

Abstract
Spinal cord injury (SCI) causes loss of neurological function and, depending upon the severity of injury, may lead to paralysis. Currently, no FDA approved pharmacotherapy is available for SCI. High-dose methylprednisolone is widely used, but this treatment is controversial. We have previously shown that low doses of estrogen treatment reduced inflammation, attenuated cell death, and protected axon and myelin in SCI rats, but its effectiveness in recovery of function is not known. Therefore, the goal of the current study was to investigate whether low doses of estrogen treatment post-SCI would reduce inflammation, protect cells and axons, and improve locomotor function during the chronic phase of injury. Injury (40 g•cm force) was induced at T10 in young adult male rats. Rats were treated with 10 or 100 μg 17β-estradiol for 7 days following injury and compared with vehicle treated injury and laminectomy (sham) controls. Histology (H&E), immunohistofluorescence, Doppler laser technique and Western blotting were used to monitor tissue integrity, gliosis, blood flow, angiogenesis, expression of angiogenic factors, axonal degeneration and locomotor function (BBB rating) following injury. To assess the progression of recovery, rats were sacrificed at 7, 14, or 42 days post-injury. A reduction in glial reactivity, attenuation of axonal and myelin damage, protection of cells, increased expression of angiogenic factors and microvessel growth, and improved locomotor function were found following estrogen treatment compared to vehicle treated injured rats. These results suggest that treatment with a very low dose of estrogen has significant therapeutic implications for the improvement of locomotor function in chronic SCI.
Keywords: Angiogenesis, axonal protection, chronic spinal cord injury, locomotor function, low dose estrogen, neuroprotection
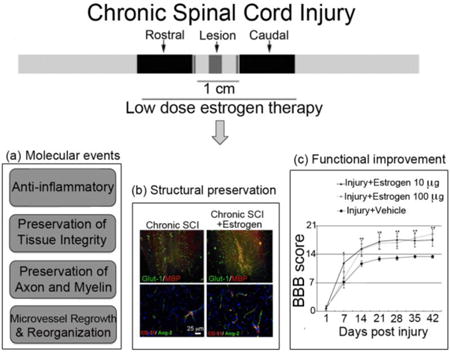
Graphical abstract
Experimental studies with low dose estrogen therapy following chronic spinal cord injury (SCI) demonstrated the potential for multi-active beneficial outcomes that could ameliorate the degenerative pathways in chronic SCI as shown in (a). Furthermore, low dose estrogen treatment resulted in the protection and preservation of cells and promotion of microvessel growth in the spinal cord, as shown in (b), and led to locomotor functional recovery as shown in (c).
Introduction
While significant advances have been made in surgical procedures for the treatment of spinal cord injury (SCI) over the years, this research has also aimed to understand the molecular mechanisms involved in the pathophysiology of SCI. In spite of these advances, no effective pharmacotherapy is currently available that will improve or restore function following SCI (Cox et al. 2015a). Thus, development of new therapeutics with maximal capacity that can improve locomotor and sensory function after SCI is critical.
SCI is a devastating problem associated with neurological and other functional deficits. Depending on the severity of injury, SCI leads to a debilitating condition, including loss of movement and sensation, often leading to paralysis and reduced longevity (Cox et al. 2015a, Varma et al. 2010, Krause & Newman 2009). Following SCI, the neurological dysfunction occurs from two very closely associated injury phases – primary and secondary. The primary injury at the initial impact (lesion) site occurs rapidly, causing irreversible necrotic cell death. In contrast, the secondary injury in the rostral and caudal penumbra adjacent to or surrounding the initial impact site develops over hours, days, and weeks involving multiple mechanisms, which include inflammation and infiltration of inflammatory cells, activation of astrocytes and microglia, and ischemia due to lack of blood supply. These different mechanisms, as well as the release of detrimental factors from activated glial and inflammatory cells, damage neurons, axons, oligodendrocytes and myelin, ultimately destroying tissue; this pathophysiological process may be targeted for therapeutic intervention (Varma et al. 2013). The multi-destructive pathways involved in the secondary injury process include ischemia due to disruption of blood vessels, oxidative damage, inflammation, glutamate excitotoxicity, calcium (Ca2+) overload, and activation of enzymes, phospholipase and proteases, including calpain and caspases, which leads to progression of neuron and oligodendrocyte death in the spinal cord (Agrawal & Fehlings 1996, Stokes et al. 1983). To overcome these destructive processes and ameliorate dysfunction following SCI with pharmacotherapeutic intervention is an enormous task. In addition, the formation of a glial scar and cavity at the lesion (injury site), which are not penetrable and block the regeneration or repair of damaged axons and myelination, presents a major obstacle and results in neurological deficits.
Because of the complex pathophysiology of SCI, developing a therapeutic strategy that can ameliorate the multi-destructive pathways involved in tissue damage has been difficult. No FDA-approved therapy is currently available for SCI. Current therapies, including widely used methylprednisolone (MP), have limited efficacy and do not restore function (Bracken et al. 1998, Hurlbert 2000). In addition, MP has many side effects and its use is controversial (Pointillart et al. 2000). To this end, investigations have recently demonstrated neuroprotective effects of steroid hormones (17β-estradiol, progesterone) in animal models of acute and chronic SCI (Elkabes & Nicot 2014, Sribnick et al. 2005, Sribnick et al. 2010, Yune et al. 2004). Although administration of non-physiological high doses of steroids, e.g. estrogen, were used in these investigations, the results indicated significant reduction of secondary injury parameters, including glial cell activation, inflammation, oxidative damage, and modulation of L-type Ca2+ channels and Na+-Ca2+ exchanger (Sribnick et al. 2005, Sribnick et al. 2010, Yune et al. 2004), while promoting angiogenesis with concomitant protection of neurons and axons and restoration of function following injury (Sribnick et al. 2010, Yune et al. 2004). Since the high doses of estrogen (100 μg-4 mg/kg) cannot be used for treatment of human SCI, we have previously shown that very low doses of estrogen (1-10 μg/kg) significantly reduced glial activation and inflammation and promoted neuroprotection in acute SCI in animals (Samantaray et al. 2011).
The current studies on the efficacy of low dose estrogen (10 μg/kg) in a chronic SCI rat model assessing recovery of function were investigated on the basis of findings described in more detail in a preceding manuscript (Samantaray et al. 2015). The estrogen-mediated attenuation of many different destructive parameters that are involved in the degenerative process in acute injury suggests that its efficacy may be tested in the chronic, long term SCI. Thus, the present report is focused on examination of the effect of low dose estrogen in recovery of locomotor function following SCI. This is essential to investigate the potential of using estrogen in SCI for clinical studies/trials. In support of estrogen as a potential therapy for SCI, we report that estrogen administration at low dose (10 μg/kg) attenuated inflammation and calpain activation; upregulated estrogen receptors, angiogenic factors, and microvessel growth; preserved axons and protected neurons; and, importantly, restored locomotor function following SCI. These pre-clinical findings may help develop appropriate therapeutic studies for using estrogen in brain and spinal cord injuries and associated neurodegenerative diseases.
Materials and Methods
Induction of SCI
Male Sprague-Dawley (Harlan Laboratories, Dublin, VA, USA) rats weighing 200-250 g were used throughout the studies. Rats were anaesthetized with ketamine (Ketaset, 80 mg/kg Pfizer, New York, NY, USA) and xylazine (AnaSed, 10 mg/kg, Lloyd Laboratories, Shenandoah, IA, USA) and a laminectomy was performed at T10. After immobilizing the spine with a spinal stereotactic device, a moderately severe SCI was induced by a modified method (Perot et al. 1987), dropping a constant weight (5 g) from a height of 8 cm onto an impounder (0.3 cm in diameter) gently placed on the spinal cord. Sham animals received laminectomy only, and buprenorphine (0.01-0.05 mg/kg) was administered for post-operative pain. A total of 90 rats were used in this study, which includes n = 3 for sham + vehicle or sham + estrogen and n = 6 for injury + vehicle or injury + estrogen (10 μg) or injury + estrogen (100 μg) sacrificed at 3 time points (7, 14, and 28 days) as well as an additional set of rats used for local spinal blood flow as specified time intervals following SCI. The rats were sacrificed by decapitation under anesthesia either 7 days, 14 days or 6 weeks post-SCI. A 1 cm long section of the spinal cord centered on the lesion site as well as 1 cm long rostral section (starting 0.5 cm rostral from the impact site) and a 1 cm long caudal penumbra section (starting 0.5 cm caudal to the impact site) were taken for all groups so that the location and size remained the same for the quantitative analysis, i.e., to determine the effects of estrogen on the pathophysiological changes. ARRIVE experimental guidelines were followed along with institutional approval (AR#2079, Medical University of South Carolina) for care and handling of experimental animals during the course of this study.
Estrogen treatment in chronic SCI rats
Water-soluble estrogen, in the form of 17β-estradiol (10 or 100 μg/kg), was dissolved in saline, and vehicle treated rats received an equivalent volume of saline. All rats received a bolus tail vein injection of low dose estrogen or vehicle at 15 minutes post-injury to facilitate a rise of estrogen in the blood to levels high enough for therapeutic efficacy. Estrogen (10 or 100 μg/kg/day) levels were maintained by daily injections for 1 week post-SCI. Motor function was examined at 24 hours after surgery and weekly thereafter for 6 weeks following a controlled blinded protocol.
