

Abstract
Unlike conventional T cells, innate CD8 T cells develop a memory-like phenotype in the thymus and immediately respond upon antigen stimulation, similar to memory T cells. The development of innate CD8 T cells in the thymus is known to require IL-4, which upregulates Eomesodermin (Eomes). These features are similar to that of virtual memory CD8 T cells and IL-4-induced memory-like CD8 T cells generated in the peripheral tissues. However, the relationship between these cell types has not been clearly documented. In the present study, IL-4-induced memory-like CD8 T cells generated in the peripheral tissues were compared with innate CD8 T cells in terms of phenotype and function. When an IL-4/anti-IL-4 antibody complex (IL-4C) was injected into C57BL/6 mice daily for 7 days, the EomeshiCXCR3 + CD8 T cell population was markedly increased in the peripheral lymphoid organs and blood. These cells were generated from naïve CD8 T cells or accumulated via the expansion of pre-existing CD44hiCXCR3 + CD8 T cells. Initially, the majority of these CXCR3 + CD8 T cells expressed low levels of CD44, which was followed by the conversion to the CD44hi phenotype. This conversion was associated with the acquisition of enhanced effector function. After discontinuation of IL-4C treatment, Eomes expression levels gradually decreased in CXCR3 + CD8 T cells. Taken together, the results of this study demonstrate that IL-4-induced memory-like CD8 T cells generated in the peripheral lymphoid tissues are phenotypically and functionally similar to the innate CD8 T cells generated in the thymus.
Keywords: IL-4, Memory-like CD8 T cells, Innate CD8 T cells, CXCR3
INTRODUCTION
Unlike conventional T cells, some T cells develop a memory-like phenotype in the thymus and immediately respond upon antigen stimulation. These T cells, termed innate CD8 T cells, include NKT cells, T-T CD4 (or T-CD4) T cells, H2-M3 specific T cells, mucosal-associated invariant T (MAIT) cells, CD8αα + intraepithelial T cells, and innate CD8 T cells expressing Eomesodermin (Eomes) (1,2,3).
Eomes + innate CD8 T cells were initially discovered in mice deficient in T cell signaling molecules or transcription factors such as Itk, Klf2, Cbp, or Id3 (3,4,5). A large fraction of CD8 single positive (SP) thymocytes in these mice expresses memory markers such as CD44 and CD122, and IFN-γ. These cells expressed significant amounts of Eomes, which is dependent on the IL-4 produced by innate T cells expressing PLZF (promyelocytic leukemia zinc finger protein) (4,6). For this reason, these innate T cells were called IL-4-induced innate CD8 T cells (7). Eomes + innate CD8 T cells were also identified in CIITA transgenic (CIITATg) mice (3) and wild-type (WT) BALB/c mice (6). In plck-CIITATg C57BL/6 (B6) mice, where the proximal lck promoter-driven expression of CIITA (MHC class II transactivator) induced the expression of major MHC class II in thymocytes and T cells, MHC class II dependent thymocyte-thymocyte (T-T) interactions allowed the generation of an innate CD4 T cell, called T-T CD4 T cells (8). High quantities of Eomes + CD8 T cells were detected in the thymus of these CIITATg mice. The development of these cells was dependent on PLZF + T-T CD4 T cells (9), while PLZF + NKT cells drove the generation of these innate CD8 T cells in WT BALB/c mice (6). An Eomes + CD8 T cell population with an innate phenotype was also found in human fetal thymus and spleen (9).
In addition to innate CD8 T cell generation in an IL-4 rich intrathymic environment, similar cells have also been found in peripheral tissues of WT mice (10,11). Using MHC/peptide tetramers, a subpopulation of antigen-specific CD8 T cells bearing memory markers such as CD44, CD122, and Ly6C were found in unimmunized mice (10). Their presence in germ-free mice supported the hypothesis that these cells acquired a memory-like phenotype even in the absence of antigen stimulation. These antigen-inexperienced memory phenotype CD8 T cells have been called virtual memory (VM) CD8 T cells (10,11,12). Generation of VM CD8 T cells is dependent on endogenous IL-4 (11).
