

Abstract
Background
Vibrio alginolyticus is ubiquitous in marine and estuarine environments. In 2012–2013, SXT/R391-like integrative conjugative elements (ICEs) in environmental V. alginolyticus strains were discovered and found to occur in 8.9 % of 192 V. alginolyticus strains, which suggests that V. alginolyticus may be a natural pool possessing resourceful ICEs. However, complete ICE sequences originating from this bacterium have not been reported, which represents a significant barrier to characterizing the ICEs of this bacterium and exploring their relationships with other ICEs. In the present study, we acquired six ICE sequences from five V. alginolyticus strains and performed a comparative analysis of these ICE genomes.
Results
A sequence analysis showed that there were only 14 variable bases dispersed between ICEValE0601 and ICEValHN492. ICEValE0601 and ICEValHN492 were treated as the same ICE. ICEValA056-1, ICEValE0601 and ICEValHN492 integrate into the 5′ end of the host’s prfC gene, and their Int and Xis share at least 97 % identity with their counterparts from SXT. ICEValE0601 or ICEValHN492 contain 50 of 52 conserved core genes in the SXT/R391 ICEs (not s025 or s026). ICEValA056-2, ICEValHN396 and ICEValHN437 have a different tRNA-ser integration site and a distinct int/xis module; however, the remaining backbone genes are highly similar to their counterparts in SXT/R391 ICEs. DNA sequences inserted into hotspot and variable regions of the ICEs are of various sizes. The variable genes of six ICEs encode a large array of functions to bestow various adaptive abilities upon their hosts, and only ICEValA056-1 contains drug-resistant genes. Many variable genes have orthologous and functionally related genes to those found in SXT/R391 ICEs, such as genes coding for a toxin-antitoxin system, a restriction-modification system, helicases and endonucleases. Six ICEs also contain a large number of unique genes or gene clusters that were not found in other ICEs. Six ICEs harbor more abundant transposase genes compared with other parts of their host genomes. A phylogenetic analysis indicated that transposase genes in these ICEs are highly diverse.
Conclusions
ICEValA056-1, ICEValE0601 and ICEValHN492 are typical members of the SXT/R391 family. ICEValA056-2, ICEValHN396 and ICEValHN437 form a new atypical group belonging to the SXT/R391 family. In addition to the many genes found to be present in other ICEs, six ICEs contain a large number of unique genes or gene clusters that were not found in other ICEs. ICEs may serve as a carrier for transposable genetic elements (TEs) and largely facilitate the dissemination of TEs.
Electronic supplementary material
The online version of this article (doi:10.1186/s12866-016-0692-9) contains supplementary material, which is available to authorized users.
Keywords: Vibrio alginolyticus, Integrative Conjugative Elements (ICEs), Comparative genomics, Transposable genetic Elements (TEs)
Background
Integrative conjugative elements (ICEs) are self-transmissible mobile genetic elements (MGEs) that play a major role in gene flow in bacterial populations [1]. To date, 460 ICEs have been identified (http://db-mml.sjtu.edu.cn/ICEberg) in a variety of Gram-positive and Gram-negative bacteria [2]. Among them, the SXT/R391 family of ICEs is one of the largest and most studied ICE families and currently consists of 89 members [2, 3]. In 1996, SXT/R391 ICEs were first described in Vibrio cholerae, the etiologic agent of the diarrheal disease cholerae, and in Providencia rettgeri, which was isolated in 1972 [4, 5]. Since then, SXT/R391 ICEs were widely found to be prevalent in the 7th pandemic isolates of V. cholerae, other environmental Vibrio species and in some Enterobacteriaceae species [6–10]. A comparative genomic analysis of 13 widely distributed SXT/R391 ICEs indicated that all of these ICEs consist of the same syntenous and nearly identical 52 core genes, whereas other families of closely related mobile elements, such as lambdoid and T4-like phages, exhibit greater diversity [6]. Thus, SXT/R391 ICEs may be a relatively recent creation of evolution [6]. Five hotspots (HS1–HS5) and four variable regions (VR I–IV) are interspaced into the conserved backbone of ICEs, and these variable DNAs generally code for resistance to antibiotics, heavy metals and bacteriophage infection, toxin/antitoxin (TA) systems or c-di-GMP synthesis [6, 7, 11–14], which confer hosts adaptive functions to various environments.
V. alginolyticus is ubiquitous in marine and estuarine environments and is the most common Vibrio species isolated in southern coastal areas of China [15, 16]. V. alginolyticus has received increasing attention because some strains are pathogenic to humans and aquatic animals and have caused huge economic losses [17, 18]. In 2012–2013, SXT/R391-like ICEs in environmental V. alginolyticus strains were first discovered [9, 13, 19] and found to occur in 8.9 % of 192 V. alginolyticus strains [19]. Due to the ubiquity of V. alginolyticus in marine and estuarine environments and because ICEs can transfer at a relatively high frequency between phylogenetically distant species [7, 20–22], it is natural to speculate that V. alginolyticus may represent a reservoir for SXT/R391-like ICEs. These results also raise concerns that ICEs will further disseminate and thereby increase the risk of dissemination of multidrug-resistant gene clusters. However, until now no complete ICE sequences in V. alginolyticus have been reported. This represents a substantial barrier in characterizing the ICEs of this bacterium to further address the problems mentioned above.
In this study, we acquired six ICE sequences from five V. alginolyticus strains including a strain harboring two coresident ICEs to perform a comparative analysis of these ICE genomes. This work will shed light on the characteristics of the ICEs in environmental Vibrio species and provides new knowledge of the considerable diversity of genes and potential accessory functions encoded by the variable DNA in these ICEs.
Results
Assembly and annotation of the ICEs in V. alginolyticus
To obtain the complete sequences of six ICEs, high-throughput HiSeq 2000 sequencing of the genomes of V. alginolyticus strains, A056, E0601, HN396, HN437 and HN492 (Table 1), was carried out. Fifty-six, 93, 117, 133 and 103 scaffolds sequences were, respectively, obtained, assembled and annotated using the RAST annotation pipeline [23]. Genome annotation revealed one ICE sequence in each genome of strains E0601, HN396, HN437 and HN492; strain A056 contains two coexistent ICEs. The alignment and assembly of these ICE-related scaffolds were manually performed using the sequences of SXT (the representative ICE of the SXT/R391 family), ICEVchInd4 and ICEVchMex1 as references. Gap filling was carried out through PCR followed by sequencing, which yielded the complete sequences of ICEValA056-1, ICEValA056-2, ICEValE0601, ICEValHN396, ICEValHN437 and ICEValHN492. All ICE sequences were deposited in GenBank under the accession numbers KR231688-KR231689 and KT072768-KT072771.
Table 1.