Assessment of locomotor function (BBB scoring)
Animals were graded based on the BBB scale (Basso et al. 1995) 24 hours after surgery and weekly thereafter for 6 weeks, as previously reported (Sribnick et al. 2010). These outcome measurements were supplemented by histochemical and biochemical endpoints. Trained personnel performed the blinded BBB assessment.
Western blot analysis of angiogenesis markers
Approximately 50-100 mg samples of spinal cords were homogenized in 50 mM Tris buffer (pH 7.4) containing 5 mM EGTA and 1 mM PMSF. Proteins were separated on a 4-20% linear gradient SDS-PAGE. Proteins were transferred onto nitrocellulose membranes for analysis and specific proteins were detected using an enhanced chemiluminescence (ECL) system (Life Technologies, NY). Primary antibodies used for Western blot analysis were anti-Angpo-2, and anti-tie-2 antibodies (Santa Cruz Biotechnology, Santa Cruz, CA, USA) at 1:250 dilutions in blocking buffer to identify increases in angiogenic markers. Protein bands were quantified using an Alpha Innotech FluorChem FC2. The amount of protein was calculated as percent compared to sham vehicle ± SEM.
Histology and immunofluorescent staining
H & E staining was performed on paraffin embedded longitudinal sections to assess the cavity volumes on days 7 or 14 post-SCI rat spinal cord according to the method previously described (Sribnick et al. 2010).
Spinal cords were embedded in Tissue-Tek OCT freezing media, cut into 5 μm cross sections, and fixed with 95% ethanol for 10 minutes. After blocking sections were incubated with primary antibody overnight, they were rinsed in PBS and incubated with a mix of the appropriate secondary antibodies. Sections were then rinsed, mounted with VectaShield mounting media, and visualized as previously described (Samantaray et al. 2011). Immunofluorescence studies included identification of cells with specific marker antibodies (Chemicon) for astrocytes (GFAP, 1:400) or macrophages/microglia (OX-42, 1:100). Axonal damage was detected using 1:1000 dilution of de-NFP (SMI 311, BioLegend) or 1:400 of MBP (Santa Cruz). Immunofluorescent methods of determining microvessel growth and angiogenic markers were employed as follows. Spinal cord sections were cut into 10 μm cross sections at –20°C on a cryostat, air-dried, and fixed with 10% methanol-free formaldehyde for 10 minutes. For antigen retrieval, sections were processed in an ethanol:acetic acid (2:1) solution at -20°C for 10 minutes. After washing with PBS, the sections were treated for 5 minutes in 0.2% Triton-X in distilled H2O, washed in PBS, and blocked for 1 h at room temperature in 1.0% cold water fish gelatin (Sigma) to reduce non-specific staining. Rabbit anti-GLUT-1 (1:1000, Chemicon) antibody specific for a glucose transporter found on vascular endothelium was used to stain for endothelial cells. Rabbit antibodies to Tie-2, bFGF, and Angpo-2 (1:200, Santa Cruz), or VEGF (1:400, Chemicon) were used to label angiogenic factors. Primary antibodies were removed and sections were washed with PBS. Sections were then incubated with secondary antibodies containing anti-rabbit IgG-Alexa-Flour 488.
Quantification with circular probes
In order to quantify Glut-1 IR, an alternate method using unbiased circular probes was used. Immunohistofluorescent Glut-1 images were overlaid on top of a frame with randomly placed circles of uniform diameter. In each frame, the number of times the Glut-1 stained vessel intersected the circumference of the circles were quantified. At least 3 independent images were quantified for each rat spinal cord, and the average was calculated within each experimental group. The averages were compared using ANOVA (p < 0.05 or 0.01).
Laser doppler flowmetry
Laser-scanning imaging was used to measure local spinal blood flow (LSBF) immediately before, immediately after, and 3hrs post-SCI in accordance with a previously published method (Li et al. 2007). Laser-scanning imaging measurements and analysis were performed using the PeriScan® system and LDPIwin 2® (Perimed AB, Stockholm, Sweden). In laser-scanning imaging, the ‘single mode’ with medium resolution was used to scan the photo image of LSBF by pointing the laser beam directly at the center of the impact site and scanning a 30 mm × 10 mm area. The distance from the spinal cord was determined by the beam intensity, which was adjusted to 7.5- 8.0 (∼6-7 cm). The conventional ‘duplex mode’ was used to record the Doppler image with the laser beam pointed to the same ischemic core.
Statistical analysis
Optical density (OD) of protein immunoreactive bands obtained from Western blotting of angiogenic markers or the immunoreactive pixels of the histofluorescence data were analyzed with NIH Image J. Results were assessed in Graphpad Prism version 6 and compared by using one-way ANOVA with Bonferroni test for multiple comparisons. Data were expressed as mean ± SEM (n = 3-6). The difference between SCI or sham spinal cords with or without estrogen treatment was considered significant at p < 0.05.
Results
Low dose estrogen therapy reduces gliosis following chronic SCI
Although inflammatory events were decreased in spinal cord tissue from estrogen treated animals following acute SCI (Sribnick et al. 2005, Xie et al. 2002, Samantaray et al. 2015), further experiments were performed to determine if microgliosis and astrogliosis remained inhibited following chronic SCI (day 42). In the current investigation, rats did not receive estrogen therapy beyond 7 days post-SCI. The extent of gliosis was examined by immunofluorescent staining using OX-42 and GFAP antibodies for microglia and astrocytes, respectively. Our results showed increased staining of microglial cells and their processes, indicating gliosis, and reactive cells. OX-42 (marker for macrophage/microglia) and GFAP (marker for astrocytes) staining were increased 3.2 fold and 2.6 fold, respectively, in lesion from SCI animals treated with vehicle alone compared to sham (control). Treatment with estrogen (10 μg/kg) significantly increased immunoreactive gliosis, which was markedly attenuated in the lesion of vehicle treated SCI rats (Fig. 1b) and suggested that the inflammatory response elicited by reactive gliosis could be controlled by low dose estrogen therapy (Fig. 1).
Fig. 1.
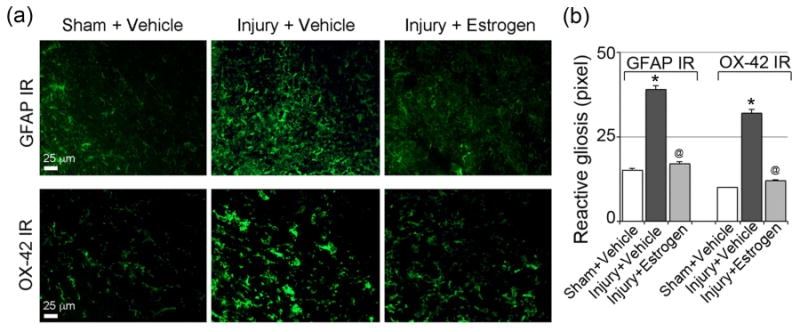
Low dose estrogen therapy reduces inflammatory events following chronic SCI. (a) Thin frozen cross sections (5 μm) from the lesion and caudal penumbra spinal cord tissues (6 weeks-post SCI) were stained with GFAP antibody as a marker for astrogliosis and OX-42 antibody as a marker of microgliosis. Representative images from lesion spinal cord are presented (200× magnification). Caudal penumbra images were omitted because there were no significant differences among the different groups at 6 weeks post-SCI in GFAP IR (immunoreactivity). In lesion spinal cord, GFAP IR was significantly higher in chronic SCI rats (day 42), which was significantly attenuated in estrogen treated group. OX-42 IR was also significantly higher in the lesion spinal cord of chronic SCI rats (day 42) compared to sham. Restored microglial morphology, similar to that seen in sham, was noted in SCI rat spinal cords treated with estrogen (10 μg/kg), (n > 3). (b) Quantification of the GFAP IR and OX-42 IR clearly indicated significant reactive gliosis (*P < 0.05 compared to control) in chronic SCI cord; 10 μg estrogen therapy attenuated this effect (@P < 0.05 compared to injury vehicle).
Low dose estrogen therapy preserves tissue integrity following chronic SCI
In order to examine the preservation of tissue and characterize protection of axons and myelin following estrogen therapy in chronic SCI rats, H&E staining was performed on spinal cord tissue from control and vehicle or estrogen treated chronic SCI rats. Visible differences in the diameter and axonal density of the spinal cord between estrogen treated (10 μg/kg) and vehicle treated SCI rats were apparent on days 7 and 14 post-SCI (Fig. 2). Edema and inflammation were reduced and markedly less tissue damage was seen in estrogen treated rats when compared with the vehicle treated rats. These findings suggest a pivotal role for estrogen in preserving tissue and myelinated axons for functional recovery following SCI (Fig. 2).
Fig. 2.
