II formation in pellets. Specifically, on day 14, the dp-8 group exhibited the highest COL II content, and the CSA-supplemented groups exhibited a higher COL II level overall compared with the CSA-free group. The GAG content, when observed under Alcian blue staining (shown in Fig. 2e–h) revealed no statistical difference between CSA-free and CSA-treated chondrocyte pellets. This can be confirmed by measuring GAGs by using the dimethyl methylene blue method. In Fig. 3a, the total amount of GAGs is shown, whereas in Fig. 3b, GAG levels with respect to the DNA level are shown. The data indicated that the GAG content of all groups continued to increase until day 10, and then decreased on day 14, with no significant difference emerging between them.
Fig. 2.

a–d H&E stains (×400, scale bar 100 μm) of the pellet culture on day 14. e–h Alcian blue stains (×400, scale bar 100 μm) of the pellet culture on day 14, and immunochemistry stains of collagen type II (×63, blue collagen type II; red the cell nucleus) of the pellet culture on day 14. a, e, and i CSA-free medium group; b, f, and j medium group supplemented with dp-2 CSA; c, g, and k medium group supplemented with dp-4 CSA; d, h, and l medium supplemented with dp-8 CSA. n represents the collagen type II protein area percentage of fluorescent immunochemistry
Fig. 3.

a Content of glycosaminoglycans (GAGs) (n = 6), b GAG/DNA distribution (n = 6). Dp0 CSA-free treatment group, Dp2 dp2-CSA treatment group, Dp4 dp4-CSA treatment group, and Dp8 dp8-treatment group
Gene expression profiles
Aggrecan (AGC) is a characteristic marker for chondrogenesis. We found AGC expression to have upregulated for the dp-8 groups, especially on day 10 and day 14 of the pellet culture, as shown in Fig. 4a. No significant differences emerged among the AGC gene expression levels on days 7, 10, and 14, and for these groups (other than dp-8), the expression levels were relatively constant during the 2-week cultivation. This finding may seem contrary to the GAG content results displayed in Fig. 3, which shows that all groups had similar GAG levels. One explanation for this discrepancy is that AGC gene expression does not correspond exactly to the GAG level, where GAG components other than aggrecans are involved. In this part of the study, dp-8 was found to have a positive effect on the chondrogenic AGC gene expression.
Fig. 4.

a Aggrecan gene expression in chondrocyte pellets, b Collagen Type II gene expression in chondrocyte pellets, c Collagen Type I gene expression in chondrocyte pellets, d Collagen Type II/Collagen Type I gene expression ratio (n = 5) in chondrocyte pellets. Significance: # P < 0.05 compared with dp2-CSA and CSA-free groups; $ P < 0.05 compared with the dp4-CSA and CSA-free groups; * P < 0.05 compared with the dp8-CSA and CSA-free groups. The relative quantification was reflects the change in gene expression in the experimental groups relative to the CSA-free treatment group on day 3
Figure 4b–d show the effects of CSA on COL I and COL II gene expression levels. The gene expression of COL II, which is one gene that is essential for chondrogenesis, was found to be influenced by CSA. As shown in Fig. 4b, all of the groups, including the CSA-free group, exhibited gradual increases in COL II expression during the 2-week culture. The highest COL II expression occurred on day 14 in the CSA-free group, whereas a higher molecular weight of CSA oligosaccharides produced a higher expression among the CSA-supplement groups (dp-8 > dp-4 = dp-2).
The apparent upregulation of COL II genes in the CSA-free group on day 14 seems to contradict the assumption that CSA can enhance chondrogenesis. However, when examining COL I gene expression, we found that the CSA-free group exhibited a more enhanced expression of this gene on day 14 compared with the other groups (Fig. 4c). Based on the ratio of COL II/COL I expression (a substantially clearer indicator of chondrogenesis) in these groups (Fig. 4d), this ratio was obviously superior for CSA-supplemented groups than for the CSA-free group. The differently sized CSA oligosaccharides performed largely similarly; the dp-8 groups had the highest ratios on day 10, and even higher ratios on day 14.
In Fig. 5, data are presented for the expression levels of enzymes that regulate the degradation of the cartilage ECM. This may further elucidate the mechanisms of CSA effects on chondrocytes. On day 14, the CSA-free group had a slightly higher ADAMTS-4 gene expression level and an extremely high (40-fold greater) level of ADAMTS-5 gene expression compared with the CSA-supplemented groups, as shown in Fig. 5a, b. In addition, the CSA-free group exhibited a substantially stronger upregulation (greater than tenfold) of MMP1 and MMP13 genes compared with the CSA-supplemented groups, as shown in Fig. 5d. Figure 5c shows the expression levels of TIMP3 on day 14. The TIMPs constitute a family of MMP inhibitors. The TIMP3 inhibitor can also inhibit ADAMTS-4 and ADAMTS-5. The expression of TIMP3 was considerably upregulated in the CSA-supplemented groups on day 14, with the dp-2 group exhibiting the highest level. Overall, these data showed that CSA oligosaccharides hinder the expression of degrading enzymes, and upregulate the enzymes that inhibit their degrading counterparts. This means that CSA oligosaccharides favor ECM synthesis and chondrogenesis. The various sizes of CSA oligosaccharides may have distinct effects such as dp-2 enhancing TIMP3 and dp-8 downregulating most degrading enzymes more than the CSA oligosaccharides of other sizes; however, their exact molecular mechanism remains unknown.
