

Abstract
Isoflurane (ISO) exhibits neuroprotective effects against inflammation and apoptosis. However, the role of ISO in motoneuronal apoptosis induced by activated microglia remains poorly studied. We investigated the protective effects of ISO on the apoptosis of ventral spinal cord 4.1 (VSC4.1) motoneurons induced by lipopolysaccharide (LPS)-activated BV-2 microglia. Results indicated that ISO inhibited NF-κB activation and pro-inflammatory cytokine release in LPS-treated BV-2 microglia. Conditioned medium (CM) from activated BV-2 cells treated by ISO directly prevented VSC4.1 motoneurons from LPS-CM-induced neuronal apoptosis, as determined by the following: reductions in caspase-8, caspase-9, and caspase-3 activities; downregulation of pro-apoptotic procaspase-8, cleaved (cl)-caspase-8, procaspase-9, cl-caspase-9, caspase-3, cl-caspase-3, Bid, Bax, and cytochrome c expression; and upregulation of anti-apoptotic Bcl-2 expression in LPS-CM-cultured VSC4.1 motoneurons. Findings demonstrated that ISO inhibits BV-2 microglia activation and alleviates VSC4.1 motoneuronal apoptosis induced by microglial activation. These effects suggest that ISO can be used as an alternative agent for reducing neuronal apoptosis.
Keywords: Isoflurane, LPS, BV-2 microglia, VSC4.1 motoneuron, NF-κB, apoptosis
Introduction
Neurodegenerative diseases, such as amyotrophic lateral sclerosis (ALS) and spinal cord injury (SCI), lead to progressive and selective death of motoneurons [1]. The mechanisms of motoneuron death in both ALS and SCI involve inflammation, ischemia, free radical injury, excitotoxicity, and protease activation [2]. Despite significantly improved therapies for these disorders, no effective interventions for preventing excessive neuroinflammation and progressive neurodegeneration have been studied, particularly in motoneurons. Thus, discovering a new therapeutic agent for promoting motoneuron survival and enhancing neurological outcomes is urgently imperative.
Microglia, the resident macrophage-like cells within the central nervous system (CNS), results in neuron injury and death in the pathology of many neurodegenerative diseases [3]. Microglia-mediated cytotoxicity significantly contributes to the pathogenesis of neurodegeneration [4]. Microglia can be activated by various pathological states, including CNS injury, ischemia, and endotoxin (LPS). Microglia activation can exert synergistic effects on neurotoxicity by activating NF-κB signaling via release of pro-inflammatory mediators, such as tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and IL-6, which mediate neuronal cell death, particularly in neurodegenerative diseases [5-7]. Pro-inflammatory cytokines may cause secondary damage to motoneurons, leading to decreased function and recovery in SCI patients [8]. Thus, modulation of microglia-mediated inflammatory responses is important in the development of a new therapeutic agent against neurodegenerative diseases.
Isoflurane (ISO) is a commonly used inhaled anesthetic with anti-inflammatory, anti-oxidative, and anti-apoptotic properties through its inhibition of NF-κB signaling and subsequent reduction of pro-inflammatory cytokines and chemokines [9-11]. ISO post-treatment prevents neonatal rat hypoxic-ischemic brain injury by impeding cell apoptosis triggered by inflammatory stress [12]. However, whether ISO elicits protective effects on motoneurons insulted by activated microglial cells remains unclear.
In this study, we demonstrate that ISO inhibits nuclear factor (NF)-κB activation and pro-inflammatory cytokine release in lipopolysaccharide (LPS)-activated BV-2 microglia. Moreover, ISO reduces the apoptosis of ventral spinal cord 4.1 (VSC4.1) motoneurons stimulated with LPS-conditioned medium (CM) in vitro. Our results suggest that ISO can be used as a potential therapeutic agent against neurodegenerative diseases.
Materials and methods
Reagents
LPS (Escherichia coli serotype 055:B5) was obtained from Sigma-Aldrich (St. Louis, MO, USA). ISO was supplied by Baxter Healthcare Corporation (Deerfield, IL, USA). All reagents for cell culture were purchased from Gibco BRL (Grand Island, NY, USA). Rabbit anti-mouse NF-κB p65, phospho-IκB-α, procaspase-8, cleaved (cl)-caspase-8, procaspase-9, cl-caspase-9, procaspase-3, and cl-caspase-3 polyclonal antibodies (pAbs) were obtained from Santa Cruz Biotechnology (Dallas, TX, USA). Rabbit anti-mouse Bax, Bcl-2, and cytochrome c pAbs were supplied by Cell Signaling Technology Inc. (Danvers, MA, USA). Rabbit anti-mouse b-actin and histone-3 monoclonal antibodies (mAbs) were procured from Sigma. Horseradish peroxidase-conjugated anti-rabbit IgG was obtained from Chemicon (Temecula, CA, USA). All other chemicals were purchased from Sigma-Aldrich, unless otherwise stated.
