

Abstract
Background
The blacklegged tick Ixodes scapularis transmits Borrelia burgdorferi (sensu stricto) in eastern North America; however, the agent of Lyme disease is not the sole pathogen harbored by the blacklegged tick. The blacklegged tick is expanding its range into areas of southern Canada such as Ontario, an area where exposure to blacklegged tick bites and tick-borne pathogens is increasing. We performed a systematic review to evaluate the public health risks posed by expanding blacklegged tick populations and their associated pathogens.
Methods
We followed PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines for conducting our systematic review. We searched Ovid MEDLINE, Embase, BIOSIS, Scopus and Environment Complete databases for studies published from 2000 through 2015, using subject headings and keywords that included “Ixodes scapularis”, “Rickettsia”, “Borrelia”, “Anaplasma”, “Babesia” and “pathogen.” Two reviewers screened titles and abstracts against eligibility criteria (i.e. studies that included field-collected blacklegged ticks and studies that did not focus solely on B. burgdorferi) and performed quality assessments on eligible studies.
Results
Seventy-eight studies were included in the final review, 72 were from the US and eight were from Canada (two studies included blacklegged ticks from both countries). Sixty-four (82 %) studies met ≥ 75 % of the quality assessment criteria. Blacklegged ticks harbored 91 distinct taxa, 16 of these are tick-transmitted human pathogens, including species of Anaplasma, Babesia, Bartonella, Borrelia, Ehrlichia, Rickettsia, Theileria and Flavivirus. Organism richness was highest in the Northeast (Connecticut, New York) and Upper Midwest US (Wisconsin); however, organism richness was dependent on sampling effort. The primary tick-borne pathogens of public health concern in Ontario, due to the geographic proximity or historical detection in Ontario, are Anaplasma phagocytophilum, Babesia microti, B. burgdorferi, Borrelia miyamotoi, deer tick virus and Ehrlichia muris-like sp. Aside from B. burgdorferi and to a much lesser concern A. phagocytophilum, these pathogens are not immediate concerns to public health in Ontario; rather they represent future threats as the distribution of vectors and pathogens continue to proliferate.
Conclusions
Our review is the first systematic assessment of the literature on the human pathogens associated with the blacklegged tick. As Lyme disease awareness continues to increase, it is an opportune time to document the full spectrum of human pathogens transmittable by blacklegged ticks.
Electronic supplementary material
The online version of this article (doi:10.1186/s13071-016-1529-y) contains supplementary material, which is available to authorized users.
Keywords: Bacteria, Blacklegged ticks, Infectious disease, Ixodes, Parasites, Pathogens, Public health, Symbionts, Vector-borne, Viruses, Zoonoses
Background
The blacklegged tick Ixodes scapularis is the vector of Borrelia burgdorferi (sensu stricto) (agent of Lyme disease) in eastern North America. Blacklegged ticks are three-host, non-nidiculous ticks with larvae and nymphs that feed on small rodents (e.g. white-footed mouse) and passerine birds, and adults that feed on large mammals (white-tailed deer, humans). The blacklegged tick’s range has been expanding northward from its precinctive habitats in the Northeast and Upper Midwest US over the last several decades [1–3]. In Ontario, this range expansion has not been uniform; spreading primarily into suburban and rural areas with mixed deciduous forests, where vertebrate hosts are abundant and local climate is favourable to blacklegged tick survival [4]. Until the mid-1990s, B. burgdorferi and Babesia microti (babesiosis) were the only pathogens known to be transmitted by blacklegged ticks [5–7]. Blacklegged ticks were soon implicated as vectors of Anaplasma phagocytophilum (anaplasmosis), and more recently, Borrelia miyamotoi (B. miyamotoi disease) and deer tick virus (DTV; DTV encephalitis) [8–10]. Uncommon in Ontario’s blacklegged ticks, so far, these pathogens are likely to become more prevalent in the future, as has been the case in recently-invaded jurisdictions such as Maine [11].
In order to assess the public health risks due to blacklegged ticks, an understanding of blacklegged tick-associated organisms is essential, particularly their distribution, prevalence and capacity to cause human disease. We performed a systematic review of the scientific literature to identify organisms (targeting human pathogens) associated with blacklegged ticks in eastern North America and to assess which organisms pose a threat to the health of Ontarians.
Methods
Search strategy
We followed PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines for conducting our systematic review [12]. With the assistance of Public Health Ontario’s Library Services, we developed our primary search strategy in Medline and customized it for other databases to account for database-specific vocabulary and functionality. Our search used subject headings and keywords that included “North America”, “Ixodes scapularis”, “Rickettsia”, “Bartonella”, “Borrelia”, “Anaplasma”, “Babesia”, “Powassan”, “pathogen” and “blacklegged tick.” We conducted a systematic review of English-language studies using five electronic databases: Ovid MEDLINE(R) In-Process & Other Non-Indexed Citations and Ovid MEDLINE(R) (Ovid platform: 1 January 1995 to 20 April 2015); Embase (Ovid platform: 1 January 1996 to Week 16, 2015); Scopus (1 January 1999 to 20 April 2015); Environment Complete (EBSCOhost Research Databases: 1 January 1995 to 20 April 2015); and BIOSIS Previews (Ovid platform: 1 January 2002 to Week 20, 2015). All searches are current as of 20 April 2015 (full search strategy for Ovid Medline, Additional file 1).
Study selection
Studies included in the review were required to meet the following eligibility criteria: (i) included field-collected I. scapularis from Canada or US; (ii) published on or after 1 January 1995; and (iii) did not test solely for B. burgdorferi. We limited studies to those that tested for at least one organism besides B. burgdorferi as our emphasis was on the incidence and prevalence of organisms other than B. burgdorferi. Studies that focused on experimental models–in the absence of field-collected blacklegged ticks–were excluded (e.g. modeling studies, studies with experimental infections of hosts or ticks), as were studies concentrating on human case reports, patient treatment or blood-donor screening (as these did not specifically link an organism to field-collected blacklegged ticks). We limited the final number of studies to those published on or after 1 January 2000; we did not identify any additional organisms from studies published from 1 January 1995 through 31 December 1999. Two reviewers independently screened titles and abstracts against eligibility criteria and differences were resolved by consensus (Mark P Nelder, Nina Jain Sheehan) (Fig. 1). We excluded one study because it did not identify the state(s) where the ticks were collected [13].
Fig. 1.
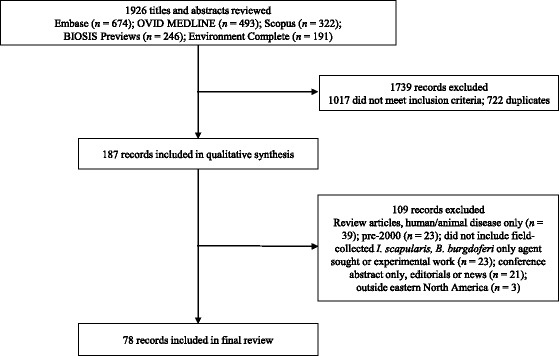
Literature search and study selection
Data extraction and quality assessment
We populated a data extraction table with the study’s first author, year(s) of study, location of study, tick or host collection methods, organisms tested for (gene targets), key organism detection methods, prevalence of organisms (by tick stage), explicit mention of positive and negative pathogen controls, acknowledgement of sequence submissions to GenBank (where the organism identification was uncertain) (Additional file 2).
To evaluate the quality of eligible studies and to reduce the risk of bias, critical appraisals for each study were completed by two independent reviewers and disagreements resolved by consensus (Curtis B Russell, Mark P Nelder; Additional file 3). We completed critical appraisals of studies using the Public Health Ontario Meta-tool for Quality Appraisal of Public Health Evidence, a composite tool based on multiple underlying instruments (e.g. STROBE, STrengthening the Reporting of OBservational studies in Epidemiology) [14]. We assessed all studies based upon relevancy, reliability, validity and applicability. The independent reviewers did not calculate an overall quality score, in keeping with the agreement in the literature [12].
Data synthesis and analysis
We structured study outcomes by organism detected, jurisdiction (province or state) where detected and crude prevalence (Table 1; Figs. 2 and 3). Pathogen prevalence is reported as a crude prevalence, indicating the prevalence reported includes pooled tick stages (larvae, nymphs and adults), along with pooled engorgement status, collection methods and pathogen detection methods. To determine what tick-borne diseases are reportable to public health officials, we reviewed reportable disease lists for Anaplasma, Babesia, Bartonella, Borrelia, DTV, Ehrlichia and Rickettsia in Canada (Manitoba, Ontario and Quebec) and the US (Illinois, Indiana, Michigan, Minnesota, New York, Ohio, Pennsylvania and Wisconsin) (Table 2). We created maps presenting crude prevalence (total positive ticks/total ticks tested in a province or state) for the pathogens of concern (Figs. 2 and 3) using the Environmental Systems Research Institute (ESRI) ArcMap Geographic Information System software (v10.3.1), manually choosing data classes, cut-off values and map colours for each of the generated maps. We calculated Pearson product-moment correlation coefficients (significance determined by linear regressions) to test the linear association between the number of organisms detected and the number of (i) studies conducted in a jurisdiction, (ii) ticks tested in a jurisdiction and (iii) organisms sought in a jurisdiction.
Table 1.
Summary of tick-borne, reportable pathogens and diseases for select provinces and statesa
| Country (province or state) | Arboviruses | Anaplasma | Babesia | Bartonella | Borrelia | Ehrlichia | Rickettsia |
|---|---|---|---|---|---|---|---|
| Canada | |||||||
| Ontario [120] | Encephalitis (primary viral) | NR | NR | NR | Lyme disease (B. burgdorferi) | NR | NR |
| Manitoba [121] | NR | Anaplasmosis (A. phagocytophilum) | Babesiosis (Babesia spp.) | NR | Lyme disease (B. burgdorferi) | NR | NR |
| Quebec [122] | Encephalitis (arthropod-borne) | NR | NR | NR | Lyme disease (B. burgdorferi) | Ehrlichiosis (E. chaffeensis) | NR |
| US | |||||||
| Illinois [123] | Arboviruses | Anaplasmosis | Babesiosis | NR | Lyme disease | Ehrlichiosis | RMSF |
| Indiana [124] | Encephalitis (POWV) | A. phagocytophilum | Babesiosis (Babesia spp.) | NR | Lyme disease (B. burgdorferi) | Ehrlichiosis (E. chaffeensis) | RMSF, other spotted fevers (Rickettsia spp.) |
| Michigan [125] | Arboviral encephalitides (POWV) | Anaplasmosis (A. phagocytophilum) | Babesiosis (Ba. microti) | Cat-scratch fever (Bartonella spp.) | Lyme disease (B. burgdorferi) | Ehrlichia spp. | Spotted fevers (Rickettsia spp.) |
| Minnesota [126] | Arboviral disease (POWV) | Anaplasmosis (A. phagocytophilum) | Babesiosis (Babesia spp.) | Cat-scratch disease (Bartonella spp.) | Lyme disease (B. burgdorferi) | Ehrlichiosis (E. chaffeensis, E. muris-like sp. and E. ewingii) | RMSF, other spotted fevers (R. rickettsii), Rickettsia spp. |
| New York [127] | Arboviral infection (POWV) | Anaplasmosis (A. phagocytophilum) | Babesiosis (Babesia spp.) | NR | Lyme disease (B. burgdorferi) | Ehrlichiosis (Ehrlichia spp.) | RMSF, other spotted fevers (R. akari, R. rickettsii) |
| Ohio [128] | Arbovirus (POWV) | Anaplasmosis | Babesiosis | NR | Lyme disease | Ehrlichiosis | RMSF, other spotted fevers (Rickettsia spp.) |
| Pennsylvania [129] | Arboviruses | Anaplasmosis | NR | NR | Lyme disease | Ehrlichiosis | RMSF, rickettsialpox (Rickettsia spp.) |
| Wisconsin [130] | Arboviral infection (encephalitis, meningitis) | Anaplasmosis | Babesiosis | NR | Lyme disease | Ehrlichiosis | RMSF |
aNR, not reportable; RMSF, Rocky Mountain spotted fever; POWV, Powassan encephalitis virus
Fig. 2.
