

ABSTRACT
Reviews on whole body human cold adaptation generally do not distinguish between population studies and dedicated acclimation studies, leading to confusing results. Population studies show that indigenous black Africans have reduced shivering thermogenesis in the cold and poor cold induced vasodilation in fingers and toes compared to Caucasians and Inuit. About 40,000 y after humans left Africa, natives in cold terrestrial areas seems to have developed not only behavioral adaptations, but also physiological adaptations to cold. Dedicated studies show that repeated whole body exposure of individual volunteers, mainly Caucasians, to severe cold results in reduced cold sensation but no major physiological changes. Repeated cold water immersion seems to slightly reduce metabolic heat production, while repeated exposure to milder cold conditions shows some increase in metabolic heat production, in particular non-shivering thermogenesis. In conclusion, human cold adaptation in the form of increased metabolism and insulation seems to have occurred during recent evolution in populations, but cannot be developed during a lifetime in cold conditions as encountered in temperate and arctic regions. Therefore, we mainly depend on our behavioral skills to live in and survive the cold.
KEYWORDS: acclimation, adaptation, brown fat, cold, cold induced vasodilation; cold water immersion; human, non-shivering thermogenesis, shivering
Introduction
The most widely accepted view of geographic origin and early migration of humans is that they originate from tropical Africa and started to disperse over the world only about 40,000 y ago.1
Since high temperatures dominate in that area, one can assume that at that time humans possessed optimal behavioral and physiological mechanisms to cope with heat and less developed physiological and behavioral mechanisms to cope with cold as encountered in temperate and arctic regions. Even though it is well documented that climatic changes occurred in tropical regions, seasonal variation in ambient temperature is blunted compared to temperate climates and heat stress dominates.2 40,000 y is a relatively short time span in evolutionary terms and it is therefore interesting to investigate if current modern humans are still tropical animals. What mechanisms do we have to cope with cold and do they differ from mechanisms that we supposed to have had 40,000 y ago?
Both tropical and (Ant)Arctic climates are challenging climates for humans due to extreme heat and cold respectively. It is assumed that moderate climates with ambient temperatures of around 21°C need minimal human energy investment in comparison to heat and cold exposure.3 However, it is good to realize that human protection from adverse performance and health outcomes is required already in temperate climates due to daily and seasonal variations in temperature, and not only for temperature extremes.
Since we are not able to compare the population living 40,000 y ago with the current population, we have to make some assumptions in an attempt to make comparisons. One assumption is that humans of the current population of central Africa possess comparable thermoregulatory mechanisms as humans 40,000 y ago. This assumption is defendable since at least part of the African population continued to live under similar climatological circumstances. Therefore we can compare the heat and cold coping mechanisms of the current population of tropical Africa with people living in cold areas for millennia, in order to learn about the adaptivea mechanisms that have occurred. Another way to investigate adaptations is to compare Caucasians to the population originating from Africa that is currently living in colder areas, such as the black Americans.
Finally, experimental studies on repeated exposure to cold may elucidate the mechanisms to acclimatize. It is the purpose of this review to contribute to the discussion if and how humans adapt to cold, including population studies ánd dedicated cold acclimation studies. In this review exposure to cold is categorized as severe, moderate and mild according to the thermal stressor that includes both the medium (at a given temperature cold water exposure is more severe that cold air exposure) and the temperature of the medium. Thermal strain is the reaction of the body to the cold exposure often quantified by core body temperature.
The human thermoregulatory system relies on behavior and on physiological responses for thermal homeostasis.10 Our physiological mechanisms are limited: basically, thermal balance in humans is maintained by vasodilation/vasoconstriction of the skin and peripheral tissues within the so-called thermo-neutral zone.11 We have one extra physiological mechanism in the heat (sweat evaporation) and 2 extra mechanisms in the cold (shivering- [ST] and nonshivering-thermogenesis [NST]). Humans are good sweaters with maximal values observed exceeding 3.5 l/hour.12 Since the heat of vaporization of water is high, this leads to a cooling power of over 2500 W! Moreover, the sweating capacity adapts very well to the demand: 10 weeks of heat acclimation can double sweat production.13 This acquired additional cooling power is maintained for several weeks, even when not exposed to heat anymore.14 In conclusion, our thermal response to heat and our adaptation capabilities to heat are well developed. This review will focus on our capabilities to counteract cold exposure, which are less effective, at least on the long term.
Human adaptation to thermal extremes is not only an academic question, but important to assess the impact of climate change on mortality and morbidity.4 It is predicted that we will face more thermal extremes in the future, and the role of adaptation is essential to understand its impact. Some studies even predict the extinction of human populations that live in extremely hot climates in a few decades,5 but they hardly take human adaptation into account. On the other side, it is not unlikely that Northern Europe may experience cooling due to the thermohaline circulation6 and then it is good to know if and to what extent we can adapt to cold. Another important question is if workers are better protected against cold after repeated cold exposure. Occupational work is expected to increase in cold areas due to the exploration of natural gas (over 30% of world gas reserve is located in the Arctic area) and the opening of the waterway north of Russia. Similar questions arise in the area of sports, where running, skiing or skating in extreme cold is increasingly popular: does it have any benefits to expose oneself to cold prior to the sports event in order to be optimally prepared? Therefore, this review focuses on the capability of humans to adapt to cold.