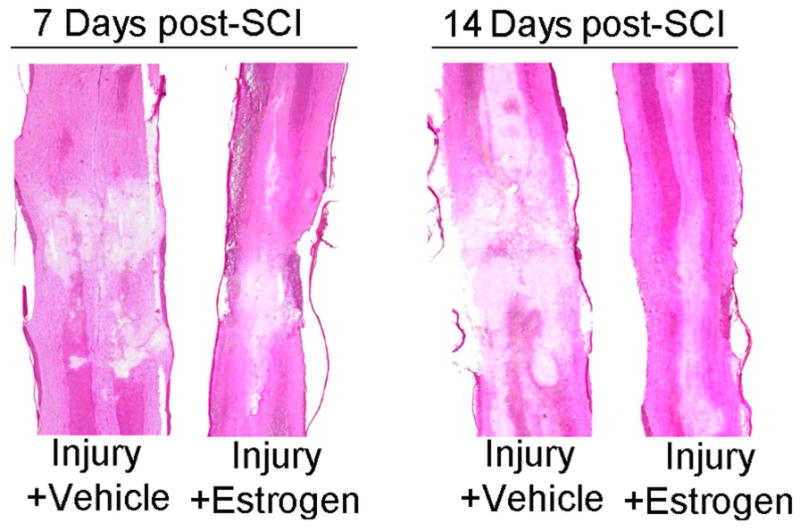
Low dose estrogen therapy preserves tissue integrity following chronic SCI. Thin paraffin embedded sections of SCI (7 and 14 days post-SCI) rat spinal cord were H&E stained and the vehicle treated and the estrogen (10 μg/kg) treated groups were compared. At 14 days post SCI, rat spinal cords show greater loss of tissue integrity than at 7 days post-SCI, presumably due to the secondary injury, and estrogen therapy (10 μg/kg), (n > 3) protected at both the time points.
Low dose estrogen therapy protects axons and reduces axonal degeneration following chronic SCI
The extent of axonal damage was analyzed in spinal cord sections from sham and SCI rats that received estrogen (10-100 μg) or vehicle (saline) using antibody against dephosphorylated neurofilament (de-NFP, marker for degenerated axons). De-NFP expression was markedly increased in both the lesion and caudal penumbra following injury in chronic spinal cord tissue of vehicle treated rats when compared with sham rats (Fig. 3a, b, c). In contrast, the deNFP staining was significantly decreased in the lesion and caudal penumbra of estrogen treated chronic SCI rats compared to vehicle. Quantification of deNFP pixels (red) showed markedly reduced axonal degeneration (>40%) in the penumbra of estrogen treated chronic rats compared with vehicle treated chronic SCI rats (Fig. 3b). The attenuating effects of estrogen in the lesion of SCI rats were also significant, indicating estrogen treatment preserved axonal integrity in chronic SCI, which would help improve the motor function.
Fig. 3.
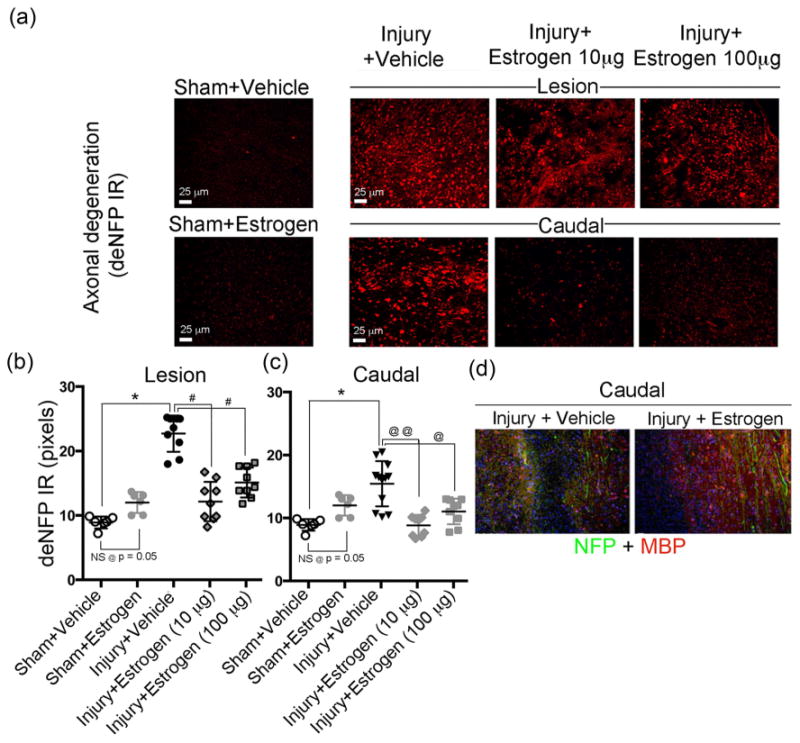
Low dose estrogen therapy reduces axonal degeneration and preserves axons following chronic SCI. Thin frozen cross sections (5 μm) were obtained from lesion and caudal penumbra spinal cord tissues (6 weeks post-SCI). Sections were stained with antibody recognizing deNFP. (a) Representative images from the four treatment groups (200× magnification). (b, c) Quantification of deNFP fluorescence for determination of axonal damage. Pixels were quantified using ImageJ. Sham + vehicle and sham + estrogen showed no significant difference at p = 0.05. Significant differences from sham values were indicated by *P < 0.001; compared to injury + vehicle (lesion) as #P < 0.01, and compared to injury + vehicle (caudal penumbra) as @P < 0.05 or @@P < 0.01. (c) Immunofluorescent staining of caudal penumbra of spinal cord tissue from SCI rats following treatment with vehicle or estrogen (10 μg) for nuclei with Hoescht stain (blue), axons with NFP antibody (green), and myelin or myelin forming cells with MBP antibody (red) (100× magnification). n = 3.
Next, the state of axons and myelin were examined in spinal cord penumbra slices. These were labeled with Hoechst staining for nuclei (blue), neurofilament protein (NFP) antibody for axons (green), and myelin basic protein (MBP) antibody for myelin/myelin forming cells (red) in vehicle and estrogen treated rats. Results from immunostained slices indicated substantially less MBP-positive cells and NFP-positive fibers in the vehicle treated tissue. In contrast, there were more myelinated axons (green/yellow) and myelin-forming cell (red) preservation in the estrogen treated rat spinal cord (Fig. 3d), suggesting protective effects provided by estrogen when compared to vehicle treatment.
Decreases in blood flow and increases in angiogenic markers following chronic SCI
Ischemic conditions following SCI result in damage to the blood vessels, and in an effort to repair the damage, increased angiogenesis occurs in the days following SCI. Our findings using Laser Doppler Imaging suggest that blood flow to the spinal cord was significantly reduced both immediately after injury and at 3 hours post-injury in SCI rats by 78% in the core region, when compared with sham rats (Fig. 4a). The time-course of increase in angiogenic markers was examined following SCI to determine when treatment with angiogenic factors, which theoretically would increase microvessel growth, would occur. Western blot analysis of angiogenic factors using spinal cord tissues from sham and SCI rats on 3 and 7 days post-SCI revealed that the angiogenic markers Ang-2 and Tie-2 were increased as early as day 3 in SCI tissue when compared with sham rats and remained elevated until day 7 post-SCI (Fig. 4b). Immunohistolabeling of angiogenic markers was also performed on sham and SCI spinal cord sections that contained the lesion and surrounding penumbra using antibodies against Ang-2, Tie-2, VEGF, bFGF, and counterstained with the endothelial marker CD-31 (Fig. 4b, c). Very little expression of Ang-2 and Tie-2 were seen in sham rats, however, all the angiogeneic markers were increased in areas surrounding the lesion in the spinal cord at 7 days post-SCI. The co-localization of the vessel endothelial marker CD-31 and angiogenic factors Tie-2 and VEGF, as well as an increase in their levels, were also detected in microvessels on day 7 following injury, compared with sham (Fig. 4c). These data demonstrate microvessel remodeling after SCI.
Fig. 4.
