

Abstract
Chromosomes are not randomly disposed in the nucleus but instead occupy discrete sub-nuclear domains, referred to as chromosome territories. The molecular mechanisms that underlie the formation of chromosome territories and how they are regulated during the cell cycle remain largely unknown. Here, we have developed two different chromosome-painting approaches to address how chromosome territories are organized in the fission yeast model organism. We show that condensin frequently associates RNA polymerase III-transcribed genes (tRNA and 5S rRNA) that are present on the same chromosomes, and that the disruption of these associations by condensin mutations significantly compromises the chromosome territory arrangement. We also find that condensin-dependent intra-chromosomal gene associations and chromosome territories are co-regulated during the cell cycle. For example, condensin-directed gene associations occur to the least degree during S phase, with the chromosomal overlap becoming largest. In clear contrast, condensin-directed gene associations become tighter in other cell-cycle phases, especially during mitosis, with the overlap between the different chromosomes being smaller. This study suggests that condensin-driven intra-chromosomal gene associations contribute to the organization and regulation of chromosome territories during the cell cycle.
INTRODUCTION
Global territorial arrangement of chromosomes is observed in a variety of eukaryotic organisms ranging from yeast to humans, and individual chromosomes are stably separated during interphase (1–8). This territorial chromosomal organization is tightly linked to gene regulation, as it has been shown that genes loop out from chromosome territories upon transcriptional activation (9,10).
Recent genomic studies have revealed that genomes are highly ordered by a hierarchy of organizing events, ranging from gene associations to topologically associating domain (TAD) formation (11–14). The condensin and cohesin complexes consisting of the structural maintenance of chromosomes proteins are known to function in mitotic chromosome assembly and in holding sister chromatids together, respectively (15–17). These complexes are also deeply involved in many aspects of the genome-organizing processes. For instance, it has been proposed that condensin and cohesin direct gene associations, enhancer–promoter interactions, and TAD formation in different species (18–23). Moreover, condensin has recently been implicated in the organization of chromosome territories in Drosophila melanogaster and Caenorhabditis elegans (24,25). A critical question that remains is how condensin mediates and regulates the territorial organization of chromosomes.
Using fission yeast as a model system, we have previously reported on condensin-mediated genome organizations and demonstrated that RNA polymerase III-transcribed (Pol III) genes, such as tRNA and 5S rRNA genes dispersed across the chromosomal arms, associate with centromeres (19,26). We now show that condensin preferentially associates Pol III genes present on the same chromosomes, and that the inhibition of these intra-chromosomal gene associations by condensin mutations disrupts the territorial arrangement of chromosomes. We also show that condensin-directed gene associations are coordinated with the chromosome territory organization throughout the cell cycle.
MATERIALS AND METHODS
Culture conditions
Fission yeast cells were cultured in yeast-extract adenine (YEA) medium. Strain constructions were performed using conventional genetic crosses. The cdc25-22 mutant was used for the cell-cycle synchronization as previously described (27). Alternatively, exponentially growing cells were arrested in S phase by culturing in YEA medium containing 11 mM hydroxyurea (HU) for 4 h.
Microscopic analysis
Fluorescent in situ hybridization (FISH) experiments were performed as previously described (28). Fission yeast chromosomes (Bio-Rad) were separated on 0.8% low melting point (LMP) agarose gel (Life Technologies) using CHEF Mapper XA Pulsed Field Electrophoresis System (Bio-Rad). Electrophoresis was performed at 14°C for 48 h in 1× Tris-acetate EDTA (TAE) buffer at 1.5 V/cm and 120° angle with a 20-min switch time. The respective chromosomes were purified using Qiaquick gel extraction Kit (Qiagen) and amplified using Illustra GenomiPhi V2 DNA Amplification Kit (GE Healthcare). The chromosome-specific FISH probes were also generated using polymerase chain reaction (PCR)-amplified DNA fragments. Fifty-six and forty-six pairs of primers were used for amplification of 5 kb DNA fragments derived from the chromosomes 1 and 2, respectively (Supplementary Tables S1 and S2). DNA fragments were labeled by incorporation of Cy3-dCTP, Cy5-dCTP (GE Healthcare) or Alexa Fluor 488-dUTP (Life Technologies) with a random primer DNA labeling kit (Takara). FISH images were captured by a Zeiss Axioimager Z1 fluorescence microscope with an oil immersion objective lens (Plan Apochromat, 100×, NA 1.4, Zeiss). The images were acquired at 0.2 μm intervals in the z-axis controlled by Axiovision 4.6.3 software (Zeiss). More than 100 cells were analyzed for every microscopic experiment. In the analysis on locus-specific FISH data, the distance between two loci was measured for each sample, and the data for the two groups were subjected to the non-parametric Mann–Whitney U test. When analyzing chromosome-painting FISH results, the percentage of the overlapped area between two chromosomes was estimated as shown in Supplementary Figure S3, and the data from wild-type (WT) and condensin mutant cells were compared by the non-parametric Mann–Whitney U test. In the Mann–Whitney U test, all the values from two groups were ranked from low to high regardless of groups, and the mean ranks in the two groups were compared.