Fig. 5.

a Disintegrin and Metalloproteinase with Thrombospondins, Tissue Inhibitors of Matrix Metalloproteinase 3, and the Metalloproteinase family gene expression by chondrocyte pellets. a ADAMST-4, b ADAMST-5, c TIMP3, d the gene expression of the MMP family on day 14 (n = 5), Significance: # P < 0.05 compared with the dp2-CSA and CSA-free groups; $ P < 0.05 compared with the dp4-CSA and CSA-free groups; and *P < 0.05 compared with the dp8-CSA and CSA-free groups. The relative quantification reflects the change in gene expression in the experimental groups relative to the CSA-free treatment group on day 3
Discussion
The GAGs, such as glucosamine-like substances and CS, have recently been investigated for their possible beneficial effects on OA. A common observation is that these substances seem to facilitate cartilage repair by reducing inflammation and blocking the loss of PGs (Kupcsik et al. 2010; Derfoul et al. 2007; Kagita et al. 2010). However, no comprehensive mechanism by which GAGs and CS can be administered to moderate the side effects of OA has been elucidated. Furthermore, previous studies have focused on large-molecular-weight GAGs and CS. This study focused instead on low-molecular-weight CSAs, and our findings revealed that low-molecular-weight CSAs helped chondrocytes maintain their normal phenotype.
A previous report indicated that high-molecular-weight CSAs did not affect COL II expression, but that high-molecular-weight CSC could enhance it (Nishimoto et al. 2005). In our study, low-molecular-weight CSAs increased the ratio of COL II/COL I gene expression levels, and the dp-8 CSA was most effective in stimulating aggrecan and COL II gene expression.
This study presented the quantitative data regarding the downregulating effects of CSA oligosaccharides on ADAMTS-5 and ADAMTS-4 gene expression, which were reported as a key factor in instability-induced knee OA in mice (Flannery 2010). We found that different molecular weights of CSAs can upregulate TIMP-3, thereby reducing the activity of various ADAMSTs (Troeberg et al. 2008). The MMP-1 gene can degrade collagen, and MMP-13 has a particular affinity for type II collagen, but can also cleave aggrecan (Fosang et al. 1996). The MMP-13 gene from chondrocytes selectively enhanced degradation of type II collagen in OA cartilage (Billinghurst et al. 1997). The MMP-1 and MMP-13 genes were further inhibited in low-molecular-weight CSA treatment groups in this experiment compared with CS treatments in previous studies (Legendre et al. 2008; Chan et al. 2005).
We investigated the mechanisms through which heparin sulfate or hyaluronic acid influences ECM synthesis and other biological events. The binding of these polymeric molecules to cell surface receptors, or growth factors, is believed to be crucial for their activity. Thus, oligosaccharides, particularly those shorter than 10 sugar units, do not typically bind growth factors. However, based on the experiments conducted in our laboratory, CSA or CSC oligosaccharides demonstrate a high activity in stimulating hyaline cartilage-related genes, in addition to the synthesis of the related ECM on chondrocytes or mesenchymal stem cells (Wu et al. 2010; Chen et al. 2011a, b). In this study, we grew chondrocytes as pellets, and the control (CSA-free) group exhibited a higher COL II gene expression level compared with the CSA-added groups. However, the CSA-treatment groups showed a higher COL II protein accumulation than the CSA-free group in this study, probably because of a lower ECM-degrading enzyme expression such as of the ADAMTS and MMP gene families.
In summary, chondrocyte pellets that we cultured in a 2 % fetal bovine serum medium exhibited the gene expression of ADAMTS and MMPs over time. Adding low-molecular-weight CSAs inhibited ADAMTS and MMP expression, and increased TIMP3 gene expression. The dp-8 CSA treatment group exhibited more COL II and aggrecan gene expression compared with the other CSA-treatment groups. Based on these findings, the potential use of CSA oligosaccharides in cartilage therapy is promising.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Table 1. Gel permeation chromatography (GPC) of digest GAGs. (PDF 93 kb)
Acknowledgments
This research was partially supported by funding from the Ministry of Economic Affairs, under Grant 91-EC-17-A-17-S1-0009.
