

Abstract
Zinc (Zn2+) is required for numerous cellular functions. As such, the homeostasis and distribution of intracellular zinc can influence cellular metabolism and signaling. However, the exact distribution of free zinc within live cells remains elusive. Previously we showed the release of zinc from thapsigargin/IP3-sensitive endoplasmic reticulum (ER) storage in cortical neurons. In the present study, we investigated if other cellular organelles also contain free chelatable zinc and function as organelle storage for zinc. To identify free zinc within the organelles, live cells were co-stained with Zinpyr-1, a zinc fluorescent dye, and organelle-specific fluorescent dyes (MitoFluor Red 589: mitochondria; ER Tracker Red: endoplasmic reticulum; BODIPY TR ceramide: Golgi apparatus; Syto Red 64: nucleus). We examined organelles that represent potential storing sites for intracellular zinc. We showed that zinc fluorescence staining was co-localized with MitoFluor Red 589, ER Tracker Red, and BODIPY TR ceramide respectively, suggesting the presence of free zinc in mitochondria, endoplasmic reticulum, and the Golgi apparatus. On the other hand, cytosol and nucleus had nearly no detectable zinc fluorescence. It is known that nucleus contains high amount of zinc binding proteins that have high zinc binding affinity. The absence of zinc fluorescence suggests that there is little free zinc in these two regions. It also indicates that the zinc fluorescence detected in mitochondria, ER and Golgi apparatus represents free chelatable zinc. Taken together, our results support that these organelles are potential zinc storing organelles during cellular zinc homeostasis.
Keywords: Zinc, mitochondria, endoplasmic reticulum, Golgi apparatus, organelles
Introduction
Zinc is an essential element in human body regulating many physiological processes. It is involved in DNA and protein synthesis, hormone packaging, mitosis, apoptosis, and many other functions [1]. Zinc is found within the cells in both a protein-bound state and as a free or chelatable divalent ion. In the protein-bound form, zinc, as a structural and functional component, is essential for the activity of various cytosolic and nuclear enzymes [2]. The free zinc is of special interest, because it can easily exchange between different compartments of the cell and bind to numerous proteins, thereby altering their biological activity [3]. The concentration of intracellular free zinc is strictly regulated, which is known as zinc homeostasis [4]. However, the mechanisms responsible for regulating zinc homeostasis are not well established. On the other hand, abnormal zinc homeostasis has been implicated in numerous clinical manifestations, including degenerative diseases, diabetes, Crohn’s diseases, Alzheimer’s and Parkinson’s diseases, wound healing, and impaired growth and development [5-8]. Therefore, understanding how zinc homeostasis is maintained is both biological and clinical important.
The available literature suggests that the maintenance of the zinc homeostasis is achieved generally through two main mechanisms: Zinc transporting proteins and zinc-buffer system. The former contains two large metal-transporter families, zinc transporters (ZnTs) and Zrt-Irt-like proteins (ZIPs) that actively transport zinc among cytosol, discrete intracellular compartments, and extracellular medium [9]. The second important part of zinc homeostasis is so called cellular zinc storing or buffering system that regulate zinc availability and accumulation in response to metabolic demand. Perhaps the best characterized storage of zinc is the membrane-bound vesicles that are found in a large variety of cell types such as pancreatic β cells, subtype cortical neurons, prostate secretary cells, blood platelets, etc. [10-12]. They secrete zinc as an ionic intercellular signal. However, the identity of other membrane-bound organelles as zinc-storing sites is still elusive.
It has been surmised for some time that cellular membrane-bound organelles may be critical in modulating bioavailability of cellular zinc. Emerging evidence has shown that zinc transporters, both ZnTs and ZIPs, are also found on plasma membrane of cellular organelles providing molecular evidence that zinc are transported among discrete subcellular compartments [13-16]. However, there has been no direct observation of the presence of free chelatable zinc in the organelles that were in question. In this study, we measured the intracellular zinc with a fluorescent zinc indicator that detects chelatable free zinc, as fluorescent probes typically consist of a zinc-chelating unit and a fluorescent reporter. To characterize zinc staining in discrete organelles, we also stained the same cells with organelle-specific fluorescence indicators for mitochondria, endoplasmic reticulum (ER), Golgi apparatus and nucleus respectively. The presence of free zinc within an organelle was evidenced by the colocalization of zinc sensitive- and organelle sensitive-fluorescence. Our results showed that zinc fluorescence was distributed in mitochondria, ER and Golgi apparatus (but not detectible in the nucleus), suggesting that these organelles are potential sites of zinc buffers and storage of zinc homeostasis.