The memory-like CD8 T cell population is also expanded in mice administered with an IL-4/anti-IL-4 antibody complex (IL-4C) (13). IL-4C induces an innate CD8 T cell-like phenotype in peripheral CD8 T cells, which is characterized by elevated expression levels of CD44, CD122, CXCR3, and Eomes. However, the relationship between these three types of memory-like CD8 T cells (Eomes + innate CD8 T cells, VM CD8 T cells, and IL-4-induced memory-like CD8 T cells) has not been clearly documented. In the present study, IL-4-induced memory-like CD8 T cells were compared with innate CD8 T cell in terms of their phenotype and function.
MATERIALS AND METHODS
Mice
B6, BALB/c, IL-4 -/- B6, OT-I B6, and CD45.1 + B6 mice were purchased from Jackson Laboratories (Bar Harbor, ME, USA). B6 mice were thymectomized at 6 weeks of age and maintained until 8 weeks of age. plck-CIITATg mice were generated in the Seoul National University College of Medicine (8). All mice were bred and maintained under specific pathogen-free conditions in the Biomedical Center for Animal Resource Development at the Seoul National University. All experiments were approved by the Institutional Animal Care and Use Committee of the Institute of Laboratory Animal Resource at the Seoul National University, Korea.
Administration of IL-4 and anti-IL-4 antibody in vivo
Based on a previously reported protocol (14), a mixture of 1.5 µg mouse IL-4 (Peprotech, Princeton, NJ, USA) and 50 µg anti-IL-4 antibody (11B11; Bio X Cell, West Lebanon, NH, USA) was intraperitoneally injected into mice daily. After 7 days of treatment, lymphoid organs were analyzed. For the adoptive transfer, donor CD8 T cells were isolated from CD45.1 + B6 mouse splenocytes using anti-CD8 microbeads and magnetic sorting (MACS; Miltenyi Biotec, Auburn, CA, USA). Next, these CD8 T cells were sorted into CD44lowCXCR3 - and CD44hiCXCR3 + populations using a FACSAria (BD Bioscience, San Jose, CA, USA) and then labeled with Cell Trace Violet (CTV, Life Technologies, Waltham, MA, USA) for tracing. Each recipient B6 mouse received 3×106 sorted cells via lateral tail vein injection, followed by IL-4C treatment one day later.
Flow cytometry analysis
Fluorochrome-or biotin-labeled monoclonal antibodies against the following antigens: CD8 (53-6.7), CD44 (IM7), CD62L (MEL-14), CD124 (mIL4R-M1), CXCR3 (CXCR3-173), and CD24 (M1/69) were purchased from BD Bioscience (San Jose, CA, USA), eBioscience (San Diego, CA, USA), and BioLegend (San Diego, CA, USA). Single-cell suspensions were labeled with antibodies for 30 min at 4℃. For intracellular labeling, prepared cells were resuspended in a mixture of fixation and permeabilization buffers from the Foxp3 staining buffer kit (eBioscience, San Diego, CA, USA). Then, intracellular labeling was performed using antibody Eomes (BD Bioscience, San Diego, CA, USA). Flow cytometry was performed on a FACSCalibur and LSR Fortessa (Becton Bioscience, Mountain View, CA, USA). The data were analyzed using the FlowJo software (TreeStar, Ashland, OR, USA).
For the intracellular cytokine assay, CD8 T cells isolated from mouse spleens were stimulated with 50 ng/mL phorbol 12-myristate 13-acetate (PMA) and 1.5 µM ionomycin (Sigma-Aldrich, St Louis, MO, USA) for 4 h at 37℃ in a CO2 incubator, followed by an additional incubation in the presence of 10 µg/mL brefeldin A (Sigma-Aldrich, St Louis, MO, USA) for 2 h. Cultured cells were labeled with anti-CD8 and anti-CXCR3, followed by fixation, permeabilization, and intracellular cytokine labeling with anti-IFN-γ.
Graft-versus-host disease model
Recipient BALB/c mice were exposed to 800 rad of total body irradiation from a [137Cs] source. Donor cells were prepared from the spleen of B6 mice injected with IL-4C daily for 7 days. CD8 T cells were selected by magnetic sorting (MACS; Miltenyi Biotec, Auburn, CA, USA), followed by sorting into CD44lowCXCR3 + and CD44hiCXCR3 + cell populations using a FACSAria (BD Bioscience, San Jose, CA, USA) and CTV labeling. Each recipient mouse received 2×105 sorted cells mixed with 3×106 T cell-depleted bone marrow cells from WT B6 mice. The splenocytes were analyzed 8 weeks later.