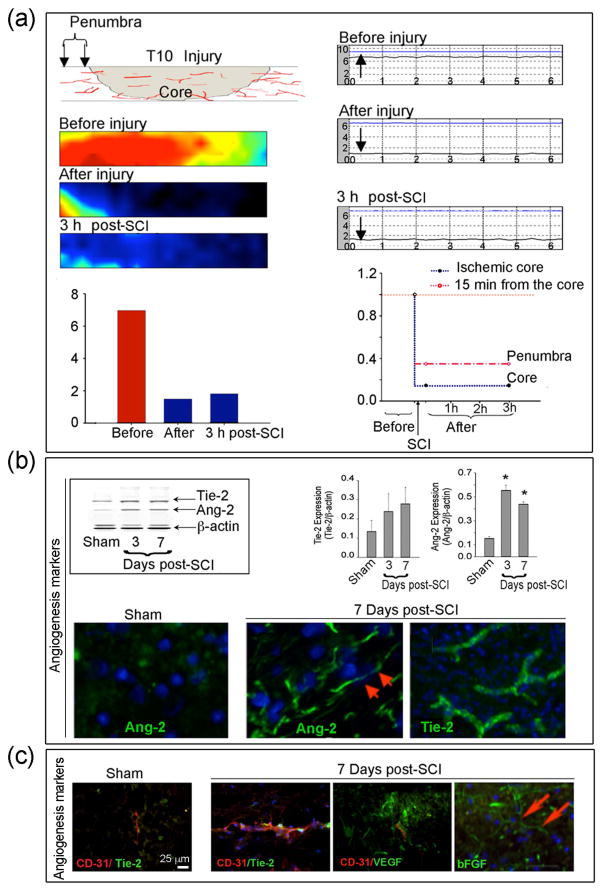
Local blood flow and angiogenesis markers alter following SCI. Local blood flow was measured using Laser scanning imaging or the conventional Laser Doppler single point measurements. (a) Diagram of the scanning spots at injury core (Tx-Lx), 15 mm from the center of core is defined as penumbra. (Left panels) Blood flow Laser image scanning before injury, soon after and 3 h after injury with quantification of the blood flow image in the lower left panel. The average of normal blood flow (7 unit), drastically dropped by 78% after SCI in the core region. The change in the penumbra region was relatively small. After 3 hr, the average blood flow slightly increased, by 26% of the normal. (Right panels) Single point Laser Doppler scanning before, immediately after, and 3 h after SCI. In the core region, the normal blood flow was about 7.37 (voltage unit), which dropped by 75% after SCI. There was no further change after 3 h in the core region. Quantification of the Laser Doppler scanning shown as normal blood flow defined as 1 (red dash line), the percentage of blood flow compare to normal was demonstrated in the core region (blue dash line) and penumbra (pink dash line), respectively. Post-SCI blood flow in the penumbra region (15 mm from center of ischemic core) showed less but significant decrease (67% reduction) and there was no obvious change after 3 h. n = 3. (b) Expression of angiogenic factors following SCI. Western blot analysis of angiogenic factors in the penumbra spinal cord in SCI animals at days 3 and 7 compared to sham. Representative immunoblots for Ang-2 and Tie-2 expression in the left panel and quantitative data from n = 3 in the right panel as Mean ± SEM. (Lower panel) Angiogenic markers were increased following SCI. Spinal cord lesion sections (1 cm) were removed from sham and SCI rats on day 7 following injury for immunofluorescent labeling of angiogenic markers, shown in green, using antibody against Ang-2 and Tie-2. Cell nuclei were stained with Bis-benzimide (blue). Red arrows indicate increased angiogenic marker staining following SCI. n = 3 (400× magnification). (c) Expression of angiogenic factors in endothelial cells after SCI. Immunofluorescence staining shows co-localization of vessel endothelial marker CD31 and angiogenic factors Tie-2 and VEGF at 7 days after SCI. bFGF another marker for angiogenesis was also increased 7 days post-SCI. Blue indicates Hoechst staining of all cells. n = 3 (400× magnification).
Low dose estrogen improves microvessel growth and protects myelin and myelin forming cells
The effects of estrogen therapy on angiogenic and microvessel growth markers were examined in spinal cord penumbra at various time-points via immunohistochemical labeling. For immunofluorescence, GLUT-1 antibody (green) was used as a marker of microcapillaries and myelin basic protein (MBP) antibody for myelin and myelin forming cells (red) (Fig. 5a). The extent of microvessel growth was examined in the penumbra of estrogen (10 μg) treated rats at day 10 post-SCI. Growth of microvessels in estrogen treated penumbra of injured rats was substantially greater than vehicle treated chronic SCI rats. Preservation of myelin (red) or MBP and myelin forming cells were also greater in estrogen treated versus vehicle treated chronic SCI rats, as shown in Fig. 5a, indicating a neuroprotective role for estrogen. On day 14, increases in both angiogenesis (Ang-2) and endothelial cell growth (CD31) were also demonstrated in spinal cord from the estrogen treated SCI rats, when compared with vehicle treated chronic SCI rats. These data, measured at selective time-points, demonstrated the feasibility of measuring angiogenic markers at different time-points following SCI.
Fig. 5.
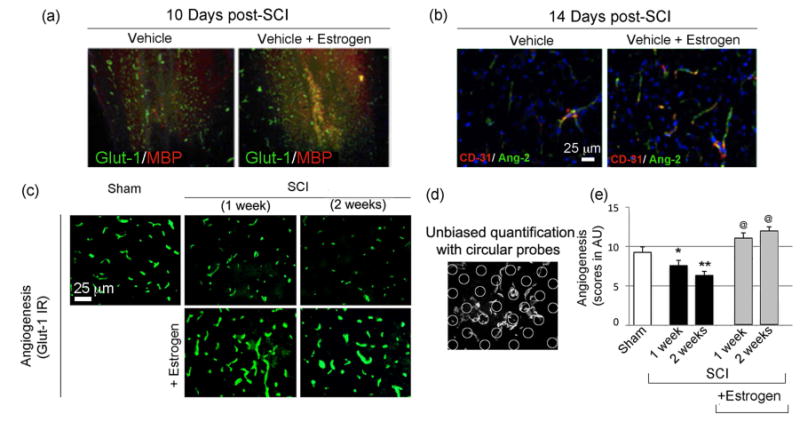
Low dose estrogen therapy increases angiogenesis and microvessel growth following SCI. (a-b) Spinal cord penumbra from vehicle and estrogen treated SCI rats were labeled with Glut-1 antibody (green) and MBP antibody (red); or CD31 (green) and Ang-2 (red) at 14 days post-SCI. Sham rats are not shown. n ≥ 3. (c) On 7 and 14 days post-SCI, profiles of Glut-1, a marker for angiogenesis was tested for estrogen efficacies. Spinal cord penumbra samples from the ventral horn of sham, vehicle, and estrogen treated SCI rats were labeled with Glut-1 antibody (green) at 7 and 14 days following SCI. n = 3 (200× magnification). (d) Quantitative evaluation of Glut-1 staining with circular probes and applying stereological principles as points of intersection of the vessel, as indicated by Glut-1 staining. The arbitrary circles were counted in at least four images per sham or SCI rats with or without estrogen treatment and were tested for statistical significance. (e) Quantification of Glut-1 IR indicated a progressive loss over a time period of 1-2 weeks. SEM *p < 0.05 or **p < 0.01 compared to sham and @p < 0.05 compared to injury + vehicle. (n ≥ 3).
Next, microvessel growth was examined using Glut-1 antibody as a marker for microcapillaries in rats following SCI. Our results on day 7 and 14 post-SCI indicated significant reduction of microcapillaires in the vehicle treated SCI penumbra compared to sham (Fig. 5c-e). In contrast, markedly increased Glut-1+ microcapillaries were seen in the penumbra of estrogen treated SCI rats compared with vehicle treated chronic SCI (Fig. 5c-e). Our findings suggest that estrogen treatment substantially improves microvessel growth in chronic SCI rats.
Low dose estrogen therapy improves motor function following chronic SCI
Previous studies with low dose estrogen therapy were performed in an acute model of SCI to determine if the secondary events that occur within hours of injury would become attenuated. These studies were used to select the lowest optimal dose of estrogen needed to reduce secondary damage. However, one of the main focuses of any SCI therapy is to ultimately improve motor function; thus, chronic SCI experiments are used to determine if rats recover following injury. Sham and SCI rats received a bolus tail vein injection of estrogen (10 or 100 μg/kg) or vehicle (saline) at 15 min post-injury. In addition, constant dose of estrogen (10 or 100 μg/kg/day) or vehicle were maintained for 7 days post-SCI. Rats were followed for 6 weeks post-injury and BBB scores were recorded every 7 days until sacrifice under anesthesia on day 42 (Fig. 6). BBB scores were compared in vehicle treated rats, and the two estrogen treated groups of SCI rats. In the injury + vehicle group, there was a steep rise of scores on days 7 and 14, respectively, compared to day 1. This spontaneous recovery in injury + vehicle rats plateaued at a median score of 13 from days 21 to 42. A significant difference (p < 0.05) was noted between estrogen and vehicle treated rats from days 7 to 42 with the 10 μg and from days 14 to 42 with 100 μg, respectively, indicating the low dose estrogen therapy improved motor function after chronic SCI (Fig 6).
Fig. 6.
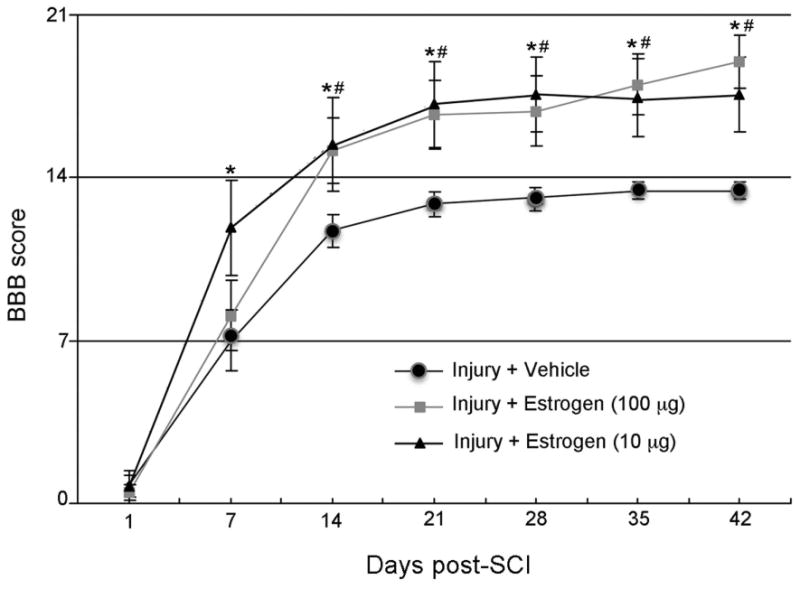
Low dose estrogen therapy improves motor function following SCI. SCI rats treated with estrogen (10 or 100 μg/kg) or vehicle were followed for 42 days post-injury. Rats were graded weekly using the BBB scale to examine motor function following injury. *P < 0.05 for estrogen (10μg) or #P < 0.05 estrogen (100μg) compared to injury + vehicle. n ≥ 7.