Microglial cell culture and treatment
An immortalized mouse BV-2 microglial cell line was purchased from Union Cell Resource Center, Chinese Academy of Medical Sciences (Beijing, China). The cell line was characterized by the provider using a gene profiling analysis, and the cells were used no later than 6 months after receipt. The cells were routinely grown in Dulbecco’s modified Eagle’s medium (DMEM) containing 10% heat-inactivated fetal bovine serum (FBS), streptomycin (10 μg/ml), and penicillin (10 U/ml) at 37°C under 5% CO2, and cells from passages 2-4 were used for experiments. Cells were seeded in 6-well plates, incubated overnight, and then subjected to LPS (100 ng/ml) or control (Ctrl) culture media treatment for 0.5 h. The cells were subsequently exposed to room air (RA) with or without ISO for 0.5 h at 2 L/min in a metabolic chamber (Columbus Instruments, Columbus, OH, USA). During ISO exposure, the ISO concentration was continuously verified by sampling the exhaust gas with a Datex Capnomac (SOMA Technology Inc., Cheshire, CT, USA) [13]. Four treatment groups were established, namely, Ctrl + RA, Ctrl + ISO, LPS + RA, and LPS + ISO.
VSC4.1 motoneuron cell culture and CM treatment
VSC4.1 motoneurons were formed by fusion of dissociated embryonic rat ventral spinal cord neuron with mouse N18TG2 neuroblastoma cells [14]. VSC4.1 cells were grown in 24-well plates containing a DMEM/F12 medium with HEPES, pyridoxine, and NaHCO3, supplemented with 2% Sato’s components, 1% penicillin/streptomycin, and 2% FBS. CMs were collected as previously described [15]. After 24 h of LPS or Ctrl treatment, the supernatants of BV-2 cells in the four groups was collected, centrifuged at 1000 g for 5 min, filtered through 0.22 µm pore-diameter Millipore filters, and dialyzed using the Slide-A-Lyzer Dialysis Cassette (Pierce Biotechnology Inc., Rockford, IL) overnight to exclude any possible influence of LPS. The fresh media with CMs at 1:1 were supplemented to incubate VSC4.1 cells for the subsequent experiments.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay
Cell viability was measured using an MTT assay. BV-2 cells (1×104 cells/well) were seeded in 96-well plates and treated with different concentrations of ISO (0.7%, 1.4%, and 2.1%) for 24 h. MTT (0.5 mg/ml) was then added into each well and incubated at 37°C for 4 h. The supernatants were removed, and 150 µl of dimethylsulfoxide was added to each well. The plates were immediately read at 540 nm with the use of a microplate reader (BioTek Instruments Inc., Winooski, VT, USA). For VSC4.1 cell viability assay, cells (1×104 cells/well) were cultured in a proliferative medium with various CMs and measured by MTT assay as described in BV-2 cells.
Lactate dehydrogenase (LDH) assay
The membrane integrity of BV-2 and VSC4.1 cells was evaluated using Thermo Scientific Pierce LDH Cytotoxicity Assay Kits (Thermo Scientific, Rockford, IL, USA) in accordance with the manufacturer’s instructions. Assays were performed after a 7-day culture to evaluate the membrane integrity of the cells, and data were recorded at a wave length of 490 and 680 nm by using a microplate reader (BioTek).
Quantitative real-time PCR (qPCR)
Total RNA from BV-2 cells was isolated and extracted using TRIzol Reagent (Invitrogen, Carlsbad, CA, USA) in accordance with the manufacturer’s instructions. cDNA was generated using a reverse transcription kit (Promega, Madison, WI, USA). qPCR was performed using a standard iQ™ SYBR Green Supermix Kit (Bio-Rad, Berkeley, USA), and PCR-specific amplification was assessed using Mastercycler® ep realplex (Eppendorf, Hamburg, Germany). Glycerinaldehyd-3-phosphate dehydrogenase (GAPDH) was used as an endogenous control. Relative mRNA levels were calculated by the comparative 2-ΔΔCt method [16]. The primers used for PCR amplification were as follows: for TNF-α, forward 5’-TTGACCTCAGCGCTGAGTTG-3’ and reverse 5’-CCTGTAGCCCACGTCGTAG C-3’; for IL-1β, forward 5’-CAGGATGAGGACATGAGCACC-3’ and reverse 5’-CTCTGCAGACTCA AACTCCAC-3’; for IL-6, forward 5’-GTACTCCAGAAGACCAGAGG-3’ and reverse 5’-TGCTGGTG ACAACCACGGCC-3’; for GAPDH, forward 5’-AGGTCGGTGTGAACGGATTTG-3’ and reverse 5’-TGTAGACCATGTAGTTGAGGTCA-3’.
Enzyme-linked immunosorbent assay (ELISA)
Production of TNF-α, IL-1β, and IL-6 in the supernatants of the cultured BV-2 cells was measured with commercial ELISA kits (R&D, Minneapolis, MN, USA) in accordance with the manufacturer’s instructions. The absorbance was read at 490 nm on a microplate scanner (BioTek), and the samples were run in triplicate.
Western blot analysis
Cytosolic and nuclear proteins of BV-2 cells were prepared using a Nuclear Extract Kit (Active Motif, Carlsbad, CA, USA). NF-κB p65 levels were quantified in nuclear fractions. Phospho-IκB-α level was quantified in cytosolic fractions. Proteins in VSC4.1 cells were prepared using RIPA lysis buffer (Beyotime Biotechnology, Jiangsu, China). The ultimate extracts were boiled, separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis, electrotransferred onto nitrocellulose membranes, and immunoblotted with primary antibodies. Equivalent sample loading was confirmed by probing with anti-β-actin mAb and anti-lamin B mAb. Detection was performed using an enhanced chemiluminescence assay kit (Pierce, Rockford, IL, USA). Standards and samples were run in triplicate in accordance with the manufacturer’s instructions.