Statistical analyses
All data were analyzed by a two-way ANOVA using the GraphPad Prism software (GraphPad Software, CA, USA). The bar graphs represent the mean±standard deviation (SD).
RESULTS
Generation of IL-4-induced CXCR3 + CD8 T cells
To compare the phenotype and function of Eomes + innate CD8 T cells and IL-4-induced memory-like T cells arising from peripheral tissues, CIITATg mice, IL-4C-treated mice, and WT mice (control) were used. As previous reported (9), we found that the proportion and absolute number of fully matured CD24low CD8 SP thymocytes increased in CIITATg mice when compared to WT mice (Fig. 1A), most of which exhibited a characteristic CD44hiCXCR3 + phenotype (Fig. 1B). A substantial fraction of CD44hi memory CD8 T cells in the spleen of WT mice also expressed CXCR3, and CD44lowCXCR3 + CD8 T cell populations were also detected in low numbers (Fig. 1B). To obtain the IL-4-induced memory-like T cells, B6 mice were injected with IL-4C daily for 7 days, and CD8 T cells were isolated from the thymus and spleen of mice on the eighth day. Consistent with a previous report (13), IL-4C treatment resulted in an increase in CD8 T cells (Fig. 1A) and the accumulation of a CD44lowCXCR3 + cell population (Fig. 1B) in the spleen. Of note, the CXCR3 + CD8 SP cell population was also expanded in the thymus of IL-4C-treated mice (Fig. 1B). This was associated with an approximate two-fold increase in the number of fully matured CD24low CD8 SP thymocytes (Fig. 1A). However, the IL-4C-induced accumulation of CXCR3 + CD8 T cells was also observed in the spleen of thymectomized mice (Fig. 1C), indicating that peripheral accumulation of these cells was not a result of emigration from the thymus. The effect of the IL-4C treatment was also observed in a monoclonal TCR system. For this experiment, CD45.1 + OT-I mice, which have ovalbumin-specific monoclonal TCR CD8 T cells, were injected with IL-4C daily for 7 days. Similar to the polyclonal system, both CD44hiCXCR3 + and CD44lowCXCR3 + OT-I cells were markedly increased in the spleen of the IL-4C-treated recipients (Fig. 1D).
Figure 1. Generation of a memory-like CD8 T cell population by IL-4 and anti-IL-4 antibody complex injections. (A & B) Comparison of memory-like CD8 T cells in CIITA-transgenic (CIITATg) mice and IL-4C treated B6 mice. WT B6 mice were injected with IL-4C daily for 7 days. On day 8, CD4 and CD8 T cell populations among CD24low mature thymocytes and total splenocytes (A) and the expression levels of CD44 and CXCR3 on CD8 T cells from the thymus and spleen (B) were compared with those of WT and CIITATg mice. Representative flow cytometry data (left panel) and summarized graph (n=3, right panel) from two independent experiments are shown. Numbers in the plots indicate the percentages of cells in each quadrant. The bars represent mean±SD. *p<0.05; **p<0.01; ***p <0.001. (C) Thymus-independent accumulation of memory-like CD8 T cells in IL-4C-treated mice. WT and thymectomized B6 mice were injected with IL-4C daily for 7 days. CD8 T cells were isolated from the spleen to compare CD44 and CXCR3 expression levels with those of untreated mice. Representative flow cytometry data from two independent experiments are shown. Numbers in the plots indicate the percentages of cells in each quadrant. (D) Generation of memory-like CD8 T cells in monoclonal TCR transgenic mice. OT-1 TCR transgenic mice were treated with IL-4C for 7 days. CD8 T cells were isolated from the spleen, and then labeled with anti-CD44 and anti-CXCR3 antibodies. Representative flow cytometry data from three independent experiments are shown. Numbers in the plots indicate the percentages of cells in each quadrant.

