

Abstract
Retinoid x receptors (RXRs) are master regulators that control cell growth, differentiation, and survival and form heterodimers with many other family members. Here we show that treatment with the RXR agonist IRX4204 enhances the differentiation of CD4+ T cells into inducible regulatory T cells (iTreg) and suppresses the development of T helper (Th) 17 cells in vitro. Furthermore in a murine model of multiple sclerosis (experimental autoimmune encephalomyelitis (EAE)), treatment with IRX4204 profoundly attenuates both active and Th17-mediated passive disease. In the periphery, treatment with IRX4204 is associated with decreased numbers of CD4+ T cells that produce pro-inflammatory cytokines. In addition, CD4+ T cells express decreased levels of Ki-67 and increased expression of CTLA-4. Our findings demonstrate IRX4204 treatment during EAE results in immune modulation and profound attenuation of disease severity.
Keywords: Experimental autoimmune encephalomyelitis (EAE), retinoid x receptors (RXR), rexinoid, regulatory T cell (Treg), T helper 17 (Th17)
Introduction
Multiple sclerosis is a complex autoimmune disease involving central nervous system (CNS) inflammation, demyelination, and axonal loss and affects about one million people worldwide [1]. The most commonly used model to study multiple sclerosis is EAE. In C57BL/6 mice, disease results in a CNS inflammatory disease characterized by progressive ascending paralysis. In this model, myelin-specific CD4+ T cells are generated and differentiate into Th1 and Th17 cells in the periphery [2,3]. Subsequent work indicates that Th17 cells expressing CCR6 initially enter the CNS [4], and that they may become Th1 cells [5]. Both Th1 and Th17 cells can cause activation of microglia that further contribute to neuroinflammation [2].
RXRs are a family of three nuclear receptor isotypes that are key regulators of cell growth, differentiation, and survival during inflammation, cancer, and metabolic disorders [6,7]. Under basal conditions, CD4+ T cells and other hematopoietic cell types express RXRα, and CNS tissue expresses both RXRβ and RXRγ [8-13]. RXRs can form homodimers and homotetramers with each other as well as heterodimers with several other nuclear receptors including (but not limited to) retinoic acid receptors (RARs), vitamin D receptor (VDR), liver x receptors (LXRs), perixosome proliferator-associated receptors (PPARs), nuclear receptor related 1 protein (Nurr1, Nr4a2), and nerve growth factor IB (NGFIB, Nur77, Nr4a1). Of these potential heterodimer partners, LXRs, PPARs, Nurr1, and Nur77 are permissive, meaning that ligand binding of RXR typically causes full activation of the entire heterodimer. In contrast, the RXR-VDR pair is non-permissive and can only be activated by ligand binding to VDR. In addition, RXR-RAR heterodimers are a special case in that they are conditionally permissive. Initial activation of the complex must occur by RAR ligation; however, the subsequent binding of a RXR ligand can cause additive and/or synergistic effects on the activity of the entire heterodimer [6,14,15]. In addition, RXR heterodimers are thought to often function as co-repressors in the absence of ligand. Consequently ligand binding can result in the loss of a co-repressor complex to allow gene transcription [7,14].
Ligands of RXRs include natural compounds such as 9-cis retinoic acid (RA) and docosahexaenoic acid, organotins such as tributyltin, and synthetic agonists termed rexinoids. Each can have distinct effects due to their unique structures and subsequent affinities to RXR [14,16]. For example, 9-cis RA and docosahexaenoic acid can also bind RARs and PPARs, respectively [15,17-19]. In contrast, tributyltin binds RXR at nanomolar concentrations resulting in activation of RXR-PPAR heterodimers; however, this activity may contribute to its high toxicity [20]. In order to study the effect of rexinoids on CD4+ T cell differentiation in vitro as well as in vivo in the context of EAE, we used the rexinoid IRX4204 (originally designated as AGN194204), which has high affinity binding for RXRs and does not transactivate RXR-RAR and RXR-PPARγ heterodimers [21,22]. These findings are relevant for the potential therapeutic use of this and other second generation rexinoids in multiple sclerosis patients.
Materials and methods
Mice
C57BL/6 mice were purchased from Charles River Laboratories and were used at 7-9 weeks of age in a specific pathogen free facility at the Geisel School of Medicine at Dartmouth. Experiments were performed under protocols approved by the Institutional Animal Care and Use Committee of Dartmouth College.
EAE immunization and clinical evaluation
For active EAE, mice were immunized subcutaneously with 175 µg MOG35-55 (Anaspec) emulsified in Incompete Freund’s Adjuvant (Sigma-Aldrich) supplemented with 0.4 mg Mycobacterium tuberculosis H37 (Difco Laboratories) on day 0 and given 200 ng pertussis toxin (List Biologicals) intraperitoneally (IP) on day 0 and 2. For passive EAE, donor mice were immunized as for active EAE except without pertussis toxin, and were sacrificed on day 7 after immunization to obtain spleens. These were homogenized, depleted of red blood cells, and cultured with 20 µg/ml MOG35-55 and 40 ng/ml IL-23 (eBioscience) for 4 days. At the end of culture 7-9 million live CD4+ T cells/mouse were injected intravenously as well as 200 ng pertussis toxin IP. Except where indicated otherwise, mice were treated daily with 200 µg IRX4204 (Io Therapeutics) or vehicle control starting at the indicated time point.