RESULTS
Intra-chromosomal associations of Pol III gene loci
We previously reported that condensin binds to Pol III genes and mediates associations between Pol III gene loci and centromeres in fission yeast (19,26). If condensin binds to Pol III genes and centromeres and mediates their associations, then Pol III genes also should associate with one another. To explore this possibility, we performed FISH experiments and examined associations between Pol III gene loci. The c417 Pol III gene locus contains three tRNA and two 5S rRNA genes within a 15 kb region. On average, the Pol III gene loci we investigated consist of 1–2 Pol III genes within 10–20 kb regions. We observed that the two Pol III gene loci (c10H11 and c417) on the chromosome 3 were frequently positioned in proximity, reflecting their association (Figure 1A). Using a live-cell imaging, we have previously shown that condensin-mediated associations are highly transient during interphase, but gene loci and centromeres migrate together in a coordinated fashion when associated through condensin (29). Since condensin-mediated gene association is a transient event, the association is scored as a frequency of nearby localization of gene loci. In clear contrast, the two Pol III gene loci (c10H11 and c354) on the different chromosomes were infrequently co-localized, and the localization pattern was significantly different from that of the two Pol III gene loci on the same chromosome (P < 0.001, Mann–Whitney U test), suggesting that the Pol III gene loci on the same chromosome tend to associate (Figure 1A and B).
Figure 1.

Frequent co-localization between Pol III gene loci present on same chromosomes. (A) FISH visualization of the Pol III gene loci located on the same and different chromosomes. The c10H11 Pol III gene locus on the chromosome 3 was co-visualized with either the c417 Pol III gene locus (chr3) or the c354 Pol III gene locus (chr2). Representative FISH images are shown on top. Centers of FISH foci were defined as positions of the loci. The distance between the two loci was measured in more than 100 cells and assigned to a 0.1 μm bin. (B) Simplified representation of the FISH results shown in panel A. The distance between the two loci was binned into one of the three categories (right). This is to simplify a representation of the FISH data, and statistical analysis was performed based on the original datasets (the Mann–Whitney U test). (C) Positions of the gene loci visualized by FISH experiments. The cosmid clones (yellow) were used to prepare FISH probes. The cosmid names in shade indicate the negative control loci without Pol III genes, and this representation is consistent in other figures. Positions of Pol III genes (tRNA and 5S rRNA) are also shown. (D) Pairs of the indicated gene loci were co-visualized by FISH, and FISH data were summarized as described in panel B. Numbers of Pol III genes present at the gene loci are annotated beneath the cosmid clones. (E and F) The Pol III genes (tRNAasn and 5S rRNA) were inserted to the c162 non-Pol III gene locus. The c162 control locus with and without the insertion of the Pol III genes was co-visualized with the c10H11 Pol III gene locus (E). FISH data were summarized as described in panel B (F).
We next investigated whether the intra-chromosomal gene association we observed is a common feature of many other Pol III genes distributed across the genome (Figure 1C). We co-visualized combinations of the Pol III gene loci and the non-Pol III gene loci (Figure 1C). There are no Pol III genes located within 100 kb from the c887, c1020 and c162 negative control loci. We consistently observed that the five pairs of the Pol III gene loci (c417-c736, c354-c1677, c354-c800, c354-c428 and c417-c10H11) on the same chromosomes were frequently positioned in proximity, whereas the two pairs of the Pol III gene loci (c354-c27D7 and c417-c354) on the different chromosomes were mainly separated (P < 0.001, Mann–Whitney U test; Figure 1B and D). Moreover, the localization patterns between the non-Pol III gene loci and the Pol III gene loci (c162-c736 and c1020-c417) on the same chromosomes were not clearly different from that of the c162 non-Pol III gene loci and the c354 Pol III gene loci on the different chromosomes (Figure 1D). These negative control data were similar to the localization patterns of the Pol III gene loci (c354-c27D7 and c417-c354) on the different chromosomes, suggesting that the Pol III gene loci on the different chromosomes rarely or do not associate (Figure 1B and D). More importantly, these results suggest that Pol III gene loci on the same chromosomes, but not on the different chromosomes, frequently associate.
If Pol III genes are responsible for intra-chromosomal associations, a non-Pol III gene locus should associate with other Pol III gene loci when Pol III genes are ectopically inserted. In this regard, we found that the c162 Pol III gene locus consisting of the insertion of the two Pol III genes (tRNAasn and 5S rRNA) frequently co-localized with the c10H11 Pol III gene locus (Figure 1E and F), which was significantly different from the localization pattern of the c162 locus without the insertion (P < 0.001, Mann–Whitney U test). This localization pattern was similar to that of the two Pol III gene loci (c417 and c10H11) on the same chromosome (Figure 1B). These results further demonstrate that Pol III genes mediate associations of gene loci present on same chromosomes.
Associations between Pol III gene loci relative to centromeres
We next examined whether co-localization of Pol III gene loci on same chromosomes occurs at centromeres. We observed that the Pol III gene loci (c417 and c10H11) present on the same chromosome frequently co-localized near centromeres (category I) and away from centromeres (category II), while one of the Pol III gene loci was independently positioned near centromeres (category III) in the remaining cell population (Supplementary Figure S1A and S1B). In contrast, the Pol III gene loci (c354 and c10H11) on the different chromosomes predominantly showed the localization pattern of category III (Supplementary Figure S1B). These results indicate that the association between the Pol III gene loci on the same chromosome can occur at and away from centromeres, whereas Pol III gene loci on the different chromosomes preferentially associate with centromeres. We speculate that there is a potential mechanism to prevent the co-localization of Pol III genes on different chromosomes at centromeres.