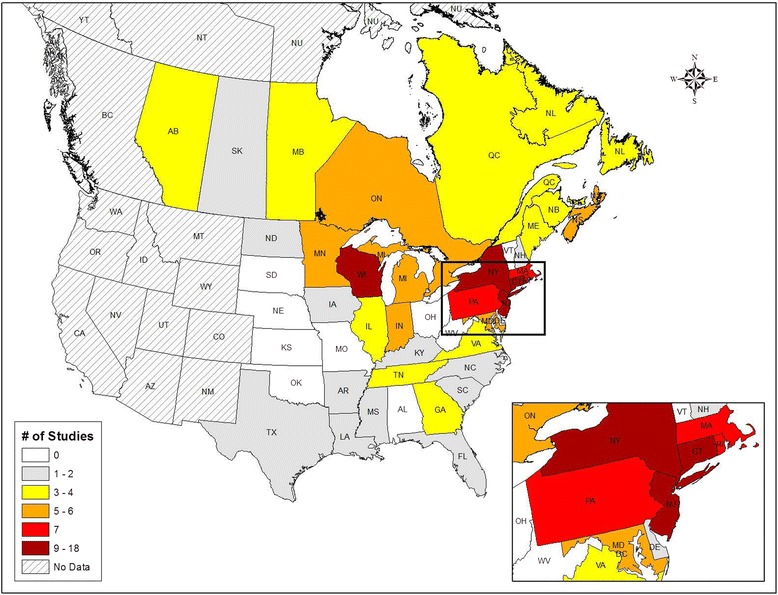
Number of studies performed in each province or state from reviewed studies (2000–2015). No Data, no studies from these jurisdictions were included in the review
Fig. 3.
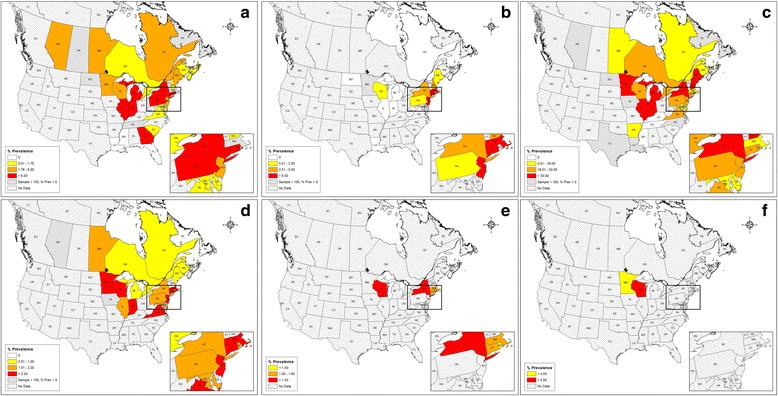
Distribution and crude prevalence for human pathogens transmitted by Ixodes scapularis in eastern North America (2000–2015). a Anaplasma phagocytophilum, b Babesia microti, c Borrelia burgdorferi, d Borrelia miyamotoi, e DTV/POWV, f Ehrlichia muris-like
Table 2.
Distribution of Ixodes scapularis organisms in eastern North America, from 78 studies reviewed (2000–2015)
| Organism | Jurisdiction where organism detected (Canadian provinces and US states)a |
|---|---|
| Tick-borne organisms | |
| Alphaproteobacteria | |
| Anaplasmataceae | |
| Anaplasma phagocytophilum b,c | Canada: AB, MB, NB, NL, NS, ON, PE, QC, SK; US: CT, DE, GA, IL, IN, MA, MD, ME, MI, MN, NC, ND, NH, NJ, NY, PA, RI, SC, TN, VA, WI |
| Ehrlichia chaffeensis b | US: NY |
| Ehrlichia ewingii b | US: TN |
| Ehrlichia Panola Mountain sp.b | US: TN |
| Ehrlichia muris-like sp. (or E. sp. nr. muris)b | US: MN, WI |
| Bartonellaceae | |
| Bartonella henselae b | US: NJ, NY, PA |
| Unidentified/uncharacterized Bartonella spp. | US: MD, NJ, PA |
| Rickettsiaceae | |
| Candidatus Rickettsia cooleyi (or cooleyi-like)d | US: FL, GA, TN, TX |
| Candidatus Rickettsia IRS4 | US: MA |
| Candidatus Rickettsia amblyommii | US: TN |
| Rickettsia buchneri d | US: MN |
| Rickettsia massiliae/Rickettsia sp. Bar 29 | US: NC |
| Rickettsia parkeri b | US: LA |
| Rickettsia peacockii d | US: TX |
| Rickettsia endosymbiont of Ixodes scapularis d | US: CT, IN, LA, MA, ME, NY, PA, TN, WI |
| Rickettsia sp. Is-I | US: GA |
| Rickettsia sp. TR-39 | US: FL, GA |
| Unidentified/uncharacterized Rickettsia spp. | US: MD, NC, TN |
| Wolbachia sp. | US: CT |
| Spirochaetes | |
| Spirochaetaceae | |
| Borrelia andersonii b | US: MI |
| Borrelia bissettii b | US: LA |
| Borrelia burgdorferi s.s.b | Canada: AB, MB, NB, NS, NL, ON, PE, QC; US: AR, CT, DE, GA, IA, IL, IN, LA, MA, MD, ME, MI, MN, NC, ND, NH, NJ, NY, PA, RI, SC, TX, VA, WI |
| Borrelia kurtenbachii b | Canada: NS; US: NY |
| Borrelia lonestari b | US: AR, MA, NY |
| Borrelia miyamotoi b | Canada: AB, MB, NB, NS, ON, PE, QC; US: CT, IA, IL, IN, MA, MD, ME, MI, MN, NJ, NY, PA, RI, VA, WI |
| Unidentified/uncharacterized Borrelia sp. | US: NY |
| Piroplasmida | |
| Babesiidae | |
| Babesia microti b | US: CT, MA, MD, ME, MN, NH, NJ, NY, PA, WI |
| Babesia odocoilei | US: IN, IL, ME, MI, MS, PA, TN, WI |
| Theileriidae | |
| Theileria cervi | US: TN |
| Viruses | |
| Bunyaviridae | |
| Blacklegged tick phlebovirus | US: NY |
| South Bay virus | US: NY |
| Flaviviridae | |
| Deer tick virus (POWV lineage II)b | US: CT, MA, NY, RI, WI |
| Powassan virus (POWV lineage I)b | US: NY |
| POWV (not subtyped)b | US: NY |
| Reoviridae | |
| St. Croix River viruse | US: WI |
| Other | |
| Ixodes scapularis mononegaviralese | US: NY |
| Non-vector-borne, gut/surface orgnaisms | |
| Alphaproteobacteria | |
| Bradyrhizobiaceae | |
| Afipia broomeae f | US: NY |
| Afipia felis f | US: CT |
| Caulobacteraceae | |
| Uncultured bacterium 1 | US: CT |
| Uncultured bacterium 2 | US: CT |
| Methylobacteriaceae | |
| Methylobacterium mesophilicum f | US: CT |
| Methylobacterium sp. strain G296-5 | US: CT |
| Uncultured bacterium 3 | US: CT |
| Sphingomonadaceae | |
| Sphingomonas elodea f | US: CT |
| Sphingomonas melonis f | US: CT |
| Sphingomonas sp. | US: CT |
| Sphingomonas sp. strain BF2 | US: CT |
| Sphingomonas sp. AV069 | US: CT |
| Sphingopyxis alaskensis | US: CT |
| Sphingobacterium sp. | US: NY |
| Betaproteobacteria | |
| Burkholderiaceae | |
| Burkholderia sp. | US: CT |
| Ralstonia mannitolilytica f | US: NY |
| Comamonadaceae | |
| Delftia acidovorans f | US: CT |
| Delftia acidovorans WDL34f | US: CT |
| Uncultured bacterium 4 | US: CT |
| Uncultured bacterium 5 | US: CT |
| Gammaproteobacteria | |
| Enterobacteriaceae | |
| Enterobacter asburiae f | US: NY |
| Escherichia coli f | US: CT |
| Photorhabdus sp. | US: NY |
| Raoultella sp. | US: NY |
| Shigella sp. | US: NY |
| Uncultured gamma protebacterium | US: NY |
| Acinetobacter sp. strain phenon 4 | US: CT |
| Acinetobacter sp. 1 | US: CT |
| Acinetobacter sp. 2 | US: NY |
| Pseudomonadaceae | |
| Pseudomonas fluorescens f | US: NY |
| Pseudomonas sp. 1 | US: NY |
| Pseudomonas sp. 2 | US: NY |
| Pseudomonas sp. 3 | US: NY |
| Symbiont cf. Pseudomonas | US: NY |
| Xanthomonadaceae | |
| Stenotrophomonas maltophilia f | US: CT |
| Uncultured Stenotrophomonas sp. | US: NY |
| Stenotrophomonas sp. | US: NY |
| Actinobacteria | |
| Mycobacteriaceae | |
| Mycobacterium manitobense f | US: CT |
| Nocardiaceae | |
| Rhodococcus erythropolis strain NVIf | US: CT |
| Rhodococcus sp. | US: NY |
| Williamsia sp. | US: NY |
| Propionibacteriaceae | |
| Propionibacterium acnes f | US: CT |
| Firmicutes | |
| Bacillaceae | |
| Bacillus sp. | US: CT |
| Streptococcaceae | |
| Uncultured Streptococcus sp. | US: CT |
| Unclassified Bacteria | |
| Uncultured bacterium 6 | US: CT |
| Uncultured bacterium 7 | US: CT |
| Uncultured bacterium 8 | US: CT |
| Uncultured bacterium 9 | US: CT |
| Uncultured bacterium 10 | US: CT |
| Uncultured bacterium 11 | US: CT |
| Uncultured bacterium 12 | US: CT |
| Uncultured bacterium 13 | US: CT |
| Plastid clone | US: CT |
| Uncultured bacterium 14 | US: NY |
| Uncultured/unidentified bacterium | US: NY |
aCanada: AB, Alberta; MB, Manitoba; NB, New Brunswick; NL, Newfoundland; NS, Nova Scotia; ON, Ontario; PE, Prince Edward Island; SK, Saskatchewan; QC, Quebec. US: AR, Arkansas; CT, Connecticut; DE, Delaware; FL, Florida; GA, Georgia; IL, Illinois; IN, Indiana; IA, Iowa; LA, Louisiana; ME, Maine; MD, Maryland; MA, Massachusetts; MI, Michigan; MN, Minnesota; MS, Mississippi; NH, New Hampshire; NJ, New Jersey; NY, New York; NC, North Carolina; ND, North Dakota; PA, Pennsylvania; RI, Rhode Island; SC, South Carolina; TN, Tennessee; TX, Texas; VT, Vermont; VA, Virginia; WI, Wisconsin
bKnown or putative human pathogen, B. andersoni [131]; B. bissettii [132]; B. kurtenbachii [133]; B. lonestari [134]; R. parkeri [91]
cBoth the Ap-ha strain (human disease strain) and the Ap-variant-1 strain were pooled during data extraction, since not all studies distinguished between the two
dKnown or putative endosymbiont of Ix. scapularis
eLikely a viral genome integrated into I. scapularis genome [135, 136]
fHuman pathogen, but likely not transmitted by blacklegged ticks, gut organisms
Results
Study characteristics
Seventy-eight studies were included in our final review (Additional file 2) [4, 8, 15–90]. Eight studies included blacklegged ticks from Canada and 72 studies included samples from the US (two studies included Canadian and US blacklegged ticks). In Canada, Ontario (n = 6) was sampled the most frequently, followed by Nova Scotia (n = 5) (Fig. 2). In the US, New York (n = 18) was sampled the most frequently, followed by Wisconsin (n = 12), New Jersey (n = 10), Connecticut (n = 9), Massachusetts (n = 7), Pennsylvania (n = 7) and Rhode Island (n = 7) (Fig. 2). Most Canadian studies did not provide details on the sub-provincial regions where tick collection occurred; however, the areas sampled most frequently were southern regions of Ontario and Nova Scotia. In the US, counties that were sampled the most were Dutchess (New York) (n = 11), Westchester (New York) (n = 9), Monmouth (New Jersey) (n = 6) and New London (Connecticut) (n = 6). No studies included blacklegged ticks from Alabama, Kansas, Missouri, Ohio, Oklahoma, South Dakota, Vermont or West Virginia. From 2000 through 2007, three studies were published per year (range = 2–4); from 2009 through 2015, 6.8 studies were published per year (range = 1–19).