NF-κB DNA-binding activity assay
The NF-κB DNA-binding activity was quantified using a TransAM NF-κB p65 Transcription Factor Assay Kit (Active Motif). Nuclear extracts of the BV-2 cells were prepared using the Nuclear Extraction Kit (Active Motif). All standards and samples were analyzed in triplicate in accordance with the manufacturer’s instructions.
Apoptosis assay by flow cytometry
The apoptosis of VSC4.1 cells was assessed by Annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) staining. The cells were harvested, centrifuged, and resuspended in binding buffer. Approximately 10 µl of ready-to-use Annexin V-FITC (BD Bioscience, MA, USA) was added into the mixture, incubated at 37°C for 15 min, and counterstained with 5 µl of PI in the dark for 30 min. Annexin V-FITC and PI fluorescence were assessed by BD FACSCalibur flow cytometry (BD Bioscience, MA, USA). Results were analyzed with the CellQuest software (BD Bioscience). Each sample was prepared in triplicate.
Nucleosomal fragmentation assay
VSC4.1 cell apoptosis was quantified using a nucleosomal fragmentation kit (Cell Death Detection ELISA PLUS; Roche Applied Science, Indianapolis, IN, USA) in accordance with the manufacturer’s protocol. The absorbance values were normalized with reference to control-treated cells to derive a nucleosomal enrichment factor.
Mitochondrial membrane potential analysis
The loss of mitochondrial potential was measured using the fluorescent probe JC-1 (MitoPT, Immunohistochemistry Technologies, Bloomington, MN, USA). The VSC4.1 cells were stained with JC-1, which exhibits potential-dependent accumulation in the mitochondria. At low membrane potentials, JC-1 is present as a monomer and produces green fluorescence (emission at 527 nm). At high membrane potentials or concentrations, JC-1 forms J aggregates (emission at 590 nm) and produces red fluorescence. Fluorescence was measured in a fluorescent plate reader (BioTek).
Quantitative caspase-8, caspase-9, and caspase-3 activity assays
Activity of caspase-8, caspase-9, and caspase-3 in VSC4.1 cells was measured with commercial caspase colorimetric assay kits (Biovision, Palo Alto, CA, USA) in accordance with the manufacturer’s instructions. The colorimetric assays were based on the hydrolysis of the amino acid chains acetyl-Ile-Glu-Thr-Asp p-nitroanilide (Ac-IETD-pNA) by caspase-8, acetyl-Leu-Glu-His-Asp p-nitroanilide (Ac-LEHD-pNA) by caspase-9, and acetyl-Asp-Glu-Val-Asp p-nitroanilide (Ac-DEVD-pNA) by caspase-3, resulting in the release of pNA. Proteolytic reactions were carried out in an extraction buffer containing 200 μg of cytosolic protein extract and 40 μM Ac-IETD-pNA, 40 μM Ac-LEHD-pNA, or 40 μM Ac-DEVD-pNA. The reaction mixtures were incubated at room temperature for 2 h, and the formation of pNA was measured at 405 nm by using a microplate reader (BioTek). Experiments were performed in triplicate.
Statistical analysis
All values are expressed as means ± SD. Differences were analyzed using one-way ANOVA, followed by Tukey post-hoc tests. All analyses were performed using a Prism software package (version 5.01; GraphPad Software, Inc., San Diego, CA, USA). P < 0.05 was considered statistically significant.
Results
Toxicity of ISO to BV-2 microglial cells
We first investigated the effect of ISO on the viability of BV-2 microglial cells. 1.4% ISO exhibited no significant effect on the viability of BV-2 cells. However, the number of survival cells was markedly reduced by 2.1% ISO (Figure 1A). LDH assay showed that cell membrane integrity was obviously impaired by 2.1% ISO, whereas 1.4% ISO showed no significant damage of cell membrane integrity (Figure 1B). These results suggested that ISO conferred toxicity to BV-2 cells at higher concentrations. Thus, we selected the 1.4% ISO dose for the subsequent experiments.
Figure 1.
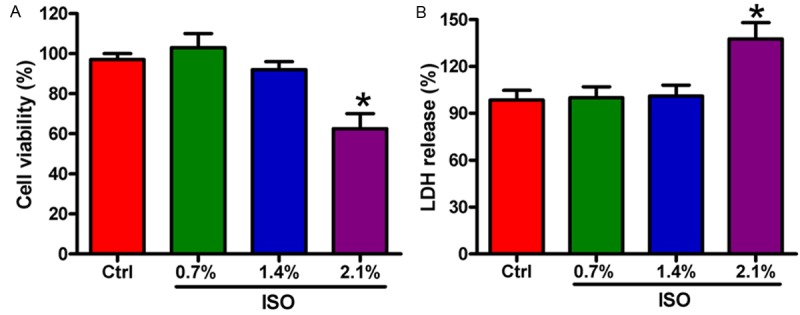
Effects of ISO on the viability and LDH release of BV-2 cells. A. BV-2 cell viability was measured by MTT assay with varying concentrations of ISO treatment. B. LDH release of BV-2 cells was measured in the extracellular medium. Data are means ± SD. *P < 0.05 vs. control (Ctrl) group.