Intra-chromosomal gene associations mediated by condensin
We next investigated whether intra-chromosomal gene associations are dependent upon condensin activity. The cut3-477 and cut14-208 condensin mutants are commonly used by fission yeast researchers, and condensin function in these mutants is disrupted at the restrictive temperature (30). We observed that the associations between the Pol III gene loci (c417-c10H11 and c354-c800) on the same chromosomes were significantly disrupted by the cut3-477 and cut14-208 mutations (P < 0.001, Mann–Whitney U test; Figure 2A). In contrast, the localization patterns of the Pol III gene loci and non-Pol III gene loci (c10H11-c162 and c354-c887) on the same chromosomes as well as the non-Pol III gene loci (c1020-c162 and c887-c162) on the same and different chromosomes were not significantly affected by the condensin mutations (Figure 2B and C). These results suggest that the condensin mutations specifically disrupt intra-chromosomal associations between Pol III gene loci.
Figure 2.

Intra-chromosomal associations between Pol III genes mediated by condensin. (A) Disruption of intra-chromosomal gene associations by the cut3–477 and cut14–208 condensin mutations. The cut3–477 and cut14–208 mutants were cultured at the restrictive temperature (36°C) for 2 and 1 h, respectively, when most cells were in interphase. Wild-type cells (WT) were also cultured at 36°C for 2 h. The c417 and c10H11 Pol III gene loci located on the chromosome 3 as well as the c354 and c800 Pol III gene loci on the chromosome 2 were co-visualized by FISH. Representative FISH images are shown on top. FISH data were summarized as described in Figure 1B. (B) The c10H11 Pol III gene locus and the162 non-Pol III gene locus on the chromosome 3 were co-visualized in the indicated strains (left). The c354 Pol III gene locus and the c887 non-Pol III gene locus on the chromosome 2 were also investigated by FISH (right). (C) The two pairs of the non-Pol III gene loci, c1020-c162 (chr3) and c887-c162 (chr2 and chr3), were analyzed by FISH in WT and condensin mutant cells. (D) The two pairs of the Pol III gene loci, c417-c10H11 (chr3) and c354-c800 (chr2), were visualized in WT, cut3–477 and cut3–477 sfc3–1 cells.
We previously found that the sfc3-1 point mutation in the sfc3 gene encoding the Pol III transcription factor TFIIIC subunit promotes condensin localization at Pol III genes (19). We observed that the associations of the Pol III gene loci (c10H11-c417 and c800-c354) on the same chromosomes were significantly disrupted by the cut3–477 condensin mutation but restored in the cut3-477 sfc3-1 double mutant (P < 0.001, Mann–Whitney U test; Figure 2D). We predict that the cut3-477 condensin mutation partially decreases condensin activity, while the sfc3-1 mutation can suppress the condensin defect in intra-chromosomal gene associations by promoting condensin binding to Pol III genes. These results collectively indicate that condensin mediates intra-chromosomal associations of Pol III gene loci.
Painting the whole fission yeast chromosomes
To analyze chromosome territories in the fission yeast nucleus, we established a FISH approach to visualize the entire chromosomes (see ‘Materials and Methods’ section). In brief, we purified the respective chromosomes after pulsed-field gel electrophoresis (PFGE) and prepared FISH probes that recognize the entire chromosomes (Figure 3A). As an experimental control, we first applied these FISH probes to prophase-arrested cells treated by carbendazim, a microtubule-destabilizing agent, resulting in the artificially heavily compacted chromosomes. We observed that the FISH probes almost exclusively visualized the individual chromosomes, suggesting that the prepared FISH probes can be used for chromosome painting (Supplementary Figure S2). We then analyzed interphase cells and observed that the three yeast chromosomes were not dispersed so as to occupy the entire nucleus, indicating a territorial organization of the fission yeast chromosomes during interphase (Figure 3B).
Figure 3.

Visualization of the entire fission yeast chromosomes. (A) The fission yeast chromosomes were separated by PFGE and purified from agarose gel. The recovered DNA was used for preparation of chromosome-specific FISH probes. (B) The fission yeast chromosomes were co-visualized using PFGE-derived FISH probes. (C) Positions of FISH probes prepared by PCR amplification. Red and green bars indicate positions of DNA fragments prepared by PCR. PCR-derived DNA fragments were mixed to generate chromosome-specific FISH probes. (D) The fission yeast chromosomes 1 (red) and 2 (green) were co-visualized using PCR-derived FISH probes.
PFGE-derived FISH probes potentially recognize repetitive sequences including centromeric repeats, tRNA and 5S rRNA genes, and long terminal repeat retrotransposons commonly present on all of the fission yeast chromosomes. To precisely estimate chromosome territories, we developed an alternative FISH approach. We prepared 56 and 46 DNA fragments specific to the chromosomes 1 and 2, respectively, by PCR (Figure 3C). These DNA fragments were designed not to contain any repeat sequences and were distributed throughout the chromosomes at 100 kb intervals. These PCR-derived DNA fragments were mixed to prepare chromosome-specific FISH probes. We observed that the chromosomes 1 and 2 were not spread throughout the nucleus, again indicating the territorial organization of the fission yeast chromosomes (Figure 3D). These FISH approaches work comparably for chromosome painting and are useful for probing chromosome territories in the fission yeast model system.