First the basic mechanisms to cope with cold will be discussed, followed by differences between populations living in hot and in cold areas. In most reviews on cold adaptation7-10 studies regarding population differences are intertwined with acclimation studies (in line with the definition in the Glossary of terms11). This may lead to confusion and therefore this review starts with discussing the results of population studies followed by studies on acclimation to cold. When required, small excursions will be made to the effect of heat exposure on humans. A recent review on heat adaptation provides extended information on adaptation to heat.12
Mechanisms to cope with cold
Cold induced thermogenesis can be divided in NST and ST. In a cold environment, heat production increases by 10–30 W during the initial first minutes without any increase of muscle activity.13 Later, extra heat is generated by involuntary contractions of skeletal muscles (shivering). Shivering occurs when core and skin temperature surpass a certain threshold and may produce heat equivalent to about 4 times resting metabolism14 (Fig. 1). It is hard to quantify the non-shivering component during shivering, but there is some evidence that this component continues in severe cold.15 It is important to note that shivering and profuse sweating are limited in time due to exhaustion of glycogen16 and water resources,17 respectively.
Figure 1.
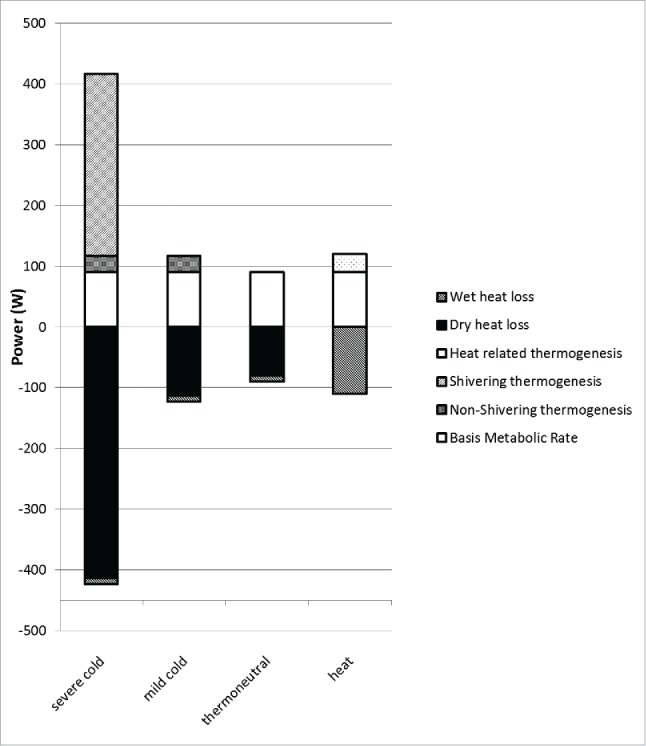
Heat balance in Watts for exposure to severe cold, mild cold, thermoneutrality and heat. Please note the contribution of non-shivering thermogenesis in mild cold and shivering and non-shivering thermogenesis in severe cold. Basal metabolic rate of 90 W is for young healthy males.110 Non-shivering thermogenesis is estimated at 27 W.111 Shivering thermogenesis (300 W) is based on work of Benzinger.14 Wet heat loss in cold/thermoneutrality is equal to ‘perspiratio insensibilis’ of about 10 W.112 In the heat, wet heat loss equals heat production since dry heat loss is negligible when skin temperature equals ambient temperature. The extra heat production of 30 W in the heat is based on recent measurements in Van Marken Lichtenbelt's lab, that will be submitted for publication. Dry heat loss in cold and thermoneutrality is equaled to the difference in the heat balance.
The prolonged skin and tissue vasoconstriction in the cold leads to reduced peripheral tissue temperatures. When the temperature in the fingers and toes becomes sufficiently low and when the core is sufficiently warm paradoxical cold induced vasodilation (CIVD) occurs likely through the opening of arterio-venous anastomoses.18 There is some evidence that this mechanism protects against cold injuries.19
The behavioral mechanisms of temperature regulation are complex. The basic adaptations are activity, body posture, and the creation and selection of a microclimate by means of clothing and shelters. We adapt our body posture to minimize heat loss. For instance, newborns show skin exposure of 79% when in thermal comfort and 69% when exposed to a cool environment.20 More specifically and more importantly in the cold, physical activity may increase to generate more heat. Clothing is selected in such a way that thermal equilibrium can be maintained. A population study showed that people tend to select clothing which insulation values corresponds to those estimated/or calculated to provide thermal equilibrium (ISO11079).21 This thermal need exceeds cultural aspects of clothing as has been shown in India, where the clothing style was shown to depend not only on cultural but also on climatic conditions.22 Finally shelter can be seeked for instance to reduce the effect of wind. In the heat, physical activity, clothing insulation and exposure to heat may be reduced, for instance by seeking shade. Clothing choice is also determined by the ability to reduce radiative heat strain.23 Adequate clothing systems and well insulated or heated homes and/or vehicles enable good protection against cold.