Discussion
While methylprednisolone is widely used to treat SCI (Bracken et al. 1998), its limited efficacy and numerous side effects make its use highly controversial (Hurlbert 2000, Pointillart et al. 2000). Although many agents have been shown to be beneficial in experimental SCI, few have been found to be efficacious in clinical settings, including GM-1 ganglioside (Geisler et al. 1991a, Geisler et al. 1991b), nimodipine (Pointillart et al. 2000), and gacyclidine (Tator 2006, Varma 2009). Thus, new therapeutic strategies must be developed to treat SCI. Therapy targeting one mechanism was not found to be beneficial, but estrogen with its multiple actions seems to have rendered a neuroprotective effect in SCI by inhibiting many of the destructive pathways, including calpain/caspase-mediated cell death pathways, inflammatory responses, and others. The attenuation of these destructive pathways by very low doses of estrogen in acute SCI (Samantaray et al. 2015) suggests its neuroprotective capability in ameliorating dysfunction in long term (chronic) SCI as well as estrogen's potential as a therapy in human SCI.
In our present study, the efficacy of estrogen therapy was examined using very low doses administered by IV injection in chronic SCI in rat at intervals of 7, 10, 14, and 42 days after injury. The long-term goal of this study was to investigate whether post-injury low dose estrogen (10 μg/kg) treatment can confirm the functional recovery previously demonstrated at 42 days after injury with high dose estrogen (4 mg/kg) treatment (Sribnick et al. 2010, Yune et al. 2004, Elkabes & Nicot 2014). Thus, we report that the use of low dose estrogen restored locomotor function following SCI by attenuating many of the destructive pathways, as mentioned above, and promoting angiogenesis, microvessel growth, and restoring blood supply essential for cell survival. Importantly, the locomotor function was significantly improved following treatment of chronic SCI rats with a low dose of estrogen. Therefore, the beneficial effects found in this study with low dose estrogen indicate it may be used as a treatment strategy for chronic human SCI.
The mechanical damage to the spinal cord caused by traumatic insult occurs immediately; disruption of the integrity of tissue within the area of damage (lesion site and penumbra) extends with time following injury (Sribnick et al. 2005, Banik et al. 1980, Hsu et al. 1985, Sharma et al. 1993, Bresnahan 1978, Balentine 1978). Cells in the lesion tissue undergo irreversible necrotic death while cells in the penumbra are partially damaged and apoptotic, and the apoptotic pathway may be reversible if treated with proper agents in time following injury (Ray et al. 2011, Elkabes & Nicot 2014, Sribnick et al. 2010, Sribnick et al. 2009). The current study demonstrated that such events occur and, for the first time, that integrity of the injured tissue at 14 days post-injury was decimated compared with that at 7 days. In contrast, the integrity of tissue in the injured area was significantly recovered/preserved by estrogen treatment not only at 7 days by reducing edema, swelling and other secondary injury events, but also markedly at 14 days of treatment. Thus, maintaining the structural integrity of injured tissue is pivotal for recovery of function in chronic SCI.
Inflammation is a common factor in neurodegeneration in CNS diseases and injuries. Following SCI, activation of resident glial cells, astrogliosis and microgliosis, and infiltration of neurotrophils are found in the lesion as well as in the lesion penumbra (Popovich et al. 1997, Sribnick et al. 2010, Ritz & Hausmann 2008). Activated astrocytes, microglia, and infiltrating inflammatory cells have been found to produce increased levels of pro-inflammatory cytokines and chemokines, including interleukins IL-12 and IL-1β, TNFα, MCP-1, and CCL-2 in spinal cord following injury (Ritz & Hausmann 2008, Cuzzocrea et al. 2008, Cox et al. 2015b). These factors not only destroy cells, they also degrade axons and myelin within hours following injury (Bresnahan 1978, Banik et al. 1980). As has been previously shown in CNS injury, the neuroprotective effects of estrogen attenuate several destructive pathways, including inflammation and macrophage infiltration and gliosis (Sribnick et al. 2010, Sribnick et al. 2005, Yune et al. 2004).
Although various inflammatory factors contribute to cell damage in SCI, other secondary injury components, particularly increased Ca2+ and Ca2+ mediated events have been found to play a pivotal role in neuron/oligodendrocyte death and axon and myelin degeneration in SCI as well as in TBI (Sribnick et al. 2005, Sribnick et al. 2007, Wingrave et al. 2003, Springer et al. 1997). Increased calpain has been found to be significantly involved in tissue degeneration in SCI, and treatment of animals with calpain inhibitor following SCI and TBI protected cells and inhibited calpain and caspase activity (Sribnick et al. 2005, Wingrave et al. 2003, Springer et al. 1997, Saatman et al. 1996, Yu et al. 2008). Further, treatment of SCI animals with doses of estrogen (17β-estradiol) ranging from 300μg-4mg/kg provided neuroprotection with a reduction in the number of TUNEL positive cells (Yune et al. 2004, Sribnick et al. 2005, Cuzzocrea et al. 2008, Lee et al. 2012). The number of apoptotic cells was also diminished when treated with a very low dose (10 μg/kg) of estrogen at different times following injury (Samantaray et al. 2011, Samantaray et al. 2015). With the increase in calpain activity following SCI, caspase activities (caspase-3, caspase-9) and other pro-apoptotic markers (e.g., Bax) were elevated. In contrast, these factors, including calpain, were reduced following treatment with estrogen (Samantaray et al. 2011, Lee et al. 2012, Yune et al. 2004, Sribnick et al. 2010, Sribnick et al. 2005, Samantaray et al. 2015). These apoptotic factors were reduced while anti-apoptotic Bcl-2 was increased after treatment, changing the Bax:Bcl-2 ratio and leading to cell protection. However, it is important to note that estrogen modulation of Ca2+ channels may affect the concentration of Ca2+ and reduce calpain activation and its subsequent degenerative processes (Sribnick et al. 2009, Mermelstein et al. 1996). Also, the final executioner of cell death, caspase-3, is a substrate of calpain, and thus, estrogen will elicit its neuroprotective effect by controlling/inhibiting calpain activity that is upstream of caspase-3 (Blomgren et al. 2001). Estrogen has been found to protect cells from the damaging effects of various insulting agents in vitro (Behl et al. 1995, Singer et al. 1999, Sur et al. 2003). Treatment with estrogen was shown to protect primary cortical neurons from glutamate-induced excitotoxicity with restoration of electrophysiological function (Sribnick et al. 2009, Sribnick et al. 2004).
NF-κB, the transcription factor following activation, has an important role in the development of inflammation, and estrogen treatment has been shown to inhibit its activation in vitro and in vivo in SCI (Sribnick et al. 2009, Bruce-Keller et al. 2000, Vegeto et al. 2003) as well as in ischemia in rats (Wen et al. 2004). Calpain is involved in the activation and translocation of NF-κB to nucleus and activation of NF-κB has been found in SCI (Schaecher et al. 2004, Bethea et al. 1998). It is not clear whether inhibition of NF-κB activation by estrogen, as demonstrated in SCI and in vitro, is mediated by estrogen receptors ERα and ERβ. The levels of ERα and ERβ are reduced in SCI lesion and penumbra concomitant with increased activity of calpain and NF-κB and other inflammatory components. Nonetheless, the increased level of estrogen receptors ERα and ERβ in the SCI rats after treatment with estrogen has been implicated in the estrogen-mediated attenuation of inflammatory processes (Sribnick et al. 2005, Sribnick et al. 2006, Sribnick et al. 2007). Since ERα and ERβ are substrates of calpain, inhibition of calpain activity by estrogen treatment would be expected to upregulate expression of both estrogen receptors to prevent inflammation in SCI. However, recent studies have shown that estrogen and its receptors are involved in reducing the ischemic damage and attenuating other destructive mechanisms in the spinal cord following injury (Sribnick et al. 2006, Vegeto et al. 2003, Dubal et al. 2001). The receptor-mediated neuroprotective effect of estrogen has been abrogated by using estrogen receptor antagonist Fulvestrant (ICI 182,780) not only in SCI (Das et al., unpublished), but also in experimental autoimmune encephalitis (EAE), an animal model of multiple sclerosis (MS) (Tiwari-Woodruff et al. 2007, Wu et al. 2013, Giraud et al. 2010) and in motoneuron cells in vitro (Das et al. 2011, Smith et al. 2011).
The increases in calpain activity found in the lesion and penumbra of acute and chronic SCI were attenuated not only by calpain inhibitor treatment, but also by estrogen administration. The axonal, myelin, and cytoskeletal proteins that are important for maintaining cellular integrity and of axon-myelin structural unit functionality (e.g., neurofilament proteins (NFPs), MBP, microtubular protein, and spectrin) are all substrates of calpain (Ray et al. 2003, Sorimachi et al. 1997, Takahashi 1990). Following SCI, the concomitant degradation/loss of these proteins will cause axonal and cellular disintegration leading to loss of function, which was prevented by treatment with estrogen (Sribnick et al. 2010, Yune et al. 2004). Thus, the estrogen treatment not only provided protection to neurons and oligodendrocytes, but also preserved axons and myelin, as demonstrated by deNFP and MBP staining (Fig. 3, 5), which did help recover locomotor function after SCI.