Mice were scored as follows: 1 - limp tail, 1.5 - limp tail and paresis in one rear limb, 2 - limp tail and paralysis on one rear limb, 2.5 - limp tail, paresis one rear limb, and paralysis in other rear limb, 3 - limp tail and paralysis in both rear limbs, and 3.5 - limb tail, rear limb paralysis, and frontal limb paresis. Mice were euthanized at indicated time points, score 3.5, or when they lost 15% of starting body weight (whichever comes first).
Cell Isolation and culture
Spleen and lymph nodes were homogenized with glass slides. CNS tissue was digested with 0.1 mg/ml DNase I (Roche) and 0.16 mg/mL Liberase DL (Roche) in plain RPMI at 37°C for 45 minutes, homogenized, mixed to make 30% Percoll (GE Healthcare), and spun down to form a gradient. Top (myelin) layer of gradient was removed, and cells were washed. Cells were counted on a Nexcelom cellometer and further processed for culture or staining.
For CD4+ T cell culture, lymphocytes were processed using EasySep Mouse Naïve CD4+ T cell Isolation Kit (Stemcell Technologies). Resulting naïve CD4+ T cells (purity 95% or greater) were cultured with irradiated antigen presenting cells and 5 µg/ml soluble anti-CD3 (BioXCell) in RPMI (Corning) supplemented with 10% fetal bovine serum (Hyclone), 50 mM 2-mercaptoethanol, and penicillin/streptomycin/L-glutamine. Th17 cultures included 1 ng/ml TGF-β, 10 ng/mL IL-6, 10 ng/mL IL-1β (Peprotech), 10 µg/mL anti-IFNγ, and 10 µg/mL anti-IL-4 (BioXCell). Inducible Treg cultures included 0, 0.5, 1.0, and 2.0 ng/ml TGF-β and 100 units/mL IL-2 (Peprotech). Where indicated, cultures were supplemented with IRX4204 at 1, 10, or 100 nM or an equivalent volume of vehicle control matched to the 100 nM IRX4204 volume.
Flow cytometry
Cells were stained with either Zombie NIR (Biolegend) or LIVE/DEAD Fixable Yellow stain (Invitrogen) for 20 minutes on ice, washed, stained with surface antibodies, washed, re-suspended, and filtered. For apoptosis staining, cells were stained according to manufacturer’s directions using Annexin V Apoptosis Detection Kit with 7-AAD (Biolegend). For nuclear staining, cells were stained using Transcription Factor Staining Buffer Set (eBioscience). For intracellular staining, samples were re-stimulated with 100 ng/mL phorbol 12-myristate 13-acetate and 500 ng/ml ionomycin (Calbiochem) in the presence of Protein Transport Inhibitor Cocktail (eBioscience) for 5 hours. Cells were then stained using Intracellular Fixation and Permeabilization Buffer Set (eBioscience) for intracellular cytokines or the Transcription Factor Buffer Set (BD Bioscience) for intracellular cytokines and transcription factor staining. Antibodies used were purchased from Biolegend, eBioscience, and BD Bioscience. Samples were run on a modified FACScan (BD Bioscience) running CellQuest (BD Bioscience) and Rainbow (Cytek) software or on a MACSQuant (Miltenyi Biotec) and analyzed with FlowJo software (Tree Star).
Statistical analysis
Graphs were made and analyzed using GraphPad Prism software. Results of iTreg cultures were analyzed by two-way ANOVA and Sidak’s multiple comparison test. Results of Th17 cultures were compared using one-way ANOVA and Dunnett’s multiple comparison test. EAE curves were compared doing a Mann-Whitney U test with the vehicle control treated group. All other analysis of two groups was done using a Student’s t test.
Results
RXR activation by IRX4204 promotes iTreg formation and inhibits Th17 generation
Both 9-cis RA and all-trans retinoic acid (ATRA) enhance iTreg formation and inhibit Th17 differentiation [23-28]. However, unlike ATRA, which only binds to RARs, 9-cis RA can bind both RARs and RXRs. Therefore to determine if RXR activation can mediate these effects, we cultured naïve CD4+ T cells with TGFβ and IL-2, which promotes the generation of iTreg, in the presence of vehicle or 100 nM of IRX4204. Similar to reported effects of ATRA and 9-cis RA, IRX4204 can enhance the differentiation of CD4+ T cells to Foxp3+ T cells when TGFβ is present at 1 and 2 ng/mL (Figure 1A). To evaluate if RXR activation can suppress the differentiation of Th17 cells, naïve T cells were cultured with TGF-β, IL-6, IL-1β, anti-IFNγ, and anti-IL-4 in the presence of vehicle or 1, 10, or 100 nM IRX4204. After re-stimulation CD4+ T cells from Th17 cultures with IRX4204 had decreased production of IL-17A and TNFα (Figure 1B). We also found that RXR activation under Th17 conditions increases CD4+ T cell expression of FoxP3. In addition, we skewed naïve CD4+ T cell differentiation to the Th1 lineage with IL-12 and anti-IL-4 and the Th2 lineage with IL-4 and anti-IFNγ in the presence of vehicle and IRX4204. Supernatants showed similar levels of IFNg in Th1 cultures and IL-4, IL-5, and IL-13 in Th2 cultures, suggesting that IRX4204 does not impact the initial differentiation of these lineages (data not shown).