Quality assessment
Twenty studies (26 % or 20 out of 78) met 100 % of quality assessment criteria and an additional 44 studies (56 %) met 75 % of criteria (Additional file 3). Thirty-three studies (42 %) collected blacklegged ticks by dragging or flagging; 21 (27 %) collected ticks from mammal or bird hosts; 15 (19 %) used a combination of dragging and animal collections; and nine (12 %) did not report collection methods or employed other techniques. Thirty-three studies (42 %) explicitly reported positive and negative controls; four (5 %) reported negative controls only; 10 (13 %) reported positive controls only; and 31 (40 %) did not report any controls (Additional file 2). Six studies (38 % or six out of 16) included GenBank accession numbers for gene sequenced PCR products where species identification was indeterminate.
Descriptive analysis
Blacklegged ticks harbored 91 distinct taxa. Sixteen of these organisms are tick-transmitted human pathogens; the remainder is primarily gut or surface microbes of blacklegged ticks and not transmittable to humans or symbionts of blacklegged ticks (Table 1). The highest number of organisms detected in the Canadian-sampled blacklegged ticks were from Nova Scotia (n = 4; i.e. at least four separate identifications of the organisms sought in the ticks sampled), followed by three each in Alberta, Manitoba, New Brunswick, Ontario and Prince Edward Island (Table 2). US-sampled blacklegged ticks harbored 91 organisms. The highest number of organisms were detected in Connecticut (n = 42), followed by New York (n = 36), Wisconsin (n = 9), Tennessee (n = 9), Massachusetts (n = 8) and Pennsylvania (n = 8). Studies in different jurisdictions sought varying numbers of pathogens and often used various detection methods. The mean ± SE number of studies per jurisdiction (4.2 ± 0.60; median = 3; n = 78) was positively correlated (r = 0.77, n = 37, P < 0.0001) with the mean ± SE number of organisms detected per jurisdiction (5.2 ± 1.09; median = 4; n = 78). Similarly, the number of blacklegged ticks tested in a jurisdiction (mean = 2,797 ± 1,022.6; median = 696) was positively correlated with the number of organisms detected per jurisdiction (r = 0.72, n = 37, P < 0.0001). In the studies reviewed, researchers performed 149 unique sampling events, where a sampling event is the testing of blacklegged ticks for a specific organism in a specific jurisdiction. The number of organisms sought per sampling event (mean = 2.8 ± 0.29; median = 2; n = 149) was positively correlated (r = 0.95, n = 149, P < 0.0001) with the number of organisms detected per sampling event (mean = 2.3 ± 0.28; median = 2; n = 149). We performed multiple tests on the same dataset, however, since these P-values are very small, adjusting for multiple testing would not alter the statistical significance of the results.
Thirty-two distinct organisms transmitted by ticks were reported in the studies reviewed (Table 1). Five species of Anaplasmataceae were detected; A. phagocytophilum was most prevalent in Rhode Island (21.3 %; n = 684), Georgia (17.3 %; n = 910), Connecticut (13.0 %; n = 454), Indiana (10.8; n = 712), New York (8.9 %; n = 25,098), Michigan (7.9 %; n = 444), Pennsylvania (6.5 %; n = 1,559) and Illinois (6.5 %; n = 278) (Fig. 3; Table 1). Ehrlichia muris-like sp. was restricted to Wisconsin (5.4 %; n = 4,066) and Minnesota (3.0 %; n = 534) (Fig. 3). Blacklegged ticks harbored one Bartonellaceae species, Bartonella henselae, which was only found in ticks from New Jersey (100 %; n = 1), New York (2.3 %; n = 88) and Pennsylvania (3.1 %; n = 544). Blacklegged ticks contained 12 Rickettsia taxa; Rickettsia endosymbiont of Ixodes scapularis, a blacklegged tick obligate endosymbiont (non-pathogenic), was the most prevalent taxa, particularly in Pennsylvania (64.9 %; n = 94), Indiana (63.0 %; n = 100), Tennessee (51.1 %; n = 47) and Maine (46.0 %; n = 100). Seven out of 12 Rickettsia species were restricted geographically to the Southern US and the known human pathogen Rickettsia parkeri was limited to Louisiana (16.6 %; n = 18). Ticks sampled across all 78 studies contained six species of Borrelia. Borrelia burgdorferi was most prevalent in New Hampshire (52.3 %; n = 509), Maine (49.1 %; n = 10,004), Minnesota (47.2 %; n = 803), Indiana (45.3 % n = 506), Michigan (39.5 %; n = 696), Illinois (33.9 %; n = 460) and New York (31.6 %; n = 21,363). Borrelia miyamotoi was most prevalent in Connecticut (4.7 %; n = 1,226), Indiana (4.1 %; n = 487), Virginia (3.5 %; n = 173), Minnesota (2.9 %; n =700), New Jersey (2.7 %; n = 765), Massachusetts (2.5 %; n = 159) and Wisconsin (2.4 %; n = 3,151) (Fig. 3). Blacklegged ticks harbored three Apicomplexan parasites; Ba. microti was most prevalent in Connecticut (6.7 %; n = 1,198), New Jersey (6.5 %; n = 1.195) and Massachusetts (5.3 %; n = 851) (Fig. 3). Several viruses were detected in blacklegged ticks; however, only DTV and Powassan virus (POWV) are of concern as human pathogens. Deer tick virus/POWV was most common in New York (3.4 %; n = 91) (Fig. 3). Pathogens sought for but not detected in any samples included Coxiella burnetii (New York, n = 88), Francisella tularensis (New York, n = 374; Florida, n = 52; Georgia, n = 13) and Rickettsia rickettsii (New York, n = 88) (Additional file 2).
Two of the studies reported 55 additional bacteria taxa, common genera were Acinetobacter (Enterobacteriaceae), Afipia (Alphaproteobacteria), Pseudomonas (Pseudomonadaceae), Sphingomonas (Sphingomonadaceae) and Stenotrophomonas (Xanthomonadaceae) [19, 64]. While most of the bacteria identified are commonly found in the environment (i.e., soil, water), some are associated with mammals, such as E. coli, Shigella sp. and Streptococcus sp. These additional bacteria are likely a part of the microbial community (as commensal organisms or environmental contaminants) of the blacklegged tick’s gut or surface and not necessarily transmitted or maintained by blacklegged ticks.
The mandatory reporting requirements of pathogens varied among the jurisdictions reviewed. Deer tick virus is typically reportable as an arboviral infection in most of the jurisdictions reviewed except for Manitoba (Table 2). In Canada, anaplasmosis is reportable only in Manitoba, yet reportable in all US states reviewed. In Canada, babesiosis is reportable only in Manitoba, but in all US states reviewed except Pennsylvania. Bartonellosis was not reportable in any Canadian jurisdiction and reportable only in Michigan and Minnesota in the US. Lyme disease (as B. burgdorferi) is reportable in all jurisdictions reviewed; however, infection by other Borrelia species is not. In Canada, only Quebec requires reporting of ehrlichiosis, but ehrlichiosis is reportable in all US states reviewed. In Canada, rickettsial infections are not reportable, but are reportable in all US states reviewed.