Features of the ICEs in Vibrio alginolyticus strains
| ICE | Host strain | Sources | Sites and year of isolation | Size (bp) | Identity to IntSXT (%) | Integration site | Resistant genes | Notable variable genes code forb | GenBank accession number |
|---|---|---|---|---|---|---|---|---|---|
| ICEValA056-1 | A056 | Litopeneaus vannamei | Zhanjiang, Guangdong, China, 2003 | 89004 | 97 | prfC | strBA, sul2 | type III RM system, diguanylate cyclase, acriflavin resistance protein, membrane-fusion protein | KR231688 |
| ICEValA056-2 | 103826 | N a | tRNA-ser | N | type I RM system, Fic family protein, HigA, MosTA, choline uptake protein, mechanosensitive channel regulation, calcium/sodium proton antiporter, oxaloacetate and citrate metabolism, PrrABCD | KR231689 | |||
| ICEValE0601 | E0601 | Seawater | Yangjiang, Guangdong, China, 2006 | 106165 | 99 | prfC | N | HipBA, Flp pilus assembly system, P pilus assembly system, type III RM system, threonine efflux protein, choline uptake protein, diguanylate cyclase, DDE endonuclease, mechanosensitive channel regulation, RNA-dependent DNA polymerases | KT072768 |
| ICEValHN492 | HN492 | Seawater | Sanyan, Hainan, China, 2008 | 106164 | 99 | prfC | N | KT072769 | |
| ICEValHN396 | HN396 | Seawater | Qinzhou, Guangxi, China, 2008 | 86687 | N | tRNA-ser | N | type II RM system, Fic family protein, HigA, diguanylate cyclase, aerotaxis sensor, chemotaxis sensor, DNA recombination-mediator, DDE endonuclease | KT072770 |
| ICEValHN437 | HN437 | Seawater | Haikou, Hainan, China, 2008 | 94290 | N | tRNA-ser | N | type I RM system, Fic family protein, HigA, organic hydroperoxide resistance, DDE endonuclease, nucleotide metabolism, phage lysin, rhamnose metabolism, sulfate assimilation, synthesis of capsular polysaccharide | KT072771 |
a N represents that compared genes lack similarity or that no resistant genes were found
bThe functions conferred by unique gene contents that are not found in other ICEs are indicated in boldface
General genetic structures of the ICEs in V. alginolyticus
The genomes of six ICEs were analyzed and compared in this study (Table 1). It is very rare to find two ICEs (ICEValA056-1 and ICEValA056-2) coexisting in one strain (V. alginolyticus A056). The entire length of ICEValA056-1 is 83.4 Kb, coding for 83 open reading frames (ORFs). Among them, 64 ORFs are present in SXT and 67 ORFs in ICEVflind1 (Additional file 1: Table S1). ICEValA056-1 contains a syntenous set of 52 conserved core genes in the SXT/R391 ICEs (Fig. 1). Similar to SXT, specific genes of ICEValA056-1 are inserted into five hotspots (HS1-HS5) and a VR III region, thereby disrupting rumB (Fig. 1). The largest variable DNA insertions are 10.0 and 12.4 Kb, occurring in the VR III and HS5 regions, respectively. ICEValA056-2 has a size of 103.8 Kb, including 108 ORFs (Additional file 2: Table S2). Of the 108 ORFs, 54 are present in SXT, ICEVchMex1 or ICEVchBan8 (an atypical SXT/R391-related ICE), respectively. The largest variable DNAs (22.3 and 12.4 Kb) are inserted into HS3 and HS4, respectively.
Fig. 1.
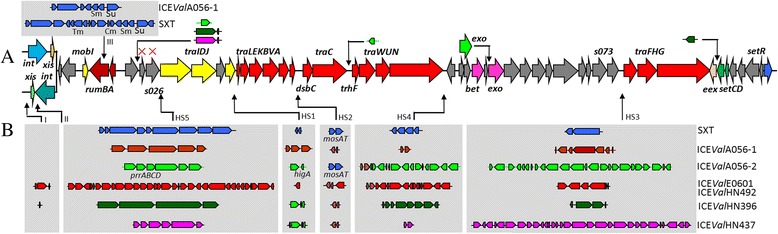
Structural comparison between SXT and six ICEs from V. alginolyticus. a Schematic representation of the 52 core genes of SXT/R391 ICEs. Left lower part is the normal int/xis module of SXT/R391 ICEs, and the upper left part shows the alternative int/xis modules of ICEValA056-2, ICEValHN396 and ICEValHN437. Atypical insertions of ORFs of these ICEs and an extra copy of exo gene (pale green) of ICEValA056-2 are indicated with fold-down arrows. The absences of s025 and s026 in ICEValE0601/ICEValHN492 are shown with red crosses. b Variable ICE regions are shown with colors according to the elements in which they were originally described: SXT (blue), ICEValA056-1 (dark red), ICEValA056-2 (pale green), ICEValE0601/ICEValHN492 (red), ICEValHN396 (dark green) and ICEValHN437 (pink). Thin arrows indicate the sites of insertion for each VR region (I, II and III) and HS1–HS5 represent hotspots 1–5. Some notable genes in these regions are marked. Tm, trimethoprim cassette; Cm, chloramphenicol cassette; Sm, streptomycin cassette; Su, sulfamethoxazole cassette; mosAT, toxin-antitoxin system; prrABCD, an novel operon including a RM system and anticodon nuclease gene. Variable region III is shown at the top of the backbone of SXT/R391 ICEs for saving space
A sequence analysis showed only 14 variable bases dispersed between ICEValE0601 (106.2 Kb) and ICEValHN492 (106.2 Kb) and that all of the genes of ICEValE0601 code amino acid sequences that are identical with the counterparts in ICEValHN492. Therefore, ICEValE0601 and ICEValHN492 were treated as the same ICE. Each of ICEValE0601 and ICEValHN492 contains 113 open ORFs (Additional file 3: Table S3). Among them, 52 ORFs are present in SXT or ICEVchInd4. Variable DNAs of ICEValE0601 or ICEValHN492 are 54.8 Kb in length, which are distributed between five hotspots (HS1-HS5) and VR I (Fig. 1). The largest insertions of variable DNAs are 28.8 Kb and 12.1 Kb, occurring in the HS5 and HS4 regions, respectively.
ICEValHN396 (86.7 Kb) includes 79 ORFs (Additional file 4: Table S4), of which 51 and 54 ORFs are present in SXT and ICEVchICDC-1307, respectively. Variable DNAs of ICEValHN396 with a total size of 31.0 Kb are distributed in five HS regions and the VR I region. The largest variable DNAs are 16.5 and 7.5 Kb and are located in the HS5 and HS4 regions, respectively. ICEValHN437 (94.3 Kb) contains 100 ORFs (Additional file 5: Table S5), of which 51 and 52 ORFs are present in SXT and ICEVchBan9, respectively. Variable DNAs of ICEValHN437 have a total size of 40.1 Kb, which are distributed in five HS regions. The largest variable DNAs are 28.2 and 8.9 Kb, occurring in the HS3 and HS5 regions, respectively.
Core genes of the ICEs from V. alginolyticus
ICEValA056-1 contains a syntenous set of 52 conserved core genes in the SXT/R391 ICEs (Fig. 1 and Additional file 1: Table S1), which are responsible for machinery involved in conjugative transfer, integration/excision and their regulation. Either ICEValE0601 or ICEValHN492 contain 50 out of 52 conserved core genes in the SXT/R391 ICEs (with the exception of genes s025 and s026, which code for predicted proteins of unknown function) (Fig. 1, Additional file 3: Table S3 and Additional file 4: Table S4). S025 and S026 from ICEValA056-1, ICEValA056-2, ICEValHN396 and ICEValHN437 have low identity values (<90 %) with their counterparts from SXT. ICEValA056-1, ICEValE0601 and ICEValHN492 integrate into the 5′ end of the host’s prfC gene targeted by SXT/R391 ICEs [6], and their Int and Xis share at least 97 % identity with their counterparts from SXT.