Figure 1.
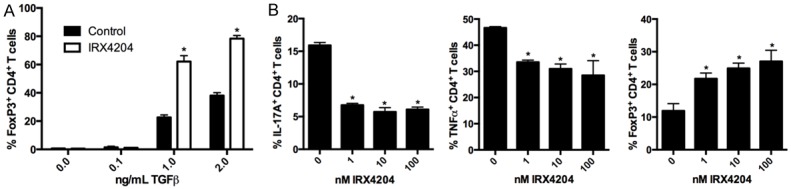
IRX4204 promotes iTreg and inhibits Th17 formation in vitro. Naïve CD4+ T cells were cultured under iTreg (A) or Th17 (B) differentiation conditions for four days in the presence of vehicle control (0 nM group) or IRX4204 at 100 nM or indicated concentration. At the end of culture cells were assessed for FoxP3 expression and cytokine production after re-stimulation and graphed as Mean ± SD. Asterik (*) indicates the group is statistically significant from the controls. Results are representative of 3 independent experiments.
Treatment with RXR agonist IRX4204 ameliorates active EAE
As IRX4204 can impair Th17 and promote iTreg differentiation in vitro, we examined if the administration of IRX4204 could impact on the development of EAE. For this, mice immunized with MOG35-55 peptide emulsified in CFA and pertussis toxin were monitored daily for disease development. On day 7 after immunization they were divided into groups of approximately equal score and treated daily by IP injection with 50, 100, or 200 µg of IRX4204 or a vehicle control. At all dosing levels a statistically significant decrease in disease severity was observed with IRX4204 treatment; however, the greatest differences seen were with the highest dose. This treatment regimen profoundly ameliorated disease, preventing its progression for the duration of the experiment (Figure 2A). On the basis of these findings, further effects of IRX4204 were studied using the highest dosage regimen.
Figure 2.

Treatment with IRX4204 inhibits active EAE. Mice were immunized to induce EAE and treated daily with 50, 100, or 200 ug of IRX4204 or vehicle control IP starting day 7 after immunization. Scores were monitored over time and P-values in comparison to the vehicle treated group are shown. Results are representative of 2 independent experiments with n=6-8 mice/group (A). At the end of experiment vehicle and 200 ug IRX4204 treated groups were used for further analysis. Cells from lymph nodes draining the immunization site and the CNS were pooled to assess total numbers of CD4+ and expression of FoxP3, CCR6, and CD49d in the periphery (B). Cells from lymph node (C) and the CNS (D) were also re-stimulated to assess cytokine production by FACS. Significant P-values are shown. Results using 200 ug dosing are representative of 3 independent experiments with n=6-8 mice/group.
At the end of the experiment mice treated with vehicle or IRX4204 were used for further analysis of peripheral and CNS-infiltrating CD4+ T cells. Consistent with the observed reduced disease severity, total numbers of CD4+ T cells in the periphery were reduced (Figure 2B). However, in contrast to our in vitro findings of enhanced Treg frequency in the presence of IRX4204, treatment in vivo resulted in a reduced frequency of Tregs (Figure 2B). Reflective of the reduction of total numbers of CD4+ T cells, we also observed significant reductions in the total numbers of CD4+ T cells producing IL-17A, IFNγ, and TNFα after re-stimulation (Figure 2C). In the CNS the mean number of CD4+ T cells tended to decrease with IRX4204 treatment though it did not reach statistical significance (Figure 2D). As such, there was also minimal impact on the frequency of CD4+ T cells expressing the CNS homing markers CCR6 and CD49d in the periphery (Figure 2B). As EAE severity is shown to be dependent on the ability of Th17 cells to become Th1 cells [29], we also re-stimulated CNS-infiltrating CD4+ T cells to assess their ability to produce IL-17A and IFNγ. We saw no differences with IRX4204 treatment by frequency (Figure 2D) or numbers (data not shown) of CD4+ T cells producing IL-17A, IFNγ, or both of these two cytokines, suggesting IRX4204 does not impact on the plasticity of Th17 cells to become Th1 in the CNS.
Treatment with IRX4204 attenuates Th17-mediated passive EAE and modulates peripheral CD4+ T cell phenotype and function
We also tested the effects of systemic treatment with IRX4204 on passive EAE mediated by encephalitogenic CD4+ T cells skewed toward a Th17 phenotype. In this model, treatment began on day 1 and continued daily for the duration of the experiment. Similar to our findings with active EAE, mice treated with RXR agonist are protected from disease development (Figure 3A).
Figure 3.