Methods
Cell culture
HeLa cell line and pancreatic β-cell line HIT-T15 were purchased from ATCC (Manassas, VA, USA). HeLa cells were cultured in EMEM (Gibco) supplemented with 4% FBS at 37°C in a 5% CO2, 95% air. HIT-T15 cells were maintained in RPMI 1640 (Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine serum and 5% dialyzed horse serum at 37°C in a 5% CO2, 95% air. Seven-day-old rat primary cortical neuron cells were gift from Dr. Robert Colvin (Ohio University) [17]. Cells were cultured on a petri dish with a glass bottom of 35 mm diameter (MatTek Corporation, Ashland, MA, USA).
Zinc labeling
To detect intracellular labile zinc, we used Zinpyr-1 (Sigma-Aldrich, St. Louis, MO, USA), a selective fluorescent sensor for zinc that is membrane permeable [18]. Fluorescent probes for zinc typically consist of a zinc-chelating unit and a fluorescent reporter. Zinpyr-1 has the di-2-picolylamine (DPA) ligand as the zinc-chelating unit. DPA ligand has no measurable affinity for Ca2+ or Mg2+. Zinpyr-1 easily permeates through the membrane making it an ideal candidate to study Zn2+ in live cells. Zinpyr-1 has zinc sensitivity in nM range (significantly lower than binding affinity of most proteins to zinc) and its fluorescence increases three fold upon zinc binding [18]. Cells were incubated with 2.5-10 μm Zinpyr-1 for 10-30 min at 37°C in 5% CO2. After incubation, cells were washed three times with Hank’s Balanced Salt Solution (HBSS, Life Technologies, Grand Island, NY, USA), and then visualized by fluorescence microscopy.
Mitochondria-zinc co-labeling
Labeling of mitochondria was performed by incubating cells with MitoFluor Red 589 (Molecular Probes, Eugene, OR, USA). HeLa cells and 7-day-old primary cortical neurons were incubated with 1 ml of dye solution (250 nM of MitoFluor Red 589 and 10 μM of Zinpyr-1 dissolved in HBSS) at 37°C, 5% CO2 for 20 min. After incubation, the cells were washed three times with HBSS.
ER-zinc co-labeling
We used ER Tracker Red (Molecular Probes, Eugene, OR, USA) to label the ER in HeLa cells. HeLa cells were incubated with 1 ml of dye solution (1 μM of ER Tracker Red and 10 μM of Zinpyr-1 dissolved in HBSS) at 37°C, 5% CO2 for 20 min. After incubation, the cells were washed three times with HBSS.
Golgi-zinc co-labeling
BODIPY TR Ceramide (Molecular Probes, Eugene, OR, USA) was used to visualize the Golgi apparatus. 2-day-old HeLa cells and one-day-old HIT-T15 cells were incubated with 1.5 μM BODIPY TR Ceramide at 4°C for 30 min. Cells were then washed with ice-old HBSS and incubated with 2.5 μM Zinpyr-1 for further 20 min at 37°C. Cells were washed three times before imaging.
Nucleus-zinc co-labeling
Labeling of the nucleus was performed with SYTO Red 64 (Molecular Probes, Eugene, OR, USA). HeLa cells were first incubated with 10 μM Zinpyr-1 for 10 min and then with 2.5 μM SYTO Red 64 for 10 min at 37°C, 5% CO2. The cells were washed and imaged in HBSS.
Fluorescence Imaging
Fluorescent signals were detected using a customized fluorescence microscope and Zeiss LSM 510 confocal microscope. All the images were captured using ImagePro Plus software (Media Cybernetics, Silver Spring, MD, USA) at exposures of about 1 s (For excitation: blue filter: 470/70 nm, green filter: 517/30 nm; for emission: green filter-517/30 nm, red filter-620/40 nm; Filters were purchased from Chroma). ImagePro plugins were used to enhance contrast, adjust the brightness and improve the resolution of the images. Regions of interest were marked and cropped using ImageJ software. Fluorescence intensities were calculated for unprocessed images using ImageJ. Co-localization was measured with ImagePro as Pearson’s correlation value. 3D images were collected using Zeiss LSM 510 confocal microscope (Oberkochen, Germany) with 100X magnification, and analyzed with Zeiss imaging software.