ICEValA056-2, ICEValHN396 and ICEValHN437 integrate into 3′ end of the tRNA-ser gene (Table 1). This locus was reported to be most commonly targeted by genomic islands or by ICEVchBan8 from a clinical non-toxigenic isolate V. cholerae O37 MZO-3 [24, 25]. int and xis genes of three ICEs locate on the same strand, whereas int and xis genes of SXT/R391 ICEs locate on different strands. Besides, the two genes lack similarity with their counterparts in the SXT/R391 ICEs. However, when the three ICEs are compared with ICEVchBan8, there are three obvious differences between two of the ICE types: (1) int genes of ICEValA056-2, ICEValHN396 and ICEValHN437 only have 90 % identity with the intBan8 on the amino acids level, which is substantially lower than the 98 % average identity among SXT/R391 ICEs; (2) int followed by xis locates at the 5′ end of three ICEs, whereas int and xis of ICEVchBan8 locate at the 3′ end of the ICE; and (3) rumB genes of three ICEs remain intact without any insertion, but ICEVchBan8 has two parts of a truncated rumB gene. BLASTN searches using the complete sequences of ICEValA056-2, ICEValHN396 and ICEValHN437 as queries also indicated that the most closely related ICEs were ICEVchMex1, ICEVchICDC-1307 and ICEVchBan9, respectively. These sequences are members of the SXT/R391 ICEs. To provide a detailed description of the similarities and differences between the three aforementioned ICEs and their closely related ICEs, representative SXT and ICEVchBan8, whole-sequence comparisons of these ICEs were performed using WebACT [26]. The highly similar areas (the same orientation, red) between ICEValA056-2 and ICEVchMex1 (55.7 Kb) or between ICEValA056-2 and SXT (50.1 Kb) are substantially larger than those between ICEValA056-2 and ICEVchBan8 (43.8 Kb) (Fig. 2). Similarity values between ICEValA056-2 and ICEVchMex1 (red areas) are generally higher than those between ICEValA056-2 and SXT or ICEVchBan8 (Fig. 2). Three inversion areas of ICEVchBan8 clearly highlight major rearrangements in it (blue, Fig. 2). Sequence comparisons between ICEValHN396 and SXT, ICEVchICDC-1307 (a member of SXT/R391) or ICEVchBan8 and the comparisons between ICEValHN437 and SXT, ICEVchBan9 or ICEVchBan8 also exhibit similar profiles with the comparison between ICEValA056-2 and related ICEs (Additional file 6: Figure S1 and Additional file 7: Figure S2). Therefore, the analysis demonstrated that ICEValA056-2, ICEValHN396 and ICEValHN437 were more similar to genuine SXT/391 ICEs than to ICEVchBan8, with the exception of the int/xis module.
Fig. 2.
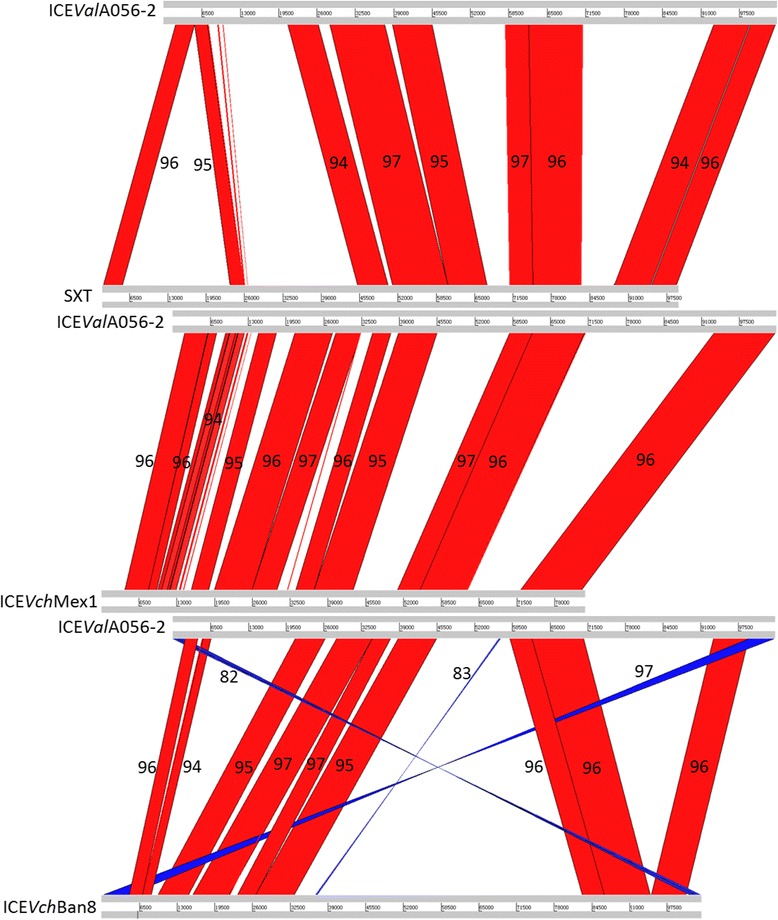
Alignment generated using WebACT for ICEValA056-2, SXT, ICEVchMex1 and ICEVchBan8. Comparisons between the regions < 80 bp are filtered. Numbers show the identity values of the compared regions. Red areas indicate homologous regions; blue areas indicate inversions
To further demonstrate differences in the int/xis modules contributed by ICEValA056-2, ICEValHN396 and ICEValHN437, we created phylogenetic trees based on respective nucleotide sequences to clarify their evolution. traI, one of most divergent core genes in SXT/R391 ICEs [6], was used as a reference gene. The trees for int and xis were extremely similar (Fig. 3a and b). In both trees, the ICEs were segregated into two evolutionarily distinct groups, whereas traI exhibited a completely different branching pattern in which all of the ICEs could not be clustered into different groups (Fig. 3c). In the trees for int and xis, though four ICEs were grouped into the same big branch, ICEVchBan8 in each tree formed an independent clade that was much distant from the clade comprised of ICEValA056-2, ICEValHN396 and ICEValHN437. ICEValA056-2, ICEValHN396 and ICEValHN437 have a very similar core genetic structure; however, each has very different variable DNAs (Fig. 2, Additional file 2: Table S2, Additional file 4: Table S4 and Additional file 5: Table S5), and the strains containing ICEValA056-2, ICEValHN396 and ICEValHN437 were isolated in different time and location. Further data mining in GenBank using their int genes as queries indicated that V. vulnificus CladeA-yb158 (LBNN01000013.1) also potentially contained a complete ICE that was similar to the three ICEs. These results strongly suggest that the occurrence of ICEValA056-2, ICEValHN396 and ICEValHN437 is not a rare event.
Fig. 3.