Treatment with IRX4204 inhibits Th17-mediated passive EAE. Mice were given encephalotigenic CD4+ T cells and pertussis toxin on day 0 and were treated with 200ug of IRX4204 or vehicle control IP daily starting day 1 after immunization (A). On day 10 after immunization, cells from spleen were analyzed to determine CD4+ T cell numbers (B), Treg frequency (B), early and late apoptosis (C), and homing marker expression (D). Significant P-values are indicated. Results are representative of 3 independent experiments (n=6 mice/group).
Mice were assessed on day 10 after immunization (about peak of disease in vehicle-treated mice) for effects on CD4+ T cells. In the spleen, CD4+ T cell total numbers are similar between vehicle and IRX4204 treated groups (Figure 3B). Although we observed no differences in cell viability with the usage of IRX4204 in vitro, another group reported that IRX4204 decreases CD4+ T cell apoptosis [30]. Therefore we investigated if IRX4204 can influence CD4+ T cell apoptosis in vivo in the context of EAE. We found that early (Annexin V+ 7AAD-) and late (Annexin V+ 7AAD+) apoptosis in CD4+ T cells was not significantly different with treatment (Figure 3C). IRX4204 treatment is also associated with slightly elevated expression of two CNS homing markers expressed on CD4+ T cells, CCR6 and CD49d (Figure 3D), suggesting the observed phenotype of decreased disease severity is not due to inhibited migration to the CNS.
Studies with 9-cis RA and docasohexaenoic acid suggest that Tregs may be targets of RXR activation [27,31,32]. In addition, deletion of Nurr1 and Nur77, two potential heterodimer partners for RXR, implicates them as regulators of Treg development and function [33-35]. Furthermore we observed that IRX4204 treatment is associated with a decreased Treg frequency in both active and Th17-mediated passive EAE (Figure 3B). In an attempt to reconcile this with the observed reduced disease severity seen with treatment, an analysis of several markers associated with Treg phenotype and function was done. Expression of CD25, which is associated with Treg stability, increased slightly but significantly with IRX4204 treatment. Neuropillin-1 (Nrp-1) and Helios, markers associated with natural Tregs as well as Treg activation, were significantly decreased. IRX4204 treatment also decreased expression of Ki-67, a marker of mitotic activity, and the co-stimulatory marker GITR. Three additional markers that contribute to Treg function, CTLA-4, LAP, and CD39, were unchanged with treatment. Overall these results suggest that Tregs are less activated with IRX4204 treatment.
As IRX4204 treatment during EAE attenuates disease severity and does not result in enhanced Treg phenotype, we also analyzed the FoxP3- CD4+ T cell population (Figure 4B). We found that CTLA-4 was significantly increased in the IRX4204-treated group. This expression can allow for both cell intrinsic and extrinsic inhibition of CD4+ T cell responses [36,37]. Consistent with this finding, the activation markers CD25, Nrp-1, Helios, and GITR were reduced on FoxP3- CD4+ T cells with IRX4204 treatment. Treatment also decreased mitotic activity in FoxP3- CD4+ T cells as assessed by Ki-67. This suggests that systemic IRX4204 treatment decreases activation of FoxP3- CD4+ T cells.
Figure 4.

CD4+ T cells from mice passively immunized for EAE are less activated with IRX4204 treatment. On day 10 after passive EAE immunization, spleens were analyzed by FACS for expression of the indicated markers. Analysis of both FoxP3+ (A) and FoxP3- (B) CD4+ T cells is shown. Significant P-values are shown. Results are representative of 3 independent experiments (n=6 mice/group).
To further assess CD4+ T cell function, cells from spleens and CNS were re-stimulated in vitro and assessed for the expression of cytokines and transcription factors. In the periphery we found that IRX4204 inhibited Th17 cell numbers in vivo as assessed by IL-17A production; however, RORγt expression was not significantly reduced (Figure 5). In addition, the number of CD4+ T cells producing IFNγ and TNFα was reduced. Consistent with the observed reduction in IFNγ, CD4+ T cells from mice receiving IRX4204 treatment also expressed significantly less T-bet. We also observed that the mean number of CD4+ T cells producing IL-4 tends to increase with treatment although this fails to reach statistical significance. In the CNS we found that the number of infiltrating CD4+ T cells was not significantly reduced with IRX4204 treatment (Figure 6). In addition, no differences in the frequency of IL-17A and IFNγ cytokine production were observed. However, RORγt expression by CD4 T cells was reduced in mice treated with IRX4204, suggesting that the Th17 lineage may be less stable in the CNS.
Figure 5.

IRX4204 treated mice passively immunized for EAE have reduced numbers of CD4+ T cells pro-inflammatory cytokines in the periphery. On day 10 after passive EAE immunization, spleens were taken and re-stimulated for expression of cytokines and transcription factors. Significant P-values are shown. Results are representative of 3 independent experiments (n=6 mice/group).
Figure 6.