Results
We used HeLa cell line to determine and characterize the intracellular distribution of free chelatable zinc. This widely used cell line is ideal for fluorescent microscopic observation. It is well attached to the glass-bottom cell plates, big in size, and grows into a single layer with flat cell morphology. Moreover, HeLa cells have been used to study zinc response [19-21]. As shown in Figure 1, after staining with Zinpyr-1, a selective fluorescence zinc indicator, HeLa cell showed an uneven distribution of zinc-dependent fluorescence throughout the entire cell, with high amounts of free zinc detected in what looked like cellular compartments that encircle the nuclear. The staining also proceeded to cellular processes. On the other hand, cytosol and nucleus had nearly no detectable zinc fluorescence. Next, to further study zinc distribution, we stained the cells with organelle specific fluorescence indicators to determine whether free zinc was present in the membrane bound organelles, which were evidenced by the colocalization of fluorescence zinc signals and organelle specific fluorescence signals.
Figure 1.
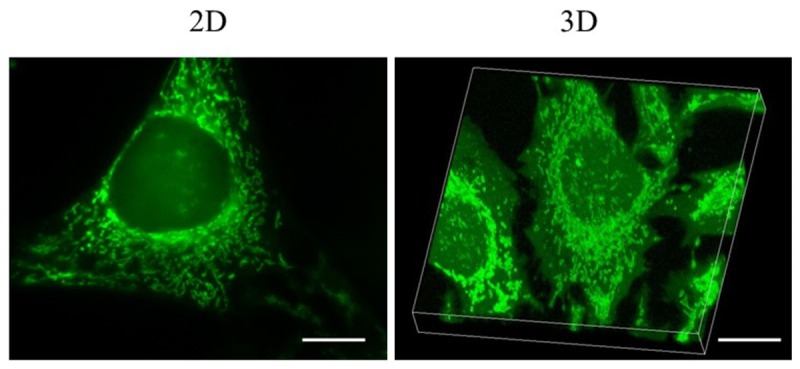
Cellular zinc distribution in HeLa cells shows compartmentalized pattern. Images of HeLa cells labeled with the intracellular fluorescent zinc indicators. Living HeLa cells were incubated with 10 mM of fluorescent zinc indicator Zinpyr-1. 2D images were captured using a customized fluorescence microscope equipped with a 100X/1.1NA objective lens. 3D images were captured using a LSM510 confocal microscope equipped with a 100X/1.3NA objective lens. Scale bars indicate 10 µm.
Zinc in mitochondria
First, we assessed whether mitochondria was one of the storage sites for intracellular free zinc. HeLa cells were co-stained with Zinpyr-1 and MitoFluor Red 589 for 20 min. Mitochondria in HeLa cells resemble squiggly lines occupying the major portion of cytosol. The red fluorescence from MitoFluor Red 589 highly co-localized with the green fluorescence from Zinpyr-1. This suggests that mitochondria are involved in the storage of intracellular free zinc (Figure 2A). HeLa cells contain more than thousands of mitochondria which, in live imaging, tend to appear as mitochondrial clustering. Therefore, while HeLa cells can provide an overall picture for the co-localization of mitochondria and zinc, they cannot be used to visualize individual mitochondria.
Figure 2.
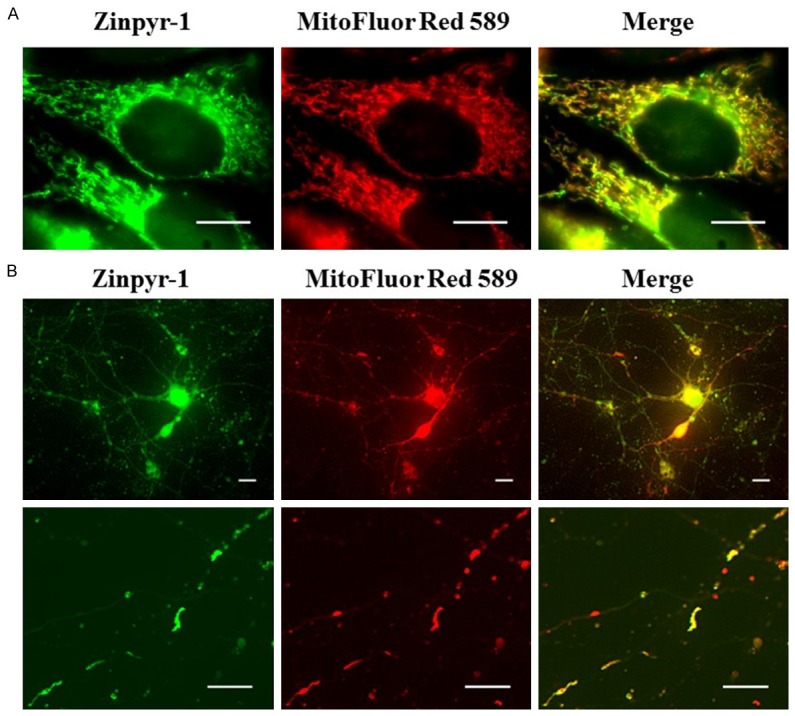
Zinc is localized to mitochondria in HeLa cells and cortical neurons. Images of HeLa cells or cortical neurons co-labeled with Zinpyr-1and a live cell marker for the mitochondria. A. Zinpyr-1 co-localizes with the mitochondria marker MitoFluor Red 589 in HeLa cells. B. Zinpyr-1 co-localizes with MitoFluor Red 589 in rat primary cultured cortical neurons. HeLa cells or rat primary cortical neurons were dual-stained with 15 mM Zinpyr-1 and 250 nM MitoFluor Red 589. Scale bars indicate 10 µm.