The behavioral and physiological responses to heat and cold should be considered as closely interwoven. There is evidence that the feeling of (dis)comfort drives the behavioral actions.24,25 Since the feeling of (dis)comfort is linked to skin temperature, also correlations between skin temperature and thermal behavior are observed.26
Comparison between populations
Before turning to cold adaptation, first a few words about differences between populations in heat coping mechanisms. A comparison between 10 morphological and physically matched tropical males (Malaysia) with 10 males from a moderate temperature region (Japan) showed that the tropical indigenes showed more stability in core temperature during heat exposure and less thirst.27 The temperature of the extremities during rest was lower in the tropical residents prior to heat exposure; heat was stored in the periphery of the body during exposure to heat so that the core temperature did not have to rise that much. We could not find studies that compared the ability to vasodilate in the heat for physically and morphologically matched black and white people. However, there are indications that some of the excellent skills to function in the heat, such as sweat loss, may have been reduced after people migrated to colder areas: it has been shown that people with a white skin have significantly less mixed apocrine-eccrine sweat glands than people with a black skin28 and that people living outside tropical areas have fewer sweat glands than people living in those areas.29 A brute force analysis of the genes of 862 subjects living at different areas of our planet showed that the genes ADRA2A and ADRA2C that may play a role in vasoconstriction in response to cold and pain stimuli, show increased presence in relatively cold areas.30 From these experiments it can be deduced that African people may have better heat loss mechanisms.
With respect to cold adaptation, 4 types of cold adaptation are distinguished: metabolic, insulative, hypothermic and the combination of the latter (insulative hypothermic).7,31 Mäkinen, however, confines herself to metabolic and insulative adaptation.8,32 Also, Sawka et al. mention only these 2 types of cold adaptation, but add the term ‘habituation’ that stands for blunted shivering, bunted cutaneous vasoconstrictor response, or both.9 Similarly, in a study on NST, insulative and metabolic responses were distinguished.33 Hypothermic adaptation can be considered as increased tolerance to cold without corresponding physiological changes. Therefore, we use the classification of Mäkinen in metabolic and insulative adaptation. It is suggested that the different types of cold adaptation responses are dependent on the degree of whole-body cooling and that metabolic adaptation occurs when it can compensate increased heat loss.34 If not, then insulative mechanisms tend to prevail.34 There are, however, considerable interindividual differences in the response to cold in which increased metabolism tends to dominate in lean subjects and insulative responses in obese subjects.33,35
Metabolic cold adaptation
If people indigenous to cold areas increase heat production in response to cold exposure to a larger extent than people from the tropics, they can be considered metabolically cold adapted. Metabolic adaptation, can go hand in hand with higher mean skin temperature without an increase in body core temperature.7 Metabolic cold adaptation is indeed present in people from temperate and arctic regions, since the shivering response to cold is considerably higher compared to African people. This, however, does not answer the question if those adaptations are phenotypic of genotypic. Adams and Covino36 subjected nude Negro, Caucasian and Eskimo males to an air temperature of 17°C. After 55 minutes of exposure, both the Caucasian and Eskimo subjects demonstrated an average rise in metabolism of 26 W/m2 body surface area above control levels of 46 and 64 W/m2, respectively. Negro subjects showed an increase in body heat production of only 12 W/m2 after 85 minutes in the cold room.
It can be argued that the discomfort associated with severe cold exposure triggers behavioral avoidance mechanisms and that therefore prolonged exposure to severe cold is more rare than prolonged exposure to mild cold3 Exposure to mild cold leads to NST.37 In rodents NST can be attributed to brown adipose tissue (BAT).38 In the meantime it is shown that also adult humans possess active cold inducible BAT,39-42 but far less than rodents.43 Brown fat combustion in humans seems to be an important heat source in NST,38,44 at least BAT activity is related to NST (e.g. ref 45). Brown fat is also present in people living in the tropics.46 Retrospective studies indicate that there are no differences in brown adipose tissue prevalence between black and whites47 Bakker et al.,48 however, showed that Asians living in Europe had lower BAT prevalence and exhibited a poorer shivering and non-shivering response to cold than Caucasians of similar age and BMI. On the other hand, subjects living in polar regions have higher BMI, and likely more white fat for body energy reserves and insulation.49 This cannot be explained by less exercise,50 but by body composition51 and food intake.49 It has been shown that more body fat is negatively related to NST44 and even that NST increases when losing weight.52 More studies are needed to elucidate differences in NST and BAT prevalence between populations.
However, there appear to be differences in basic metabolic rate (BMR) between populations. Although Rodahl observed no differences in BMR between Eskimos and whites53 more recent studies by Leonard et al.54 clearly show that indigenous circumpolar populations have 3–19 % higher BMR for males and females than predicted on the basis of people in temperate climates. When adjusted for body composition initially no differences in BMR were observed between Asians and Caucasians55 but later Bakker et al.48 observed that Caucasians had higher BMR values than Asians living in Europe. Black inhabitants of Nigeria had lower resting metabolic rate than black Americans in a study in 2002,56 but the same group observed no differences in another sample reported in the year 2000.57 Thus, it seems that whites and in particular indigenous circumpolar populations generate more heat than African natives at rest.