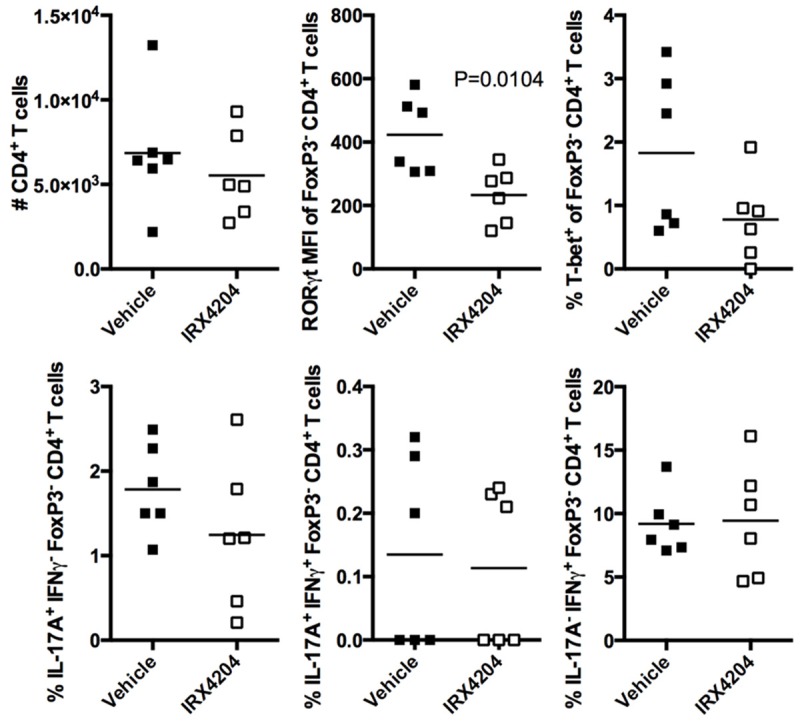
CD4+ T cells from the CNS of passively immunized EAE mice receiving IRX4204 treatment have responses similar to controls. On day 10 after immunization, CNS tissue was processed for lymphocytes and re-stimulated for expression of cytokines and transcription factors. Significant P-values are shown. Results are representative of 3 independent experiments (n=6 mice/group).
Discussion
The findings presented here show that IRX4204 treatment profoundly inhibits both active and passive EAE, suggesting it may be beneficial for the treatment of multiple sclerosis. Previous studies using the RXR agonists 9-cis RA, docosahexaenoic acid, and tributyltin reported relatively modest amelioration of active EAE [38-40]. Considering the differences in the specificity of these agonists, our findings suggest that activation of RXR-RAR and RXR-PPARγ are not necessary for attenuation of disease. However, activation of other RXR heterodimers such as RXR-Nurr1 may be. Indeed, treatment with isoxazolo-pyridinone 7e, an activator of the Nurr1 signaling pathway, attenuates active but not passive EAE [41]. This result would be consistent with some but not all of the effects of IRX4204 being through RXR-Nurr1 activation.
Studies utilizing a T cell lineage restricted dominant negative RAR implicate RA binding to RAR as playing a prominent role in directing Th1 fate commitment in models of allograft tolerance, oral tolerance, and infection and diminishing Th17 responses in response to alloantigen [42,43]. Therefore we hypothesized that IRX4204 binding of RXR may influence CD4+ T lineage development and activation in a similar manner as ligand binding to RAR. During EAE we observed that IRX4204 treatment results in reductions in IFNγ, IL-17A, TNFα, and T-bet expression in peripheral CD4+ T cells. This suggests RXR may be important in stabilizing Th1 and Th17 lineage commitment. Moreover, we also observe that FoxP3+ and FoxP3- CD4+ T cells from IRX4204 treated mice have decreased activation manifesting as reduced Ki-67, GITR, Nrp-1, and Helios expression. Because CTLA-4 is known to have cell extrinsic effects [36,37], the increased expression of CTLA-4 on FoxP3- CD4+ T cells we observe may also be contributing to this phenotype. This is also consistent with the finding that treatment with docosahexaenoic acid in an atopic dermatitis model causes increases in CTLA-4 mRNA in CD4+ T cells and attenuates disease severity [31]. Overall, our data suggests that ligand binding of RXR limits CD4+ T cell activation and destabilizes Th1 and Th17 lineage commitment during active and passive EAE.
As differences in CD4+ T cells were not as striking in the CNS with IRX4204 treatment, it is certainly possible that the main target of its activity in this tissue may be on additional cell types. Work with IRX4204 in vitro has shown that it reduces IL-6 and nitric oxide production by a macrophage cell line in response to lipopolysaccharide [44]. It is possible that it may be able to have similar effects on CNS macrophages and/or microglia in vivo. In addition, 9-cis RA promotes regeneration of myelin sheath neurons in a toxin-induced model of demyelination [12]. If this ability is mediated through RXR, and not RAR, it is possible IRX4204 may also have the same ability during EAE. This will be the subject of future studies examining the mechanism of action of IRX4204 treatment during neuroinflammation.
Acknowledgements
This work was supported by grants from the National Institute of Health (R01AI11405901A1 and R43AI112512). In addition, portions of this work were performed with the help of Dartmouth’s Immune Monitoring and Flow Cytometry Resource and Irradiation, Preclinical Imaging, and Microscopy Resource.