One of the major causes of cell and tissue damage in the area surrounding the injury site is the ischemic conditions brought about by disruption of or damage to blood vessels following trauma. Thus, cell damage may occur due to lack of blood supply, depriving cells of the nutrition needed for survival, as shown in SCI and TBI (Arai et al. 2009). Disruption of the vasculature of the spinal cord and tissue edema reduce transport of oxygen, growth factors, and other nutrients to the injured lesion and penumbra, which will lead to degeneration of tissue (Holtz et al. 1990). Therefore, promoting angiogenesis is critical to repair the damage and functional recovery. Although endogenous factors may make an effort to repair the damage, increased detrimental factors in the damaged area may be overwhelming and hence, hinder that effort. Nonetheless, upregulation or supplementation of endogenous agents, such as estrogen in the damaged area, may boost the repair process. The finding of increased angiogenic factors (Ang-2, Tie-2) that occur 3-7 days after SCI led us to hypothesize that treatment with these factors would increase microvessel growth. Estrogen is known to play a critical pro-angiogenic role in the physiology of the female reproductive system where recurrent neo-vascularization is a normal process. Angiogenesis is mediated by increased VEGF, bFGF, ERα, nitric oxide, and others, and some of these markers of angiogenesis are increased days after SCI and ischemia (Duckles & Krause 2003, Kato et al. 1997). Other angiogenic proteins (e.g., Tie-1, Tie-2, Ang-1, Ang-2) are expressed in endothelial cells and increased in ischemia (Wei et al. 2005, Wei et al. 2001, Wei et al. 2003) as well as in SCI (present report). The mechanisms of estrogen promoting angiogenesis in SCI have been suggested to be mediated by estrogen receptors (ERα, ERβ) and VEGF receptors (Flt-1, Flk-1) (Sribnick et al. 2006). Thus, the present study's finding that angiogenesis and microvessel growth increased following treatment with low dose estrogen at 10-14 days after injury may have a significant role to play in the protection of neurons and oligodendrocytes. This indicates that the neuroprotective role of estrogen may also lead to preservation of axons and myelin, which is essential for recovery of locomotor function. This estrogen-mediated neuroprotection seen early on (within 2 weeks) in acute and sub-chronic injury significantly contributed to recovery of function at 7-14 days and continued to 42 days with doses as low as 10 μg of estrogen. Treatment with this low dose has been found to be as good as higher doses and safer, since the lower dose will help reduce side effects. Since estrogen effects start earlier in SCI, the 10 μg dose likely reached optimal efficacy compared to higher doses. Moreover, our earlier report showed lower effects on cell death and inflammation following treatment with 5 μg estrogen compared to 10 or 100 μg estrogen (Samantaray et al. 2011).
Formation of an impenetrable scar in the lesion of SCI is one of the main obstacles for axonal growth and recovery of function. Although estrogen treatment reduced edema and reactive astrogliosis, prevented neurofilament and microtubule protein degradation, and improved function following SCI (Sribnick et al. 2010, Siriphorn et al. 2012, Chaovipoch et al. 2006), it is not known whether this steroid hormone has any effect on compromising the scar. However, one may speculate that functional recovery might have been able to be promoted, if the scar were compromised by estrogen, preserving axonal neurofilament proteins and particularly microtubule protein, which is thought to be involved in axon growth and perhaps guidance. Such stabilization of axonal proteins, reduction of astrogliosis by estrogen, and reduction of the scar by other agents have been found hospitable for axonal growth and improved function (Ruschel et al. 2015, Zhu et al. 2015, Soderblom et al. 2013, Saraceno et al. 2010). Nonetheless, as demonstrated in the current study, treatment with low doses of estrogen improves locomotor function following SCI and may be a potential therapeutic agent for treatment of chronic human SCI.
Conclusion
Treatment with high non-physiological dose of estrogen has been found to improve locomotor function following SCI [see review (Elkabes & Nicot 2014)]. Thus, for clinical relevance, the present study examined whether treatment with low doses of estrogen (10 μg of 17β-estradiol) would improve locomotor function following injury and reduce side effects. We found that such treatment decreased inflammation, tissue damage, gliosis, neuron and oligodendrocyte death and calpain activity in addition to preserving axons and myelin, increasing angiogenesis and improving locomotor function. The multi-active effects of low dose estrogen on different parameters, including protection of cells, preservation of the axon-myelin tract and increase of angiogenesis, may have contributed to improving locomotor function following injury. Although high dose non-physiological estrogen treatment has been found to improve function in rodent SCI, its use will have many unwanted side effects in humans, including cancer, deep vein thrombosis, feminizing effects in males. It will therefore never be used in human clinical trials. Thus, this pre-clinical study demonstrating recovery of locomotor function with low dose estrogen may have significant clinical implications for treatment of SCI.
Acknowledgments
This work was supported in part by funding from NIH (R01-NS3166, R01-NS45967); Veterans Administration (1IOBX00126; 1IOBX002349); South Carolina State Spinal Cord Research Fund (SCIRF-2015P-01; SCIRF-2015P-04); Department of Neurosurgery, MUSC; and MUSC-CTSA program. We also thank Dr. Kelly Guyton for her valuable contribution to this work. The authors declare no conflict of interest.
Abbreviations
- BBB
Basso, Beattie, and Bresnahan
- MP
methylprednisolone
- OCT
Optimum cutting temperature
- T10
Thoracic 10
- SCI
Spinal cord injury
References
- Agrawal SK, Fehlings MG. Mechanisms of secondary injury to spinal cord axons in vitro: role of Na+, Na(+)-K(+)-ATPase, the Na(+)-H+ exchanger, and the Na(+)-Ca2+ exchanger. J Neurosci. 1996;16:545–552. doi: 10.1523/JNEUROSCI.16-02-00545.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arai K, Jin G, Navaratna D, Lo EH. Brain angiogenesis in developmental and pathological processes: neurovascular injury and angiogenic recovery after stroke. FEBS J. 2009;276:4644–4652. doi: 10.1111/j.1742-4658.2009.07176.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balentine JD. Pathology of experimental spinal cord trauma. II. Ultrastructure of axons and myelin. Lab Invest. 1978;39:254–266. [PubMed] [Google Scholar]
- Banik NL, Powers JM, Hogan EL. The effects of spinal cord trauma on myelin. J Neuropathol Exp Neurol. 1980;39:232–244. doi: 10.1097/00005072-198005000-00002. [DOI] [PubMed] [Google Scholar]
- Basso DM, Beattie MS, Bresnahan JC. A sensitive and reliable locomotor rating scale for open field testing in rats. J Neurotrauma. 1995;12:1–21. doi: 10.1089/neu.1995.12.1. [DOI] [PubMed] [Google Scholar]
- Behl C, Widmann M, Trapp T, Holsboer F. 17-beta estradiol protects neurons from oxidative stress-induced cell death in vitro. Biochem Biophys Res Commun. 1995;216:473–482. doi: 10.1006/bbrc.1995.2647. [DOI] [PubMed] [Google Scholar]
- Bethea JR, Castro M, Keane RW, Lee TT, Dietrich WD, Yezierski RP. Traumatic spinal cord injury induces nuclear factor-kappaB activation. J Neurosci. 1998;18:3251–3260. doi: 10.1523/JNEUROSCI.18-09-03251.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blomgren K, Zhu C, Wang X, Karlsson JO, Leverin AL, Bahr BA, Mallard C, Hagberg H. Synergistic activation of caspase-3 by m-calpain after neonatal hypoxia-ischemia: a mechanism of pathological apoptosis? The Journal of biological chemistry. 2001;276:10191–10198. doi: 10.1074/jbc.M007807200. [DOI] [PubMed] [Google Scholar]
- Bracken MB, Shepard MJ, Holford TR, et al. Methylprednisolone or tirilazad mesylate administration after acute spinal cord injury: 1-year follow up. Results of the third National Acute Spinal Cord Injury randomized controlled trial. Journal of neurosurgery. 1998;89:699–706. doi: 10.3171/jns.1998.89.5.0699. [DOI] [PubMed] [Google Scholar]
- Bresnahan JC. An electron-microscopic analysis of axonal alterations following blunt contusion of the spinal cord of the rhesus monkey (Macaca mulatta) Journal of the neurological sciences. 1978;37:59–82. doi: 10.1016/0022-510x(78)90228-9. [DOI] [PubMed] [Google Scholar]