In summary, population studies indicate that people living in Africa, i.e. the Negroid population, have better heat loss (sweating) capacities, but have blunted metabolic responses to cold exposure. One may conclude that in the 40,000 y that people inhabit cold areas, part of the heat loss capability in the heat is lost and that a more effective ST is developed. For NST the number of studies are insufficient for conclusions. It seems that BMR of cold indigenous people is higher than tropical counterparts.
Insulative cold adaptation
Insulative adaptations are characterized by a lower skin temperature when exposed to cold with unchanged metabolic rates and core temperatures.7 This type of adaptation is shown in Aboriginals that sleep in cold deserts,58 but does not apply to bushmen in the Kalahari desert; the natives make sure that the fire is warm enough to create ambient temperatures that are acceptable and do not disturb night rest.59 Evidence for ethnic differences in insulative adaptation is scarce.
Research on human cold adaptation by anthropologists focused on the relation between body size and the physical environment and was mainly performed before 1975.10 It has been observed that the surface to volume ratio of people living in colder areas is lower, thus contributing to reduced heat loss.60,61 The magnitude is about 6 cm2/kg body weight for 10 degrees difference in latitude. It has been shown, however, that nutrition patterns interfere with this relationship:62 the relationship between body surface area and ambient temperature was much better in 195363 than in 199862 since more people adopted the western food style leading to body size changes.
It is assumed that European Neanderthals were well equipped with cold defense mechanisms. Apart from metabolic adaptations, including brown adipose tissue for NST, they are supposed to have had insulative adaptation, such as strong vasoconstriction in the skin, hairy skin to reduce heat loss, localized cold-induced vasodilation and cold shock proteins.64 It is predicted that some Neanderthals would have covered up to 80% of their body.65 There is some evidence that the Neanderthals, who lived in Europe 180,000 y ago until extinction 30,000 y ago,66 exchanged DNA with the modern Cro-Magnon human.67 This may have led to the presence of the aforementioned cold defense mechanisms (but not a hairy skin) currently found in Inuit and Caucasians that are present to a lesser extent in black African people nowadays.
It is well-documented that reactions to local cold, like immersion of hands in cold water, show considerable differences between blacks and whites. Several studies indicate that Negroes show slower and reduced CIVD responses.68-70 Purkayastha et al. showed that subjects living in polar regions had superior CIVD responses over tropical natives.71,72
In summary, it is not possible to conclude from the available studies if differences in passive (subcutaneous fat) and active (vasoconstriction of the skin and peripheral tissues) insulation exist between different populations in the world. Cold induced vasodilation is more pronounced in cold indigenous populations.
Acclimation to cold
Cold acclimation procedures
There are different levels of whole body cold acclimation, depending on the degree of the cold exposure. Two levels can be distinguished: 1. Severe cold exposure, leading to a drop in mean skin, tissue and body core temperature generally evoked using repeated cold water immersions, 2. Moderate cold exposure, leading to decrease in skin and tissue temperatures with no or minor drop in body core temperature, generally evoked using repeated cold air exposure. While it is undisputed that humans have excellent mechanisms to acclimatize to heat,73-78 the acclimation capabilities to cold are a topic of controversy (e.g.,79). Next to whole body acclimation, the changes in peripheral (hands, feet) blood flow are also a matter of dispute.80
Acclimation to repeated severe cold exposure
Table 1 summarizes the studies regarding the effect of repeated cold exposure on metabolism, thermal insulation and peripheral blood flow (the CIVD response).
Table 1.
Studies regarding the effect of repeated cold water immersion on metabolism, thermal insulation and peripheral blood flow (cold induced vasodilation (CIVD)).
| Authors | Year | #subjects | #immersion days | duration (min) | water temp. (°C) | exercise | metabolism change | insulation change | CIVD |
|---|---|---|---|---|---|---|---|---|---|
| Young et al. | 1986 | 7M | 25 | 90 | 18 | none | Initial 20% drop (10 min) during cold air exposure | increased Tre-Tsk gradient | |
| Bittel | 1987 | 9M | 32–40 | 60–180 | 10–15 | none | no change | increased Tre-Tsk gradient | |
| Budd et al. | 1993 | 6M | 10 | 30–60 | 15 | none | 12% reduction | increased tissue insulation | inconclusive |
| Jansky et al. | 1995 | 26M | 12–18 | 60 | 14 | none | 23% reduction at end of immersion | no change | |
| Golden&Tipton | 1998 | 16M | 10 | 40 | 15 | 8 exercise, 8 none | 22% reduction at Tb of 33°C | no change | |
| O'Brien et al. | 2000 | 14M | 25 | 60 | 20 | 7 exercise, 7 none | no change in cold water or cold air | no change | Reduced |
| Stocks et al. | 2001 | 7M | 15 | 90 | 18 | none | 18% reduction, no difference during exercise in pre- and posttest | no change | |
| Zeyl et al. | 2004 | 12M | 15 | 60–90 | 18 | none | not measured | no change | |
| Wakabayashi et al. | 2012 | 7M | 12 | 60 | 26 | none | no difference | no change | Reduced |
| Tipton et al. | 2013 | 7M | 5 | 45 | 12 | none | 29% reduction for ΔTre <1.2°C | no change | |
| Brazaitis et al. | 2014 | 14M | 17 | 170 or Tre <35.5°C | 14 | none | 22% reduction | no change |
Note: In the study of Bittel, subjects were wearing neoprene suits.