Disclosure of conflict of interest
E.C.N., R.J.N., and R.A.S.C have patent applications for the use of IRX4204 in the treatment of autoimmune diseases. R.J.N. is an advisor and shareholder of Io Therapeutics. R.A.S.C. is a director, scientific officer, and shareholder of Io Therapeutics, which is developing IRX4204 for clinical use.
References
- 1.Nylander A, Hafler DA. Multiple sclerosis. J Clin Invest. 2012;122:1180–1188. doi: 10.1172/JCI58649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Goverman J. Autoimmune T cell responses in the central nervous system. Nat Rev Immunol. 2009;9:393–407. doi: 10.1038/nri2550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Robinson AP, Harp CT, Noronha A, Miller SD. The experimental autoimmune encephalomyelitis (EAE) model of MS: utility for understanding disease pathophysiology and treatment. Handb Clin Neurol. 2014;122:173–189. doi: 10.1016/B978-0-444-52001-2.00008-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Reboldi A, Coisne C, Baumjohann D, Benvenuto F, Bottinelli D, Lira S, Uccelli A, Lanzavecchia A, Engelhardt B, Sallusto F. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nat Immunol. 2009;10:514–523. doi: 10.1038/ni.1716. [DOI] [PubMed] [Google Scholar]
- 5.Hirota K, Duarte JH, Veldhoen M, Hornsby E, Li Y, Cua DJ, Ahlfors H, Wilhelm C, Tolaini M, Menzel U, Garefalaki A, Potocnik AJ, Stockinger B. Fate mapping of IL-17-producing T cells in inflammatory responses. Nat Immunol. 2011;12:255–263. doi: 10.1038/ni.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Altucci L, Leibowitz MD, Ogilvie KM, de Lera AR, Gronemeyer H. RAR and RXR modulation in cancer and metabolic disease. Nat Rev Drug Discov. 2007;6:793–810. doi: 10.1038/nrd2397. [DOI] [PubMed] [Google Scholar]
- 7.Kiss M, Czimmerer Z, Nagy L. The role of lipid-activated nuclear receptors in shaping macrophage and dendritic cell function: From physiology to pathology. J Allergy Clin Immunol. 2013;132:264–286. doi: 10.1016/j.jaci.2013.05.044. [DOI] [PubMed] [Google Scholar]
- 8.Du X, Tabeta K, Mann N, Crozat K, Mudd S, Beutler B. An essential role for Rxr alpha in the development of Th2 responses. Eur J Immunol. 2005;35:3414–3423. doi: 10.1002/eji.200535366. [DOI] [PubMed] [Google Scholar]
- 9.Stephensen CB, Borowsky AD, Lloyd KC. Disruption of Rxra gene in thymocytes and T lymphocytes modestly alters lymphocyte frequencies, proliferation, survival and T helper type 1/type 2 balance. Immunology. 2007;121:484–498. doi: 10.1111/j.1365-2567.2007.02595.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Nunez V, Alameda D, Rico D, Mota R, Gonzalo P, Cedenilla M, Fischer T, Bosca L, Glass CK, Arroyo AG, Ricote M. Retinoid X receptor alpha controls innate inflammatory responses through the up-regulation of chemokine expression. Proc Natl Acad Sci U S A. 2010;107:10626–10631. doi: 10.1073/pnas.0913545107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ricote M, Snyder CS, Leung HY, Chen J, Chien KR, Glass CK. Normal hematopoiesis after conditional targeting of RXRalpha in murine hematopoietic stem/progenitor cells. J Leukoc Biol. 2006;80:850–861. doi: 10.1189/jlb.0206097. [DOI] [PubMed] [Google Scholar]
- 12.Huang JK, Jarjour AA, Nait Oumesmar B, Kerninon C, Williams A, Krezel W, Kagechika H, Bauer J, Zhao C, Baron-Van Evercooren A, Chambon P, Ffrench-Constant C, Franklin RJ. Retinoid X receptor gamma signaling accelerates CNS remyelination. Nat Neurosci. 2010;14:45–53. doi: 10.1038/nn.2702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bookout AL, Jeong Y, Downes M, Yu RT, Evans RM, Mangelsdorf DJ. Anatomical profiling of nuclear receptor expression reveals a hierarchical transcriptional network. Cell. 2006;126:789–799. doi: 10.1016/j.cell.2006.06.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dawson MI, Xia Z. The retinoid X receptors and their ligands. Biochim Biophys Acta. 2012;1821:21–56. doi: 10.1016/j.bbalip.2011.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lefebvre P, Benomar Y, Staels B. Retinoid X receptors: common heterodimerization partners with distinct functions. Trends Endocrinol Metab. 2010;21:676–683. doi: 10.1016/j.tem.2010.06.009. [DOI] [PubMed] [Google Scholar]