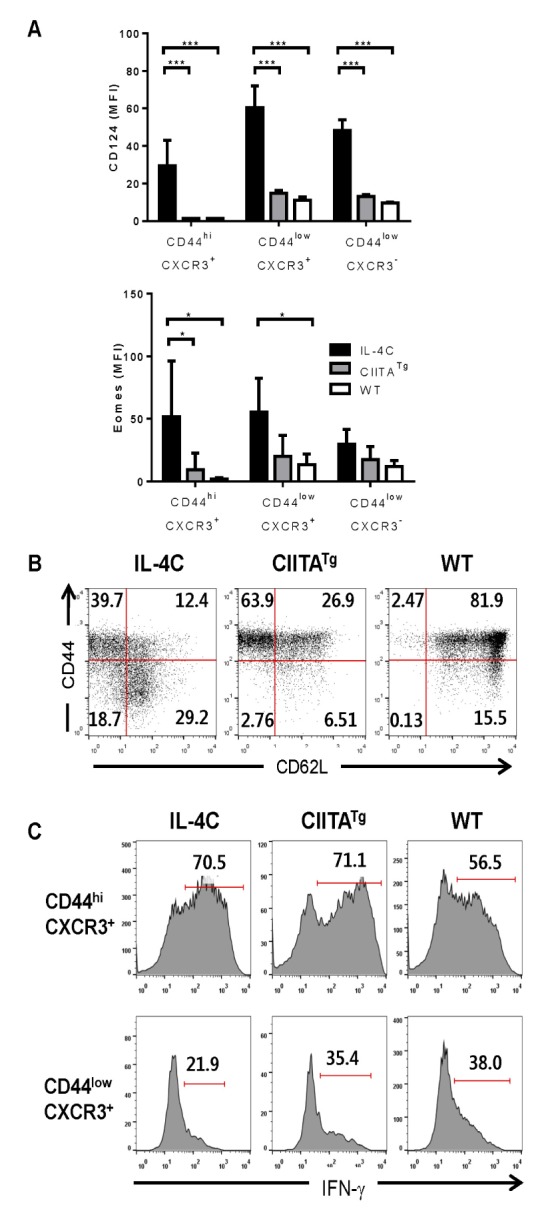
When other memory markers were examined, both CD44hiCXCR3 + and CD44lowCXCR3 + cell populations from IL-4C-treated mice were found to have higher expression levels of both CD124 (IL-4 receptor α subunit) and Eomes, when compared to Eomes + innate CD8 T cells from CIITATg mice and to naïve CD8 T cells from WT mice (Fig. 2A). Moreover, CD44lowCXCR3 + CD8 T cells expressed CD62L, which functions as a lymphoid tissue homing receptor, while CD44hiCXCR3 + cell populations contained both central memory (CD44hiCD62L + ) and effector memory (CD44hiCD62L - ) T cells (Fig. 2B). When CD44hiCXCR3 + CD8 T cells were stimulated with PMA and ionomycin, the majority of cells immediately produced IFN-γ, whereas CD44lowCXCR3 + CD8 T cells isolated from three different mice showed low IFN-γ production capacity (Fig. 2C).
Figure 2. Phenotype and function of memory-like CD8 T cells in IL-4C-treated mice. Splenic CD8 T cells were isolated from IL-4C-treated mice, and the expression levels of the indicated markers (A & B) or IFN-γ production (C) were compared with those of CIITA-transgenic (CIITATg) and untreated WT mice. Summarized graph (n=3, A) and representative data from three independent experiments (B & C) are shown. The bars represent mean±SD. Numbers in the plots indicate the percentages of cells producing IFN-γ. *p< 0.05; ***p<0.001.