Metabolic acclimation
Repeated exposure to severe cold to investigate the effects of metabolic adaptations is generally performed in cold water. Water is a good medium for cold exposure since it conducts heat much better than air and has a high thermal capacity. Water temperatures in the studies range from 1281 to 26°C.82 Most studies observe a paradoxical reduction in total metabolism, generally measured using analysis of expired air (Table 1). Two studies show no reduced metabolism upon repeated cold exposure, which in one case may be attributed to the relatively high water temperature.82 The other study observed no differences in metabolism for cold water exposure after 25 cold water immersions.83 All other studies reported decreases in metabolic rate of about 20% due to repeated cold water immersions.79,81,84-88 Brazaitis et al. immersed 14 male subjects 17 consecutive days in 14°C for 170 minutes or until core temperature dropped below 35.5°C.79 The latter occurred more often and the duration until this temperature was reached dropped from about 130 minutes to 80 minutes in 13 d. Since cold water immersion is uncomfortable and since the subjects may have noticed that they were rewarded with a lift out of the cold water when the core temperature reached 35.5°C, it cannot be excluded that they intentionally reduced shivering thermogenesis. Thermogenesis can be influenced by many factors through the sympathetic system including mental status.89 However, it is obvious that these adaptations to repeated severe cold exposure do not contribute to survivability in the cold. Tipton et al. showed that the reduced metabolic response was confined to mild cooling only.81 They immersed 7 subjects 5 consecutive days in 12°C water for 45 minutes and compared the results before and after this intervention. When rectal temperature dropped more than 1°C differences in metabolism disappeared. This points at a protective increase in metabolism when core temperature becomes too low. Stocks et al. immersed 7 males for 15 d in 18°C water and also observed a drop in metabolism.88 Generally, cold adaptation is investigated in passive subjects, i.e., with the lowest possible metabolism to represent the worst case in the cold. Some studies also investigated the effect of repeated cold exposure on exercise performance in the cold. It is observed that during exercise following cold exposure, the differences in metabolism disappeared.88 This is in line with earlier observations reviewed by Shephard90 and with earlier research of Golden and Tipton.87 In summary, repeated severe cold exposure leads to reduced metabolism, in particular when the body core does not become too cold.
Insulative acclimation
Table 1 shows that increased insulation was only observed in 3 studies. Young et al. observed an increased gradient between rectal and mean skin temperature,84 a finding that could not be reproduced.83 Bittel observed a decrease in heat debt from 8.4 to 5.7 kJ/kg due to repeated cold water immersion.35 The third study observed increased vasoconstriction in the peripheral tissue, not the skin, after repeated cold water immersions.85 In summary, it seems that repeated cold water immersions hardly leads to increased thermal insulation.
Cooler skin temperatures were observed in 10 adult males during the initial month in Antarctica when exposed to a standard cold stress of 10°C for 2 hours as compared to results in Australia. This type of adaptation is called insulative adaptation. It is however difficult to exclude the effect of fatter food in Arctic and Antarctic areas on skin insulation in this type of research. This confounder was not present in a nice study of Imamura.91 She exposed Japanese women to the cold Japanese winter with mini-skirts or long skirts. In mini-skirts the legs were more exposed to cold than in long skirts. Using MRI she showed that the subjects with mini-skirts developed more fat at the legs at the end of the winter period. The studies did not report on changes in vasoconstriction capacity, influencing the active insulative properties of the body.
Other changes due to repeated cold exposure
Repeated exposure to cold alters the activity of several proteins in the body.92 Cold stress changes the lipid composition of cellular membranes, and suppresses the rate of protein synthesis and cell proliferation.93 Although most studies dealt with tissue temperatures below 20°C, mild hypothermia (32°C) can change the cell's response to subsequent stresses as well. For example, cold exposed rats increase the expression of constitutive and inducible members of the HSP70 gene family in association with increased expression of the mitochondrial uncoupling protein in brown adipose tissue (BAT).94 Thus, protein changes due to cold exposure can lead to increased metabolism. Cold shock proteins may prevent cell damage. Therefore, controlled studies investigating the effect of repeated cold exposure on the prevalence of cold shock proteins can attribute to reduced problems in the cold. Unfortunately, most efforts are dedicated to marine animals.