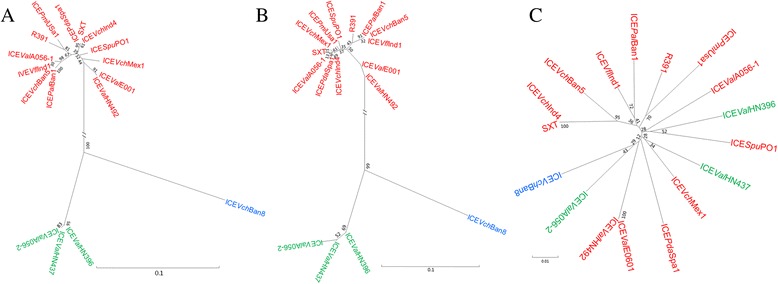
Phylogenetic trees of int (a), xis (b) and traI (c) genes from six ICEs, some randomly selected SXT/R391 ICEs and ICEVchBan8. The trees were constructed using the neighbor-joining method. Bootstrap values were obtained after 1000 repetitions
Five ICEs (with the exception of ICEValA056-1) in this study maintain intact rumB genes without disruption by one typical transposon carrying drug-resistant cassettes. Moreover, it is unusual that ICEValA056-2 contains another version of the exo gene located upstream of a standard exo gene (Fig. 1). The two exo genes share 99 % similarity at the amino acid level.
Usual gene contents of the hotspots and variable regions of the ICEs in V. alginolyticus
The variable genes of six ICEs encode a large array of functions to bestow various adaptive abilities upon their hosts (Table 1, Additional file 1: Tables S1, Additional file 2: Table S2, Additional file 3: Table S3, Additional file 4: Table S4 and Additional file 5: Table S5). A substantial proportion of variable genes have orthologous genes and functionally related genes found in SXT/R391 ICEs, which confer functions typically found in other ICEs. For instances, mosA/T, hipA/B and sole higA (antitoxin gene to higB) encode for toxin-antitoxin (TA) systems that promote ICE maintenance by killing or severely inhibiting the growth of cells that have lost the element [6, 11]. Many genes encode for diverse restriction-modification (RM) systems, helicases and endonucleases. ICEValA056-1, ICEValE0601 and ICEValHN396 contain the genes coding diguanylate cyclase, which are also found in ICEVchMex1, ICEVflInd1 and ICEVchMoz3 [12]. These genes are involved in biofilm formation, motility and virulence in several organisms [27, 28]. Five ICEs with the exception of ICEValA056-2 contain an acetyltransferase gene that is always flanked by a conserved hypothetical gene in HS2 (Fig. 2), and similar genes were only found in ICEVchMex1 and R391. ICEValA056-1 has a truncated copy of a drug-resistant gene cluster in VR III region that is owned by SXT. Similar truncations were also observed in other ICEs such as ICEPalBan1 and ICEVchInd5 [6]. When BLASTN was performed with the whole sequences of any variable DNA regions in ICEValA056-1, highly similar DNA sequences (>98 %) could be found in other ICEs (Additional file 1: Table S1). However, this feature is not obvious in the other five ICEs.
Unique gene contents of the hotspots and variable regions in the ICEs of V. alginolyticus
With the exception of the abovementioned usual genes in hotspots and variable regions shared by other ICEs, six ICEs also contain a large number of unique genes or gene clusters that were not found in other ICEs (Table 1, Additional file 1: Tables S1, Additional file 2: Table S2, Additional file 3: Table S3, Additional file 4: Table S4 and Additional file 5: Table S5). Here, we only analyzed some of their notable genes. A large number of variable genes of ICEValA056-2 code for proteins that are involved in metabolism and transport. The functions of these proteins include choline uptake, the stabilization of the MscS mechano sensitive channel, calcium/sodium proton antiport and alternative metabolic pathways for oxaloacetate and citrate. A novel intact prrABCD operon-like structure is present in the HS5 region of ICEValA056-2 (Fig. 1). No similar continuous structure was found in the other bacterial genomes according to a BLASTN search. prrA, prrB and prrD code for a type I DNA RM system that is similar to hsdMSR in Escherichia coli [29], whereas prrC codes for a putative anticodon nuclease. A related prrABCD system has been reported in E. coli CTr5X [30, 31]. PrrA, PrrB and PrrC of CTr5X only share 82, 33 and 74 % similarity with their counterparts in ICEValA056-2, respectively, whereas the PrrD lacks similarity with the PrrD encoded by ICEValA056-2. Overall, most of the variable genes in ICEValA056-2 could not be found in any other ICEs. Two genes coding for transposases were found to insert in two atypical sites (between s024 and s025 and between traC and trhF) (Fig. 1).
ICEValE0601 bears a large gene cluster composed of at least 22 genes (14.7 Kb) coding for a complex pilus assembly system. A putative threonine efflux gene and a related gene coding hydroxyphenylpyruvate dioxygenase were discovered in ICEValE0601, which were predicted to contribute to amino acid transport and metabolism. A gene coding for MscS mechanosensitive channel protein and a homologous gene to yrbG coding for calcium/sodium proton antiporter occur in ICEValE0601, which were predicted to participate in the regulation of osmotic homeostasis protecting the cell from acute decreases in the osmotic environment. ICEValE0601 contains two dissimilar genes that respectively encode for RNA-dependent DNA polymerase participating in DNA replication, recombination and repair.
ICEValHN396 includes a gene cluster composed of at least six genes. These genes code for a bacterial chemotaxis apparatus that enables the bacteria to direct their movement away from unfavorable chemical stimuli and towards favorable chemical compounds [32, 33]. ICEValHN396 contains the gene dprA and accessory gene recQ. Both ICEValHN396 and ICValHN437 have an atypical insertion of a small hypothetical gene flanked by a transposase gene between s024 and s025 (Fig. 1). In addition to this, ICEValHN396 includes another atypical insertion of a gene coding DDE endonuclease between eex and setC (Fig. 1).
The largest insertion (with a size of 28.2 Kb) occurs in the HS3 region of ICEValHN437 and consists of 30 genes with various functions. The HS3 region contains five known genes coding for UDP-glucose dehydrogenase, UTP-glucose-1-phosphate uridylyltransferase, alpha-L-Rha alpha-1,3-L- rhamnosyltransferase, glycosyl transferase and capsular polysaccharide ABC transporter, respectively. All five genes are involved in the metabolism of nucleotide and carbohydrate and synthesis of capsular polysaccharide. Five known genes are adjoined and dispersed by eight hypothetical genes, among which four hypothetical genes lack homologous genes in GenBank. Based on these features, eight hypothetical genes may be related to nucleotide and carbohydrate metabolism and the synthesis of capsular polysaccharide. The HS3 region also includes a gene cluster comprised of five genes coding for sulfate permease, sulfate adenylyltransferase subunit 2, adenylylsulfate kinase, acetyltransferase and sulfate adenylyltransferase subunit, respectively. Thus, this gene cluster is likely responsible for the metabolic assimilation of sulfur from inorganic sulfate. A sole gene (osmC) is coupled by two transposase genes to form a typical transposon structure. osmC codes for organic hydroperoxide resistance protein; this gene thus likely confers its host with an alternative function to regulate oxidative stress. The genes similar to osmC have never previously been found in Vibrio species.
Intensive distribution of transposase genes in the ICEs from V. alginolyticus
Notably, the ICEs from these V. alginolyticus strains contain abundant transposase genes (Additional file 1: Tables S1, Additional file 2: Table S2, Additional file 3: Table S3, Additional file 4: Table S4 and Additional file 5: Table S5), with an average of over 7 transposase genes per ICE. A similar phenomenon was observed in other SXT/R391 ICEs, such as SXT, ICEVchBan5, ICEVflInd1 and ICEPdaSpa1. The annotations for genomic scaffold sequences of five strains, A056, E0601, HN396, HN437 and HN492, showed that they contained 38, 50, 32, 48 and 43 transposase genes, respectively. The construction of a phylogenetic tree based on the transposase genes in these ICEs failed because they are so highly distinct that no common sites can be found. Blast searches showed that the genes closely matching these distinct transposase genes have extensive sources.