Discussion
Our systematic review of 78 North American studies published since 2000 documented that blacklegged ticks are associated with 91 distinct organisms. Sixteen of these organisms are tick-transmitted human pathogens. The remainder of the organisms are non-pathogenic gut or surface biota (as commensal organisms or environmental contaminants), intracellular symbionts of ticks or have unknown pathologies in vertebrates. Blacklegged ticks are the principal vectors of six of these human pathogens: A. phagocytophilum, Ba. microti, B. burgdorferi, B. miyamotoi, DTV and Ehrlichia muris-like sp. Due to studies documenting their proximity to, or detection in, Ontario, the six pathogens represent the primary risks in the province, with two presenting more immediate threats than others due to their recent detection in parts of Ontario (i.e. A. phagocytophilum, B. burgdorferi). Similar to our review, 20 human pathogens were reported from Europe’s sister taxa to the blacklegged tick, Ixodes ricinus [91, 92]. Given the blacklegged tick’s liberal feeding behavior and propensity to bite humans, continued identification of blacklegged tick-pathogen relationships and assessing their public health implications is justified.
Blacklegged ticks transmit two of the Anaplasmataceae bacteria reported in the review (i.e. A. phagocytophilum and Ehrlichia muris-like sp.). The number of human granulocytic anaplasmosis cases has increased in the US from less than 200 cases in 1997 to over 2,700 cases in 2013, with most cases reported from Massachusetts, Minnesota, New York and Wisconsin [93, 94]. Patients infected with A. phagocytophilum display (in order of decreasing incidence) malaise, fever, myalgia, headache, arthralgia and nausea [95]. Anaplasma phagocytophilum is relatively rare (< 0.5 %) in Ontario blacklegged ticks and we are not aware of any anaplasmosis cases diagnosed from the province [4]. Ehrlichia muris-like sp. is restricted to Wisconsin and Minnesota; however, this pathogen has not been widely tested for elsewhere. Little is known about this pathogen; nevertheless, it has been identified in 73 patients from Indiana, Michigan, Minnesota, North Dakota and Wisconsin (all cases reported tick-bite exposures from Minnesota or Wisconsin) displaying fever, malaise, headache and myalgia [96]. Considering A. phagocytophilum is present in Ontario and the distribution of Ehrlichia muris-like sp. is not well-characterized, awareness of these pathogens by public health officials is warranted.
We expected a relatively high number of rickettsial agents to be reported from blacklegged ticks, given the extensive association of Rickettsia with hematophagous invertebrates such as fleas (Siphonaptera) and other ticks (Ixodidae, Argasidae) [91]. Human disease has not been associated with the majority of rickettsial organisms identified in this review; however, careful clinical and epidemiological studies can lead to some of these being identified as human pathogens (e.g. R. parkeri) [97]. Currently, there is no evidence demonstrating blacklegged ticks as effective vectors of Rickettsia; however, further investigation is justified where unexplained cases of spotted fever occur with tick-bite histories.
The reviewed studies reported that blacklegged ticks harbor six species of Borrelia, two of which are primarily reported from blacklegged ticks (i.e. B. burgdorferi and B. miyamotoi). Borrelia burgdorferi was widely distributed throughout the range of the blacklegged tick (at least in the ticks examined in these studies), including Ontario (prevalence = 16 % of blacklegged ticks), with the highest prevalence (> 30 %) in the Northeast and Midwest US. Studies focusing solely on B. burgdorferi were not included in review; therefore, caution must be used when interpreting the prevalence of B. burgdorferi. Patients with acute B. burgdorferi infection usually display erythema migrans (bulls-eye rash), fever, myalgia, headache, arthralgia, neck stiffness and arthritis [98]. In Ontario, the Lyme disease incidence rate has increased from 0.2 (2002) to 1.6 cases per 100,000 population (2014) [99]. Borrelia miyamotoi is a newly recognized human pathogen, which based on the reviewed studies has a widespread distribution in North America (Ontario prevalence < 0.5 %). Recently, 53 patients with Lyme disease-like symptoms from Connecticut, Massachusetts, New Jersey, New York and Rhode Island showed serological evidence for B. miyamotoi infection [100, 101]. Patients with B. miyamotoi disease display fever, fatigue, headache, myalgia, chills and nausea [10, 102]. To our knowledge, B. miyamotoi disease has not been diagnosed in Ontario. Since our literature search was performed, a new blacklegged tick-transmitted Borrelia has been associated with human disease in Wisconsin, Candidatus Borrelia mayonii [103, 104]. Borrelia burgdorferi has rightfully been the Borrelia of concern to public health; however, other Borrelia species are beginning to emerge as additional threats to public health.
While the focus of this systematic review was organisms of public health significance, it is important to note the superficially benign microbes and symbionts associated with blacklegged ticks. While outside the scope of this review, the gut microorganisms of blacklegged ticks can moderate the colonization of human pathogens in blacklegged ticks, such as with B. burgdorferi [105, 106]. Several of the gut or surface bacteria (e.g. Stenotrophomonas maltophilia and Rhodococcus erythropolis) associated with opportunistic infections in humans, especially in immunocompromised individuals [107–109]. Our understanding of the role that symbionts play in the biology of blacklegged ticks and the blacklegged tick’s ability to transmit pathogens is poorly understood and represents an opportunity for future research. Symbionts, such as Rickettsia endosymbiont of Ixodes scapularis and Wolbachia sp. (closely related to W. pipientis supergroup A) were reported from the studies reviewed. Wolbachia symbionts are involved in the manipulation of arthropod reproduction (e.g. cytoplasmic incompatibility and parthenogenesis) in other arthropods such as Drosophila and parasitoid wasps [110]. Understanding how the entire tick microbiome regulates pathogen acquisition and transmission is a burgeoning field, undoubtedly providing insights into potential blacklegged tick management options. In addition, a better understanding of Ix. scapularis symbionts could lead to novel management tools.
In the reviewed studies, Ba. microti was most common in blacklegged ticks collected from the Coastal Atlantic states of the Northeast US. In the US, the number of babesiosis cases has increased from approximately 1,100 (2011) to 1,800 (2013), with most cases reported from Connecticut, Massachusetts, New Jersey and New York [94, 111]. Patients with babesiosis display fever, fatigue, headache, chills and arthralgia [112]. While Ba. microti could eventually occupy the same distribution of B. burgdorferi, it will do so at a slower rate due to several factors. For example, blacklegged tick transmission of Ba. microti to reservoir hosts is not as efficient when compared to B. burgdorferi [113, 114]. In addition, the survival of Ba. microti is low in overwintering blacklegged tick nymphs when compared to B. burgdorferi [115]. Babesia microti has not been detected in Ontario blacklegged ticks (in the published literature) or identified as the cause of locally acquired disease, to our knowledge.
Deer tick virus (POWV lineage II) was the most common viral agent detected in the studies reviewed, a viral species serologically indistinguishable from POWV (POWV lineage I) but with unique nucleotide and amino acid sequences. In 2009, DTV was the cause of a fatal case of encephalitis from New York, the first report implicating DTV as an agent of human disease [116]. Soon after this initial case report, additional cases were reported in New York that displayed fever, malaise, confusion, seizure, headache, rash and vomiting [117]. There is no evidence for the presence of DTV in the Ontario’s blacklegged ticks; however, field research is underway to determine its presence (Curtis B Russell, unpublished data).
Describing pathogens associated with blacklegged ticks is only the first step towards appreciating the role of this tick as North America’s most important vector of public health significance. The majority of pathogens and intracellular symbionts reported here have poorly understood natural histories and, for the most part, we know little of their maintenance in nature, their potential or preferred vertebrate reservoirs and, in some cases, their ability to cause disease in vertebrates. While symbionts and non-human pathogens are relatively benign, it is important to understand their ecology as well, because they likely play roles in determining the distribution and prevalence of the human pathogens discussed through competitive exclusion [118]. In Ontario blacklegged ticks, only a handful of pathogens have been sought after and there is an opportunity to better understand their ecology (e.g. Babesia, Ehrlichia, Rickettsia, DTV). Understanding the complex epidemiology of the human pathogens transmittable by blacklegged ticks will require longitudinal, ecological (host and vector competence) and epidemiological studies in endemic and emerging areas.
While comprehensive, our systematic review does have several limitations. We did not perform a search of the grey literature (publications on testing results from public health or government institutions); therefore, our results might be biased towards primary research with positive results due to publication bias. In Canada, blacklegged ticks collected by passive surveillance were likely used in multiple studies for pathogen detection; however, an examination of the studies shows that overlap (same ticks used in separate studies) involved only two studies each from 2007 through 2012 and was limited to the detection of A. phagocytophilum and B. burgdorferi. Another limitation is that studies were undertaken where pathogen prevalence or disease burden are high, meaning prevalence does not represent a uniform value across a province or state. The heterogeneity among studies in terms of collection methods, blacklegged tick stages assayed, engorgement levels of ticks, molecular methods used or gene targets undoubtedly influenced the crude prevalences we reported. For example, we expect that ticks collected from reservoir hosts (pathogen acquisition can also vary by host type and tick stage) would have a higher pathogen load, compared to host-seeking ticks collected by dragging. The number of organisms detected in a jurisdiction is dependent upon sampling effort and testing methods; therefore, little-studied areas may indeed hold more blacklegged tick- organisms. The presence of pathogens in neighboring jurisdictions or in Ontario itself provides the basis for assessing the risks of blacklegged tick-transmitted infections in Ontario, but this method does not allow for an accurate estimate of when these pathogens will be a threat to Ontario. As far as we know, blacklegged ticks transmit and maintain all the underscored pathogens within a rodent-mammal-tick cycle in temperate, mixed deciduous forests. Given the pathogens share a common transmission cycle, these pathogens should eventually occur throughout the blacklegged tick’s range. The review identified that most research has occurred in relatively small geographic areas, representing an opportunity to determine pathogen incidence and prevalence outside of highly sampled regions. As research continues to detect pathogens in blacklegged ticks and reservoir hosts, we expect the number of pathogens to increase. In addition, comprehensive studies, seeking all possible pathogens using standardized methods, are needed for better comparison of pathogen prevalence across northeastern North America. Our review is the first systematic assessment of literature, identifying and bringing together the scattered knowledge of human pathogens associated with the blacklegged tick.
Conclusions
Viewed as a mere pest until the mid-1970s, the blacklegged tick was not known to transmit pathogens to humans or other animals [119]. In the following 40 years, the blacklegged tick has turned into the most important vector in North America. Our review has identified several human pathogens besides B. burgdorferi that are transmittable by blacklegged ticks, yet we have not demonstrated disease in Ontario at this time for A. phagocytophilum, Ba. microti, B. miyamotoi, DTV or Ehrlichia muris-like sp. While most of these pathogens do not represent immediate public health threats in Ontario, there is an opportunity to ensure mitigation efforts are in place prior to their arrival. The growing public and physician awareness concerning Lyme disease provides a catalyst upon which to synergize awareness of other tick-borne diseases. In addition, surveillance of pathogens in field-collected blacklegged ticks will establish baseline data and inform local risk assessments. Public health in Ontario needs to remain vigilant because Ontario (i) has a relatively higher population of at-risk people in Canada, (ii) has blacklegged tick populations that continue to expand and (iii) is positioned next to jurisdictions where these pathogens occur.