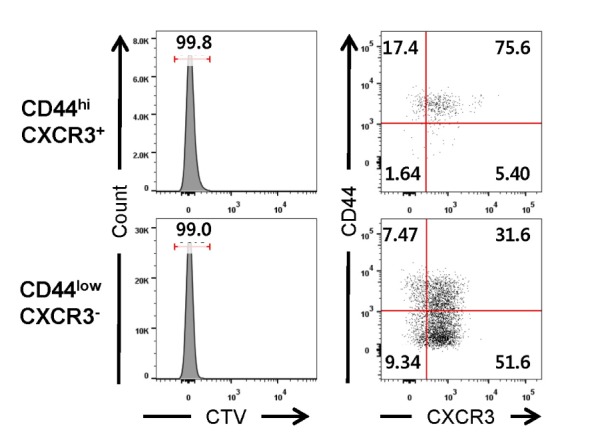
Next, we investigated whether IL-4-induced memorylike CD8 T cells arose from naïve T cells and/or accumuaccumulated via expansion of preexisting CD44hiCXCR3 + cells, most of which are known to be VM T cells (15). For this purpose, CD44lowCXCR3 - (naïve) or CD44hiCXCR3 + CD8 T cells were adoptively transferred into B6 mice, followed by IL-4C treatment. Donor cells were isolated from the spleen of CD45.1 + B6 mice and labeled with CTV prior to the adoptive transfer. In these B6 mice, IL-4C treatment induced the proliferation (CTVlow) of CD44lowCXCR3 - and CD44hiCXCR3 + CD8 T cells, and simultaneously, caused the differentiation of naïve CD8 T cells into both CD44lowCXCR3 + and CD44hiCXCR3 + cells during their proliferation (Fig. 3). These results indicate that the IL-4C treatment is able to induce the generation of memory-like CD8 T cells from naïve CD8 T cells, as well as expand pre-existing CD44hiCXCR3 + cells.
Figure 3. Generation of memory-like CD8 T cells from naïve CD8 T cells CD44hiCXCR3 + (memory phenotype) and CD44lowCXCR3 - (naïve) cells sorted from CD45.1 + B6 splenocytes were labeled with CTV and transferred into CD45.2 + B6 mice via intravenous injection. The recipient mice were injected with IL-4C. The phenotypic changes and proliferation of transferred CD8 T cells in the spleen were analyzed. Numbers in the plots indicate the percentages of cells in each population or quadrant.
Maintenance and maturation of IL-4-induced CXCR3 + CD8 T cells
To determine how long CXCR3 + CD8 T cells could be maintained after cessation of IL-4C treatment, peripheral blood mononuclear cells were analyzed for approximately 1 month post-treatment. In IL-4C-treated mice, the total CXCR3 + cell population reached its peak size on the eighth day after discontinuation of IL-4C treatment, and then gradually decreased over the follow-up period (Fig. 4A). The decrease in the total CXCR3 + cell population was associated with a contraction in the CD44low fraction. In contrast, the CD44hi fraction slowly increased and eventually the majority of CXCR3 + CD8 T cells exhibited a CD44hi phenotype (by the 29th day) (Fig. 4A). These results suggest that CD44lowCXCR3 + CD8 T cells might represent a transient stage between cells with a CD44lowCXCR3 - naïve and CD44hiCXCR3 + memory phenotype. We also measured Eomes expression levels in the CXCR3 + population during the follow-up period. As previously shown, IL-4C treatment significantly upregulated Eomes expression in CXCR3 + CD8 T cells; however, these levels rapidly declined after discontinuation of IL-4C treatment, until basal levels were reached (Fig. 4B).
Figure 4. Conversion of CD44lowCXCR3 + CD8 T cells to the CD44hi phenotype. (A & B) Effect that discontinuing IL-4C treatment has on the CXCR3 + CD8 T cell population. B6 mice were treated with IL-4C for 7 days, and the percentage of total CXCR3 + , CD44hiCXCR3 + , and CD44lowCXCR3 + cell populations in peripheral blood CD8 T cells (A) and Eomes expression levels in CXCR3 + CD8 T cells (B) were analyzed by flow cytometry on the indicated days after treatment. Data summarized from two experiments are shown. (C & D) Conversion of CD44lowCXCR3 + CD8 T cells to the CD44hi phenotype by allo-stimulation. Splenic CD8 T cells from IL-4C-treated B6 mice were sorted into CD44hiCXCR3 + and CD44lowCXCR3 + populations by FACSAria and labeled with CTV. A total of 2×105 CTV-labeled cells from each population were mixed with 3×106 T cell-depleted bone marrow cells from untreated B6 mice, and transferred into irradiated BALB/c mice through intravenous injection. After the transfer, body weight was monitored every week (C). On day 14 after the cell transfer, splenocytes from the recipient BALB/c mice were labeled with antibodies against H-2Kb, CD8, CD44 and CXCR3, and the expression levels of CD44 and CXCR3 in CTVlow H-2Kb+ donor CD8 T cells that had proliferated were analyzed by flow cytometry (D). Numbers in the plots indicate the percentage of cells in each quadrant.