Discussion
SXT/R391 ICEs widely distribute in various marine-sourced bacteria around the world [34]; however, there were no records indicating the emergence of any ICEs in China until we confirmed the presence of SXT/R391-like ICEs in some V. alginolyticus strains in 2012 [19]. To fully understand the features and functions of ICEs in V. alginolyticus, we acquired complete sequences of six ICEs in five randomly selected V. alginolyticus strains. Strains E0601 and HN492 contain extremely similar ICEs (ICEValE0601 and ICEValHN492) that were isolated from various places (linear distance between two sites is nearly 500 km) in 2006 and 2008, respectively. Similar situations have been observed in the strains containing highly similar SXTMO10 and ICEVchInd4 and the strains containing extremely similar ICEVchBan9 and ICEVchMoz10; however, these were isolated at distant places and in different years [6]. With the exception of extreme similarity between ICEValE0601 and ICEValHN492, a high genetic diversity between strains HN492 and E0601 was exhibited through their genomic comparison (data not shown), which indicated that they are different strains. These results strengthened the speculation by Wozniak et al. that some ICEs have recently spread [6] because they have not demonstrated obvious divergent evolution.
Regarding the core genes of the six ICEs, ICEValE0601 and ICEValHN492 lack s025 and s026. In another case, the absence of the core genes s024, s025 and s026 in the atypical element ICEVchBan8 has been observed [24]. Additionally, S025 and S026 from these ICEs have relatively low identity values with their counterparts from SXT. The significant divergence of S026 had been analyzed; however, S025 has not [6]. Beaber et al. showed that the deletion of five cores gene from rumB to s026 (including s024 and s025) had no detectable influence on SXT excision or transfer [20]. These results indicated that though most SXT/R391- or SXT/R391-related ICEs have the core genes s024, s025 and s026, they are not necessary for the maintenance and transfer of ICEs. Thus, these genes can bear more variation due to spontaneous mutations and are potentially lost during recombination events.
ICEValA056-1, ICEValE0601 and ICEValHN492 integrate into the 5′ end of the host’s prfC gene targeted by SXT/R391 ICEs [6], and their Int and Xis share at least 97 % identity with their counterparts from SXT. Together with the fact that they share 48 other conserved core genes (with the exception of s025 and s026), these three ICEs are easily classified into the SXT/R391 family based on the strict definition of SXT/R391 ICEs [34].
The classification of ICEValA056-2, ICEValHN396 and ICEValHN437 presents difficulties because they exhibit a different int/xis module from that in SXT/R391 ICEs and because they integrate into another chromosomal site, the tRNA-ser gene. However, comparative genomics demonstrated that ICEValA056-2, ICEValHN396 and ICEValHN437 were more similar to genuine SXT/391 ICEs than to ICEVchBan8 with the exception of the int/xis module. A phylogenetic analysis showed that though int and xis of ICEVchBan8 were grouped into the same large branch, they formed a very independent clade in the trees of int and xis. These results clearly show that ICEValA056-2, ICEValHN396 and ICEValHN437 form a group that is independent from ICEVchBan8. However, these three ICEs cannot be classed into a new ICE family only based on the different int/xis module and integration site because they retain 50 of 52 conserved core genes in the same organization, which are highly similar to those in SXT/R391 ICEs (with the exception of s025 and s026). traI is one of the most divergent core genes in SXT/R391 ICEs [6] and is theoretically more likely to cluster these ICEs into different groups; however, the opposite is true. Based on these facts and other considerations, we prefer to classify ICEValA056-2, ICEValHN396 and ICEValHN43 into an atypical group of SXT/R391 ICEs. This classification will enrich our knowledge to SXT/R391 ICEs and increase the diversity of SXT/R391 ICEs. Because they are the members of an atypical group in SXT/R391 ICEs, it is interesting to speculate on how these atypical ICEs were generated. ICEs have three discrete functional modules that govern integration/excision, conjugative transfer (DNA processing and mating pair formation) and regulation [2, 3]. Therefore, the primary int/xis module as a whole was likely replaced by the current int/xis module; primary ICEs could have also been replaced by SXT/R391 ICEs, leaving only int/xis modules through potential recombination. The latter appears more reasonable because three ICEs integrate into the tRNA-ser site that SXT/R391 ICEs never inserts into. Inter-ICE recombination was observed between tandem SXT/R391 ICEs, yielding hybrid ICEs with considerable frequency [35]. Comparative genomics also supports the assertion that inter-ICE recombination is commonplace [6].
Our results revealed that the ICEs analyzed in environmental strains generally retain intact rumB genes. The rumB and adjacent rumA genes encode a UN repair DNA polymerase and a UV repair protein, respectively [36]. Sunlight-emitted UV light represents a major source of genotoxic stress to bacteria in the environment [37]; they therefore tend to conserve ICEs devoid of antibiotic resistance genes by retaining a functional rumBA compared with clinical strains not exposed to UV but to antibiotics [38]. Among the six ICEs from V. alginolyticus, only ICEValA056-1 contains drug-resistant genes carrying a transposon. Similar results were also found in another report in which only 3 of 11 environmental Vibrio strains containing ICEs exhibited resistance to sulfamethoxazole and streptomycin, the typical resistance carried on SXT [9]. Additionally, most drug-resistant cassettes in ICEs are carried by transposons, which increases the likelihood that these drug-resistant cassettes were captured after the acquisition of the ICEs by primary hosts. These results demonstrated that drug resistance was not a necessary function of ICEs, especially in environmental bacteria. However, nearly all of the ICEs discovered contain diverse RM systems, helicases and endonucleases, which likely protect hosts from invasion by foreign DNA (including phage infection) and/or promote the integrity of the ICE genome during its transfer between hosts [6]. Thus, the initial biological significance of ICEs to hosts is to provide protection from invasion by phages and plasmids and even from DNA lesions due to environmental factors.
A sequence analysis of VR and HS regions of six ICEs indicated that unbalanced insertions of variable DNAs are very common. For instance, the HS3 region of ICEValHN437 occupies 70 % of variable DNAs. ICEVchBan8 has a similar situation, and nearly all variable DNAs are distributed in its HS4 region [24]. Though five hotspots of the ICEs in V. alginolyticus strains have variable sizes, the insertional sizes in HS1 never exceed 4 Kb and remain relatively stable. A similar phenomenon was observed in 12 out of 13 typical SXT/R391 ICEs with the exception of ICEVflInd1 [6]. Regarding the contents of variable genes, the six ICEs also contain a larger number of variable genes that confer various functions to their hosts. Here, we only discuss some unique genes discovered in these six ICEs. ICEValE0601 bears a large gene cluster coding for a complex pilus assembly system. Pili are involved in adhesion to host cells, motility and DNA exchange [39, 40], and pili are often crucial virulence factors in pathogenic bacteria because they mediate attachment to and infection of target cells and are involved in evasion of the host immune system [41]. ICEValHN396 contains the gene dprA, which codes for DNA recombination mediator protein A. This gene is predicted to bind cooperatively to single-stranded DNA (ssDNA) and to interact with RecA [42, 43]. In the process, DprA-RecA-ssDNA filaments are produced; these filaments catalyze the homology-dependent formation of joint molecules [42, 43]. DprA is a new member of the recombination-mediator protein family and performs natural bacterial transformation [42–44]. This indicates that ICEs not only increase the genetic diversity of their hosts in their integration but also have the ability to facilitate bacterial gene exchange via natural bacterial transformation. Many alternatively metabolic pathways were discovered in six ICEs. As a member of bacterioplankton, V. alginolyticus widely distributes in various marine environments through ocean currents. Increased metabolic pathways could potentially strengthen the fitness of V. alginolyticus in various environments.