- 16.Perez E, Bourguet W, Gronemeyer H, de Lera AR. Modulation of RXR function through ligand design. Biochim Biophys Acta. 2012;1821:57–69. doi: 10.1016/j.bbalip.2011.04.003. [DOI] [PubMed] [Google Scholar]
- 17.Heyman RA, Mangelsdorf DJ, Dyck JA, Stein RB, Eichele G, Evans RM, Thaller C. 9-cis retinoic acid is a high affinity ligand for the retinoid X receptor. Cell. 1992;68:397–406. doi: 10.1016/0092-8674(92)90479-v. [DOI] [PubMed] [Google Scholar]
- 18.Levin AA, Sturzenbecker LJ, Kazmer S, Bosakowski T, Huselton C, Allenby G, Speck J, Kratzeisen C, Rosenberger M, Lovey A, et al. 9-cis retinoic acid stereoisomer binds and activates the nuclear receptor RXR alpha. Nature. 1992;355:359–61. doi: 10.1038/355359a0. [DOI] [PubMed] [Google Scholar]
- 19.de Urquiza AM, Liu S, Sjoberg M, Zetterstrom RH, Griffiths W, Sjovall J, Perlmann T. Docosahexaenoic acid, a ligand for the retinoid X receptor in mouse brain. Science. 2000;290:2140–2144. doi: 10.1126/science.290.5499.2140. [DOI] [PubMed] [Google Scholar]
- 20.le Maire A, Grimaldi M, Roecklin D, Dagnino S, Vivat-Hannah V, Balaguer P, Bourguet W. Activation of RXR-PPAR heterodimers by organotin environmental endocrine disruptors. EMBO Rep. 2009;10:367–373. doi: 10.1038/embor.2009.8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vuligonda V, Thacher SM, Chandraratna RA. Enantioselective syntheses of potent retinoid X receptor ligands: differential biological activities of individual antipodes. J Med Chem. 2001;44:2298–2303. doi: 10.1021/jm0100584. [DOI] [PubMed] [Google Scholar]
- 22.Li X, Hansen PA, Xi L, Chandraratna RA, Burant CF. Distinct mechanisms of glucose lowering by specific agonists for peroxisomal proliferator activated receptor gamma and retinoic acid X receptors. J Biol Chem. 2005;280:38317–38327. doi: 10.1074/jbc.M505853200. [DOI] [PubMed] [Google Scholar]
- 23.Schambach F, Schupp M, Lazar MA, Reiner SL. Activation of retinoic acid receptor-alpha favours regulatory T cell induction at the expense of IL-17-secreting T helper cell differentiation. Eur J Immunol. 2007;37:2396–2399. doi: 10.1002/eji.200737621. [DOI] [PubMed] [Google Scholar]
- 24.Sun CM, Hall JA, Blank RB, Bouladoux N, Oukka M, Mora JR, Belkaid Y. Small intestine lamina propria dendritic cells promote de novo generation of Foxp3 T reg cells via retinoic acid. J Exp Med. 2007;204:1775–1785. doi: 10.1084/jem.20070602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Coombes JL, Siddiqui KR, Arancibia-Carcamo CV, Hall J, Sun CM, Belkaid Y, Powrie F. A functionally specialized population of mucosal CD103+ DCs induces Foxp3+ regulatory T cells via a TGF-{beta} and retinoic acid dependent mechanism. J Exp Med. 2007;204:1757–1764. doi: 10.1084/jem.20070590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Benson MJ, Pino-Lagos K, Rosemblatt M, Noelle RJ. All-trans retinoic acid mediates enhanced T reg cell growth, differentiation, and gut homing in the face of high levels of co-stimulation. J Exp Med. 2007;204:1765–1774. doi: 10.1084/jem.20070719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Mucida D, Park Y, Kim G, Turovskaya O, Scott I, Kronenberg M, Cheroutre H. Reciprocal TH17 and regulatory T cell differentiation mediated by retinoic acid. Science. 2007;317:256–260. doi: 10.1126/science.1145697. [DOI] [PubMed] [Google Scholar]
- 28.Elias KM, Laurence A, Davidson TS, Stephens G, Kanno Y, Shevach EM, O’Shea JJ. Retinoic acid inhibits Th17 polarization and enhances FoxP3 expression through a Stat-3/Stat-5 independent signaling pathway. Blood. 2008;111:1013–1020. doi: 10.1182/blood-2007-06-096438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wang Y, Godec J, Ben-Aissa K, Cui K, Zhao K, Pucsek AB, Lee YK, Weaver CT, Yagi R, Lazarevic V. The transcription factors T-bet and Runx are required for the ontogeny of pathogenic interferon-gamma-producing T helper 17 cells. Immunity. 2014;40:355–366. doi: 10.1016/j.immuni.2014.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rasooly R, Schuster GU, Gregg JP, Xiao JH, Chandraratna RA, Stephensen CB. Retinoid x receptor agonists increase bcl2a1 expression and decrease apoptosis of naive T lymphocytes. J Immunol. 2005;175:7916–7929. doi: 10.4049/jimmunol.175.12.7916. [DOI] [PubMed] [Google Scholar]