Acknowledgements
We thank PHO’s Library Services (Susan Massarella, Beata Pach) for developing search strategies. We thank Rajitha Liyanage and Lauren Ramsey (PHO) for bibliographical support. Additionally, we thank three anonymous reviewers that provided valuable comments to improve the manuscript.
Abbreviations
- ESRI
Environmental Systems Research Institute
- DTV
deer tick virus
- PCR
polymerase chain reaction
- POWV
Powassan encephalitis virus
- PRISMA
Preferred Reporting Items for Systematic Reviews
- NR
not reportable
- RMSF
Rocky Mountain spotted fever
- STROBE
Strengthening the Reporting of OBservational Studies in Epidemiology
Additional files
Search strategy example using Ovid MEDLINE(R) In-Process & Other Non-Indexed Citations and Ovid MEDLINE(R) 1 January 1995 to 20 April 2015. (DOCX 26 kb)
Summary of organisms detected in Ixodes scapularis collected throughout eastern North America, from 78 studies reviewed (2000–2015). (DOCX 117 kb)
Critical appraisal of 78 studies in review. (DOCX 61 kb)
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
MPN designed and conceived the systematic review, screened titles and abstracts against eligibility criteria, performed critical appraisals, extracted data from studies and wrote the manuscript. CBR performed critical appraisals of eligible primary studies and helped write the manuscript. NJS screened titles and abstracts against eligibility criteria. YL performed statistical analyses. SJ prepared the maps. BS, SM, SNP, LF and DS participated in the study design and helped draft the manuscript. All authors read and approved the final manuscript.
References
- 1.Ogden NH, St-Onge L, Barker IK, Brazeau S, Bigras-Poulin M, Charron DF, et al. Risk maps for range expansion of the Lyme disease vector, Ixodes scapularis, in Canada now and with climate change. Int J Health Geogr. 2008, doi: 10.1186/1476-072X-7-24. [DOI] [PMC free article] [PubMed]
- 2.Spielman A. The emergence of Lyme disease and human babesiosis in a changing environment. Ann N Y Acad Sci. 1994;740:146–56. doi: 10.1111/j.1749-6632.1994.tb19865.x. [DOI] [PubMed] [Google Scholar]
- 3.Leighton PA, Koffi JK, Pelcat Y, Lindsay LR, Ogden NH. Predicting the speed of tick invasion: an empirical model of range expansion for the Lyme disease vector Ixodes scapularis in Canada. J Appl Ecol. 2012;49:457–64. doi: 10.1111/j.1365-2664.2012.02112.x. [DOI] [Google Scholar]
- 4.Nelder MP, Russell C, Lindsay LR, Dhar B, Patel SN, Johnson S, et al. Population-based passive tick surveillance and detection of expanding foci of blacklegged ticks Ixodes scapularis and the Lyme disease agent Borrelia burgdorferi in Ontario, Canada. PLoS One. 2014;9:e105358. [DOI] [PMC free article] [PubMed]
- 5.Pancholi P, Kolbert CP, Mitchell PD, Reed KD, Dumler JS, Bakken JS, et al. Ixodes dammini as a potential vector of human granulocytic ehrlichiosis. J Infect Dis. 1995;172:1007–12. doi: 10.1093/infdis/172.4.1007. [DOI] [PubMed] [Google Scholar]
- 6.Burgdorfer W, Barbour AG, Hayes SF, Benach JL, Grunwaldt E, Davis JP. Lyme disease-a tick-borne spirochetosis? Science. 1982;216:1317–9. doi: 10.1126/science.7043737. [DOI] [PubMed] [Google Scholar]
- 7.Steere AC, Grodzicki RL, Kornblatt AN, Craft JE, Barbour AG, Burgdorfer W, et al. The spirochetal etiology of Lyme disease. N Engl J Med. 1983;308:733–40. doi: 10.1056/NEJM198303313081301. [DOI] [PubMed] [Google Scholar]
- 8.Dupuis AP, Peters RJ, Prusinski MA, Falco RC, Ostfeld RS, Kramer LD. Isolation of deer tick virus (Powassan virus, lineage II) from Ixodes scapularis and detection of antibody in vertebrate hosts sampled in the Hudson Valley, New York State. Parasit Vectors. 2013, doi: 10.1186/1756-3305-6-185. [DOI] [PMC free article] [PubMed]
- 9.Telford SR, Dawson JE, Katavolos P, Warner CK, Kolbert CP, Persing DH. Perpetuation of the agent of human granulocytic ehrlichiosis in a deer tick-rodent cycle. Proc Natl Acad Sci U S A. 1996;93:6209–14. doi: 10.1073/pnas.93.12.6209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Molloy PJ, Telford SR, Chowdri HR, Lepore TJ, Gugliotta JL, Weeks KE, et al. Borrelia miyamotoi disease in the Northeastern United States: a case series. Ann Intern Med. 2015;163:91–8. doi: 10.7326/M15-0333. [DOI] [PubMed] [Google Scholar]
- 11.Robinson S. Infectious disease epidemiology report: tick-borne diseases, Maine - 2013. 2014. [Google Scholar]
- 12.Liberati A, Altman DG, Tetzlaff J, Mulrow C, Gøtzsche PC, Ioannidis JP, et al. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. Ann Intern Med. 2009;339:b2700. doi: 10.7326/0003-4819-151-4-200908180-00136. [DOI] [PubMed] [Google Scholar]
- 13.Stromdahl EY, Evans SR, O'Brien JJ, Gutierrez AG. Prevalence of infection in ticks submitted to the human tick test kit program of the U.S. Army Center for Health Promotion and Preventive Medicine. J Med Entomol. 2001;38:67–74. doi: 10.1603/0022-2585-38.1.67. [DOI] [PubMed] [Google Scholar]
- 14.Ontario Agency for Health Protection and Promotion (Public Health Ontario) Meta-tool for quality appraisal of public health evidence: PHO MetaQAT. Toronto: Queen’s Printer for Ontario; 2015. [Google Scholar]
- 15.Swanson KI, Norris DE. Co-circulating microorganisms in questing Ixodes scapularis nymphs in Maryland. J Vector Ecol. 2007;32:243–51. doi: 10.3376/1081-1710(2007)32[243:CMIQIS]2.0.CO;2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Aliota MT, Dupuis AP, Wilczek MP, Peters RJ, Ostfeld RS, Kramer LD. The prevalence of zoonotic tick-borne pathogens in Ixodes scapularis collected in the Hudson Valley, New York State. Vector Borne Zoonotic Dis. 2014;14:245–50. doi: 10.1089/vbz.2013.1475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Anderson JF, Armstrong PM. Prevalence and genetic characterization of Powassan virus strains infecting Ixodes scapularis in Connecticut. Am J Trop Med Hyg. 2012;87:754–9. doi: 10.4269/ajtmh.2012.12-0294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Barbour AG, Bunikis J, Travinsky B, Hoen AG, Diuk-Wasser MA, Fish D, et al. Niche partitioning of Borrelia burgdorferi and Borrelia miyamotoi in the same tick vector and mammalian reservoir species. Am J Trop Med Hyg. 2009;81:1120–31. doi: 10.4269/ajtmh.2009.09-0208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Benson MJ, Gawronski JD, Eveleigh DE, Benson DR. Intracellular symbionts and other bacteria associated with deer ticks (Ixodes scapularis) from Nantucket and Wellfleet, Cape Cod, Massachusetts. Appl Environ Microbiol. 2004;70:616–20. doi: 10.1128/AEM.70.1.616-620.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bouchard C, Leighton PA, Beauchamp G, Nguon S, Trudel L, Milord F, et al. Harvested white-tailed deer as sentinel hosts for early establishing Ixodes scapularis populations and risk from vector-borne zoonoses in southeastern Canada. J Med Entomol. 2013;50:384–93. doi: 10.1603/ME12093. [DOI] [PubMed] [Google Scholar]
- 21.Brackney DE, Nofchissey RA, Fitzpatrick KA, Brown IK, Ebel GD. Stable prevalence of Powassan virus in Ixodes scapularis in a northern Wisconsin focus. Am J Trop Med Hyg. 2008;79:971–3. [PMC free article] [PubMed] [Google Scholar]
- 22.Cherepko J, Berry GJ, Keeler SP, Huffman JE. Prevalence of Borrelia burgdorferi, Bartonella spp., Bartonella henselae, Babesia microti and Anaplasma phagocytophila in Ixodes scapularis ticks collected in Monroe County, Pennsylvania, show a risk for co- and tri-infections. J Pa Acad Sci. 2010;84:74–8. [Google Scholar]
- 23.Cohen SB, Yabsley MJ, Freye JD, Dunlap BG, Rowland ME, Huang J, et al. Prevalence of Ehrlichia chaffeensis and Ehrlichia ewingii in ticks from Tennessee. Vector Borne Zoonotic Dis. 2010;10:435–40. doi: 10.1089/vbz.2009.0058. [DOI] [PubMed] [Google Scholar]
- 24.Courtney JW, Dryden RL, Montgomery J, Schneider BS, Smith G, Massung RF. Molecular characterization of Anaplasma phagocytophilum and Borrelia burgdorferi in Ixodes scapularis ticks from Pennsylvania. J Clin Microbiol. 2003;41:1569–73. doi: 10.1128/JCM.41.4.1569-1573.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Crowder CD, Carolan HE, Rounds MA, Honig V, Mothes B, Haag H, et al. Prevalence of Borrelia miyamotoi in Ixodes ticks in Europe and the United States. Emerg Infect Dis. 2014;20:1678–82. doi: 10.3201/eid2010.131583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Curran KL, Kidd JB, Vassallo J, Van Meter VL. Borrelia burgdorferi and the causative agent of human granulocytic ehrlichiosis in deer ticks. Delaware Emerg Infect Dis. 2000;6:408–11. doi: 10.3201/eid0604.000417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Dibernardo A, Cote T, Ogden NH, Lindsay LR. The prevalence of Borrelia miyamotoi infection, and co-infections with other Borrelia spp. in Ixodes scapularis ticks collected in Canada. Parasit Vectors. 2014;doi: 10.1186/1756-3305-7-183. [DOI] [PMC free article] [PubMed]