In our study, abundant transposase genes were discovered in six ICEs from V. alginolyticus and other ICEs. Transposase genes are the most abundant genes in both completely sequenced genomes and environmental metagenomes and are also the most ubiquitous in metagenomes [45, 46]. Previous statistics from 630 bacterial genomes by Aziz et al. also showed that the average number of transposase genes in bacteria was 40 per genome [46]. Considering this average value and the size of the ICEs (generally less than 150 Kb), transposase genes may have a more intensive distribution in these ICEs than in any other parts of their host genomes. Transposase genes encode DNA-binding enzymes that catalyze ‘cut-and-paste’ or ‘copy-and-paste’ reactions to promote the integration of DNA segments into new sites [47]. Most of the described transposase genes are the core parts of transposable genetic elements (TEs), consisting of insertion sequence (IS) elements and their resulting transposon elements (Tn), which play an important role in bacterial fitness and evolution through horizontal gene transfer [48–50]. Based on these analyses, these transposase genes likely integrated into the hotspots or variable regions of ICEs after the creation of the ICEs. The acquisition of TEs does not always benefit hosts; TEs can exhibit detrimental effects by inactivating housekeeping genes or impairing the chromosome’s structure due to transposition to unsuitable sites [26, 48, 51, 52]. The accommodation of various TEs into one relatively unimportant genomic locus exhibits fewer hazards than that into multiple dispersed loci. ICEs can represent an ideal target for the ‘foothold’ of these TEs due to their insertional sites and containing capability. This may explain why transposase genes occur intensively in ICEs. ICEs have sophisticated transfer machinery and can transfer between genetically distant species at a relatively high frequency of 10−5-10−6 per donor or recipient [7, 20, 22, 53]; consequently, ICEs may serve as a carrier for TEs and largely facilitate the dissemination of TEs.
Conclusions
In this study, we acquired complete sequences for six new ICEs in V. alginolyticus strains. A comparative genomic analysis showed that ICEValA056-1, ICEValE0601 and ICEValHN492 are typical members of the SXT/R391 family. ICEValA056-2, ICEValHN396 and ICEValHN437 represent an atypical group in SXT/R391 family. The variable genes of six ICEs encode a large array of functions to bestow various adaptive abilities upon their hosts. Many variable genes have orthologous and functionally related genes found in SXT/R391 ICEs. Six ICEs also contain a large number of unique genes or gene clusters that were not found in other ICEs. Diverse transposase genes intensively occur in the ICEs, and ICEs may serve as a carrier for TEs and largely facilitate the dissemination of TEs.
Methods
Genome sequencing and assemblies of six ICEs from V. alginolyticus strains
Five V. alginolyticus strains, A056, E0601, HN396, HN437 and HN492 (their sources are listed in Table 1), were sequenced on the HiSeq 2000 sequencing platform (BGI, China) with a paired-end 2 × 100-nucleotide (nt) procedure. De novo assemblies were performed through the CLC Genomics Work Bench and SeqMan [54].
Genome sequences of V. alginolyticus strains were annotated using the RAST annotation pipeline [23]. The scaffolds containing fragments of ICE elements were singled out and aligned manually with ICE sequences of SXT (AY055428.1), ICEVchInd4 (GQ463141.1) and ICEVchMex1 (GQ463143.1) as references. Gaps between two scaffolds were closed using manual editing via PCR amplification and sequencing.
PCR conditions
PCR primers are listed in Additional file 8: Table S6. All PCRs for gap filling were performed in a 50-μl reaction containing 1 μl of genomic DNA, 0.4 μM of each primer, 5 μl of 10 × PCR buffer, 0.2 mM dNTP and 1 U of Taq DNA polymerase (Takara, China). The amplification program consisted of an initial denaturation at 94 °C for 4 min, 30 cycles of denaturation at 94 °C for 20 s, annealing at 54–58 °C for 30 s, and extension at 72 °C for 1–2 min with a final extension at 72 °C for 7 min.
Bioinformatics
Related ICEs sequences were searched via BLASTN using the entire sequence of each ICE as a query. The ICEValA056-2, ICEValHN396 and ICEValHN437 sequences were compared with related ICE sequences using WebACT [26]. Similarities in nucleotide and protein sequences for two ICEs were determined as the % nucleotide or amino acid identity with related ICEs by employing BLASTN and BLASTP. The genes analyzed were aligned using ClustalW in MEGA 6.0, and phylogenetic trees were constructed using the neighbor-joining method in MEGA 6.0. The reliability of each tree was subjected to a bootstrap test of 1000 repetitions.
Acknowledgments
The research was supported by a grant from the Natural Science Foundation of China (31370149), the Comprehensive Strategic Cooperation Project of Guangdong Province and Chinese Academy of Sciences (2012B091100269) and Knowledge Innovation Program of CAS (KSCX2-EW-G-12B).
Additional files
ORFs in ICEValA056-1 and their similarity with related ICEs. (DOCX 21 kb)
ORFs in ICEValA056-2 and their similarity with related ICEs. (DOCX 22 kb)
ORFs in ICEValE0601 (ICEValHN492) and their similarity with related ICEs. (DOCX 21 kb)
ORFs in ICEValHN396 and their similarity with related ICEs. (DOCX 19 kb)
ORFs in ICEValHN437 and their similarity with related ICEs (DOCX 19 kb)
Alignment generated using WebACT of ICEValHN396, SXT, ICEVchICDC-1307 and ICEVchBan8. Comparisons between the regions < 80 bp are filtered. Numbers show the identity values of the compared regions. Red areas indicate homologous regions; blue areas indicate inversions. (TIF 522 kb)
Alignment generated using WebACT of ICEValHN437, SXT, ICEVchBan9 and ICEVchBan8. Comparisons between the regions < 80 bp are filtered. Numbers show the identity values of the compared regions. Red areas indicate homologous regions; blue areas indicate inversions. (TIF 493 kb)
Primers used for gap filling in this study. (DOCX 15 kb)
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
PL wrote the manuscript and designed the experiments. XYH performed analysis of bioinformatics. YHW carried out PCRs to fill all the gaps between scaffolds. QTL performed partial analysis of bioinformatics. CQH guided the experiments and contributed to the writing of the manuscripts. All authors read and approved the final manuscript.