It is a general observation in all studies reported in Table 1 that the discomfort associated with cold exposure declined. In particular the initial response to cold water immersion blunted, a phenomenon called habituation and also observed after repeated showers.95 Shivering is uncomfortable and humans like to avoid discomfort. It is not unlikely that the reduced shivering response in severe cold acclimation may be attributed to avoidance of discomfort. Apart from heat production, shivering also increases the heat loss in cold water due to increased convection and may thus be undesired. Finally, the heat generating effect of shivering is relatively minor to the heat loss in cold water. Reducing shivering in cold water may not be so disastrous at all.
Acclimation to repeated moderate cold exposure
When unacclimatized rodents are placed in a cold environment, they will defend their body temperature by shivering. However, after prolonged exposure to cold, shivering will decrease while energy expenditure remains elevated.96 Shivering can under those circumstances be fully replaced by NST.38,96 In rodents the increase in NST is ascribed to brown adipose tissue (BAT).38 Already in 1961 a study by Davis showed that also humans that are repeatedly exposed to cold (12°C, 8 h/day, 31 days) show a gradual decrease in shivering while heat production remained elevated.97 At that time the tissue responsible for NST was not identified. In the meantime it is shown that also adult humans possess active cold inducible BAT.40-42 The main function of BAT is heat production (NST) and is activated when the body is exposed to cold. Cold acclimation studies show seasonal variation in NST even in the temperate climate zone.98 Interestingly, cold-activated BAT also shows seasonal variation40 and is related to outdoor temperatures.99
Recently several studies have been performed that used moderate cold acclimation to study the effect on BAT and energy metabolism (Table 2). A recent acclimation study in healthy males and females, showed that NST increased after a relatively short period of acclimation. In a 10-day acclimation study (15°C air, 6 h/d) NST increased significantly from 11% to 18 % and was also accompanied with an increase in BAT presence and activity.45 During the 10 day acclimation the subjects onset of shivering was delayed. Shivering intensity was not measured. The increase in NST after cold acclimation was confirmed in a study in type 2 diabetes.100 Another study showed that daily cold exposure of 2 hours per day at 17°C for 6 weeks also resulted in an increase in cold induced thermogenesis accompanied with an increase in BAT, and with a significant decrease in (white) fat mass.101 A study from Blondin et al. (2014), using more severe cold (10°C) during acclimation and measurements indicated an increase of cold induced oxidative capacity after a 4 week cold acclimation period.102 In this study it was not possible to discriminate between shivering and NST. Finally Lee et al. performed a cold acclimation test, by exposing the subjects to overnight cool room (19°C) for a month (5 d per week) compared to a month sleeping under thermoneutral (24°C) conditions.103 They observed increases in BAT activity and total energy expenditure, but no significant change in NST. All in all the results seem to depend on the level of control of the cold exposure and the measurement protocol. When the protocol is designed to measure NST, the results clearly show an increase in NST after acclimation. The studies also make it likely that the shivering response decreases.
Table 2.
Studies regarding the effect of mild cold exposure on metabolism and thermal insulation.
| Authors | Year | #subjects | #days | duration (h) | air temp. (°C) | BAT | metabolism | insulation |
|---|---|---|---|---|---|---|---|---|
| Davis | 1961 | 10M | 31 | 8 | 13.5 | NA | no change in EE; decrease in shivering | NA |
| Lans, van der et al. | 2013 | 9F8M | 10 | 6 | 14–15 | increase | increase NST (ΔNST: 75%) | decrease Tco-Tsk gradient; decrease skin perfusion |
| Yoneshiro et al | 2013 | 51M | 42 | 2 | 19 | increase | increase NST (ΔNST: 150%-estimated from bar graph) | NA |
| Chen et al./Lee et al. | 2013/2014 | 10F14M | 42 | 12 (overnight) | 19 | increase | increase in EE; no change in NST | NA |
| Blondin et al.* | 2014 | 6 | 28 (5d/wk) | 2 | 10* | increase | shivering intensity did not change; increase BAT oxidative capacity | NA |
| Hanssen et al. | 2015 | 8 | 10 | 6 | 14–15 | increase | increase NST | NA |
in between severe and moderate cold acclimation; use of liquid-conditioned suit.
The contribution of NST is small compared to ST. NST ranges from zero to 30% of RMR,104 depending on body composition, age and level of acclimation, while with ST the amount of heat production can be 4 times higher than resting metabolic rate. Also the reported effects of acclimation of NST capacity are limited. In the study of van der Lans et al. in healthy young males NST changed from 10.6 ± 3.2 to 20.3±12.0 Watt and in females from 11.0 ± 9.8 to 15.9 ± 10.7 Watt.45 Although this is relatively small in magnitude, it is important to note that the extra heat can be sustained over a long time period. This in contrast to shivering that declines when glycogen storage deplete,16 affects coordination and leads to fatigue and discomfort. These recent studies line up with the classical study by Scholander et al. When eight males were exposed to the cold Norwegian environment for several days with minimal thermal protection, metabolism was found to be considerable higher than before and then in controls.105
As can be seen in Table 2, not many studies on insulation have been performed. Two studies show a decrease in insulation, based on core-skin temperature gradients, and one study also shows a greater decrease in skin perfusion.