To visualize individual mitochondria, neuronal cultures were used because neuronal axons are typically 1-3 mM in diameter and are likely to house individual mitochondrion. Therefore, we visualized axonal mitochondria to assess its co-localization with zinc. Rat primary cortical neurons were cultured in vitro for seven days before being subjected to fluorescence staining. Fluorescence signal from Zinpyr-1 and MitoFluor Red 589 were detected from soma, axons and dendrites. Considering how small the neuronal processes were, stained neurons had to be exposed for longer time in order to capture the fluorescence signal from axons, this lead to over-saturation of fluorescence signal in soma (Figure 2B left top). A single axon was imaged under higher magnification. The axon contained a linear array of mitochondria (Figure 2B). These mitochondria differed in size, some resembled squiggly lines, while others were dot-like, reflects the morphological heterogeneity of mitochondria. As expected, mitochondria were highly co-localized with intracellular free zinc in the axon. Both the HeLa cells and neuronal axon mitochondria showed co-localization with free zinc, suggesting that mitochondria were zinc storing organelles containing free chelatable zinc.
Zinc in ER
To determine whether ER was one of the storage sites for zinc in HeLa cells, fresh cells were dual-labeled with Zinpyr-1 and ER tracker Red, an ER specific fluorescent probe. Images captured by fluorescence microscopy showed significant colocalization of zinc indicator and the ER Tracker Red, indicating that free zinc were located within the lumen of the ER (Figure 3). These results were consistent with our previous study [22] in which we showed that zinc was released from thapsigargin-sensitive and IP3R-mediated stores of the ER.
Figure 3.
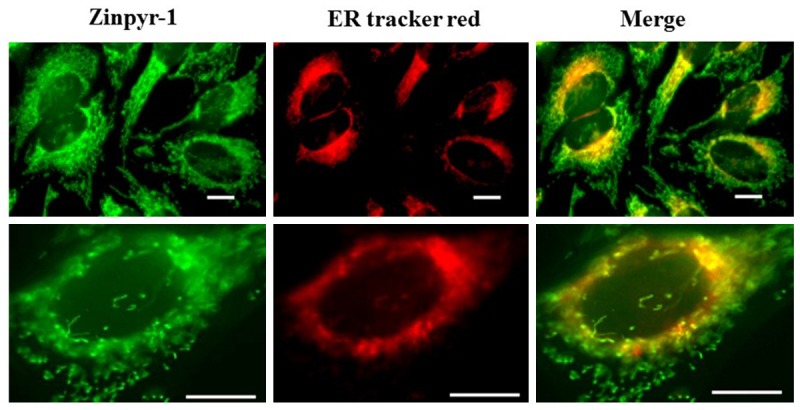
Zinc is localized to ER in HeLa cells. Images of HeLa cells co-labeled with Zinpyr-1 and ER Tracker Red, a marker for the ER. HeLa cells were double-stained with 10 mM Zinpyr-1 and 1 mM ER Tracker Red at 37°C for 20 min. Images were captured using a 40X objective (top panel) or 100X objective (bottom panel). Scale bars indicate 10 µm.