References
- 1.Wozniak RA, Waldor MK. Integrative and conjugative elements: mosaic mobile genetic elements enabling dynamic lateral gene flow. Nat Rev Microbiol. 2010;8:552–63. doi: 10.1038/nrmicro2382. [DOI] [PubMed] [Google Scholar]
- 2.Bi D, Xu Z, Harrison EM, Tai C, Wei Y, He X, et al. ICEberg: a web-based resource for integrative and conjugative elements found in bacteria. Nucleic Acids Res. 2012;40:621–6. doi: 10.1093/nar/gkr846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Garriss G, Burrus V. Integrating conjugative elements of the SXT/R391 family. In: Roberts AP, Mullany P, editors. Bacterial integrative mobile genetic elements. Austin: Landes Biosciences; 2013. pp. 217–34. [Google Scholar]
- 4.Coetzee JN, Datta N, Hedges RW. R factors from Proteus rettgeri. J Gen Microbiol. 1972;72:543–52. doi: 10.1099/00221287-72-3-543. [DOI] [PubMed] [Google Scholar]
- 5.Waldor MK, Tschape H, Mekalanos JJ. A new type of conjugative transposon encodes resistance to sulfamethoxazole, trimethoprim, and streptomycin in Vibrio cholerae O139. J Bacteriol. 1999;178:4157–65. doi: 10.1128/jb.178.14.4157-4165.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wozniak RA, Fouts DE, Spagnoletti M, Colombo MM, Ceccarelli D, Garriss G, et al. Comparative ICE genomics: insights into the evolution of the SXT/R391 family of ICEs. PLoS Genet. 2009;5:e1000786. doi: 10.1371/journal.pgen.1000786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rodriguez-Blanco A, Lemos ML, Osorio CR. Integrating conjugative elements as vectors of antibiotic, mercury, and quaternary ammonium compound resistance in marine aquaculture environments. Antimicrob Agents Chemother. 2012;56:2619–26. doi: 10.1128/AAC.05997-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Badhai J, Kumari P, Krishnan P, Ramamurthy T, Das SK. Presence of SXT integrating conjugative element in marine bacteria isolated from the mucus of the coral Fungia echinata from Andaman Sea. FEMS Microbiol Lett. 2013;338:118–23. doi: 10.1111/1574-6968.12033. [DOI] [PubMed] [Google Scholar]
- 9.Song Y, Yu P, Li B, Pan Y, Zhang X, Cong J, et al. The mosaic accessory gene structures of the SXT/R391-like integrative and conjugative elements derived from Vibrio spp. isolated from aquatic products and environment in the Yangtze River Estuary, China. BMC Microbiol. 2013;13:214. doi: 10.1186/1471-2180-13-214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Spagnoletti M, Ceccarelli D, Rieux A, Fondi M, Taviani E, Fani R, et al. Acquisition and evolution of SXT-R391 integrative conjugative elements in the seventh-pandemic Vibrio cholerae lineage. mBio. 2014;5:e01356–14. doi: 10.1128/mBio.01356-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wozniak RAF, Waldor MK. A toxin-antitoxin system promotes the maintenance of an integrative conjugative element. PLoS Genet. 2009;5:e1000439. doi: 10.1371/journal.pgen.1000439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bordeleau E, Brouillette E, Robichaud N, Burrus V. Beyond antibiotic resistance: integrating conjugative elements of the SXT/R391 family that encode novel diguanylate cyclases participate to c-di-GMP signalling in Vibrio cholerae. Environ Microbiol. 2010;l12:510–23. doi: 10.1111/j.1462-2920.2009.02094.x. [DOI] [PubMed] [Google Scholar]
- 13.Balado M, Lemos ML, Osorio CR. Integrating conjugative elements of the SXT/R391 family from fish-isolated vibrios encode restriction-modification systems that confer resistance to bacteriophages. FEMS Microbiol Ecol. 2013;l83:457–67. doi: 10.1111/1574-6941.12007. [DOI] [PubMed] [Google Scholar]
- 14.Carraro N, Poulin D, Burrus V. Replication and active partition of integrative and conjugative elements (ICEs) of the SXT/R391 family: the line between ICEs and conjugative plasmids is getting thinner. PLoS Genet. 2015;10:e1005298. doi: 10.1371/journal.pgen.1005298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen MX, Li HY, Li G, Zheng TL. Distribution of Vibrio alginolyticus-like species in Shenzhen coastal waters, China. Braz J Microbiol. 2011;l42:884–96. doi: 10.1590/S1517-83822011000300007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liu XF, Cao Y, Zhang HL, Chen YJ, Hu CJ. Complete genome sequence of Vibrio alginolyticus ATCC 17749T. Genome Announc. 2015;3:e01500–14. doi: 10.1128/genomeA.01500-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Daniels NA, Shafaie A. A review of pathogenic Vibrio infections for clinicians. Infect Med. 2000;17:665–85. [Google Scholar]
- 18.Luo P, Hu C. Vibrio alginolyticus gyrB sequence analysis and gyrB-targeted PCR identification in environmental isolates. Dis Aquat Organ. 2008;82:209–16. doi: 10.3354/dao01984. [DOI] [PubMed] [Google Scholar]
- 19.Luo P, Jiang H, Wang Y, Su T, Hu C, Ren C, Jiang X. Prevalence of mobile genetic elements and transposase genes in Vibrio alginolyticus from the southern coastal region of China and their role in horizontal gene transfer. Int Microbiol. 2012;15:201–10. doi: 10.2436/20.1501.01.172. [DOI] [PubMed] [Google Scholar]
- 20.Beaber JW, Hochhut B, Waldor MK. Genomic and functional analyses of SXT, an integrating antibiotic resistance gene transfer element derived from Vibrio cholerae. J Bacteriol. 2002;184:4259–69. doi: 10.1128/JB.184.15.4259-4269.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hochhut B, Beaber JW, Woodgate R, Waldor MK. Formation of chromosomal tandem arrays of the SXT element and R391, two conjugative chromosomally integrating elements that share an attachment site. J Bacteriol. 2001;183:1124–32. doi: 10.1128/JB.183.4.1124-1132.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kiiru JN, Saidi SM, Goddeeris BM, Wamae NC, Butaye P, Kariuki SM. Molecular characterization of Vibrio cholerae O1 strains carrying an SXT/R391-like element from cholera outbreaks in Kenya: 1994–2007. BMC Microbiol. 2009;9:275. doi: 10.1186/1471-2180-9-275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA, et al. The RAST Server: rapid annotations using subsystems technology. BMC Genomics. 2008;9:75. doi: 10.1186/1471-2164-9-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Taviani E, Spagnoletti M, Ceccarelli D, Haley BJ, Hasan NA, Chen A, et al. Genomic analysis of ICEVchBan8: An atypical genetic element in Vibrio cholerae. FEBS Lett. 2012;586:1617–21. doi: 10.1016/j.febslet.2012.03.064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Haley BJ, Grim CJ, Hasan NA, Choi SY, Chun J, Brettin TS, et al. Comparative genomic analysis reveals evidence of two novel Vibrio species closely related to V. cholerae. BMC Microbiol. 2010;10:154. doi: 10.1186/1471-2180-10-154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Abbott JC, Aanensen DM, Rutherford K, Butcher S, Spratt BG. WebACT—an online companion for the artemis comparison tool. Bioinformatics. 2005;21:3665–6. doi: 10.1093/bioinformatics/bti601. [DOI] [PubMed] [Google Scholar]