Next, CD44lowCXCR3 + or CD44hiCXCR3 + CD8 T cells isolated from the spleen of IL-4C-treated B6 mice were injected into sub-lethally irradiated BALB/c mice. Donor cells were labeled with CTV prior to the adoptive transfer for tracing. After the adoptive transfer of CD8 T cells, recipients of CD44hiCXCR3 + CD8 T cells showed a more severe reduction in body weight compared to that of the mice receiving CD44lowCXCR3 + cells, indicating that CD44hiCXCR3 + cells have higher alloreactivity than CD44lowCXCR3 + cells (Fig. 4C). The injected cells were recovered 14 days after the transfer, and the population of proliferated CTVlow cells was gated. Some CD44hiCXCR3 + donor cells maintained their phenotype in terms of CD44 and CXCR3 expression levels, even 2 weeks after the stimulation with alloantigen; however, CXCR3 expression was downregulated in the main fraction (Fig. 4D). CD44 expression was upregulated in almost all CD44lowCXCR3 + donor cells, and their phenotype was nearly identical to that of activated CD44hiCXCR3 + donor cells. These data indicate that CD44lowCXCR3 + CD8 T cells are converted into CD44hiCXCR3 + cells via TCR stimulation.
DISCUSSION
IL-4 is a common γ chain cytokine that promotes differentiation of naïve CD4 + T cells into the Th2 cell subset and inhibits the Th1 response (16,17). IL-4 is also able to regulate CD8 T cell development and function. Endogenous IL-4 stimulates CD8 T cell proliferation (14); in the thymus, IL-4 is required for the development of innate CD8 T cells (369). In addition, exogenous IL-4 induces a memory-like phenotype in peripheral CD8 T cells (13). In the present study, an exogenous IL-4 treatment upregulated CD44, CXCR3, CD124, and Eomes in peripheral CD8 T cells. Consistent with a previous report (13), IL-4 was more potent at causing the accumulation of CD44lowCXCR3 + cells than the accumulation of CD44hiCXCR3 + CD8 T cells. This effect could be associated with the expansion of CD44lowCXCR3 + cells and/or the conversion of naïve CD8 T cells to the CD44lowCXCR3 + phenotype. In WT mice, CD44lowCXCR3 + cells are rare relative to CD44lowCXCR3 - naïve and CD44hiCXCR3 + memory cell populations. However, this cell population had higher CD124 (α chain of the IL-4 receptor) expression levels than CD44hiCXCR3 + cells, suggesting that the CD44lowCXCR3 + cell population could be more susceptible to IL-4C treatment. Moreover, these cells could be generated from naïve CD8 T cells. In the spleen, the amount of CD44lowCXCR3 - naïve T cells decreased after IL-4C treatment, suggesting that naïve T cells were converted to the CXCR3 + phenotype. This was confirmed by adoptive transfer of naïve CD8 T cells into congenic mice, followed by IL-4C treatment.
After cessation of IL-4C treatment, the total CXCR3 + cell population gradually decreased over a 1-month period. However, CD44 expression levels gradually increased in these cells, and the majority of CXCR3 + CD8 T cells exhibited the CD44hi phenotype at the end of the follow-up period. CD44 expression levels in T cells increased as a result of diverse situations involving TCR signaling, like positive selection of thymocytes, homeostatic proliferation of T cells in a lymphopenic host, and activation by agonistic antigens. In the present study, a graft-versus-host disease model revealed that activation with an alloantigen promoted the differentiation of IL-4C-induced CD44lowCXCR3 + CD8 T cells into CD44hiCXCR3 - and CD44hiCXCR3 + effector/memory cells. In the absence of exogenous antigen stimulation, TCR signaling by low affinity self-antigens seemed to upregulate CD44 expression levels in CXCR3 + CD8 T cells. This conversion of CD44lowCXCR3 + cells to CD44hiCXCR3 + cells was associated with the acquisition of an enhanced effector function. CD44hiCXCR3 + cells produced significantly higher amounts of IFN-γ and exhibited higher alloreactivity than CD44lowCXCR3 + cells. Moreover, CD44lowCXCR3 + CD8 T cells expressed CD62L like naïve T cells, whereas CD44hiCXCR3 + populations contained both central memory (CD44hiCD62L + ) and effector memory (CD44hiCD62L - ) T cells. In this aspect, this step represents a maturation process.