- Bruce-Keller AJ, Keeling JL, Keller JN, Huang FF, Camondola S, Mattson MP. Antiinflammatory effects of estrogen on microglial activation. Endocrinology. 2000;141:3646–3656. doi: 10.1210/endo.141.10.7693. [DOI] [PubMed] [Google Scholar]
- Chaovipoch P, Jelks KA, Gerhold LM, West EJ, Chongthammakun S, Floyd CL. 17beta-estradiol is protective in spinal cord injury in post- and pre-menopausal rats. J Neurotrauma. 2006;23:830–852. doi: 10.1089/neu.2006.23.830. [DOI] [PubMed] [Google Scholar]
- Cox A, Varma A, Banik N. Recent advances in the pharmacologic treatment of spinal cord injury. Metabolic brain disease. 2015a;30:473–482. doi: 10.1007/s11011-014-9547-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox A, Varma A, Barry J, Vertegel A, Banik N. Nanoparticle Estrogen in Rat Spinal Cord Injury Elicits Rapid Anti-Inflammatory Effects in Plasma, Cerebrospinal Fluid, and Tissue. J Neurotrauma. 2015b doi: 10.1089/neu.2014.3730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cuzzocrea S, Genovese T, Mazzon E, et al. Effect of 17beta-estradiol on signal transduction pathways and secondary damage in experimental spinal cord trauma. Shock. 2008;29:362–371. doi: 10.1097/shk.0b013e31814545dc. [DOI] [PubMed] [Google Scholar]
- Das A, Smith JA, Gibson C, Varma AK, Ray SK, Banik NL. Estrogen receptor agonists and estrogen attenuate TNF-alpha-induced apoptosis in VSC4.1 motoneurons. The Journal of endocrinology. 2011;208:171–182. doi: 10.1677/JOE-10-0338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dubal DB, Zhu H, Yu J, Rau SW, Shughrue PJ, Merchenthaler I, Kindy MS, Wise PM. Estrogen receptor alpha, not beta, is a critical link in estradiol-mediated protection against brain injury. Proc Natl Acad Sci U S A. 2001;98:1952–1957. doi: 10.1073/pnas.041483198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duckles SP, Krause DN. Vascular and endothelial function: Role of gonadal steroids in neuronal and vascular plasticity. In: Maiese K, editor. Neuronal and Vascular Plasticity : Elucidating Basic Cellular Mechanisms for Future Therapeutic Discovery. Kluwer Press; New York: 2003. pp. 95–115. [Google Scholar]
- Elkabes S, Nicot AB. Sex steroids and neuroprotection in spinal cord injury: a review of preclinical investigations. Exp Neurol. 2014;259:28–37. doi: 10.1016/j.expneurol.2014.01.008. [DOI] [PubMed] [Google Scholar]
- Geisler FH, Dorsey FC, Coleman WP. Correction: recovery of motor function after spinal-cord injury--a randomized, placebo-controlled trial with GM-1 ganglioside. The New England journal of medicine. 1991a;325:1659–1660. [PubMed] [Google Scholar]
- Geisler FH, Dorsey FC, Coleman WP. Recovery of motor function after spinal-cord injury--a randomized, placebo-controlled trial with GM-1 ganglioside. The New England journal of medicine. 1991b;324:1829–1838. doi: 10.1056/NEJM199106273242601. [DOI] [PubMed] [Google Scholar]
- Giraud SN, Caron CM, Pham-Dinh D, Kitabgi P, Nicot AB. Estradiol inhibits ongoing autoimmune neuroinflammation and NFkappaB-dependent CCL2 expression in reactive astrocytes. Proc Natl Acad Sci U S A. 2010;107:8416–8421. doi: 10.1073/pnas.0910627107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holtz A, Nystrom B, Gerdin B. Relation between spinal cord blood flow and functional recovery after blocking weight-induced spinal cord injury in rats. Neurosurgery. 1990;26:952–957. doi: 10.1097/00006123-199006000-00005. [DOI] [PubMed] [Google Scholar]
- Hsu CY, Halushka PV, Hogan EL, Banik NL, Lee WA, Perot PL., Jr Alteration of thromboxane and prostacyclin levels in experimental spinal cord injury. Neurology. 1985;35:1003–1009. doi: 10.1212/wnl.35.7.1003. [DOI] [PubMed] [Google Scholar]
- Hurlbert RJ. Methylprednisolone for acute spinal cord injury: an inappropriate standard of care. Journal of neurosurgery. 2000;93:1–7. doi: 10.3171/spi.2000.93.1.0001. [DOI] [PubMed] [Google Scholar]
- Kato H, Kanellopoulos GK, Matsuo S, Wu YJ, Jacquin MF, Hsu CY, Kouchoukos NT, Choi DW. Neuronal apoptosis and necrosis following spinal cord ischemia in the rat. Exp Neurol. 1997;148:464–474. doi: 10.1006/exnr.1997.6707. [DOI] [PubMed] [Google Scholar]
- Krause JS, Newman SD. Clinical Outcomes After Spinal Cord Injury. In: Banik N, Ray SK, editors. Handbook of Neurochemistry and Molecular Neurobiology. Springer; New York, NY: 2009. pp. 615–632. Vol Brain and Spinal Cord Injury. [Google Scholar]
- Lee JY, Choi SY, Oh TH, Yune TY. 17beta-Estradiol inhibits apoptotic cell death of oligodendrocytes by inhibiting RhoA-JNK3 activation after spinal cord injury. Endocrinology. 2012;153:3815–3827. doi: 10.1210/en.2012-1068. [DOI] [PubMed] [Google Scholar]
- Li Y, Lu Z, Keogh CL, Yu SP, Wei L. Erythropoietin-induced neurovascular protection, angiogenesis, and cerebral blood flow restoration after focal ischemia in mice. J Cereb Blood Flow Metab. 2007;27:1043–1054. doi: 10.1038/sj.jcbfm.9600417. [DOI] [PubMed] [Google Scholar]
- Mermelstein PG, Becker JB, Surmeier DJ. Estradiol reduces calcium currents in rat neostriatal neurons via a membrane receptor. J Neurosci. 1996;16:595–604. doi: 10.1523/JNEUROSCI.16-02-00595.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perot PL, Jr, Lee WA, Hsu CY, Hogan EL, Cox RD, Gross AJ. Therapeutic model for experimental spinal cord injury in the rat: I. Mortality and motor deficit. Central nervous system trauma : journal. of the American Paralysis Association. 1987;4:149–159. doi: 10.1089/cns.1987.4.149. [DOI] [PubMed] [Google Scholar]
- Pointillart V, Petitjean ME, Wiart L, Vital JM, Lassié P, Thicoipé M, P D. Pharmacological therapy of spinal cord injury during the acute phase. Spinal Cord. 2000;38:71–76. doi: 10.1038/sj.sc.3100962. [DOI] [PubMed] [Google Scholar]
- Popovich PG, Wei P, Stokes BT. Cellular inflammatory response after spinal cord injury in Sprague-Dawley and Lewis rats. J Comp Neurol. 1997;377:443–464. doi: 10.1002/(sici)1096-9861(19970120)377:3<443::aid-cne10>3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- Ray SK, Matzelle DD, Sribnick EA, Guyton MK, Wingrave JM, Banik NL. Calpain inhibitor prevented apoptosis and maintained transcription of proteolipid protein and myelin basic protein genes in rat spinal cord injury. Journal of chemical neuroanatomy. 2003;26:119–124. doi: 10.1016/s0891-0618(03)00044-9. [DOI] [PubMed] [Google Scholar]
- Ray SK, Samantaray S, Smith JA, Matzelle DD, Das A, Banik NL. Inhibition of cysteine proteases in acute and chronic spinal cord injury. Neurotherapeutics : the journal. of the American Society for Experimental NeuroTherapeutics. 2011;8:180–186. doi: 10.1007/s13311-011-0037-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritz MF, Hausmann ON. Effect of 17beta-estradiol on functional outcome, release of cytokines, astrocyte reactivity and inflammatory spreading after spinal cord injury in male rats. Brain research. 2008;1203:177–188. doi: 10.1016/j.brainres.2008.01.091. [DOI] [PubMed] [Google Scholar]
- Ruschel J, Hellal F, Flynn KC, et al. Axonal regeneration. Systemic administration of epothilone B promotes axon regeneration after spinal cord injury. Science. 2015;348:347–352. doi: 10.1126/science.aaa2958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saatman KE, Murai H, Bartus RT, Smith DH, Hayward NJ, Perri BR, McIntosh TK. Calpain inhibitor AK295 attenuates motor and cognitive deficits following experimental brain injury in the rat. Proc Natl Acad Sci U S A. 1996;93:3428–3433. doi: 10.1073/pnas.93.8.3428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samantaray S, Das A, Matzelle DC, Yu SP, Wei L, Varma AK, Ray SK, Banik NL. (2015 ) Administration of low dose-estrogen attenuates gliosis and protects neurons in acute spinal cord injury in rats. J Neurochem. doi: 10.1111/jnc.13464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samantaray S, Smith JA, Das A, Matzelle DD, Varma AK, Ray SK, Banik NL. Low dose estrogen prevents neuronal degeneration and microglial reactivity in an acute model of spinal cord injury: effect of dosing, route of administration, and therapy delay. Neurochem Res. 2011;36:1809–1816. doi: 10.1007/s11064-011-0498-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saraceno GE, Bertolino ML, Galeano P, Romero JI, Garcia-Segura LM, Capani F. Estradiol therapy in adulthood reverses glial and neuronal alterations caused by perinatal asphyxia. Exp Neurol. 2010;223:615–622. doi: 10.1016/j.expneurol.2010.02.010. [DOI] [PubMed] [Google Scholar]
- Schaecher K, Goust JM, Banik NL. The effects of calpain inhibition on IkB alpha degradation after activation of PBMCs: identification of the calpain cleavage sites. Neurochem Res. 2004;29:1443–1451. doi: 10.1023/b:nere.0000026410.56000.dd. [DOI] [PubMed] [Google Scholar]