- 28.Diuk-Wasser MA, Liu Y, Steeves TK, Folsom-O'Keefe C, Dardick KR, Lepore T, et al. Monitoring human babesiosis emergence through vector surveillance New England. USA Emerg Infect Dis. 2014;20:225–31. doi: 10.3201/eid2002.130644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Drebot MA, Lindsay R, Barker IK, Artsob H. Characterization of a human granulocytic ehrlichiosis-like agent from Ixodes scapularis, Ontario. Canada Emerg Infect Dis. 2001;7:479–80. doi: 10.3201/eid0703.017327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ebel GD, Campbell EN, Goethert HK, Spielman A, Telford SR. Enzootic transmission of deer tick virus in New England and Wisconsin sites. Am J Trop Med Hyg. 2000;63:36–42. doi: 10.4269/ajtmh.2000.63.36. [DOI] [PubMed] [Google Scholar]
- 31.Eskow E, Rao RV, Mordechai E. Concurrent infection of the central nervous system by Borrelia burgdorferi and Bartonella henselae: evidence for a novel tick-borne disease complex. Arch Neurol. 2001;58:1357–63. doi: 10.1001/archneur.58.9.1357. [DOI] [PubMed] [Google Scholar]
- 32.Fang QQ, Mixson TR, Hughes M, Dunham B, Sapp J. Prevalence of the agent of human granulocytic ehrlichiosis in Ixodes scapularis (Acari: Ixodidae) in the coastal southeastern United States. J Med Entomol. 2002;39:251–5. doi: 10.1603/0022-2585-39.2.251. [DOI] [PubMed] [Google Scholar]
- 33.Fritzen C, Mosites E, Applegate RD, Telford SR, Huang J, Yabsley MJ, et al. Environmental investigation following the first human case of babesiosis in Tennessee. J Parasitol. 2014;100:106–9. doi: 10.1645/12-158.1. [DOI] [PubMed] [Google Scholar]
- 34.Fryxell RT, Steelman CD, Szalanski AL, Kvamme KL, Billingsley PM, Williamson PC. Survey of Borreliae in ticks, canines, and white-tailed deer from Arkansas, U.S.A. Parasit Vectors. 2012; 5:doi: 10.1186/1756-3305-5-139. [DOI] [PMC free article] [PubMed]
- 35.Goltz L, Varela-Stokes A, Goddard J. Survey of adult Ixodes scapularis Say for disease agents in Mississippi. J Vector Ecol. 2013;38:401–3. doi: 10.1111/j.1948-7134.2013.12056.x. [DOI] [PubMed] [Google Scholar]
- 36.Grant-Klein RJ, Baldwin CD, Turell MJ, Rossi CA, Li F, Lovari R, et al. Rapid identification of vector-borne flaviviruses by mass spectrometry. Mol Cell Probes. 2010;24:219–28. doi: 10.1016/j.mcp.2010.04.003. [DOI] [PubMed] [Google Scholar]
- 37.Hamer SA, Roy PL, Hickling GJ, Walker ED, Foster ES, Barber CC, et al. Zoonotic pathogens in Ixodes scapularis, Michigan. Emerg Infect Dis. 2007;13:1131–3. doi: 10.3201/eid1307.070046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hamer SA, Tsao JI, Walker ED, Mansfield LS, Foster ES, Hickling GJ. Use of tick surveys and serosurveys to evaluate pet dogs as a sentinel species for emerging Lyme disease. Am J Vet Res. 2009;70:49–56. doi: 10.2460/ajvr.70.1.49. [DOI] [PubMed] [Google Scholar]
- 39.Hamer SA, Hickling GJ, Keith R, Sidge JL, Walker ED, Tsao JI. Associations of passerine birds, rabbits, and ticks with Borrelia miyamotoi and Borrelia andersonii in Michigan, U.S.A. Parasit Vectors. 2012;5:doi: 10.1186/1756-3305-5-231. [DOI] [PMC free article] [PubMed]
- 40.Hamer SA, Goldberg TL, Kitron UD, Brawn JD, Anderson TK, Loss SR, et al. Wild birds and urban ecology of ticks and tick-borne pathogens, Chicago, Illinois, USA, 2005-2010. Emerg Infect Dis. 2012;18:1589–95. doi: 10.3201/eid1810.120511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Hamer SA, Lehrer E, Magle SB. Wild birds as sentinels for multiple zoonotic pathogens along an urban to rural gradient in greater Chicago, Illinois. Zoonoses Public Health. 2012;59:355–64. doi: 10.1111/j.1863-2378.2012.01462.x. [DOI] [PubMed] [Google Scholar]
- 42.Hamer SA, Hickling GJ, Walker ED, Tsao JI. Increased diversity of zoonotic pathogens and Borrelia burgdorferi strains in established versus incipient Ixodes scapularis populations across the Midwestern United States. Infect Genet Evol. 2014;27:531–42. doi: 10.1016/j.meegid.2014.06.003. [DOI] [PubMed] [Google Scholar]
- 43.Han GS, Stromdahl EY, Wong D, Weltman AC. Exposure to Borrelia burgdorferi and other tick-borne pathogens in Gettysburg National Military Park, South-Central Pennsylvania, 2009. Vector Borne Zoonotic Dis. 2014;14:227–33. doi: 10.1089/vbz.2013.1363. [DOI] [PubMed] [Google Scholar]
- 44.Herrin BH, Zajac AM, Little SE. Confirmation of Borrelia burgdorferi sensu stricto and Anaplasma phagocytophilum in Ixodes scapularis. Southwestern Virginia Vector Borne Zoonotic Dis. 2014;14:821–3. doi: 10.1089/vbz.2014.1661. [DOI] [PubMed] [Google Scholar]
- 45.Hersh MH, Tibbetts M, Strauss M, Ostfeld RS, Keesing F. Reservoir competence of wildlife host species for Babesia microti. Emerg Infect Dis. 2012;18:1951–7. doi: 10.3201/eid1812.111392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hersh MH, Ostfeld RS, McHenry DJ, Tibbetts M, Brunner JL, Killilea ME, et al. Co-infection of blacklegged ticks with Babesia microti and Borrelia burgdorferi is higher than expected and acquired from small mammal hosts. PLoS One. 2014;9:e99348. doi: 10.1371/journal.pone.0099348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hoen AG, Rollend LG, Papero MA, Carroll JF, Daniels TJ, Mather TN, et al. Effects of tick control by acaricide self-treatment of white-tailed deer on host-seeking tick infection prevalence and entomologic risk for Ixodes scapularis-borne pathogens. Vector Borne Zoonotic Dis. 2009;9:431–8. doi: 10.1089/vbz.2008.0155. [DOI] [PubMed] [Google Scholar]
- 48.Holman MS, Caporale DA, Goldberg J, Lacombe E, Lubelczyk C, Rand PW, et al. Anaplasma phagocytophilum, Babesia microti, and Borrelia burgdorferi in Ixodes scapularis, southern coastal Maine. Emerg Infect Dis. 2004;10:744–6. doi: 10.3201/eid1004.030566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Keesing F, McHenry DJ, Hersh M, Tibbetts M, Brunner JL, Killilea M, et al. Prevalence of human-active and variant 1 strains of the tick-borne pathogen Anaplasma phagocytophilum in hosts and forests of eastern North America. Am J Trop Med Hyg. 2014;91:302–9. doi: 10.4269/ajtmh.13-0525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kogut SJ, Thill CD, Prusinski MA, Lee JH, Backerson PB, Coleman JL, et al. Babesia microti, upstate New York. Emerg Infect Dis. 2005;11:476–8. doi: 10.3201/eid1103.040599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Krakowetz CN, Dibernardo A, Lindsay LR, Chilton NB. Two Anaplasma phagocytophilum strains in Ixodes scapularis ticks, Canada. Emerg Infect Dis. 2014;20:2064–7. doi: 10.3201/eid2012.140172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Kurtti TJ, Felsheim RF, Burkhardt NY, Oliver JD, Heu CC, Munderloh UG. Rickettsia buchneri sp. nov., a rickettsial endosymbiont of the blacklegged tick Ixodes scapularis. Int J Syst Evol Microbiol. 2015;65:965–70. doi: 10.1099/ijs.0.000047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Layfield D, Guilfoile P. The prevalence of Borrelia burgdorfieri (Spirochaetales: spirochaetaceae) and the agent of human granulocytic ehrlichiosis (Rickettsiaceae: Ehrlichieae) in Ixodes scapularis (Acari: Ixodidae) collected during 1998 and 1999 from Minnesota. J Med Entomol. 2002;39:218–20. doi: 10.1603/0022-2585-39.1.218. [DOI] [PubMed] [Google Scholar]
- 54.Lee X, Coyle DR, Johnson DK, Murphy MW, McGeehin MA, Murphy RJ, et al. Prevalence of Borrelia burgdorferi and Anaplasma phagocytophilum in Ixodes scapularis (Acari: Ixodidae) nymphs collected in managed red pine forests in Wisconsin. J Med Entomol. 2014;51:694–701. doi: 10.1603/ME13140. [DOI] [PubMed] [Google Scholar]
- 55.Leydet BF, Liang FT. Detection of Lyme Borrelia in questing Ixodes scapularis (Acari: Ixodidae) and small mammals in Louisiana. J Med Entomol. 2014;51:278–82. doi: 10.1603/ME12273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lovrich SD, Jobe DA, Kowalski TJ, Policepatil SM, Callister SM. Expansion of the Midwestern focus for human granulocytic anaplasmosis into the region surrounding La Crosse, Wisconsin. J Clin Microbiol. 2011;49:3855–9. doi: 10.1128/JCM.05025-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Margos G, Hojgaard A, Lane RS, Cornet M, Fingerle V, Rudenko N, et al. Multilocus sequence analysis of Borrelia bissettii strains from North America reveals a new Borrelia species, Borrelia kurtenbachii. Ticks Tick Borne Dis. 2010;1:151–8. [DOI] [PMC free article] [PubMed]
- 58.Massung RF, Mauel MJ, Owens JH, Allan N, Courtney JW, Stafford KC, et al. Genetic variants of Ehrlichia phagocytophila, Rhode Island and Connecticut. Emerg Infect Dis. 2002;8:467–72. doi: 10.3201/eid0805.010251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Massung RF, Mather TN, Priestley RA, Levin ML. Transmission efficiency of the AP-variant 1 strain of Anaplasma phagocytophila. Ann N Y Acad Sci. 2003;990:75–9. doi: 10.1111/j.1749-6632.2003.tb07340.x. [DOI] [PubMed] [Google Scholar]
- 60.Mays SE, Hendricks BM, Paulsen DJ, Houston AE, Trout Fryxell RT. Prevalence of five tick-borne bacterial genera in adult Ixodes scapularis removed from white-tailed deer in western Tennessee. Parasit Vectors. 2014;7:doi: 10.1186/s13071-014-0473-y. [DOI] [PMC free article] [PubMed]
- 61.McCall JW, Baker CF, Mather TN, Chester ST, McCall SD, Irwin JP, et al. The ability of a topical novel combination of fipronil, amitraz and (S)-methoprene to protect dogs from Borrelia burgdorferi and Anaplasma phagocytophilum infections transmitted by Ixodes scapularis. Vet Parasitol. 2011;179:335–42. doi: 10.1016/j.vetpar.2011.03.046. [DOI] [PubMed] [Google Scholar]