In the present study, most splenic CD8 T cells from IL-4C-treated mice showed higher levels of CD124 expression when compared to untreated CD8 T cells. This reflects the fact that most CD8 T cells received an IL-4C-induced stimulation signal. However, CXCR3 expression was not upregulated in some of the CD8 T cells. Moreover, when CD8 T cells were treated with IL-4C in vitro, CXCR3 was not upregulated (data not shown). As proposed in a previous report (18), this result suggests that other signals are required for a full innate CD8 T cell-like phenotype.
Eomes expression levels significantly increased in CD8 T cells of IL-4C-treated mice, particularly in the CXCR3 + cell populations. However, after the IL-4C treatment was discontinued, Eomes expression levels in these cell populations gradually decreased. Similarly, although CIITATg mice had abundant Eomes + CD44hiCXCR3 + innate CD8 T cells in their thymus (9), CD44hiCXCR3 + CD8 T cells from the spleen of these mice expressed only slightly higher levels of Eomes than their WT counterparts. Taken together, the results from this study suggest that IL-4 is required for the expression and maintenance of high-expressing Eomes in both centrally IL-4-induced innate CD8 T cells and peripherally IL-4-induced memory-like CD8 T cells.
Innate CD8 T cells are functionally characterized by enhanced INF-γ production and cytotoxic activity. Additionally, they could provide effective immune protection against intracellular organisms such as virus and bacteria (7,19). Based on this, we are now investigating whether IL-4C treatment could enhance antiviral immunity via generation and/or expansion of memory-like CD8 T cells.
This study conclusively demonstrated that IL-4-induced memory-like CD8 T cells that are generated in the peripheral tissues are very similar to innate CD8 T cells generated in the thymus in terms of phenotype, cytokine production, and IL-4 dependence for the maintenance of Eomes. These memory-like CD8 T cells can be generated from naïve CD8 T cells and can also be accumulated via the expansion of pre-existing VM CD8 T cells.
ACKNOWLEDGEMENTS
This work was supported by a grant of the Korea health Technology R&D Project through the Korea Health Industry Development Institute, funded by the Ministry of Health & Welfare, Republic of Korea (grant number: HI13C0954).
Abbreviations
- B6
C57BL/6
- CIITATg
CIITA transgenic
- CTV
Cell Trace Violet
- Eomes
Eomesodermin
- IL-4C
IL-4 and anti-IL-4 antibody complex
- PLZF
promyelocytic leukemia zinc finger protein
- SP
single positive
- T-T
thymocyte-thymocyte
- VM
virtual memory
- WT
wild-type
Footnotes
CONFLICTS OF INTEREST: The authors have no financial conflict of interest.
References
- 1.Berg LJ. Signalling through TEC kinases regulates conventional versus innate CD8+ T-cell development. Nat Rev Immunol. 2007;7:479–485. doi: 10.1038/nri2091. [DOI] [PubMed] [Google Scholar]
- 2.Veillette A, Dong Z, Latour S. Consequence of the SLAM-SAP signaling pathway in innate-like and conventional lymphocytes. Immunity. 2007;27:698–710. doi: 10.1016/j.immuni.2007.11.005. [DOI] [PubMed] [Google Scholar]
- 3.Lee YJ, Jameson SC, Hogquist KA. Alternative memory in the CD8 T cell lineage. Trends Immunol. 2011;32:50–56. doi: 10.1016/j.it.2010.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Atherly LO, Lucas JA, Felices M, Yin CC, Reiner SL, Berg LJ. The Tec family tyrosine kinases Itk and Rlk regulate the development of conventional CD8+ T cells. Immunity. 2006;25:79–91. doi: 10.1016/j.immuni.2006.05.012. [DOI] [PubMed] [Google Scholar]
- 5.Broussard C, Fleischacker C, Horai R, Chetana M, Venegas AM, Sharp LL, Hedrick SM, Fowlkes BJ, Schwartzberg PL. Altered development of CD8+ T cell lineages in mice deficient for the Tec kinases Itk and Rlk. Immunity. 2006;25:93–104. doi: 10.1016/j.immuni.2006.05.011. [DOI] [PubMed] [Google Scholar]