- Sharma HS, Olsson Y, Cervos-Navarro J. Early perifocal cell changes and edema in traumatic injury of the spinal cord are reduced by indomethacin, an inhibitor of prostaglandin synthesis. Experimental study in the rat. Acta neuropathologica. 1993;85:145–153. doi: 10.1007/BF00227761. [DOI] [PubMed] [Google Scholar]
- Singer CA, Figueroa-Masot XA, Batchelor RH, Dorsa DM. The mitogen-activated protein kinase pathway mediates estrogen neuroprotection after glutamate toxicity in primary cortical neurons. J Neurosci. 1999;19:2455–2463. doi: 10.1523/JNEUROSCI.19-07-02455.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siriphorn A, Dunham KA, Chompoopong S, Floyd CL. Postinjury administration of 17beta-estradiol induces protection in the gray and white matter with associated functional recovery after cervical spinal cord injury in male rats. J Comp Neurol. 2012;520:2630–2646. doi: 10.1002/cne.23056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith JA, Das A, Butler JT, Ray SK, Banik NL. Estrogen or estrogen receptor agonist inhibits lipopolysaccharide induced microglial activation and death. Neurochem Res. 2011;36:1587–1593. doi: 10.1007/s11064-010-0336-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soderblom C, Luo X, Blumenthal E, et al. Perivascular fibroblasts form the fibrotic scar after contusive spinal cord injury. J Neurosci. 2013;33:13882–13887. doi: 10.1523/JNEUROSCI.2524-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sorimachi H, Ishiura S, Suzuki K. Structure and physiological function of calpains. Biochem J. 1997;328(Pt 3):721–732. doi: 10.1042/bj3280721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Springer JE, Azbill RD, Kennedy SE, George J, Geddes JW. Rapid calpain I activation and cytoskeletal protein degradation following traumatic spinal cord injury: attenuation with riluzole pretreatment. J Neurochem. 1997;69:1592–1600. doi: 10.1046/j.1471-4159.1997.69041592.x. [DOI] [PubMed] [Google Scholar]
- Sribnick EA, Del Re AM, Ray SK, Woodward JJ, Banik NL. Estrogen attenuates glutamate-induced cell death by inhibiting Ca2+ influx through L-type voltage-gated Ca2+ channels. Brain research. 2009;1276:159–170. doi: 10.1016/j.brainres.2009.04.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sribnick EA, Matzelle DD, Banik NL, Ray SK. Direct evidence for calpain involvement in apoptotic death of neurons in spinal cord injury in rats and neuroprotection with calpain inhibitor. Neurochem Res. 2007;32:2210–2216. doi: 10.1007/s11064-007-9433-7. [DOI] [PubMed] [Google Scholar]
- Sribnick EA, Matzelle DD, Ray SK, Banik NL. Estrogen treatment of spinal cord injury attenuates calpain activation and apoptosis. J Neurosci Res. 2006;84:1064–1075. doi: 10.1002/jnr.21016. [DOI] [PubMed] [Google Scholar]
- Sribnick EA, Ray SK, Nowak MW, Li L, Banik NL. 17beta-estradiol attenuates glutamate-induced apoptosis and preserves electrophysiologic function in primary cortical neurons. J Neurosci Res. 2004;76:688–696. doi: 10.1002/jnr.20124. [DOI] [PubMed] [Google Scholar]
- Sribnick EA, Samantaray S, Das A, Smith J, Matzelle DD, Ray SK, Banik NL. Postinjury estrogen treatment of chronic spinal cord injury improves locomotor function in rats. J Neurosci Res. 2010;88:1738–1750. doi: 10.1002/jnr.22337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sribnick EA, Wingrave JM, Matzelle DD, Wilford GG, Ray SK, Banik NL. Estrogen attenuated markers of inflammation and decreased lesion volume in acute spinal cord injury in rats. J Neurosci Res. 2005;82:283–293. doi: 10.1002/jnr.20622. [DOI] [PubMed] [Google Scholar]
- Stokes BT, Fox P, Hollinden G. Extracellular calcium activity in the injured spinal cord. Exp Neurol. 1983;80:561–572. doi: 10.1016/0014-4886(83)90307-2. [DOI] [PubMed] [Google Scholar]
- Sur P, Sribnick EA, Wingrave JM, Nowak MW, Ray SK, Banik NL. Estrogen attenuates oxidative stress-induced apoptosis in C6 glial cells. Brain research. 2003;971:178–188. doi: 10.1016/s0006-8993(03)02349-7. [DOI] [PubMed] [Google Scholar]
- Takahashi K. Calpain substrate specificity. In: Mellgren R, Murachi T, editors. Intracellular Calcium-Dependent Proteolysis. CRC Press; Boca rAton, FL: 1990. pp. 55–74. [Google Scholar]
- Tator CH. Review of treatment trials in human spinal cord injury: issues, difficulties, and recommendations. Neurosurgery. 2006;59:957–982. doi: 10.1227/01.NEU.0000245591.16087.89. discussion 982-957. [DOI] [PubMed] [Google Scholar]
- Tiwari-Woodruff S, Morales LB, Lee R, Voskuhl RR. Differential neuroprotective and antiinflammatory effects of estrogen receptor (ER)alpha and ERbeta ligand treatment. Proc Natl Acad Sci U S A. 2007;104:14813–14818. doi: 10.1073/pnas.0703783104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varma A. Spinal Cord Injury - A Clinical Perspective. In: Banik N, Ray SK, editors. In: Handbook of Neurochemistry and Molecular Neurobiology. Springer; New York, NY: 2009. pp. 633–648. Vol Brain and Spinal Cord Trauma. [Google Scholar]
- Varma A, Hill EG, Nicholas J, Selassie A. Predictors of early mortality after traumatic spinal cord injury: a population-based study. Spine (Phila Pa 1976) 2010;35:778–783. doi: 10.1097/BRS.0b013e3181ba1359. [DOI] [PubMed] [Google Scholar]
- Varma AK, Das A, Wallace Gt, Barry J, Vertegel AA, Ray SK, Banik NL. Spinal cord injury: a review of current therapy, future treatments, and basic science frontiers. Neurochem Res. 2013;38:895–905. doi: 10.1007/s11064-013-0991-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vegeto E, Belcredito S, Etteri S, et al. Estrogen receptor-alpha mediates the brain antiinflammatory activity of estradiol. Proc Natl Acad Sci U S A. 2003;100:9614–9619. doi: 10.1073/pnas.1531957100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei L, Erinjeri JP, Rovainen CM, Woolsey TA. Collateral growth and angiogenesis around cortical stroke. Stroke. 2001;32:2179–2184. doi: 10.1161/hs0901.094282. [DOI] [PubMed] [Google Scholar]
- Wei L, Keogh CL, Whitaker VR, Theus MH, Yu SP. Angiogenesis and stem cell transplantation as potential treatments of cerebral ischemic stroke. Pathophysiology : the official journal of the International Society for Pathophysiology / ISP. 2005;12:47–62. doi: 10.1016/j.pathophys.2004.11.002. [DOI] [PubMed] [Google Scholar]
- Wei L, Yin K, Lee JM, Chao J, Yu SP, Lin TN, Hsu CW. Restorative potential of angiogenesis after ischemic stroke. In: Maiese K, editor. Neuronal and Vascular Plasticity. Kluwer Academic/Plenum Publishers; London: 2003. pp. 75–94. [Google Scholar]
- Wen Y, Yang S, Liu R, Perez E, Yi KD, Koulen P, Simpkins JW. Estrogen attenuates nuclear factor-kappa B activation induced by transient cerebral ischemia. Brain research. 2004;1008:147–154. doi: 10.1016/j.brainres.2004.02.019. [DOI] [PubMed] [Google Scholar]
- Wingrave JM, Schaecher KE, Sribnick EA, Wilford GG, Ray SK, Hazen-Martin DJ, Hogan EL, Banik NL. Early induction of secondary injury factors causing activation of calpain and mitochondria-mediated neuronal apoptosis following spinal cord injury in rats. J Neurosci Res. 2003;73:95–104. doi: 10.1002/jnr.10607. [DOI] [PubMed] [Google Scholar]
- Wu WF, Tan XJ, Dai YB, Krishnan V, Warner M, Gustafsson JA. Targeting estrogen receptor beta in microglia and T cells to treat experimental autoimmune encephalomyelitis. Proc Natl Acad Sci U S A. 2013;110:3543–3548. doi: 10.1073/pnas.1300313110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xie LP, Fu WX, Jin C, Dong XY, Chen WF. Negative regulation of monocyte chemoattractant protein-1 gene expression by a mouse estrogen-enhanced transcript. European journal of immunology. 2002;32:2837–2846. doi: 10.1002/1521-4141(2002010)32:10<2837::AID-IMMU2837>3.0.CO;2-V. [DOI] [PubMed] [Google Scholar]
- Yu CG, Joshi A, Geddes JW. Intraspinal MDL28170 microinjection improves functional and pathological outcome following spinal cord injury. J Neurotrauma. 2008;25:833–840. doi: 10.1089/neu.2007.0490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yune TY, Kim SJ, Lee SM, Lee YK, Oh YJ, Kim YC, Markelonis GJ, Oh TH. Systemic administration of 17beta-estradiol reduces apoptotic cell death and improves functional recovery following traumatic spinal cord injury in rats. J Neurotrauma. 2004;21:293–306. doi: 10.1089/089771504322972086. [DOI] [PubMed] [Google Scholar]
- Zhu Y, Soderblom C, Krishnan V, Ashbaugh J, Bethea JR, Lee JK. Hematogenous macrophage depletion reduces the fibrotic scar and increases axonal growth after spinal cord injury. Neurobiol Dis. 2015;74:114–125. doi: 10.1016/j.nbd.2014.10.024. [DOI] [PMC free article] [PubMed] [Google Scholar]