ISO inhibited LPS-induced NF-κB p65 activation in BV-2 cells
To confirm whether ISO-mediated inhibition of LPS-induced neuroinflammation is involved in NF-κB activation, Western blot was performed to evaluate IκB-α degradation and NF-κB p65 nuclear translocation in LPS-treated BV-2 cells. As shown in Figure 2A and 2B, both the phosphorylation of IκB in the cytosol and the nuclear accumulation of NF-κB p65 were increased after LPS stimulation. By contrast, ISO treatment significantly inhibited IκB-α phosphorylation and NF-κB p65 nuclear translocation. NF-κB DNA-binding assay also showed that ISO exposure notably reduced LPS-induced NF-κB DNA-binding activity (Figure 2C). These results indicated that ISO attenuated LPS-induced NF-κB activation in BV-2 cells.
Figure 2.
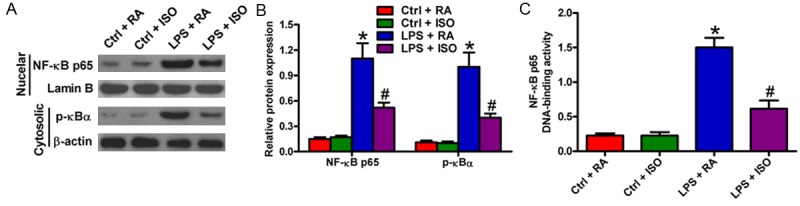
ISO inhibited LPS-induced NF-κB p65 activation in BV-2 cells. A. Representative Western blot results of NF-κB p65 expression in the nucleus and p-IκB-α expression in the cytoplasm. B. Quantitative NF-κB p65 expression against lamin B and p-IκB-α expression against b-actin. The respective densitometric analysis of protein bands results from three independent experiments. C. The NF-κB DNA-binding activity was assayed by determining the optical density. Data are means ± SD. *P < 0.05 vs. Ctrl groups; #P < 0.05 vs. LPS + RA group.
ISO reduced LPS-induced pro-inflammatory cytokine release in BV-2 cells
qPCR and ELISA assays were used to determine the mRNA expression and production of pro-inflammatory cytokines in microglia activated by LPS. The increase in the mRNA expression of TNF-α (Figure 3A), IL-1β (Figure 3B), and IL-6 (Figure 3C) was attenuated by ISO treatment in LPS-stimulated BV-2 cells. Consistently, the increased release of TNF-α (Figure 3D), IL-1β (Figure 3E), and IL-6 (Figure 3F) was reduced by ISO exposure. These results suggested that ISO treatment reduced the mRNA expression and production of pro-inflammatory cytokines in LPS-activated BV-2 cells.
Figure 3.
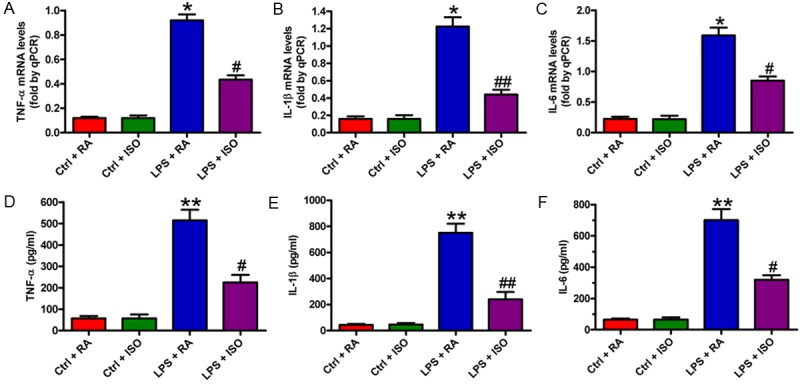
ISO reduced LPS-induced pro-inflammatory cytokines release in BV-2 cells. (A-C) qPCR analysis of mRNA expression of TNF-α (A), IL-1β (B), and IL-6 (C) was normalized against GAPDH. (D-F) The production of TNF-α (D), IL-1β (E), and IL-6 (F) in the supernatants was measured by ELISA. Data are means ± SD. *P < 0.05, **P < 0.01 vs. Ctrl groups; #P < 0.05, ##P < 0.01 vs. LPS + RA group.
CM from ISO-treated microglia promoted survival of LPS-challenged VSC4.1 cells
We investigated the effects of ISO on the survival of VSC4.1 cells simulated by LPS-activated microglia. MTT assay showed that cell viability was significantly inhibited after stimulation with LPS-CM. However, this decrease was restored by LPS + ISO-CM treatment (Figure 4A). In addition, the release of LDH in LPS + ISO-CM-treated cells was markedly decreased compared with that in the LPS-CM-challenged cells (Figure 4B). These data indicated that ISO promoted the survival of LPS-CM-cultured VSC4.1 cells.
Figure 4.
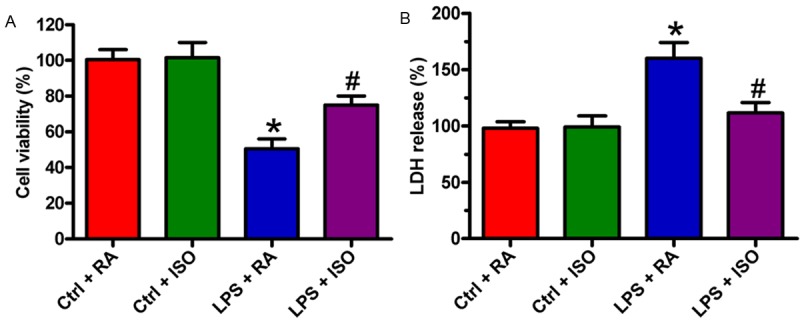
ISO promoted the survival of LPS-CM-cultured VSC4.1 cells. A. MTT assay was used to measure the viability of VSC4.1 cells cultured in the four different CMs. B. LDH release of VSC4.1 cells was measured in the extracellular medium. Data are means ± SD. *P < 0.05 vs. Ctrl groups; #P < 0.05 vs. LPS + RA group.