- 31.Han SC, Koo DH, Kang NJ, Yoon WJ, Kang GJ, Kang HK, Yoo ES. Docosahexaenoic Acid Alleviates Atopic Dermatitis by Generating Tregs and IL-10/TGF-beta-Modified Macrophages via a TGF-beta-Dependent Mechanism. J Invest Dermatol. 2015;135:1556–1564. doi: 10.1038/jid.2014.488. [DOI] [PubMed] [Google Scholar]
- 32.Yessoufou A, Ple A, Moutairou K, Hichami A, Khan NA. Docosahexaenoic acid reduces suppressive and migratory functions of CD4+CD25+ regulatory T-cells. J Lipid Res. 2009;50:2377–2388. doi: 10.1194/jlr.M900101-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sekiya T, Kashiwagi I, Inoue N, Morita R, Hori S, Waldmann H, Rudensky AY, Ichinose H, Metzger D, Chambon P, Yoshimura A. The nuclear orphan receptor Nr4a2 induces Foxp3 and regulates differentiation of CD4+ T cells. Nat Commun. 2011;2:269. doi: 10.1038/ncomms1272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Sekiya T, Kashiwagi I, Yoshida R, Fukaya T, Morita R, Kimura A, Ichinose H, Metzger D, Chambon P, Yoshimura A. Nr4a receptors are essential for thymic regulatory T cell development and immune homeostasis. Nat Immunol. 2013;14:230–237. doi: 10.1038/ni.2520. [DOI] [PubMed] [Google Scholar]
- 35.Fassett MS, Jiang W, D’Alise AM, Mathis D, Benoist C. Nuclear receptor Nr4a1 modulates both regulatory T-cell (Treg) differentiation and clonal deletion. Proc Natl Acad Sci U S A. 2012;109:3891–3896. doi: 10.1073/pnas.1200090109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wang CJ, Kenefeck R, Wardzinski L, Attridge K, Manzotti C, Schmidt EM, Qureshi OS, Sansom DM, Walker LS. Cutting edge: cell-extrinsic immune regulation by CTLA-4 expressed on conventional T cells. J Immunol. 2012;189:1118–1122. doi: 10.4049/jimmunol.1200972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Corse E, Allison JP. Cutting edge: CTLA-4 on effector T cells inhibits in trans. J Immunol. 2012;189:1123–1127. doi: 10.4049/jimmunol.1200695. [DOI] [PubMed] [Google Scholar]
- 38.Diab A, Hussain RZ, Lovett-Racke AE, Chavis JA, Drew PD, Racke MK. Ligands for the peroxisome proliferator-activated receptor-gamma and the retinoid X receptor exert additive anti-inflammatory effects on experimental autoimmune encephalomyelitis. J Neuroimmunol. 2004;148:116–126. doi: 10.1016/j.jneuroim.2003.11.010. [DOI] [PubMed] [Google Scholar]
- 39.Kong W, Yen JH, Ganea D. Docosahexaenoic acid prevents dendritic cell maturation, inhibits antigen-specific Th1/Th17 differentiation and suppresses experimental autoimmune encephalomyelitis. Brain Behav Immun. 2011;25:872–882. doi: 10.1016/j.bbi.2010.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Takeuchi H, Yokota-Nakatsuma A, Ohoka Y, Kagechika H, Kato C, Song SY, Iwata M. Retinoid X receptor agonists modulate Foxp3(+) regulatory T cell and Th17 cell differentiation with differential dependence on retinoic acid receptor activation. J Immunol. 2013;191:3725–3733. doi: 10.4049/jimmunol.1300032. [DOI] [PubMed] [Google Scholar]
- 41.Montarolo F, Raffaele C, Perga S, Martire S, Finardi A, Furlan R, Hintermann S, Bertolotto A. Effects of isoxazolo-pyridinone 7e, a potent activator of the Nurr1 signaling pathway, on experimental autoimmune encephalomyelitis in mice. PLoS One. 2014;9:e108791. doi: 10.1371/journal.pone.0108791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Brown CC, Esterhazy D, Sarde A, London M, Pullabhatla V, Osma-Garcia I, Al-Bader R, Ortiz C, Elgueta R, Arno M, de Rinaldis E, Mucida D, Lord GM, Noelle RJ. Retinoic acid is essential for Th1 cell lineage stability and prevents transition to a Th17 cell program. Immunity. 2015;42:499–511. doi: 10.1016/j.immuni.2015.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Pino-Lagos K, Guo Y, Brown C, Alexander MP, Elgueta R, Bennett KA, De Vries V, Nowak E, Blomhoff R, Sockanathan S, Chandraratna RA, Dmitrovsky E, Noelle RJ. A retinoic aciddependent checkpoint in the development of CD4+ T cell-mediated immunity. J Exp Med. 2011;208:1767–1775. doi: 10.1084/jem.20102358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Liby K, Royce DB, Risingsong R, Williams CR, Wood MD, Chandraratna RA, Sporn MB. A new rexinoid, NRX194204, prevents carcinogenesis in both the lung and mammary gland. Clin Cancer Res. 2007;13:6237–6243. doi: 10.1158/1078-0432.CCR-07-1342. [DOI] [PubMed] [Google Scholar]