Coordination between intra-chromosomal gene associations and chromosome territory organization during the cell cycle
We determined to investigate how chromosome territories are regulated during the cell cycle. Because condensin directs intra-chromosomal gene associations (Figure 2), we hypothesized that these associations contribute to the territorial organization of chromosomes during the cell cycle. To test this possibility, we first analyzed the association of the Pol III gene loci (c10H11 and c417) present on the same chromosome during the cell cycle. The cell cycle was synchronized using the temperature-sensitive cdc25-22 mutant (Figure 4A). We observed that the intra-chromosomal association was most promoted during mitosis and became infrequent during S phase (Figure 4B and C). In contrast, the c10H11 Pol III gene locus and the c162 non-Pol III gene locus, as well as the two non-Pol III gene loci (c1020 and c162), all of which are present on the same chromosome, were consistently separated during the cell cycle (Figure 4B and C). Note that the distances between the c417 Pol III gene locus and c162 negative control locus, and between the c10H11 Pol III gene locus and the c1020 non-Pol III gene loci are both 120 kb and that the paired loci (c10H11-c417, c10H11-c162 and c1020-c162) are positioned ∼1 Mb apart (Figure 1C). It has been shown that two gene loci located 120 kb apart are generally separated by 0.4–0.5 μm in the interphase nucleus, and these two loci can behave independently of each other (31). Therefore, the observed change in the intra-chromosomal association between the Pol III gene loci during the cell cycle does not simply reflect indirect effects derived from alterations in global chromosomal compaction.
Figure 4.

Intra-chromosomal gene association and chromosome territory arrangement during the cell cycle. (A) The cdc25–22 mutant was used for the cell-cycle synchronization. The cell cycle was monitored by the two indexes (binucleated anaphase cells and septation). The same procedure was employed in the following panels. (B) Intra-chromosomal gene association at the different stages of the cell cycle. The c10H11 Pol III gene locus (chr3) was co-visualized with either the c417 Pol III gene locus (chr3) or the c162 non-Pol III gene locus (chr3). The c1020 and c162 non-Pol III gene loci on the chromosome 3 were also analyzed by FISH. (C) FISH data described in panel B were summarized in a standard boxplot format. The distance between the two loci was measured in more than 200 cells. Open boxes show center quartiles, and whiskers extend to the data point which is no more than 1.5× the interquartile range from the box. (D) The painting FISH experiments with PCR-derived probes were performed using cells in different stages of the cell cycle. Representative FISH images are shown on top. The percentage overlap between the chromosomes 1 and 2 during the cell cycle was estimated as detailed in Supplementary Figure S3 and summarized as boxplots explained in panel C.
To further evaluate chromosome territories, we focused on an overlap between different chromosomes as explained in Supplementary Figure S3. We observed that the overlap between the chromosomes 1 and 2 became smallest and largest during mitosis and S phase, respectively (Figure 4D). This pattern was similar to the pattern of intra-chromosomal gene associations during the cell cycle (Figure 4C). For instance, when the condensin-dependent intra-chromosomal gene association is tightest during mitosis, the chromosomal overlap becomes smallest. In contrast, the gene association occurs in the least degree during S phase, with the chromosomal overlap being largest. Together, these results suggest that chromosome territories are regulated during the cell cycle, and that the territorial organization is coordinated with condensin-dependent intra-chromosomal gene associations rather than global chromosomal compaction.
Condensin mutations compromise chromosome territory arrangement
If condensin-directed gene associations are involved in the territorial organization of chromosomes, then inhibition of the associations by condensin mutations should disrupt chromosome territories (Figure 2A). To address this hypothesis, we performed chromosome painting using the cut3-477 and cut14-208 condensin mutants, and occasionally observed the clear separation of the chromosomes 1 and 2 in WT cells, but not in the condensin mutants (Figure 5A). More specifically, more than 20% of WT cells contained the two chromosomes almost completely separated (0–5% of the chromosomal overlap), while this clear separation was infrequent in the condensin mutants (Figure 5B). On the other hand, the chromosomes were also significantly overlapped in a subset of WT cells, implying that the degree of territorial organization can vary among fission yeast cells. Therefore, the pattern of the chromosomal overlap in a cell population needs to be considered, and we observed that the overlap patterns were significantly different between the WT and the condensin mutants (P < 0.05, Mann–Whitney U test; Figure 5C). In detail, we observed that more than 35% of the chromosomal overlap was observed in 18% of WT cells, whereas 39% of cut3-477 and 33% of cut14-208 condensin mutant cells had the similar degree of the overlap (Figure 5B). Note that asynchronous culture contains ∼75% of G2 cells, and that the observed patterns mainly reflect the chromosomal overlap during G2 phase.
Figure 5.