ISO inhibited LPS-CM-induced apoptosis of VSC4.1 cells
To further confirm the neuroprotective effects of ISO, apoptosis in VSC4.1 cells was evaluated by flow cytometry, nucleosomal fragmentation, and mitochondrial membrane potential assays. As shown in Figure 5A and 5B, VSC4.1 cells with an LPS-CM insult promoted apoptotic cell death, whereas cell apoptosis was markedly inhibited by LPS + ISO-CM treatment. The inhibitory effects of LPS + ISO-CM exposure on VSC4.1 cell apoptosis were also confirmed by nucleosomal fragmentation assay (Figure 5C). In addition, the mitochondrial membrane potential loss of the VSC4.1 cell induced by LPS-CM was markedly inhibited by LPS + ISO-CM exposure (Figure 5D). These data demonstrated that ISO attenuated LPS-CM-induced VSC4.1 cell apoptosis.
Figure 5.
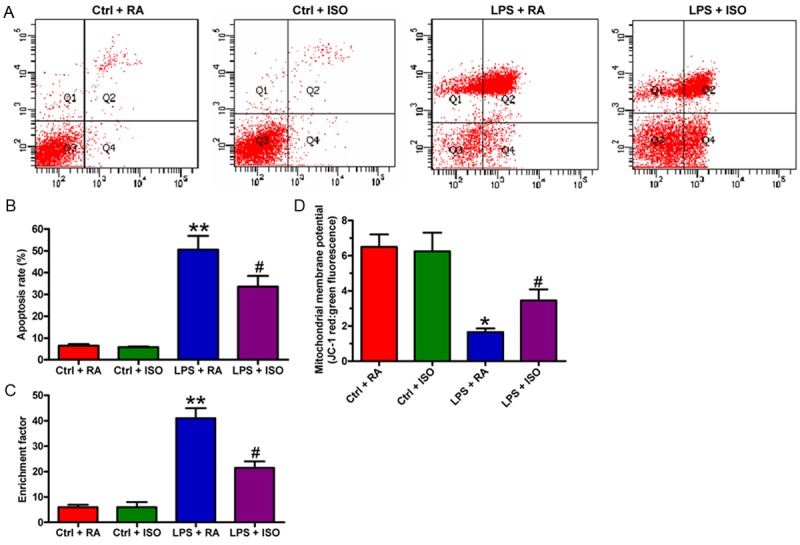
ISO inhibited LPS-CM-trigged VSC4.1 cell apoptosis. (A) Flow cytometry analysis to evaluate apoptosis in VSC4.1 cells. (B) Quantification of the apoptotic cell ratio in (A). (C) Cell apoptosis by nucleosomal fragmentation assay. (D) Changes in the mitochondrial membrane potential were used to assess cell apoptosis. Data are means ± SD. *P < 0.05, **P < 0.01 vs. Ctrl groups; #P < 0.05, ##P < 0.01 vs. LPS + RA group.
ISO reduced caspase-8, caspase-9, and caspase-3 activities of VSC4.1 cells following LPS-CM exposure
To investigate whether the apoptotic inhibition of ISO depended on caspase activation, we examined the activities of caspase-8, caspase-9, and caspase-3. As shown in Figure 6A-C, after treatment of VSC4.1 cells with LPS-CM, the activities of caspase-8, caspase-9, and caspase-3 were enhanced by about 4-, 6-, and 7-fold, respectively, compared with those of the control-CM group. However, ISO significantly attenuated these increases in caspase activities. These results suggested that ISO protected VSC4.1 cells from LPS-CM-induced apoptosis by inhibiting caspase-8, caspase-9, and caspase-3 activities.
Figure 6.
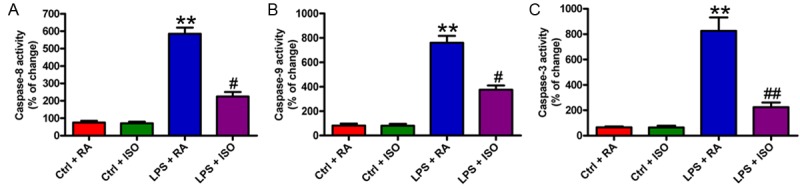
ISO reduced caspase-8, caspase-9, and caspasae-3 activities in LPS-CM-challenged VSC4.1 cells. (A-C) Caspase colorimetric assay kits were used to measure the activities of caspase-8 (A), caspase-9 (B), and caspase-3 (C) in VSC4.1 cells with four different CM treatments. Data are means ± SD. **P < 0.01 vs. Ctrl groups; #P < 0.05, ##P < 0.01 vs. LPS + RA group.