- 62.Michalski M, Rosenfield C, Erickson M, Selle R, Bates K, Essar D, et al. Anaplasma phagocytophilum in central and western Wisconsin: a molecular survey. Parasitol Res. 2006;99:694–9. doi: 10.1007/s00436-006-0217-9. [DOI] [PubMed] [Google Scholar]
- 63.Moncayo AC, Cohen SB, Fritzen CM, Huang E, Yabsley MJ, Freye JD, et al. Absence of Rickettsia rickettsii and occurrence of other spotted fever group rickettsiae in ticks from Tennessee. Am J Trop Med Hyg. 2010;83:653–7. doi: 10.4269/ajtmh.2010.09-0197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Moreno CX, Moy F, Daniels TJ, Godfrey HP, Cabello FC. Molecular analysis of microbial communities identified in different developmental stages of Ixodes scapularis ticks from Westchester and Dutchess Counties, New York. Environ Microbiol. 2006;8:761–72. doi: 10.1111/j.1462-2920.2005.00955.x. [DOI] [PubMed] [Google Scholar]
- 65.Ogden NH, Margos G, Aanensen DM, Drebot MA, Feil EJ, Hanincova K, et al. Investigation of genotypes of Borrelia burgdorferi in Ixodes scapularis ticks collected during surveillance in Canada. Appl Environ Microbiol. 2011;77:3244–54. doi: 10.1128/AEM.02636-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ogden NH, Lindsay LR, Hanincova K, Barker IK, Bigras-Poulin M, Charron DF, et al. Role of migratory birds in introduction and range expansion of Ixodes scapularis ticks and of Borrelia burgdorferi and Anaplasma phagocytophilum in Canada. Appl Environ Microbiol. 2008;74:1780–90. doi: 10.1128/AEM.01982-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Pritt BS, Sloan LM, Johnson DK, Munderloh UG, Paskewitz SM, McElroy KM, et al. Emergence of a new pathogenic Ehrlichia species, Wisconsin and Minnesota, 2009. N Engl J Med. 2011;365:422–9. doi: 10.1056/NEJMoa1010493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Prusinski MA, Kokas JE, Hukey KT, Kogut SJ, Lee J, Backenson PB. Prevalence of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae), Anaplasma phagocytophilum (Rickettsiales: Anaplasmataceae), and Babesia microti (Piroplasmida: Babesiidae) in Ixodes scapularis (Acari: Ixodidae) collected from recreational lands in the Hudson Valley Region, New York State. J Med Entomol. 2014;51:226–36. doi: 10.1603/ME13101. [DOI] [PubMed] [Google Scholar]
- 69.Roellig DM, Fang QQ. Detection of Anaplasma phagocytophilum in ixodid ticks from equine-inhabited sites in the Southeastern United States. Vector Borne Zoonotic Dis. 2012;12:330–2. doi: 10.1089/vbz.2011.0757. [DOI] [PubMed] [Google Scholar]
- 70.Rollend L, Bent SJ, Krause PJ, Usmani-Brown S, Steeves TK, States SL, et al. Quantitative PCR for detection of Babesia microti in Ixodes scapularis ticks and in human blood. Vector Borne Zoonotic Dis. 2013;13:784–90. doi: 10.1089/vbz.2011.0935. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Russart NM, Dougherty MW, Vaughan JA. Survey of ticks (Acari: Ixodidae) and tick-borne pathogens in North Dakota. J Med Entomol. 2014;51:1087–90. doi: 10.1603/ME14053. [DOI] [PubMed] [Google Scholar]
- 72.Schulze TL, Jordan RA, Healy SP, Roegner VE, Meddis M, Jahn MB, et al. Relative abundance and prevalence of selected Borrelia infections in Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae) from publicly owned lands in Monmouth County, New Jersey. J Med Entomol. 2006;43:1269–75. doi: 10.1603/0022-2585(2006)43[1269:RAAPOS]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 73.Schulze TL, Jordan RA, Healy SP, Roegner VE. Detection of Babesia microti and Borrelia burgdorferi in host-seeking Ixodes scapularis (Acari: Ixodidae) in Monmouth County, New Jersey. J Med Entomol. 2013;50:379–83. doi: 10.1603/ME12088. [DOI] [PubMed] [Google Scholar]
- 74.Scoles GA, Papero M, Beati L, Fish D. A relapsing fever group spirochete transmitted by Ixodes scapularis ticks. Vector Borne Zoonotic Dis. 2001;1:21–34. doi: 10.1089/153036601750137624. [DOI] [PubMed] [Google Scholar]
- 75.Shukla SK, Vandermause MF, Belongia EA, Reed KD, Paskewitz SM, Kazmierczak J. Importance of primer specificity for PCR detection of Anaplasma phagocytophila among Ixodes scapularis ticks from Wisconsin. J Clin Microbiol. 2003;41:4006. doi: 10.1128/JCM.41.8.4006.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Smith MP, Ponnusamy L, Jiang J, Ayyash LA, Richards AL, Apperson CS. Bacterial pathogens in ixodid ticks from a Piedmont County in North Carolina: prevalence of rickettsial organisms. Vector Borne Zoonotic Dis. 2010;10:939–52. doi: 10.1089/vbz.2009.0178. [DOI] [PubMed] [Google Scholar]
- 77.Smith RP, Jr, Elias SP, Borelli TJ, Missaghi B, York BJ, Kessler RA, et al. Human babesiosis, Maine, USA, 1995-2011. Emerg Infect Dis. 2014;20:1727–30. doi: 10.3201/eid2010.130938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Steiner FE, Pinger RR, Vann CN, Abley MJ, Sullivan B, Grindle N, et al. Detection of Anaplasma phagocytophilum and Babesia odocoilei DNA in Ixodes scapularis (Acari: Ixodidae) collected in Indiana. J Med Entomol. 2006;43:437–42. doi: 10.1603/0022-2585(2006)043[0437:DOAPAB]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 79.Steiner FE, Pinger RR, Vann CN, Grindle N, Civitello D, Clay K, et al. Infection and co-infection rates of Anaplasma phagocytophilum variants, Babesia spp., Borrelia burgdorferi, and the rickettsial endosymbiont in Ixodes scapularis (Acari: Ixodidae) from sites in Indiana, Maine, Pennsylvania, and Wisconsin. J Med Entomol. 2008;45:289–97. doi: 10.1603/0022-2585(2008)45[289:IACROA]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 80.Taft SC, Miller MK, Wright SM. Distribution of borreliae among ticks collected from eastern states. Vector Borne Zoonotic Dis. 2005;5:383–9. doi: 10.1089/vbz.2005.5.383. [DOI] [PubMed] [Google Scholar]
- 81.Telford Iii SR, Goethert HK, Cunningham JA. Prevalence of Ehrlichia muris in Wisconsin deer ticks collected during the mid 1990s. Open Microbiol J. 2011;5:18–20. doi: 10.2174/1874285801105010018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Tokarz R, Kapoor V, Samuel JE, Bouyer DH, Briese T, Lipkin WI. Detection of tick-borne pathogens by MassTag polymerase chain reaction. Vector Borne Zoonotic Dis. 2009;9:147–52. doi: 10.1089/vbz.2008.0088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Tokarz R, Jain K, Bennett A, Briese T, Lipkin WI. Assessment of polymicrobial infections in ticks in New York state. Vector Borne Zoonotic Dis. 2010;10:217–21. doi: 10.1089/vbz.2009.0036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Tokarz R, Williams SH, Sameroff S, Sanchez Leon M, Jain K, Lipkin WI. Virome analysis of Amblyomma americanum, Dermacentor variabilis, and Ixodes scapularis ticks reveals novel highly divergent vertebrate and invertebrate viruses. J Virol. 2014;88:11480–92. doi: 10.1128/JVI.01858-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Ullmann AJ, Gabitzsch ES, Schulze TL, Zeidner NS, Piesman J. Three multiplex assays for detection of Borrelia burgdorferi sensu lato and Borrelia miyamotoi sensu lato in field-collected Ixodes nymphs in North America. J Med Entomol. 2005;42:1057–62. doi: 10.1603/0022-2585(2005)042[1057:TMAFDO]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 86.Walk ST, Xu G, Stull JW, Rich SM. Correlation between tick density and pathogen endemicity, New Hampshire. Emerg Infect Dis. 2009;15:585–7. doi: 10.3201/eid1504.080940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Williamson PC, Billingsley PM, Teltow GJ, Seals JP, Turnbough MA, Atkinson SF. Borrelia, Ehrlichia, and Rickettsia spp. in ticks removed from persons, Texas, USA. Emerg Infect Dis. 2010;16:441–6. doi: 10.3201/eid1603.091333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Yabsley MJ, Nims TN, Savage MY, Durden LA. Ticks and tick-borne pathogens and putative symbionts of black bears (Ursus americanus floridanus) from Georgia and Florida. J Parasitol. 2009;95:1125–8. doi: 10.1645/GE-2111.1. [DOI] [PubMed] [Google Scholar]
- 89.Adelson ME, Rao RV, Tilton RC, Cabets K, Eskow E, Fein L, et al. Prevalence of Borrelia burgdorferi, Bartonella spp., Babesia microti, and Anaplasma phagocytophila in Ixodes scapularis ticks collected in Northern New Jersey. J Clin Microbiol. 2004;42:2799–801. doi: 10.1128/JCM.42.6.2799-2801.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Schulze TL, Jordan RA, Schulze CJ, Mixson T, Papero M. Relative encounter frequencies and prevalence of selected Borrelia, Ehrlichia, and Anaplasma infections in Amblyomma americanum and Ixodes scapularis (Acari: Ixodidae) ticks from central New Jersey. J Med Entomol. 2005;42:450–6. doi: 10.1603/0022-2585(2005)042[0966:HAOISA]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 91.Parola P, Paddock CD, Raoult D. Tick-borne rickettsioses around the world: emerging diseases challenging old concepts. Clin Microbiol Rev. 2005;18:719–56. doi: 10.1128/CMR.18.4.719-756.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Rizzoli A, Silaghi C, Obiegala A, Rudolf I, Hubálek Z, Földvári G, et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: new hazards and relevance for public health. Front Public Health. 2014;2:doi: 10.3389/fpubh.2014.00251. [DOI] [PMC free article] [PubMed]
- 93.Centers for Disease Control and Prevention. Statistics and epidemiology: annual cases of anaplasmosis in the United States. 2013. http://www.cdc.gov/anaplasmosis/stats/#casesbyyear. Accessed 11 Sept 2015.