Abbreviations
- dp-2
2-mers Oligosaccharides
- dp-4
4-mers Oligosaccharides
- dp-8
8-mers Oligosaccharides
- ADAMTS
A disintegrin and metalloproteinase with a thrombospondin type 1 motif
- AC
Articular cartilage
- COL II
Type II collagen
- COL I
Type I collagen
- CS
Chondroitin sulfate
- CSA
Chondroitin sulfate A
- CSC
Chondroitin-6-sulfate
- DS
Dermatan sulfate
- ECM
Extracellular matrix
- GAGs
Glycosaminoglycan
- MMPS
Metalloproteinase
- OA
Osteoarthritis
- PGs
Proteoglycans
- TIMP3
Tissue inhibitor of matrix metalloproteinase
References
- Aigner T, Sachse A, Gebhard PM, Roach HI. Osteoarthritis: pathobiology-targets and ways for therapeutic intervention. Adv Drug Deliv Rev. 2006;58:128–149. doi: 10.1016/j.addr.2006.01.020. [DOI] [PubMed] [Google Scholar]
- Billinghurst RC, Dahlberg L, Ionescu M, Reiner A, Bourne R, Rorabeck C, Mitchell P, Hambor J, Diekmann O, Tschesche H, Chen J, Van Wart H, Poole AR. Enhanced cleavage of type II collagen by collagenases in osteoarthritic articular cartilage. J Clin Investig. 1997;99:1534–1545. doi: 10.1172/JCI119316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Campo GM, Avenoso A, Campo S, D’Ascola A, Traina P, Sama D, Calatroni A. Glycosaminoglycans modulate inflammation and apoptosis in LPS-treated chondrocytes. J Cell Biochem. 2009;106:83–92. doi: 10.1002/jcb.21981. [DOI] [PubMed] [Google Scholar]
- Chan PS, Caron JP, Orth MW. Effect of glucosamine and chondroitin sulfate on regulation of gene expression of proteolytic enzymes and their inhibitors in interleukin-1-challenged bovine articular cartilage explants. Am J Vet Res. 2005;66:1870–1876. doi: 10.2460/ajvr.2005.66.1870. [DOI] [PubMed] [Google Scholar]
- Chen WC, Wei YH, Huang JP, Chu IM, Yao CL. Biological effects of oligosaccharide chondroitin sulfate C on human articular chondrocytes. Biomed Eng Appl Basis Commun. 2011;23:8. [Google Scholar]
- Chen WC, Yao CL, Chu IM, Wei YH. Compare the effects of chondrogenesis by culture of human mesenchymal stem cells with various type of the chondroitin sulfate C. J Biosci Bioeng. 2011;111:226–231. doi: 10.1016/j.jbiosc.2010.10.002. [DOI] [PubMed] [Google Scholar]
- Derfoul A, Miyoshi AD, Freeman DE, Tuan RS (2007) Glucosamine promotes chondrogenic phenotype in both chondrocytes and mesenchymal stem cells and inhibits MMP-13 expression and matrix degradation.Osteoarthr Cartil 15(6):646–655 [DOI] [PubMed]
- Fischer J. Drug-induced glycosaminoglycan storage: dose-dependent changes in the pattern of accumulated glycosaminoglycans in cultured bovine and human fibroblasts. Biochem Pharmacol. 1996;52:1331–1337. doi: 10.1016/S0006-2952(96)00387-5. [DOI] [PubMed] [Google Scholar]
- Flannery CR (2010) Novel therapies in OA. Curr Drug Targets 11(5):614-619 [DOI] [PubMed]
- Fosang AJ, Last K, Maciewicz RA. Aggrecan is degraded by matrix metalloproteinases in human arthritis. Evidence that matrix metalloproteinase and aggrecanase activities can be independent. J Clin Investig. 1996;98:2292–2299. doi: 10.1172/JCI119040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox BA, Stephens MM. Glucosamine/chondroitin/primorine combination therapy for osteoarthritis. Drugs Today. 2009;45:21–31. doi: 10.1358/dot.2009.45.1.1314053. [DOI] [PubMed] [Google Scholar]
- Holtzer H, Abbott J, Lash J, Holtzer S. The loss of phenotypic traits by differentiated cells in vitro, I. Dedifferentiation of cartilage cells. Proc Natl Acad Sci USA. 1960;46:1533–1542. doi: 10.1073/pnas.46.12.1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iovu M, Dumais G, du Souich P. Anti-inflammatory activity of chondroitin sulfate. Osteoarthr Cartil. 2008;16:S14–S18. doi: 10.1016/j.joca.2008.06.008. [DOI] [PubMed] [Google Scholar]