ISO reversed changes in the expression of apoptosis-associated proteins in LPS-CM-treated VSC4.1 cells
To further investigate the protective effects of ISO on VSC4.1 cells with LPS-CM treatment, the expression of apoptosis-associated proteins was analyzed by Western blot. The results showed an increase in the expression of pro-apoptotic procaspase-8, cl-caspase-8 (Figure 7A and 7B); procaspase-9, cl-caspase-9 (Figure 7C and 7D); procaspase-3, cl-caspase-3 (Figure 7E and 7F); and Bid, Bax, and cytochrome c (Figure 7G and 7H). A decrease in the anti-apoptotic Bcl-2 (Figure 7G and 7H) of VSC4.1 cells treated with LPS-CM was also shown. However, treatment with ISO reversed all changes in the proteins mentioned above (Figure 7). All data indicated that ISO inhibited LPS-CM-induced VSC4.1 cell apoptosis by downregulation of pro-apoptotic proteins and upregulation of anti-apoptotic protein.
Figure 7.
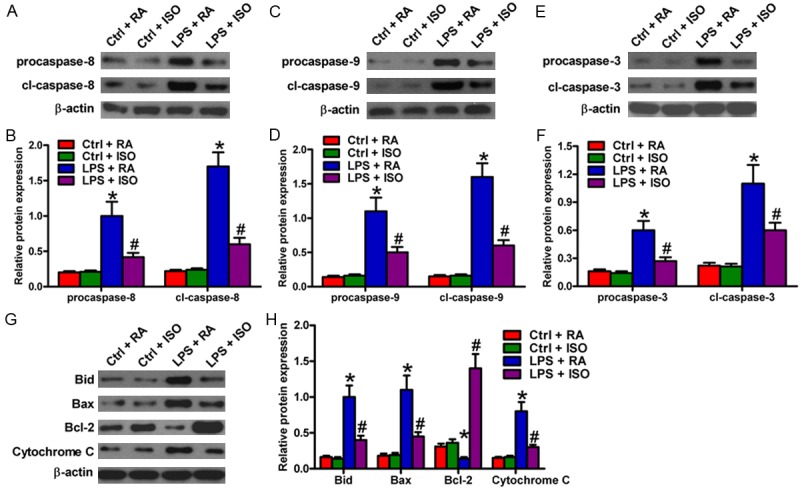
ISO reversed apoptosis-associated protein expression in LPS-CM-stimulated VSC4.1 cells. Western blot was performed to analyze the expression of (A) procaspase-8 and cl-caspase-8; (C) procaspase-9, and cl-caspase-9; (E) procaspase-3, and cl-caspase-3; (G) Bid, Bax, Bcl-2, and cytochrome c. b-actin was used as the loading control. Quantitative expression of (B) procaspase-8 and cl-caspase-8; (D) procaspase-9 and cl-caspase-9; (F) procaspase-3 and cl-caspase-3, (H) Bid, Bax, Bcl-2, and cytochrome c was normalized against β-actin. Data are means ± SD. *P < 0.05 vs. Ctrl groups; #P < 0.05 vs. LPS + RA group.
Discussion
We explored the protective effects of ISO on LPS-activated BV-2 microglia-treated VSC4.1 motoneurons. Key findings are as follows. First, ISO treatment significantly alleviated NF-κB activation and nuclear translocation as well as pro-inflammatory cytokine release in LPS-challenged BV-2 cells. Second, ISO administration promoted LPS-CM-treated VSC4.1 motoneuron survival and inhibited neuronal apoptosis. Third, ISO treatment induced caspase-8, caspase-9, and caspase-3 activities of VSC4.1 cells treated with LPS-CM. Finally, ISO treatment downregulated the expression of pro-apoptotic proteins and upregulated anti-apoptotic protein expression in LPS-CM-treated VSC4.1 cells.
Activated microglia is identified as major cytotoxic effectors and is known to play an essential role in the pathogenesis of neurodegenerative diseases [17,18]. Previous studies indicated that activated microglia inhibit axonal growth and regeneration of neurons by the release of detrimental factors [19,20]. In the present study, we showed that numerous pro-inflammatory cytokines are released following activation of BV-2 cells with LPS. CM from activated microglia containing released cytokines resulted in VSC4.1 motoneuronal death, suggesting that cytokines are the major detrimental factors released following activation of microglia in CNS diseases and are partly responsible for neuronal degeneration. However, ISO treatment significantly inhibited pro-inflammatory cytokine release from LPS-stimulated BV-2 cells and subsequently reduced VSC4.1 motoneuronal death.
Neuroinflammation is one of the most distinct features of neurodegenerative diseases. Classical NF-κB signaling involving the p65/p50 heterodimer is considered a major regulator of inflammation, driving expression of pro-inflammatory cytokines, chemokines, enzymes, and adhesion molecules, many of which are upregulated in neurodegenerative disorders. A previous study showed that activated microglia-induced motoneuronal death via the classical NF-κB pathway in ALS [21]. Notably, NF-κB plays important roles in transcriptional regulation of cytokine expression in microglia under LPS stimulation [22]. In the present study, we showed that NF-κB was activated in LPS-treated BV-2 cells, which was significantly inhibited by ISO treatment.