In conclusion, it seems that long exposures to cold air may give metabolic enhancements, in contrast to relatively short extreme cold stress of cold water immersion. Also the insulative response seems to be increased after moderate cold acclimation although the number of studies supporting this observation is small.
Local cold acclimation
The effects of local cold acclimation is recently reviewed.73 It was concluded that systematic improvements in finger and hand blood flow in the cold are neither guaranteed nor predictable, and individuals should not rely on improving peripheral cold tolerance through repeated cold exposure. The work of Nelms and Soper in which it was shown that fish filleters have better hand blood flow than controls, led to the long lasting misunderstanding that this was due to acclimation.96 The possibility that only people with warm hands take up the challenge to become fish filleter was not considered. Recent research in which 16 subjects immersed one hand and one foot in 8°C water for 15 d showed that pain became less every day (similar to the reduced discomfort observed in whole body cold immersions) but finger temperatures did not increase.97
Discussion
As can be seen in Table 1 and 2, data is generally lacking on females. Only two studies on moderately cold acclimation were carried out in both males and females.45,101 In studies with female subjects the monthly changes in core temperature have to be taken into account, which probably forms a major hurdle for the investigators. Also, the experiments are limited to healthy (exception100) young males of Caucasian ascent. It would have been interesting to have studies on acclimation to cold in the Negroid population since they have reduced mechanisms to cope with the cold and would benefit more when beneficial adaptations to repeated cold exposure would occur.
People that have to perform in the cold, either for sports or occupational, request guidelines about how to prepare for the event or work period. The review shows that repeated exposure to severe cold (e.g. cold water immersions) does not lead to beneficial physiological adaptations, but that repeated mild cold exposure may lead to increased non shivering thermogenesis. An interesting recent observation showed that time trial performance in cool circumstances increased after heat acclimation.106 After a 10 day period of heat acclimation, time trial performance of 12 cyclists improved by 8% in the heat (38°C, 30% RH) and 7% in a cool climate (13°C, 30% RH). Although more research is needed, it shows a unexpected alternative for repeated cold exposures. The review shows that repeated cold exposure leads to habituation; less pain or discomfort. The feeling of pain and discomfort requires attention that cannot be dedicated to the task. Therefore, cognitive performance in the cold is reduced.107There are some indications that habituation leads to improved cognitive performance in the cold 108 since the focus can be dedicated to the task instead of the discomfort.
In general, severe cold exposure leads to a reduced metabolism and the data on the insulative adaptations are not clear. Moderate cold acclimation results in increase in NST capacity and some studies indicate an improved insulative response. The adaptation so far is limited, but to our knowledge no studies on maximal NST capacity have been performed in humans.
Both for whole body cold acclimation studies and peripheral adaptations to cold, the field is confused by a lack of standardization in definitions and methodology. Differences in exposure duration, cooling medium, measuring methods and subject selection for instance, make it difficult to draw general conclusions.
Time course of responses to cold
The preceding analysis shows that repeated exposure to either mild or severe, total or local cold leads to perceptual and/or physiological changes that are not occurring simultaneously. The change in subjective responses occurs during the initial days of severe cold exposure. Brazaitis et al.79 showed that shivering sensation and comfort sensation was already blunted the second day; significant changes in thermal sensation occurred started at day 6. Similarly, pain sensation of an immersed hand and foot decreased exponentially.109 The first day a drop in pain score of 4.5 to 4 was observed, while pain score remained stable at 2.5 after about 5 d. The habituation response seems to precede the physiological changes in metabolism and certainly in thermal insulation although it is shown above that these responses are often not consistent. Acclimation studies during moderate cold reveal a gradual decrease of discomfort during the actual exposure.45 In contrast to heat acclimation studies, no information is available regarding the time course of changes when the repeated cold exposure is discontinued.
Recommendations for future research
Scientific studies on human cold adaptation are scarce in particular when compared to heat adaptation. At this moment, it is not possible to establish solid guidelines on how to prepare for extended stay, work or sport in the cold since there is insufficient scientific evidence available. There are indications that repeated exposure to mild cold may lead to increased thermogenesis, probably related to enhanced combustion by brown fat that go hand in hand with a decrease in thermal discomfort. On the other hand, repeated exposure to strenuous cold, i.e. cold water immersion, seems to lead to reduced heat production over time. Studies should be performed in order to validate those claims using larger numbers of subjects, investigate if gender, age and ethnicity play a role in these adaptations and if indeed the intensity of cold exposure is related to different forms of adaptation. In those studies, it is recommended not only to focus on metabolism, but also take insulative changes into account. This concerns both passive insulation quantified by the thickness of the subcutaneous fat layer and dynamic insulation quantified by the core-to-shell temperature difference.
In particular, studies are lacking that investigate local cold adaptation in vulnerable populations like elderly and people indigenous to warm areas of the world. What happens when the latter move from the tropics to cold areas in the world? How should we categorize ethnicity in this respect? What is the practical impact of the gain in non-shivering thermogenesis due to repeated exposure to mild cold? Can we increase cold-shock proteins due to repeated cold exposure and how does this translate to cold acclimation?