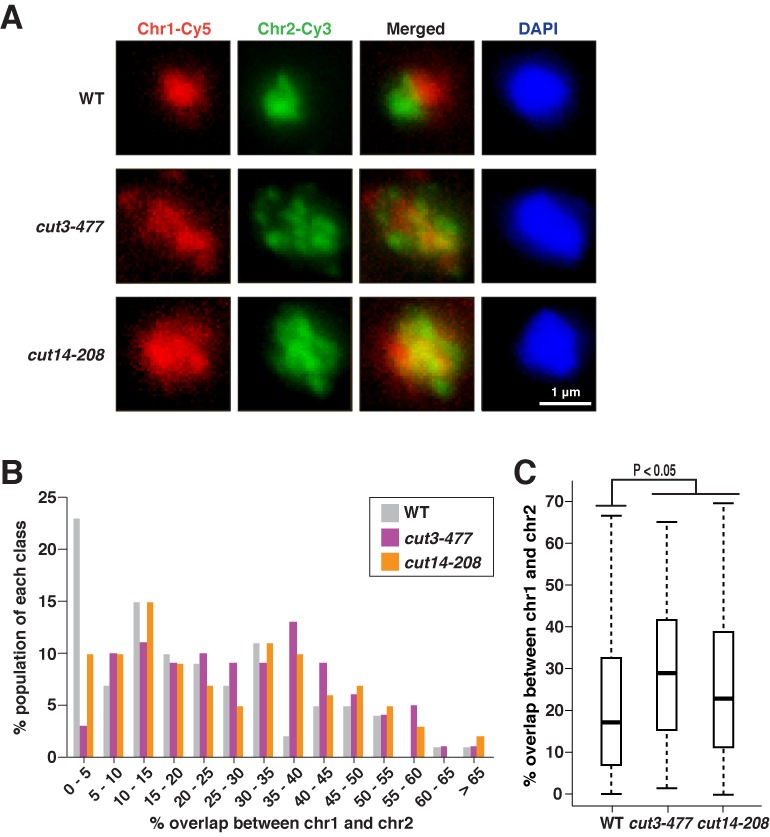
Chromosome territories compromised in condensin mutants. (A) The chromosomes 1 and 2 were co-visualized in WT, cut3–477 and cut14–208 condensin mutant cells. Cells are cultured as described in Figure 2A, and painting FISH was performed with PCR-derived probes. (B) The percentage overlap between the chromosomes 1 and 2 was estimated in the WT and condensin mutants. The observed percentage overlap was assigned to one of 5% bins. (C) The %overlap between the chromosomes 1 and 2 in the indicated strains was represented as boxplots (Figure 4C).
For the different pair of the chromosomes, we again observed a clear separation in the WT cells, but not in the condensin mutants (Supplementary Figure S4A). The patterns of the chromosomal overlap were significantly different between the WT and condensin mutants (P < 0.001, Mann–Whitney U test), and the average overlap became larger in the condensin mutants compared to the WT (Supplementary Figure S4B). We also observed that the chromosomal overlap became smaller in the cut3-477 sfc3-1 double mutant compared to the cut3-477 single mutant (Supplementary Figure S4C and S4D). Our results above suggest that the sfc3–1 mutation can suppress the condensin defect in intra-chromosomal gene associations by promoting condensin binding to Pol III genes (Figure 2D). These results collectively demonstrate that condensin participates in the organization of chromosome territories in fission yeast, and likely that condensin-directed intra-chromosomal associations of Pol III gene loci contribute to the chromosome territory arrangement.
Involvement of condensin in cell cycle-dependent organization of chromosome territories
It was still possible that the condensin mutations might cause chromosomal entanglement and segregation defects during mitosis, resulting in the disruption of chromosome territories in the following cell-cycle phases. To directly investigate roles of condensin in the chromosome territory organization during interphase, we performed HU block/release experiments with the condensin mutants (Figure 6A). The HU treatment arrests the cell cycle at S phase, and cells were subsequently cultured at the restrictive temperature to inhibit condensin activity. This procedure allows condensin mutant cells to progress through interphase, and we can specifically investigate roles of condensin in chromosome territory arrangement during interphase. As expected, the mitotic defects were observed in the cut3-477 and cut14-208 condensin mutants 1.5 h after HU release, indicating that the condensin mutants go through the normal interphase and progress to mitosis after the HU treatment (Figure 6B).
Figure 6.

Effects of condensin mutations on chromosome territory organization during S and G2 phases. (A) Schematic representation of the cell-cycle synchronization using hydroxyurea (HU). Exponentially growing cells were arrested at S phase by culturing in YEA medium containing 11 mM HU at 26°C for 4 h and further cultured at the elevated temperature (36°C), because the cut3–477 and cut14–208 condensin mutations are temperature-sensitive. Cells were released from the S-phase arrest by culturing without HU. (B) Cells prepared by the procedure described in panel A were subjected to DAPI staining. In fission yeast, mitotic defects are observed as stretched chromosomes (30). The percentage of mitotic defects is plotted against time. (C and D) S- (C) and G2-phase (D) cells were subjected to chromosome painting using PFGE-derived FISH probes, and FISH data were summarized as boxplots (Figure 4C). Representative FISH images are shown on top.
During both S and G2 phases, we observed that the patterns of the chromosomal overlap were significantly different between the WT and condensin mutants (P < 0.05, Mann–Whitney U test), and that the average chromosomal overlap was consistently enlarged in the condensin mutants compared to WT cells (Figure 6C and D). These results indicate that condensin contributes to the organization of chromosome territories during S and G2 phases. Although the chromosomal overlap becomes largest during S phase (Figure 4D), condensin seemingly still participates in the relatively loose organization of chromosome territories, suggesting the involvement of condensin in the chromosome territory organization throughout the cell cycle.