Motoneuronal death is a well-known characteristic of neurodegenerative diseases. Microglial activation is identified as a major regulator of cell apoptosis [23]. Previous studies indicated that VSC4.1 motoneuronal death is related to both extrinsic and intrinsic apoptosis pathways [24-26]. Caspase-8 is involved in the extrinsic apoptotic pathway prior to mitochondrial permeabilization and cytochrome c release [27]. Caspase-8 activation may induce cleavage of Bid to tBid, resulting in tBid translocation from the cytosol to the mitochondrial membrane to stimulate more efficient oligomerization of Bax, thereby activating the intrinsic apoptotic pathway [24]. The Bcl-2 family proteins are involved in the early stage of apoptosis, and the interaction between the pro-apoptotic Bax and the anti-apoptotic Bcl-2 may be necessary for the intrinsic apoptotic process [28]. Increased Bax/Bcl-2 ratio enhances the release of cytochrome c from mitochondria to cytosol, which activates caspase-9 and caspase-3, thereby inducing cell apoptosis [29]. We showed that exposure of VSC4.1 motoneurons to activated microglia CM significantly increased the expression and activity of caspase-8, caspase-9, and caspase-3, as well as the expression of pro-apoptotic Bid, Bax, and cytochrome c; however, such exposure decreased the expression of anti-apoptotic Bcl-2. However, ISO treatment reversed the changes mentioned above in LPS-CM-treated VSC4.1 motoneurons.
We appreciate limitations of our present studies. According to the previous report, ISO acts not only on microglia but also the neurons directly in vivo. So, we can’t exclude the possibility that ISO has a protective effect on LPS-stimulated neurons directly. However, in the future, we shall perform in vivo experiments to further verify our in vitro findings and to investigate the detailed molecular events, as well as to probe whether ISO reduces LPS-induced neuron apoptosis in vitro and in vivo.
In conclusion, we demonstrated that ISO exhibits neuroprotective effects on motoneurons under activated microglia stimulation by promoting cell survival and inhibiting cell apoptosis via its anti-inflammatory and anti-apoptotic activities. In general, these results suggest that ISO can be used as a therapeutic agent for the treatment of neurodegenerative diseases.
Disclosure of conflict of interest
None.
References
- 1.Lepore AC. Intraspinal cell transplantation for targeting cervical ventral horn in amyotrophic lateral sclerosis and traumatic spinal cord injury. J Vis Exp. 2011:18. doi: 10.3791/3069. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dong XX, Wang Y, Qin ZH. Molecular mechanisms of excitotoxicity and their relevance to pathogenesis of neurodegenerative diseases. Acta Pharmacol Sin. 2009;30:379–387. doi: 10.1038/aps.2009.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zhao W, Xie W, Xiao Q, Beers DR, Appel SH. Protective effects of an anti-inflammatory cytokine, interleukin-4, on motoneuron toxicity induced by activated microglia. J Neurochem. 2006;99:1176–1187. doi: 10.1111/j.1471-4159.2006.04172.x. [DOI] [PubMed] [Google Scholar]
- 4.Smith JA, Das A, Ray SK, Banik NL. Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Res Bull. 2012;87:10–20. doi: 10.1016/j.brainresbull.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cao Q, Li P, Lu J, Dheen ST, Kaur C, Ling EA. Nuclear factor-κB/p65 responds to changes in the Notch signaling pathway in murine BV-2 cells and in amoeboid microglia in postnatal rats treated with the γ-secretase complex blocker DAPT. J Neurosci Res. 2010;88:2701–2714. doi: 10.1002/jnr.22429. [DOI] [PubMed] [Google Scholar]
- 6.Nam KN, Koketsu M, Lee EH. 5-Chloroacetyl-2-amino-1,3-selenazoles attenuate microglial inflammatory responses through NF-kappaB inhibition. Eur J Pharmacol. 2008;589:53–57. doi: 10.1016/j.ejphar.2008.03.034. [DOI] [PubMed] [Google Scholar]
- 7.Wang F, Cui N, Yang L, Shi L, Li Q, Zhang G, Wu J, Zheng J, Jiao B. Resveratrol rescues the impairments of hippocampal neurons stimulated by microglial over-activation in vitro. Cell Mol Neurobiol. 2015;35:1003–1015. doi: 10.1007/s10571-015-0195-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gonzalez-Scarano F, Baltuch G. Microglia as mediators of inflammatory and degenerative diseases. Annu Rev Neurosci. 1999;22:219–240. doi: 10.1146/annurev.neuro.22.1.219. [DOI] [PubMed] [Google Scholar]
- 9.Li JT, Wang H, Li W, Wang LF, Hou LC, Mu JL, Liu X, Chen HJ, Xie KL, Li NL, Gao CF. Anesthetic isoflurane posttreatment attenuates experimental lung injury by inhibiting inflammation and apoptosis. Mediators Inflamm. 2013;2013:108928. doi: 10.1155/2013/108928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mu J, Xie K, Hou L, Peng D, Shang L, Ji G, Li J, Lu Y, Xiong L. Subanesthetic dose of isoflurane protects against zymosan-induced generalized inflammation and its associated acute lung injury in mice. Shock. 2010;34:183–189. doi: 10.1097/SHK.0b013e3181cffc3f. [DOI] [PubMed] [Google Scholar]