Zinc in Golgi apparatus
The Golgi apparatus is usually located near the nucleus. Here, we probed the Golgi apparatus using BODIPY TR ceramide. In HeLa cells, dual-staining with Zinpyr-1 and BODIPY TR ceramide showed that the signals from the Golgi dye co-localized with the perinuclear fluorescence of zinc dye, indicating that free zinc is localized to the Golgi apparatus (Figure 4A). As the main function of the Golgi apparatus is to process and package macromolecules such as proteins and lipids after their synthesis, it is especially prominent in cells involved in secretion, such as pancreatic cells. We used the pancreatic ß cell line, HIT-T15 to evaluate further the presence of zinc in the Golgi apparatus as a case of positive control. The Golgi apparatus in pancreatic β-cells is one of the zinc sources, because the insulin packaging and secretion requires a high amount of free zinc [23,24]. Here the HIT-T15 cells were double stained with BODIPY TR ceramide and Zinpyr-1. Bright Zinpyr-1 fluorescence emanating from the perinuclear zones, indicating the presence of high concentration of free zinc in the regions. BODIPY TR ceramide stained the same perinuclear zones, which proved that the co-localization of zinc with the Golgi apparatus (Figure 4B).
Figure 4.
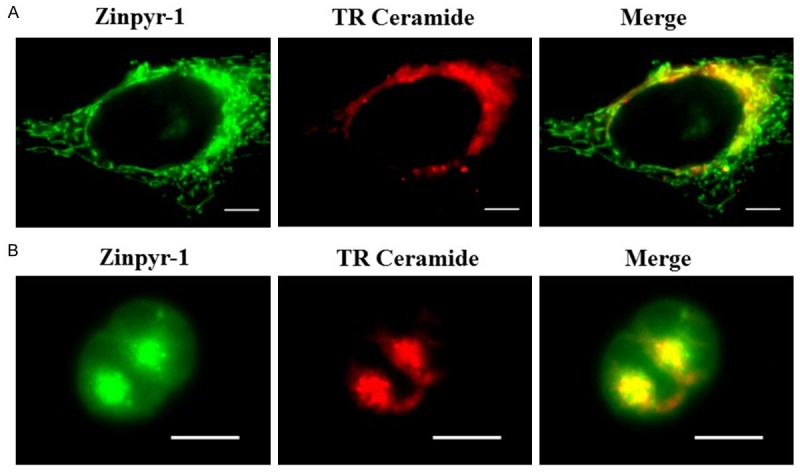
Zinc is localized to the Golgi apparatus in HeLa cells and HIT T15 cells. Images of HeLa cells or HIT T15 cells co-labeled with Zinpyr-1 and a live cell marker for the Golgi apparatus. A. Zinpyr-1 co-localizes with BODIPY TR ceramide, the marker for the Golgi apparatus, in HeLa cells. B. Zinpyr-1 co-localizes with BODIPY TR ceramide in pancreatic ß cells HIT T15. HeLa cells or HIT T15 cells were stained with 5 mM TR ceramide for 30 min at 4°C, then stained with 2.5 mM Zinpyr-1 for 30 min at 37°C. Scale bars indicate 10 µm.
Zinc was not detectable in the nucleus
Syto Red 64 was used to identify nucleus in HeLa cells because it reliably labeled nucleus in live cells. As shown in Figure 5, Syto Red 64 labeled HeLa cell nuclei with red fluorescence. The same cells were also stained with Zinpyr-1 but zinc fluorescence was not observed in the nucleus of HeLa cells. Nucleus is known for its packaging of many zinc binding proteins or zinc binding motif for hundreds transcription factors [25]. However, it didn’t surprise that zinc was not detectable within the nucleus using a fluorescence indicator. This was probably because most of the nuclear zinc was bound to nuclear proteins in a healthy cell and the concentration of free chelatable zinc was negligible.
Figure 5.
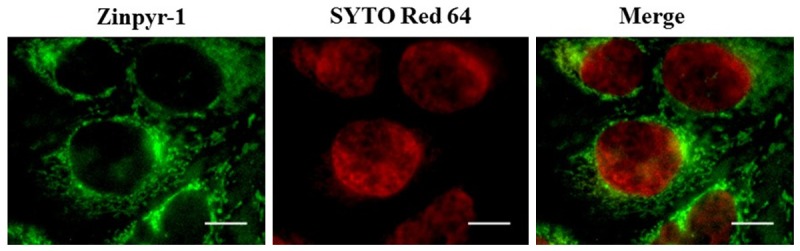
Absence of zinc staining in the nucleus. Images of HeLa co-labeled with Zinpyr-1 and Syto Red 64, a live cell marker for the nucleus. HeLa cells were stained with 10 mM Zinpyr-1 for 10 min at 37°C, and then stained with 2.5 mM SYTO Red 64 for 10 min at 37°C. Scale bars indicate 10 µm.