DISCUSSION
We show that condensin mediates intra-chromosomal gene associations, and that the condensin mutations disrupt those associations and the organization of chromosome territories. We also find that condensin-mediated gene associations are coordinated with the territorial organization during the cell cycle. These results collectively suggest that condensin-mediated associations contribute to the organization and regulation of chromosome territories during the cell cycle.
Mechanistically speaking, our study indicates that Pol III genes located on the same chromosomes associate frequently. Condensin is known to interact with another condensin molecule to mediate associations between chromatin fibers (32,33). As a result, condensin molecules present at Pol III genes can mediate associations between these genes through condensin–condensin interactions. How condensin mediates only intra-chromosomal associations remains unclear, but it has been suggested that the interaction of condensin, preferentially with another condensin molecule present nearby on the same chromatin fiber, results in mitotic chromosome assembly (34). Recently, a computational simulation study has also predicted that the condensin–condensin interaction and the resulting gene associations can mediate chromatin domain formation (35). This study suggests that condensin-directed intra-chromosomal gene associations are also involved in the territorial organization of chromosomes.
It has previously been shown that condensin plays an inhibitory role in the expression of Pol III genes (19). In this study, we find that condensin mediates intra-chromosomal associations between Pol III genes and facilitates the chromosome territory arrangement. We speculate that embedding Pol III genes in chromosome territories can potentially play an inhibitory role in their transcription. In support of this notion, it has been shown that genes loop out from chromosome territories upon transcriptional activation (9,10). Also, condensin mutations are known to cause mitotic defects, which are observed as stretched chromosomes in fission yeast cells and referred to as the Φ-shaped chromosomes phenotype (30). In this regard, we predict that the condensin-mediated intra-chromosomal gene associations help maintain chromosomal separation during the cell cycle, thereby improving the fidelity of chromosome segregation during mitosis.
How are chromosome territories governed during the cell cycle in fission yeast? It has been shown that phosphorylation of the Cut3 condensin subunit by Cdc2 results in an accumulation of condensin molecules in the mitotic nucleus (36). Additionally, phosphorylation of the Cnd2 condensin subunit promotes condensin-chromatin affinity during mitosis (37). Moreover, an epigenetic histone modification, acetylation of histone H3 lysine 56, has an inhibitory effect on condensin binding to chromatin during S phase (38). These mechanisms likely cause the stable chromatin binding of condensin during mitosis as well as condensin dissociation from chromatin during S phase, allowing condensin to mediate frequent and rare intra-chromosomal gene associations during mitosis and S phases, respectively. This modulation of condensin-directed gene associations presumably enables condensin to affect chromosome territories in a cell cycle-dependent manner. Moreover, it has recently been shown that, when condensin is stabilized or over-expressed during interphase, DAPI-dense chromatin domains, likely reflecting chromosome territories, are observed in the fly and human nuclei, implying that condensin promotes intra-chromosomal gene associations and the territorial organization of interphase chromosomes in higher eukaryotes (39,40). We thus hypothesize that the role of condensin in the organization and regulation of chromosome territories is likely conserved in other eukaryotes.
Supplementary Material
Acknowledgments
We would like to thank the Wistar Imaging Facility for microscopic analysis. We also thank Louise Showe for critically reading the manuscript and Sylvie Shaffer for editorial assistance.
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online.
FUNDING
The G. Harold & Leila Y. Mathers Charitable Foundation; NIH Director's New Innovator Award Program of the National Institutes of Health [DP2-OD004348 to K.N.]; Cancer Center Support Grant (CCSG) to The Wistar Institute [P30CA010815]. Funding for open access charge: The G. Harold & Leila Y. Mathers Charitable Foundation.
Conflict of interest statement. None declared.