- Kafienah W, Jakob M, Demarteau O, Frazer A, Barker MD, Martin I, Hollander AP. Three-dimensional tissue engineering of hyaline cartilage: comparison of adult nasal and articular chondrocytes. Tissue Eng. 2002;8:817–826. doi: 10.1089/10763270260424178. [DOI] [PubMed] [Google Scholar]
- Kagita E, Ikeda M, Wakitani S, Takagi M (2010) Effect of monosaccharides composing glycosaminoglycans on type 2 collagen accumulation in a threedimensional culture of chondrocytes. J BiosciBioeng 109(1):51–54 [DOI] [PubMed]
- Kupcsik L, Stoddart MJ, Li Z, Benneker LM, Alini M (2010) Improving chondrogenesis: potential and limitations of SOX9 gene transfer and mechanical stimulation for cartilage tissue engineering. Tissue Eng Part A 16(6):1845–1855 [DOI] [PubMed]
- Legendre F, Bauge C, Roche R, Saurel AS, Pujol JP. Chondroitin sulfate modulation of matrix and inflammatory gene expression in IL-1beta-stimulated chondrocytes–study in hypoxic alginate bead cultures. Osteoarthr Cartil. 2008;16:105–114. doi: 10.1016/j.joca.2007.05.020. [DOI] [PubMed] [Google Scholar]
- Meech R, Edelman DB, Jones FS, Makarenkova HP. The homeobox transcription factor Barx2 regulates chondrogenesis during limb development. Development. 2005;132:2135–2146. doi: 10.1242/dev.01811. [DOI] [PubMed] [Google Scholar]
- Michel BA, Stucki G, Frey D, De Vathaire F, Vignon E, Bruehlmann P, Uebelhart D. Chondroitins 4 and 6 sulfate in osteoarthritis of the knee: a randomized, controlled trial. Arthr Rheum. 2005;52:779–786. doi: 10.1002/art.20867. [DOI] [PubMed] [Google Scholar]
- Nishimoto S, Takagi M, Wakitani S, Nihira T, Yoshida T. Effect of chondroitin sulfate and hyaluronic acid on gene expression in a three-dimensional culture of chondrocytes. J Biosci Bioeng. 2005;100:123–126. doi: 10.1263/jbb.100.123. [DOI] [PubMed] [Google Scholar]
- Radhakrishnamurthy B, Jeansonne N, Berenson GS. Organization of glycosaminoglycan chains in a chondroitin sulfate-dermatan sulfate proteoglycan from bovine aorta. Biochim Biophys Acta. 1986;882:85–96. doi: 10.1016/0304-4165(86)90059-0. [DOI] [PubMed] [Google Scholar]
- Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008;3:1101–1108. doi: 10.1038/nprot.2008.73. [DOI] [PubMed] [Google Scholar]
- Sokoloff L. Osteoarthritis as a remodeling process. J Rheumatol. 1987;14:7–10. [PubMed] [Google Scholar]
- Tiraloche G, Girard C, Chouinard L, Sampalis J, Moquin L, Ionescu M, Reiner A, Poole AR, Laverty S. Effect of oral glucosamine on cartilage degradation in a rabbit model of osteoarthritis. Arthr Rheum. 2005;52:1118–1128. doi: 10.1002/art.20951. [DOI] [PubMed] [Google Scholar]
- Troeberg L, Fushimi K, Khokha R, Emonard H, Ghosh P, Nagase H (2008) Calcium pentosan polysulfate is a multifaceted exosite inhibitor of aggrecanases. FASEB J 22(10):3515–3524 [DOI] [PMC free article] [PubMed]
- Uebelhart D. Clinical review of chondroitin sulfate in osteoarthritis. Osteoarthr Cartil. 2008;16:S19–S21. doi: 10.1016/j.joca.2008.06.006. [DOI] [PubMed] [Google Scholar]
- Vinardell T, Thorpe SD, Buckley CT, Kelly DJ. Chondrogenesis and integration of mesenchymal stem cells within an in vitro cartilage defect repair model. Ann Biomed Eng. 2009;37:2556–2565. doi: 10.1007/s10439-009-9791-1. [DOI] [PubMed] [Google Scholar]
- Wu CH, Ko CS, Huang JW, Huang HJ, Chu IM. Effects of exogenous glycosaminoglycans on human chondrocytes cultivated on type II collagen scaffolds. J Mater Sci Mater Med. 2010;21:725–729. doi: 10.1007/s10856-009-3889-8. [DOI] [PubMed] [Google Scholar]
- Young AA, Smith MM, Smith SM, Cake MA, Ghosh P, Read RA, Melrose J, Sonnabend DH, Roughley PJ, Little CB. Regional assessment of articular cartilage gene expression and small proteoglycan metabolism in an animal model of osteoarthritis. Arthr Res Therapy. 2005;7:R852–R861. doi: 10.1186/ar1756. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table 1. Gel permeation chromatography (GPC) of digest GAGs. (PDF 93 kb)