Discussion
In this study, we demonstrate that zinc is present within mitochondria, ER and Golgi apparatuses (Figures 2, 3 and 4), providing the direct evidence that these organelles serve as candidates of intracellular zinc storages. We also show that zinc is not detectable in the nucleus (Figure 5). It is due to that most of the zinc is more tightly bound to nuclear proteins, and is unavailable for chelation by fluorescent zinc indicator. Therefore, the absence of zinc fluorescence in nucleus supports that zinc fluorescence detected in other organelles represents the free chelatable zinc. Taken together, our results indicate that cytosolic membrane-bound organelles such as mitochondria, ER, and Golgi apparatus contain free chelatable zinc.
While the mechanisms responsible for regulating zinc homeostasis are not well established, available data support that intracellular zinc levels are determined by the interaction of membrane zinc transporters and cytoplasmic zinc buffers [9,10]. It has been speculated that zinc is stored within intracellular organelles, however, there are few comprehensive imaging data available. Our previous study has grazed that idea when the co-localization of signals from ER Tracker Red and Zinpyr-1 was demonstrated while probing into IP3-sensitive zinc stores within cortical neurons. Since then, it has remained as the only dye-based imaging evidence for the presence of zinc within ER (Stork et al. 2010). The method used in that study served as a proof-of-principle for us to pair Zinpyr-1 with other organelle-specific dyes to probe potential intracellular Zinc stores, including ER. Results from present study support our previous observation. Zinc transporters (ZnT) ZnT5/ZnT6 hetero-oligomeric complexes are involved in ER homeostasis by transporting zinc in biological processes [15,16], and may function as bidirectional transporters [10,13,14]. These studies suggest that zinc is required for correct ER functions. Within the ER, zinc is buffered by the abundant luminal resident chaperone protein calreticulin that has multiple zinc binding sites [26].
Several studies suggest that zinc may be sequestered in mitochondria through the activation of a cation-permeable channel, the mitochondrial calcium uniporter or other unidentified-independent pathway [27-29]. Mitochondrial zinc uptake functions for an organelle storage of zinc that may provide clearance of cytosolic zinc accumulation. The latter is especially important in neurons undergoing cytotoxicity [3,30,31]. Golgi apparatus has long been speculated to pack zinc into secretory granules such as in pancreatic β-cell where zinc is co-released with insulin [24]. However, there is no direct fluorescence image of zinc, as shown in this study, that shows free chelatable zinc are presented in the Golgi apparatus. No previous evidence shows the colocalization of zinc and Golgi apparatus in none-secretary cells. Recent studies demonstrated that zinc transporters (ZnTs and ZIPs) expressed on the membrane of Golgi apparatus may be involved in Golgi homeostasis of zinc. For example, ZIP7 is a functional Zn transporter that may participate in the transport of the cytoplasmic zinc into the Golgi apparatus (Chi et al. 2006).
In our current study, zinc is considerably lower in cytosol than in the organelles, possibly, with the exception of nucleus where there is little free Zinc. Previously, our work has shown the appearance of labile zinc within the nucleus under cytotoxic conditions that promote cell death confirming the viability of the cells in our study (Stork et al. 2011). The other potential intracellular store for Zinc could be the lysosome. The lysosome’s acidity could help retain zinc in its free ionic form within its lumen [32]. In conclusion, we have used a comprehensive approach where we took all the major organelles into consideration to collect the required data on subcellular compartments of zinc. It is also necessary to quantitatively estimate zinc within various organelles and to track the change in zinc distribution during a disruption of cellular physiology.
Acknowledgements
We thank Dr. Calvin James for providing us HeLa cells and HIT-T15 cells, thanks to Dr. Robert Colvin for providing us cortical neurons. We also thank the Imaging Facility of the Ohio University Neuroscience Program for their support. We acknowledge the expert assistance of Zihui Wang and Cheng Qian.
Disclosure of conflict of interest
None.