- 27.Romling U, Gomelsky M, Galperin MY. C-di-GMP: the dawning of a novel bacterial signaling system. Mol Microbiol. 2005;57:629–39. doi: 10.1111/j.1365-2958.2005.04697.x. [DOI] [PubMed] [Google Scholar]
- 28.Jenal U, Malone J. Mechanisms of cyclic-di-GMP signaling in bacteria. Annu Rev Genet. 2006;40:385–407. doi: 10.1146/annurev.genet.40.110405.090423. [DOI] [PubMed] [Google Scholar]
- 29.Kaufmann G. Anticodon nucleases. Trends Biochem Sci. 2000;25:70–4. doi: 10.1016/S0968-0004(99)01525-X. [DOI] [PubMed] [Google Scholar]
- 30.Levitz R, Chapman D, Amitsur M, Green R, Snyder L, Kaufmann G. The optional E. coli prr locus encodes a latent form of phage T4-induced anticodon nuclease. EMBO J. 1990;9:1383–9. doi: 10.1002/j.1460-2075.1990.tb08253.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Tyndall C, Meister J, Bickle TA. The Escherichia coli prr region encodes a functional type IC DNA restriction system closely integrated with an anticodon nuclease gene. J Mol Biol. 1994;237:266–74. doi: 10.1006/jmbi.1994.1230. [DOI] [PubMed] [Google Scholar]
- 32.Wadhams GH, Armitage JP. Making sense of it all: bacterial chemotaxis. Nat Rev Mol Cell Biol. 2004;5:1024–37. doi: 10.1038/nrm1524. [DOI] [PubMed] [Google Scholar]
- 33.Sourjik V, Armitage JP. Spatial organization in bacterial chemotaxis. EMBO J. 2010;29:2724–33. doi: 10.1038/emboj.2010.178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Burrus V, Marrero J, Waldor MK. The current ICE age: biology and evolution of SXT-related integrating conjugative elements. Plasmid. 2006;55:173–83. doi: 10.1016/j.plasmid.2006.01.001. [DOI] [PubMed] [Google Scholar]
- 35.Burrus V, Waldor MK. Formation of SXT tandem arrays and SXT-R391 hybrids. J Bacteriol. 2004;186:2636–45. doi: 10.1128/JB.186.9.2636-2645.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Boltner D, MacMahon C, Pembroke JT, Strike P, Osborn AM. R391: a conjugative integrating mosaic comprised of phage, plasmid, and transposon elements. J Bacteriol. 2002;184:5158–69. doi: 10.1128/JB.184.18.5158-5169.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Charpentier X, Kay E, Schneider D, Shuman HA. Antibiotics and UV radiation induce competence for natural transformation in Legionella pneumophila. J Bacteriol. 2011;193:1114–21. doi: 10.1128/JB.01146-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Taviani E, Ceccarelli D, Lazaro N, Bani S, Cappuccinelli P, Colwell RR, et al. Environmental Vibrio spp., isolated in Mozambique, contain a polymorphic group of integrative conjugative elements and class1 integrons. FEMS Microbiol Ecol. 2008;64:45–54. doi: 10.1111/j.1574-6941.2008.00455.x. [DOI] [PubMed] [Google Scholar]
- 39.Proft T, Baker EN. Pili in Gram-negative and Gram-positive bacteria- structure, assembly and their role in disease. Cell Mol Life Sci. 2009;64:613–35. doi: 10.1007/s00018-008-8477-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Barken KB, Sünje JP, Yang L, Gjermansen M, Bertrand JJ, Klausen M, et al. Roles of type IV pili, flagellum-mediated motility and extracellular DNA in the formation of mature multicellular structures in Pseudomonas aeruginosa biofilms. Environ Microbiol. 2008;10:2331–43. doi: 10.1111/j.1462-2920.2008.01658.x. [DOI] [PubMed] [Google Scholar]
- 41.Allen WJ, Phan G, Waksman G. Pilus biogenesis at the outer membrane of Gram-negative bacterial pathogens. Curr Opin Struct Biol. 2012;22:500–6. doi: 10.1016/j.sbi.2012.02.001. [DOI] [PubMed] [Google Scholar]
- 42.Mortier-Barrière I, Velten M, Dupaigne P, Mirouze N, Piétrement O, McGovern S, et al. A key presynaptic role in transformation for a widespread bacterial protein: DprA conveys incoming ssDNA to RecA. Cell. 2007;130:824–36. doi: 10.1016/j.cell.2007.07.038. [DOI] [PubMed] [Google Scholar]
- 43.Lisboa J, Andreani J, Sanchez D, Boudes M, Collinet B, Liger D. Molecular determinants of the DprA-RecA interaction for nucleation on ssDNA. Nucleic Acids Res. 2014;42:7395–408. doi: 10.1093/nar/gku349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Smeets LC, Bijlsma JJ, Kuipers EJ, Vandenbroucke-Grauls CM, Kusters JG. The dprA gene is required for natural transformation of Helicobacter pylori. FEMS Immunol Med Microbiol. 2000;27:99–102. doi: 10.1111/j.1574-695X.2000.tb01418.x. [DOI] [PubMed] [Google Scholar]
- 45.Ochman H, DavalosL M. The nature and dynamics of bacterial genomes. Science. 2006;311:1730–3. doi: 10.1126/science.1119966. [DOI] [PubMed] [Google Scholar]
- 46.Aziz RK, Breibbart M, Edwards RA. Transposases are the most abundant, most ubiquitous genes in nature. Nucl Acids Res. 2010;38:4207–17. doi: 10.1093/nar/gkq140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rice PA, Baker TA. Comparative architecture of transposase and integrase complexes. Nat Struct Biol. 2001;8:302–7. doi: 10.1038/86166. [DOI] [PubMed] [Google Scholar]
- 48.Blot M. Transposable elements and adaptation of host bacteria. Genetica. 1994;93:5–12. doi: 10.1007/BF01435235. [DOI] [PubMed] [Google Scholar]
- 49.Syvanen M. Insertion sequences and their evolutionary role. In: de Bruijn FJ, editor. Bacterial genomes. London: Chapman & Hall; 1998. p. 213–20.
- 50.Schneider D, Lenski RE. Dynamics of insertion sequence elements during experimental evolution of bacteria. Res Microbiol. 2004;165:319–27. doi: 10.1016/j.resmic.2003.12.008. [DOI] [PubMed] [Google Scholar]
- 51.Mahillon J, Chandler M. Insertion sequences. Microbiol Mol Biol Rev. 1998;62:725–74. doi: 10.1128/mmbr.62.3.725-774.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Mills RE, Bennett EA, Iskow RC, Devine SE. Which transposable elements are active in the human genome? Trends Genet. 2007;23:183–91. doi: 10.1016/j.tig.2007.02.006. [DOI] [PubMed] [Google Scholar]
- 53.Osorio CR, Marrero J, Wozniak RA, Lemos ML, Burrus V, Waldor MK. Genomic and functional analysis of ICEPdaSpa1, a fish-pathogen -derived SXT-related integrating conjugative element that can mobilize a virulence plasmid. J Bacteriol. 2008;190:3353–61. doi: 10.1128/JB.00109-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Swindell SR, Plasterer TN. SEQMAN. Contig assembly. Methods Mol Biol. 1997;70:75–89. [PubMed] [Google Scholar]