- 94.Adams DA, Fullerton J, Jajosky R, Sharp P, Onweh DH, Schley AW, et al. Summary of notifiable diseases, United States, 2013. MMWR Morb Mortal Wkly Rep. 2015;62:1–119. [DOI] [PubMed]
- 95.Dumler JS, Choi KS, Garcia-Garcia JC, Barat NS, Scorpio DG, Garyu JW, et al. Human granulocytic anaplasmosis and Anaplasma phagocytophilum. Emerg Infect Dis. 2005;11:1828–34. doi: 10.3201/eid1112.050898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Hoang D, Johnson E, Schiffman J, Davis P, Neitzel D, Sloan L, et al. Human infection with Ehrlichia muris–like pathogen, United States, 2007-2013. Emerg Inf Dis. 2015;doi: 10.3201/eid2110.150143. [DOI] [PMC free article] [PubMed]
- 97.Paddock CD, Sumner JW, Comer JA, Zaki SR, Goldsmith CS, Goddard J, et al. Rickettsia parkeri: a newly recognized cause of spotted fever rickettsiosis in the United States. Clin Infect Dis. 2004;38:805–11. doi: 10.1086/381894. [DOI] [PubMed] [Google Scholar]
- 98.Stanek G, Wormser GP, Gray J, Strle F. Lyme borreliosis. Lancet. 2012;379:461–73. doi: 10.1016/S0140-6736(11)60103-7. [DOI] [PubMed] [Google Scholar]
- 99.Ontario Agency for Health Protection and Promotion (Public Health Ontario). Vector-borne diseases 2014 summary report. 2015. http://www.publichealthontario.ca/en/eRepository/Vector_Borne_Diseases_Summary_Report_2014.pdf. Accessed on 2 Nov 2015.
- 100.Krause PJ, Narasimhan S, Wormser GP, Barbour AG, Platonov AE, Brancato J, et al. Borrelia miyamotoi sensu lato seroreactivity and seroprevalence in the Northeastern United States. Emerg Infect Dis. 2014;20:1183–90. doi: 10.3201/eid2007.131587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Gugliotta JL, Goethert HK, Berardi VP, Telford SR. Meningoencephalitis from Borrelia miyamotoi in an immunocompromised patient. N Engl J Med. 2013;368:240–5. doi: 10.1056/NEJMoa1209039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Krause PJ, Fish D, Narasimhan S, Barbour AG. Borrelia miyamotoi infection in nature and in humans. Clin Microbiol Infect. 2015;21:631–9. doi: 10.1016/j.cmi.2015.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Pritt BS, Mead PS, Johnson DKH, Neitzel DF, Respicio-Kingry L, Davis JP, et al. Identification of a novel pathogenic Borrelia species causing Lyme borreliosis with unusually high spirochaetaemia: a descriptive study. Lancet Infect Dis. 2016;doi: 10.1016/S1473-3099(15)00464-8. [DOI] [PMC free article] [PubMed]
- 104.Dolan MC, Hojgaard A, Hoxmeier JC, Replogle AJ, Respicio-Kingry LB, Sexton C, et al. Vector competence of the blacklegged tick, Ixodes scapularis, for the recently recognized Lyme borreliosis spirochete Candidatus Borrelia mayonii. Ticks Tick Borne Dis. 2016;doi: 10.1016/j.ttbdis.2016.02.012. [DOI] [PubMed]
- 105.Narasimhan S, Rajeevan N, Liu L, Zhao YO, Heisig J, Pan J, et al. Gut microbiota of the tick vector Ixodes scapularis modulate colonization of the Lyme disease spirochete. Cell Host Microbe. 2014;15:58–71. doi: 10.1016/j.chom.2013.12.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Narasimhan S, Fikrig E. Tick microbiome: the force within. Trends Parasitol. 2015;31:315–23. doi: 10.1016/j.pt.2015.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Bagdure SR, Fisher MA, Ryan ME, Khasawneh FA. Rhodococcus erythropolis encephalitis in patient receiving rituximab. Emerg Infect Dis. 2012;18:1377–9. doi: 10.3201/eid1808.110434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Brooke JS. Stenotrophomonas maltophilia: an emerging global opportunistic pathogen. Clin Microbiol Rev. 2012;25:2–41. doi: 10.1128/CMR.00019-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Jones A, Morgan D, Walsh A, Turton J, Livermore D, Pitt T, et al. Importation of multidrug-resistant Acinetobacter spp. infections with casualties from Iraq. Lancet Infect Dis. 2006;6:317–8. [DOI] [PubMed]
- 110.Werren JH, Baldo L, Clark ME. Wolbachia: master manipulators of invertebrate biology. Nat Rev Micro. 2008;6:741–51. doi: 10.1038/nrmicro1969. [DOI] [PubMed] [Google Scholar]
- 111.Centers for Disease Control and Prevention. Babesiosis: 2013 data and statistics. 2015. http://www.cdc.gov/parasites/babesiosis/data-statistics/index.html. Accessed 10 Sep 2015.
- 112.Homer MJ, Aguilar-Delfin I, Telford SR, 3rd, Krause PJ, Persing DH. Babesiosis. Clin Microbiol Rev. 2000;13:451–69. doi: 10.1128/CMR.13.3.451-469.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Diuk-Wasser MA, Liu Y, Steeves TK, Folsom-O'Keefe C, Dardick KR, Lepore T, et al. Monitoring human babesiosis emergence through vector surveillance New England, USA. Emerg Infect Dis. 2014;20:225–31. doi: 10.3201/eid2002.130644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Mather TN, Telford SR, 3rd, Moore SI, Spielman A. Borrelia burgdorferi and Babesia microti: efficiency of transmission from reservoirs to vector ticks (Ixodes dammini) Exp Parasitol. 1990;70:55–61. doi: 10.1016/0014-4894(90)90085-Q. [DOI] [PubMed] [Google Scholar]
- 115.Piesman J, Mather TN, Dammin GJ, Telford SR, 3rd, Lastavica CC, Spielman A. Seasonal variation of transmission risk of Lyme disease and human babesiosis. Am J Epidemiol. 1987;126:1187–9. doi: 10.1093/oxfordjournals.aje.a114757. [DOI] [PubMed] [Google Scholar]
- 116.Tavakoli NP, Wang H, Dupuis M, Hull R, Ebel GD, Gilmore EJ, et al. Fatal case of deer tick virus encephalitis. N Engl J Med. 2009;360:2099–107. doi: 10.1056/NEJMoa0806326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.El Khoury MY, Camargo JF, White JL, Backenson BP, Dupuis AP, 2nd, Escuyer KL, et al. Potential role of deer tick virus in Powassan encephalitis cases in Lyme disease-endemic areas of New York, U.S.A. Emerg Infect Dis. 2013;19:1926–33. doi: 10.3201/eid1912.130903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Hudson P, Greenman J. Competition mediated by parasites: biological and theoretical progress. Trends Ecol Evol. 1998;13:387–90. doi: 10.1016/S0169-5347(98)01475-X. [DOI] [PubMed] [Google Scholar]
- 119.Strickland R, Gerriah RR, Hourrigan JL, Schubert, GO. Ticks of veterinary importance. Washington, DC: Animal and Plant Health Inspection Service, US Department of Agriculture, Agriculture Handbook No. 485; 1976.
- 120.Ministry of Health and Long-term Care. Infectious Disease Protocols, 2013. 2013. http://www.health.gov.on.ca/en/pro/programs/publichealth/oph_standards/infdispro.aspx. Accessed 10 Sep 2015.
- 121.Manitoba Health, Healthy Living and Seniors. Reporting of diseases and conditions regulation. 2009. http://web2.gov.mb.ca/laws/regs/current/_pdf-regs.php?reg=37/2009. Accessed 10 Sep 2015
- 122.Centre intégré de santé et de services sociaux. Liste pour les médecins des maladies, infections et intoxications à déclaration obligatoire (MADO) – mise à jour juillet 2005. 2005. http://publications.msss.gouv.qc.ca/acrobat/f/documentation/preventioncontrole/03-268-05.pdf. Accessed 10 Sep 2015.
- 123.Illinois Department of Public Health. Control of communicable disease code. 2014. http://www.idph.state.il.us/health/infect/id_reporting.htm. Accessed 10 Sep 2015.
- 124.Indiana Sate Department of Health. Reportable vs. non-reportable conditions/diseases/events. 2015. http://www.in.gov/isdh/23978.htm. Accessed 10 Sep 2015.
- 125.Michigan Department of Health and Human Services. Michigan's communicable disease rules. 2016. http://www.michigan.gov/documents/mdch/Reportable_Diseases_Michigan_by_Pathogen_478489_7.pdf. Accessed 2 Apr 2016.
- 126.Minnesota Department of Health. Communicable disease rule, chapter 4605. 2005. http://www.health.state.mn.us/divs/idepc/dtopics/reportable/rule/rule.html. Accessed 10 Sep 2015.
- 127.New York State Department of Health. Communicable disease reporting requirements. 2011. https://www.health.ny.gov/forms/instructions/doh-389_instructions.pdf. Accessed 2 Apr 2016.
- 128.Ohio Department of Health. Ohio’s reportable infectious diseases. 2015. http://www.odh.ohio.gov/reportablediseases. Accessed 10 Sep 2015.
- 129.Pennsylvania Department of Health. List of reportable diseases. 2012. http://www.portal.state.pa.us/portal/server.pt?open = 514&objID = 557245&mode = 2. Accessed 10 Sep 2015.
- 130.Wisconsin Department of Health Services. Communicable diseases and other notifiable conditions. 2009. http://docs.legis.wisconsin.gov/code/admin_code/dhs/110/145_a. Accessed 10 Sep 2015.
- 131.Clark KL, Leydet B, Hartman S. Lyme borreliosis in human patients in Florida and Georgia. USA Int J Med Sci. 2013;10:915–31. doi: 10.7150/ijms.6273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Girard YA, Fedorova N, Lane RS. Genetic diversity of Borrelia burgdorferi and detection of B. bissettii-like DNA in serum of north-coastal California residents. J Clin Microbiol. 2011;49:945–54. doi: 10.1128/JCM.01689-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Rudenko N, Golovchenko M, Grubhoffer L, Oliver JH., Jr Updates on Borrelia burgdorferi sensu lato complex with respect to public health. Ticks Tick Borne Dis. 2011;2:123–8. doi: 10.1016/j.ttbdis.2011.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.James AM, Liveris D, Wormser GP, Schwartz I, Montecalvo MA, Johnson BJ. Borrelia lonestari infection after a bite by an Amblyomma americanum tick. J Infect Dis. 2001;183:1810–4. doi: 10.1086/320721. [DOI] [PubMed] [Google Scholar]
- 135.Katzourakis A, Gifford RJ. Endogenous viral elements in animal genomes. PLoS Genet. 2010;6:e1001191. doi: 10.1371/journal.pgen.1001191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Bell-Sakyi L, Attoui H. Endogenous tick viruses and modulation of tick-borne pathogen growth. Front Cell Infect Microbiol. 2013;3:doi: 10.3389/fcimb.2013.00025. [DOI] [PMC free article] [PubMed]