References
- 1.Beyersmann D, Haase H. Functions of zinc in signaling, proliferation and differentiation of mammalian cells. Biometals. 2001;14:331–41. doi: 10.1023/a:1012905406548. [DOI] [PubMed] [Google Scholar]
- 2.Berg JM, Godwin HA. Lessons from Zinc-Binding Peptides. Annu Rev Biophys Biomol Struct. 1997;26:357–71. doi: 10.1146/annurev.biophys.26.1.357. [DOI] [PubMed] [Google Scholar]
- 3.Frederickson CJ, Koh JY, Bush AI. The neurobiology of zinc in health and disease. Nat Rev Neurosci. 2005;6:449–62. doi: 10.1038/nrn1671. [DOI] [PubMed] [Google Scholar]
- 4.Murakami M, Hirano T. Intracellular zinc homeostasis and zinc signaling. Cancer Sci. 2008;99:1515–22. doi: 10.1111/j.1349-7006.2008.00854.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kinlaw WB, Levine AS, Morley JE, Silvis SE, McClain CJ. Abnormal zinc metabolism in type II diabetes mellitus. Am J Med. 1983;75:273–7. doi: 10.1016/0002-9343(83)91205-6. [DOI] [PubMed] [Google Scholar]
- 6.McClain C, Soutor C, Zieve L. Zinc deficiency: a complication of Crohn’s disease. Gastroenterology. 1980;78:272–9. [PubMed] [Google Scholar]
- 7.Religa D, Strozyk D, Cherny RA, Volitakis I, Haroutunian V, Winblad B, Naslund J, Bush AI. Elevated cortical zinc in Alzheimer disease. Neurology. 2006;67:69–75. doi: 10.1212/01.wnl.0000223644.08653.b5. [DOI] [PubMed] [Google Scholar]
- 8.Rossi L, Migliaccio S, Corsi A, Marzia M, Bianco P, Teti A, Gambelli L, Cianfarani S, Paoletti F, Branca F. Reduced Growth and Skeletal Changes in Zinc-Deficient Growing Rats Are Due to Impaired Growth Plate Activity and Inanition. J Nutr. 2001;131:1142–6. doi: 10.1093/jn/131.4.1142. [DOI] [PubMed] [Google Scholar]
- 9.Cousins RJ, Liuzzi JP, Lichten LA. Mammalian Zinc Transport, Trafficking, and Signals. J Biol Chem. 2006;281:24085–9. doi: 10.1074/jbc.R600011200. [DOI] [PubMed] [Google Scholar]
- 10.Eide DJ. Zinc transporters and the cellular trafficking of zinc. Biochim Biophys Acta BBA - Mol Cell Res. 2006;1763:711–22. doi: 10.1016/j.bbamcr.2006.03.005. [DOI] [PubMed] [Google Scholar]
- 11.Marx G, Korner G, Mou X, Gorodetsky R. Packaging zinc, fibrinogen, and factor XIII in platelet α-granules. J Cell Physiol. 1993;156:437–42. doi: 10.1002/jcp.1041560302. [DOI] [PubMed] [Google Scholar]
- 12.Kelleher SL, McCormick NH, Velasquez V, Lopez V. Zinc in Specialized Secretory Tissues: Roles in the Pancreas, Prostate, and Mammary Gland. Adv Nutr Int Rev J. 2011;2:101–11. doi: 10.3945/an.110.000232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ellis CD, MacDiarmid CW, Eide DJ. Heteromeric Protein Complexes Mediate Zinc Transport into the Secretory Pathway of Eukaryotic Cells. J Biol Chem. 2005;280:28811–8. doi: 10.1074/jbc.M505500200. [DOI] [PubMed] [Google Scholar]
- 14.Ellis CD, Wang F, MacDiarmid CW, Clark S, Lyons T, Eide DJ. Zinc and the Msc2 zinc transporter protein are required for endoplasmic reticulum function. J Cell Biol. 2004;166:325–35. doi: 10.1083/jcb.200401157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ishihara K, Yamazaki T, Ishida Y, Suzuki T, Oda K, Nagao M, Yamaguchi-Iwai Y, Kambe T. Zinc Transport Complexes Contribute to the Homeostatic Maintenance of Secretory Pathway Function in Vertebrate Cells. J Biol Chem. 2006;281:17743–50. doi: 10.1074/jbc.M602470200. [DOI] [PubMed] [Google Scholar]
- 16.Suzuki T, Ishihara K, Migaki H, Ishihara K, Nagao M, Yamaguchi-Iwai Y, Kambe T. Two Different Zinc Transport Complexes of Cation Diffusion Facilitator Proteins Localized in the Secretory Pathway Operate to Activate Alkaline Phosphatases in Vertebrate Cells. J Biol Chem. 2005;280:30956–62. doi: 10.1074/jbc.M506902200. [DOI] [PubMed] [Google Scholar]