REFERENCES
- 1.Scherthan H., Bahler J., Kohli J. Dynamics of chromosome organization and pairing during meiotic prophase in fission yeast. J. Cell Biol. 1994;127:273–285. doi: 10.1083/jcb.127.2.273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Tanabe H., Muller S., Neusser M., von Hase J., Calcagno E., Cremer M., Solovei I., Cremer C., Cremer T. Evolutionary conservation of chromosome territory arrangements in cell nuclei from higher primates. Proc. Natl. Acad. Sci. U.S.A. 2002;99:4424–4429. doi: 10.1073/pnas.072618599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Walter J., Schermelleh L., Cremer M., Tashiro S., Cremer T. Chromosome order in HeLa cells changes during mitosis and early G1, but is stably maintained during subsequent interphase stages. J. Cell Biol. 2003;160:685–697. doi: 10.1083/jcb.200211103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gerlich D., Beaudouin J., Kalbfuss B., Daigle N., Eils R., Ellenberg J. Global chromosome positions are transmitted through mitosis in mammalian cells. Cell. 2003;112:751–764. doi: 10.1016/s0092-8674(03)00189-2. [DOI] [PubMed] [Google Scholar]
- 5.Pecinka A., Schubert V., Meister A., Kreth G., Klatte M., Lysak M.A., Fuchs J., Schubert I. Chromosome territory arrangement and homologous pairing in nuclei of Arabidopsis thaliana are predominantly random except for NOR-bearing chromosomes. Chromosoma. 2004;113:258–269. doi: 10.1007/s00412-004-0316-2. [DOI] [PubMed] [Google Scholar]
- 6.Bolzer A., Kreth G., Solovei I., Koehler D., Saracoglu K., Fauth C., Muller S., Eils R., Cremer C., Speicher M.R., et al. Three-dimensional maps of all chromosomes in human male fibroblast nuclei and prometaphase rosettes. PLoS Biol. 2005;3:e157. doi: 10.1371/journal.pbio.0030157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Duan Z., Andronescu M., Schutz K., McIlwain S., Kim Y.J., Lee C., Shendure J., Fields S., Blau C.A., Noble W.S. A three-dimensional model of the yeast genome. Nature. 2010;465:363–367. doi: 10.1038/nature08973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Tanizawa H., Iwasaki O., Tanaka A., Capizzi J.R., Wickramasinghe P., Lee M., Fu Z., Noma K. Mapping of long-range associations throughout the fission yeast genome reveals global genome organization linked to transcriptional regulation. Nucleic Acids Res. 2010;38:8164–8177. doi: 10.1093/nar/gkq955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Chambeyron S., Bickmore W.A. Chromatin decondensation and nuclear reorganization of the HoxB locus upon induction of transcription. Genes Dev. 2004;18:1119–1130. doi: 10.1101/gad.292104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cremer T., Cremer M., Dietzel S., Muller S., Solovei I., Fakan S. Chromosome territories–a functional nuclear landscape. Curr. Opin. Cell Biol. 2006;18:307–316. doi: 10.1016/j.ceb.2006.04.007. [DOI] [PubMed] [Google Scholar]
- 11.Lieberman-Aiden E., van Berkum N.L., Williams L., Imakaev M., Ragoczy T., Telling A., Amit I., Lajoie B.R., Sabo P.J., Dorschner M.O., et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science. 2009;326:289–293. doi: 10.1126/science.1181369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Schoenfelder S., Sexton T., Chakalova L., Cope N.F., Horton A., Andrews S., Kurukuti S., Mitchell J.A., Umlauf D., Dimitrova D.S., et al. Preferential associations between co-regulated genes reveal a transcriptional interactome in erythroid cells. Nat. Genet. 2010;42:53–61. doi: 10.1038/ng.496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dixon J.R., Selvaraj S., Yue F., Kim A., Li Y., Shen Y., Hu M., Liu J.S., Ren B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature. 2012;485:376–380. doi: 10.1038/nature11082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sexton T., Yaffe E., Kenigsberg E., Bantignies F., Leblanc B., Hoichman M., Parrinello H., Tanay A., Cavalli G. Three-dimensional folding and functional organization principles of the Drosophila genome. Cell. 2012;148:458–472. doi: 10.1016/j.cell.2012.01.010. [DOI] [PubMed] [Google Scholar]
- 15.Hirano T. Chromosome cohesion, condensation, and separation. Annu. Rev. Biochem. 2000;69:115–144. doi: 10.1146/annurev.biochem.69.1.115. [DOI] [PubMed] [Google Scholar]
- 16.Koshland D., Strunnikov A. Mitotic chromosome condensation. Annu. Rev. Cell Dev. Biol. 1996;12:305–333. doi: 10.1146/annurev.cellbio.12.1.305. [DOI] [PubMed] [Google Scholar]
- 17.Nasmyth K., Haering C.H. The structure and function of SMC and kleisin complexes. Annu. Rev. Biochem. 2005;74:595–648. doi: 10.1146/annurev.biochem.74.082803.133219. [DOI] [PubMed] [Google Scholar]
- 18.Haeusler R.A., Pratt-Hyatt M., Good P.D., Gipson T.A., Engelke D.R. Clustering of yeast tRNA genes is mediated by specific association of condensin with tRNA gene transcription complexes. Genes Dev. 2008;22:2204–2214. doi: 10.1101/gad.1675908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Iwasaki O., Tanaka A., Tanizawa H., Grewal S.I., Noma K. Centromeric localization of dispersed Pol III genes in fission yeast. Mol. Biol. Cell. 2010;21:254–265. doi: 10.1091/mbc.E09-09-0790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dowen J.M., Fan Z.P., Hnisz D., Ren G., Abraham B.J., Zhang L.N., Weintraub A.S., Schuijers J., Lee T.I., Zhao K., et al. Control of cell identity genes occurs in insulated neighborhoods in mammalian chromosomes. Cell. 2014;159:374–387. doi: 10.1016/j.cell.2014.09.