- 6.Weinreich MA, Odumade OA, Jameson SC, Hogquist KA. T cells expressing the transcription factor PLZF regulate the development of memory-like CD8+ T cells. Nat Immunol. 2010;11:709–716. doi: 10.1038/ni.1898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lee A, Park SP, Park CH, Kang BH, Park SH, Ha SJ, Jung KC. IL-4 induced innate CD8+ T cells control persistent viral infection. PLoS Pathog. 2015;11:e1005193. doi: 10.1371/journal.ppat.1005193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Choi EY, Jung KC, Park HJ, Chung DH, Song JS, Yang SD, Simpson E, Park SP. Thymocyte-thymocyte interaction for efficient positive selection and maturation of CD4 T cells. Immunity. 2005;23:387–396. doi: 10.1016/j.immuni.2005.09.005. [DOI] [PubMed] [Google Scholar]
- 9.Min HS, Lee YJ, Jeon YK, Kim EJ, Kang BH, Jung KC, Chang CH, Park SH. MHC class II-restricted interaction between thymocytes plays an essential role in the production of innate CD8+ T cells. J Immunol. 2011;186:5749–5757. doi: 10.4049/jimmunol.1002825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Haluszczak C, Akue AD, Hamilton SE, Johnson LD, Pujanauski L, Teodorovic L, Jameson SC, Kedl RM. The antigen-specific CD8+ T cell repertoire in unimmunized mice includes memory phenotype cells bearing markers of homeostatic expansion. J Exp Med. 2009;206:435–448. doi: 10.1084/jem.20081829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Akue AD, Lee JY, Jameson SC. Derivation and maintenance of virtual memory CD8 T cells. J Immunol. 2012;188:2516–2523. doi: 10.4049/jimmunol.1102213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lee JY, Hamilton SE, Akue AD, Hogquist KA, Jameson SC. Virtual memory CD8 T cells display unique functional properties. Proc Natl Acad Sci USA. 2013;110:13498–13503. doi: 10.1073/pnas.1307572110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ventre E, Brinza L, Schicklin S, Mafille J, Coupet CA, Marcais A, Djebali S, Jubin V, Walzer T, Marvel J. Negative regulation of NKG2D expression by IL-4 in memory CD8 T cells. J Immunol. 2012;189:3480–3489. doi: 10.4049/jimmunol.1102954. [DOI] [PubMed] [Google Scholar]
- 14.Morris SC, Heidorn SM, Herbert DR, Perkins C, Hildeman DA, Khodoun MV, Finkelman FD. Endogenously produced IL-4 nonredundantly stimulates CD8+ T cell proliferation. J Immunol. 2009;182:1429–1438. doi: 10.4049/jimmunol.182.3.1429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kurzweil V, LaRoche A, Oliver PM. Increased peripheral IL-4 leads to an expanded virtual memory CD8+ population. J Immunol. 2014;192:5643–5651. doi: 10.4049/jimmunol.1301755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kopf M, Legros G, Bachmann M, Lamers MC, Bluethmann H, Kohler G. Disruption of the murine Il-4 gene blocks Th2 cytokine tesponses. Nature. 1993;362:245–248. doi: 10.1038/362245a0. [DOI] [PubMed] [Google Scholar]
- 17.Ouyang W, Ranganath SH, Weindel K, Bhattacharya D, Murphy TL, Sha WC, Murphy KM. Inhibition of Th1 development mediated by GATA-3 through an IL-4-independent mechanism. Immunity. 1998;9:745–755. doi: 10.1016/s1074-7613(00)80671-8. [DOI] [PubMed] [Google Scholar]
- 18.Carty SA, Koretzky GA, Jordan MS. Interleukin-4 regulates eomesodermin in CD8 T cell development and differentiation. Plos One. 2014;9:e106659. doi: 10.1371/journal.pone.0106659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Oghumu S, Terrazas CA, Varikuti S, Kimble J, Vadia S, Yu L, Seveau S, Satoskar AR. CXCR3 expression defines a novel subset of innate CD8+ T cells that enhance immunity against bacterial infection and cancer upon stimulation with IL-15. FASEB J. 2015;29:1019–1028. doi: 10.1096/fj.14-264507. [DOI] [PMC free article] [PubMed] [Google Scholar]