- 17.Colvin RA, Lai B, Holmes WR, Lee D. Understanding metal homeostasis in primary cultured neurons. Studies using single neuron subcellular and quantitative. Metallomics. 2015;7:1111–23. doi: 10.1039/c5mt00084j. [DOI] [PubMed] [Google Scholar]
- 18.Walkup GK, Burdette SC, Lippard SJ, Tsien RY. A New Cell-Permeable Fluorescent Probe for Zn2+ J Am Chem Soc. 2000;122:5644–5. [Google Scholar]
- 19.Karin M, Herschman HR. Characterization of the Metallothioneins Induced in HeLa Cells by Dexamethasone and Zinc. Eur J Biochem. 1980;107:395–401. doi: 10.1111/j.1432-1033.1980.tb06042.x. [DOI] [PubMed] [Google Scholar]
- 20.Karin M, Andersen RD, Slater E, Smith K, Herschman HR. Metallothionein mRNA induction in HeLa cells in response to zinc or dexamethasone is a primary induction response. Nature. 1980;286:295–7. doi: 10.1038/286295a0. [DOI] [PubMed] [Google Scholar]
- 21.Danscher G. The autometallographic zinc-sulphide method. A new approach involvingin vivo creation of nanometer-sized zinc sulphide crystal lattices in zinc-enriched synaptic and secretory vesicles. Histochem J. 1996;28:361–73. doi: 10.1007/BF02331399. [DOI] [PubMed] [Google Scholar]
- 22.Stork CJ, Li YV. Zinc release from thapsigargin/IP3-sensitive stores in cultured cortical neurons. J Mol Signal. 2010;5:5. doi: 10.1186/1750-2187-5-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Slepchenko KG, James CBL, Li YV. Inhibitory effect of zinc on glucose-stimulated zinc/insulin secretion in an insulin-secreting β-cell line. Exp Physiol. 2013;98:1301–11. doi: 10.1113/expphysiol.2013.072348. [DOI] [PubMed] [Google Scholar]
- 24.Li YV. Zinc and insulin in pancreatic beta-cells. Endocrine. 2013;45:178–89. doi: 10.1007/s12020-013-0032-x. [DOI] [PubMed] [Google Scholar]
- 25.Maret W. New perspectives of zinc coordination environments in proteins. J Inorg Biochem. 2012;111:110–6. doi: 10.1016/j.jinorgbio.2011.11.018. [DOI] [PubMed] [Google Scholar]
- 26.Tan Y, Chen M, Li Z, Mabuchi K, Bouvier M. The calcium- and zinc-responsive regions of calreticulin reside strictly in the N-/C-domain. Biochim Biophys Acta. 2006;1760:745–53. doi: 10.1016/j.bbagen.2006.02.003. [DOI] [PubMed] [Google Scholar]
- 27.Csordás G, Hajnóczky G. Plasticity of Mitochondrial Calcium Signaling. J Biol Chem. 2003;278:42273–82. doi: 10.1074/jbc.M305248200. [DOI] [PubMed] [Google Scholar]
- 28.Gazaryan IG, Krasinskaya IP, Kristal BS, Brown AM. Zinc Irreversibly Damages Major Enzymes of Energy Production and Antioxidant Defense Prior to Mitochondrial Permeability Transition. J Biol Chem. 2007;282:24373–80. doi: 10.1074/jbc.M611376200. [DOI] [PubMed] [Google Scholar]
- 29.Jiang D, Sullivan PG, Sensi SL, Steward O, Weiss JH. Zn2+ Induces Permeability Transition Pore Opening and Release of Pro-apoptotic Peptides from Neuronal Mitochondria. J Biol Chem. 2001;276:47524–9. doi: 10.1074/jbc.M108834200. [DOI] [PubMed] [Google Scholar]
- 30.Medvedeva YV, Lin B, Shuttleworth CW, Weiss JH. Intracellular Zn2+ accumulation contributes to synaptic failure, mitochondrial depolarization and cell death in an acute slice oxygen glucose deprivation (OGD) model of ischemia. J Neurosci Off J Soc Neurosci. 2009;29:1105–14. doi: 10.1523/JNEUROSCI.4604-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sensi SL, Paoletti P, Bush AI, Sekler I. Zinc in the physiology and pathology of the CNS. Nat Rev Neurosci. 2009;10:780–91. doi: 10.1038/nrn2734. [DOI] [PubMed] [Google Scholar]
- 32.Roh HC, Collier S, Guthrie J, Robertson JD, Kornfeld K. Lysosome-related organelles in intestinal cells are a zinc storage site in C. elegans. Cell Metab. 2012;15:88–99. doi: 10.1016/j.cmet.2011.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]