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mizuguchi T., Fudenberg G., Mehta S., Belton J.M., Taneja N., Folco H.D., FitzGerald P., Dekker J., Mirny L., Barrowman J., et al. Cohesin-dependent globules and heterochromatin shape 3D genome architecture in S. pombe. Nature. 2014;516:432–435. doi: 10.1038/nature13833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Li L., Lyu X., Hou C., Takenaka N., Nguyen H.Q., Ong C.T., Cubenas-Potts C., Hu M., Lei E.P., Bosco G., et al. Widespread rearrangement of 3D chromatin organization underlies polycomb-mediated stress-induced silencing. Mol. Cell. 2015;58:216–231. doi: 10.1016/j.molcel.2015.02.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Crane E., Bian Q., McCord R.P., Lajoie B.R., Wheeler B.S., Ralston E.J., Uzawa S., Dekker J., Meyer B.J. Condensin-driven remodelling of X chromosome topology during dosage compensation. Nature. 2015;523:240–244. doi: 10.1038/nature14450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bauer C.R., Hartl T.A., Bosco G. Condensin II promotes the formation of chromosome territories by inducing axial compaction of polyploid interphase chromosomes. PLoS Genet. 2012;8:e1002873. doi: 10.1371/journal.pgen.1002873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lau A.C., Nabeshima K., Csankovszki G. The C. elegans dosage compensation complex mediates interphase X chromosome compaction. Epigenet. Chromatin. 2014;7:31. doi: 10.1186/1756-8935-7-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Iwasaki O., Tanizawa H., Kim K.D., Yokoyama Y., Corcoran C.J., Tanaka A., Skordalakes E., Showe L.C., Noma K.I. Interaction between TBP and condensin drives the organization and faithful segregation of mitotic chromosomes. Mol. Cell. 2015;59:755–767. doi: 10.1016/j.molcel.2015.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Alfa C., Fantes P., Hyams J., McLeod M., Warbrick E. Experiments with Fission Yeast; A Laboratory Course Manual. NY: Cold Spring Harbor Laboratory Press; 1993. [Google Scholar]
- 28.Sadaie M., Naito T., Ishikawa F. Stable inheritance of telomere chromatin structure and function in the absence of telomeric repeats. Genes Dev. 2003;17:2271–2282. doi: 10.1101/gad.1112103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kim K.D., Tanizawa H., Iwasaki O., Corcoran C.J., Capizzi J.R., Hayden J.E., Noma K. Centromeric motion facilitates the mobility of interphase genomic regions in fission yeast. J. Cell Sci. 2013;126:5271–5283. doi: 10.1242/jcs.133678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Saka Y., Sutani T., Yamashita Y., Saitoh S., Takeuchi M., Nakaseko Y., Yanagida M. Fission yeast cut3 and cut14, members of a ubiquitous protein family, are required for chromosome condensation and segregation in mitosis. EMBO J. 1994;13:4938–4952. doi: 10.1002/j.1460-2075.1994.tb06821.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bystricky K., Heun P., Gehlen L., Langowski J., Gasser S.M. Long-range compaction and flexibility of interphase chromatin in budding yeast analyzed by high-resolution imaging techniques. Proc. Natl. Acad. Sci. U.S.A. 2004;101:16495–16500. doi: 10.1073/pnas.0402766101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Hirano M., Anderson D.E., Erickson H.P., Hirano T. Bimodal activation of SMC ATPase by intra- and inter-molecular interactions. EMBO J. 2001;20:3238–3250. doi: 10.1093/emboj/20.12.3238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yoshimura S.H., Hizume K., Murakami A., Sutani T., Takeyasu K., Yanagida M. Condensin architecture and interaction with DNA: regulatory non-SMC subunits bind to the head of SMC heterodimer. Curr. Biol. 2002;12:508–513. doi: 10.1016/s0960-9822(02)00719-4. [DOI] [PubMed] [Google Scholar]
- 34.Naumova N., Imakaev M., Fudenberg G., Zhan Y., Lajoie B.R., Mirny L.A., Dekker J. Organization of the mitotic chromosome. Science. 2013;342:948–953. doi: 10.1126/science.1236083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Cheng T.M., Heeger S., Chaleil R.A., Matthews N., Stewart A., Wright J., Lim C., Bates P.A., Uhlmann F. A simple biophysical model emulates budding yeast chromosome condensation. Elife. 2015;4:e05565. doi: 10.7554/eLife.05565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sutani T., Yuasa T., Tomonaga T., Dohmae N., Takio K., Yanagida M. Fission yeast condensin complex: essential roles of non-SMC subunits for condensation and Cdc2 phosphorylation of Cut3/SMC4. Genes Dev. 1999;13:2271–2283. doi: 10.1101/gad.13.17.2271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Tada K., Susumu H., Sakuno T., Watanabe Y. Condensin association with histone H2A shapes mitotic chromosomes. Nature. 2011;474:477–483. doi: 10.1038/nature10179. [DOI] [PubMed] [Google Scholar]
- 38.Tanaka A., Tanizawa H., Sriswasdi S., Iwasaki O., Chatterjee A.G., Speicher D.W., Levin H.L., Noguchi E., Noma K. Epigenetic regulation of condensin-mediated genome organization during the cell cycle and upon DNA damage through histone H3 lysine 56 acetylation. Mol. Cell. 2012;48:532–546. doi: 10.1016/j.molcel.2012.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Buster D.W., Daniel S.G., Nguyen H.Q., Windler S.L., Skwarek L.C., Peterson M., Roberts M., Meserve J.H., Hartl T., Klebba J.E., et al. SCFSlimb ubiquitin ligase suppresses condensin II-mediated nuclear reorganization by degrading Cap-H2. J. Cell Biol. 2013;201:49–63. doi: 10.1083/jcb.201207183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Yokoyama Y., Zhu H., Zhang R., Noma K. A novel role for the condensin II complex in cellular senescence. Cell Cycle. 2015;14:2160–2170. doi: 10.1080/15384101.2015.1049778. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.