- 11.Wang H, Fan J, Li NL, Li JT, Yuan SF, Yi J, Wang L, Chen JH, Lv YG, Yao Q, Wang T, Wang YC, Ling R. A subanesthetic dose of isoflurane during postconditioning ameliorates zymosan-induced neutrophil inflammation lung injury and mortality in mice. Mediators Inflamm. 2013;2013:479628. doi: 10.1155/2013/479628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lee JJ, Li A, Jung HH, Zuo Z. Postconditioning with isoflurane reduced ischemia-induced brain injury in rats. Anesthesiology. 2008;108:1055–1062. doi: 10.1097/ALN.0b013e3181730257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Kawaraguchi Y, Horikawa YT, Murphy AN, Murray F, Miyanohara A, Ali SS, Head BP, Patel PM, Roth DM, Patel HH. Volatile anesthetics protect cancer cells against tumor necrosis factor-related apoptosis-inducing ligand-induced apoptosis via caveolins. Anesthesiology. 2011;115:499–508. doi: 10.1097/ALN.0b013e3182276d42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Smith RG, Alexianu ME, Crawford G, Nyormoi O, Stefani E, Appel SH. Cytotoxicity of immunoglobulins from amyotrophic lateral sclerosis patients on a hybrid motoneuron cell line. Proc Natl Acad Sci U S A. 1994;91:3393–3397. doi: 10.1073/pnas.91.8.3393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Song Q, Jiang Z, Li N, Liu P, Liu L, Tang M, Cheng G. Anti-inflammatory effects of three-dimensional graphene foams cultured with microglial cells. Biomaterials. 2014;35:6930–6940. doi: 10.1016/j.biomaterials.2014.05.002. [DOI] [PubMed] [Google Scholar]
- 16.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 17.Kreutzberg GW. Microglia: a sensor for pathological events in the CNS. Trends Neurosci. 1996;19:312–318. doi: 10.1016/0166-2236(96)10049-7. [DOI] [PubMed] [Google Scholar]
- 18.Nakajima K, Kohsaka S. Microglia: activation and their significance in the central nervous system. J Biochem. 2001;130:169–175. doi: 10.1093/oxfordjournals.jbchem.a002969. [DOI] [PubMed] [Google Scholar]
- 19.Horn KP, Busch SA, Hawthorne AL, van Rooijen N, Silver J. Another barrier to regeneration in the CNS: activated macrophages induce extensive retraction of dystrophic axons through direct physical interactions. J Neurosci. 2008;28:9330–9341. doi: 10.1523/JNEUROSCI.2488-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kitayama M, Ueno M, Itakura T, Yamashita T. Activated microglia inhibit axonal growth through RGMa. PLoS One. 2011;6:e25234. doi: 10.1371/journal.pone.0025234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Frakes AE, Ferraiuolo L, Haidet-Phillips AM, Schmelzer L, Braun L, Miranda CJ, Ladner KJ, Bevan AK, Foust KD, Godbout JP, Popovich PG, Guttridge DC, Kaspar BK. Microglia induce motor neuron death via the classical NF-κB pathway in amyotrophic lateral sclerosis. Neuron. 2014;81:1009–1023. doi: 10.1016/j.neuron.2014.01.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.De Miranda BR, Popichak KA, Hammond SL, Jorgensen BA, Phillips AT, Safe S, Tjalkens RB. The nurr1 activator 1,1-bis(39-indolyl)-1-(p-chlorophenyl) methane blocks inflammatory gene expression in BV-2 microglial cells by inhibiting nuclear factor kB. Mol Pharmacol. 2015;87:1021–1034. doi: 10.1124/mol.114.095398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lee P, Lee J, Kim S, Lee MS, Yagita H, Kim SY, Kim H, Suk K. NO as an autocrine mediator in the apoptosis of activated microglial cells: correlation between activation and apoptosis of microglial cells. Brain Res. 2001;892:380–385. doi: 10.1016/s0006-8993(00)03257-1. [DOI] [PubMed] [Google Scholar]
- 24.Das A, McDowell M, Pava MJ, Smith JA, Reiter RJ, Woodward JJ, Varma AK, Ray SK, Banik NL. The inhibition of apoptosis by melatonin in VSC4.1 motoneurons exposed to oxidative stress, glutamate excitotoxicity, or TNF-α toxicity involves membrane melatonin receptors. J Pineal Res. 2010;48:157–169. doi: 10.1111/j.1600-079X.2009.00739.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Das A, Smith JA, Gibson C, Varma AK, Ray SK, Banik NL. Estrogen receptor agonists and estrogen attenuate TNF-α-induced apoptosis in VSC4.1 motoneurons. J Endocrinol. 2011;208:171–182. doi: 10.1677/JOE-10-0338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.McDowell M, Das A, Smith JA, Varma AK, Ray SK, Banik NL. Neuroprotective effects of genistein in VSC4.1 motoneurons exposed to activated microglial cytokines. Neurochem Int. 2011;59:175–184. doi: 10.1016/j.neuint.2011.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lin CF, Chen CL, Chang WT, Jan MS, Hsu LJ, Wu RH, Tang MJ, Chang WC, Lin YS. Sequential caspase-2 and caspase-8 activation upstream of mitochondrial during ceramide and etoposide-induced apoptosis. J Biol Chem. 2004;279:40755–40761. doi: 10.1074/jbc.M404726200. [DOI] [PubMed] [Google Scholar]
- 28.Merry DE, Korsmeyer SJ. Bcl-2 gene family in the nervous system. Annu Rev Neurosci. 1997;20:245–267. doi: 10.1146/annurev.neuro.20.1.245. [DOI] [PubMed] [Google Scholar]
- 29.Lv CX, Fan TJ, Hu GB, Cong RS. Apoptosisinducing factor and apoptosis. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao. 2003;35:881–885. [PubMed] [Google Scholar]