Only when those questions are answered, we can establish guidelines on how to deal with cold exposure in daily life, prepare for cold expeditions, work in the cold, or athletic events in the cold. It may also learn us to what extent humans are able to survive cold spells that may occur in future years.
Conclusions
Cold adaptation responses are related to exposure type, intensity and duration, but also to various individual factors (e.g., age, gender, ethnicity, exercise, diet, medication etc.) which affect thermal balance in cold. Current reviews on cold adaptation 7-10 lack a clear distinction between population differences and between adaptations acquired during life time of a single subject. This leads to confusion. This review shows that considerable differences exist in response to cold between black Africans and Caucasians/Inuit. The latter have a more pronounced shivering response and better protective mechanisms against cold injuries of the hand and feet. These changes may be the result of genetic material handed over by the Neanderthals. If, however, we look at the effect of repeated cold exposure on physiological responses to cold in the life time of a subject, the results are less conclusive. It seems that repeated severe cold exposure (cold water immersion) leads to reduced metabolism, but repeated exposure to cold air may enhance metabolism. Some degree of insulative adaptation occurs due to repeated whole body cold exposure, but it is different to exclude the effect of a fatter diet in cold areas. The feeling of discomfort and pain subsides both in whole body cold exposure and local cold exposure, leading to altered behavior. Humans have developed excellent clothing, houses and behavioral adaptations to cold, and these seem to be tremendously more important for living under extreme conditions than our physiological mechanisms alone.
Footnotes
The terms adaptation, acclimatization, acclimation and habituation are used in line with the Glossary of terms for thermal physiology.[11] Adaptation relates to changes that reduce the physiological strain produced by stressful components of the total environment. This change may occur within the lifetime of an organism (phenotypic) or be the result of genetic selection in a species or subspecies (genotypic). Acclimati(zati)on relates to phenotypic adaptations to specified climatic components. Acclimatization refers to the natural climate while acclimation refers to an artificial climate (e.g. climatic chambers). Habituation is the reduction of responses to or perception of a repeated stimulation. The term ‘acclimation’ will be used when both natural and artificial environments may apply.
Abbreviations
- BMI
Body Mass Index
- BMR
Basic Metabolic Rate
- CIVD
Cold Induced Vasodilation
- NST
Non-shivering thermogenesis
- RH
Relative Humidity
- ST
Shivering thermogenesis
- Tb
Mean body temperature
- Tre
Rectal temperature
Disclosures of potential conflicts of interest
The authors declare no potential conflicts of interest.
References
- [1].Diamond J. Guns, Germs, and Steel. W.N.Norton, ISBN 0-393-03891-2, 1997. [Google Scholar]
- [2].Thompson LG, Mosley-Thompson E, Davis ME, Henderson KA, Brecher HH, Zagorodnov VS, Mashiotta TA, Lin P, Mikhalenko VN, Hardy DR, et al.. Kilimanjaro ice core records: evidence of holocene climate change in tropical Africa. Science 2002; 298:589-93; PMID:12386332; http://dx.doi.org/ 10.1126/science.1073198 [DOI] [PubMed] [Google Scholar]
- [3].LeBlanc J, Man in the Cold, ISBN 0-398-03429-X ed., Thomas CC, 1975. [Google Scholar]
- [4].Huynen MMTE, Martens P. Climate change effects on heat- and cold-related mortality in the Netherlands: a scenario-based integrated environmental health impact assessment. Int J Environ Res Public Health 2015; 12:13295-320; PMID:26512680; http://dx.doi.org/ 10.3390/ijerph121013295 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Bestion E, Teyssier A, Richard M, Clobert J, Cote J. Live fast, die young: experimental evidence of population extinction risk due to climate change. PloS Biol 2015; 13:1-19; http://dx.doi.org/ 10.1371/journal.pbio.1002281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Hansen B, Østerhus S, Quadfasel D, Turrell W. Already the day after tomorrow? Science 2004; 305:953-4; PMID:15310882; http://dx.doi.org/ 10.1126/science.1100085 [DOI] [PubMed] [Google Scholar]
- [7].Launay J, Savourey G. Cold adaptations. Ind Health 2009; 47:221-7; PMID:19531907; http://dx.doi.org/ 10.2486/indhealth.47.221 [DOI] [PubMed] [Google Scholar]
- [8].Mäkinen TM. Human cold exposure, adaptation, and performance in high latitude environments. Am J Hum Biol 2007; 19:155-64; http://dx.doi.org/ 10.1002/ajhb.20627 [DOI] [PubMed] [Google Scholar]
- [9].Sawka MN, Castellani JW, Pandolf KB, Young AJ.. Human adaptations to heat and cold stress, Symposium on “Blowing Hot and Cold: Protecting Against Climatic Extremes” Dresden, NATO RTO-MP-076 2001; KN4-1-KN4-15. [Google Scholar]
- [10].Steegmann AT., Jr. Human cold adaptation: an unfinished agenda. Am J Hum Biol 2007; 19:218-27; PMID:17286254; http://dx.doi.org/ 10.1002/ajhb.20614 [DOI] [PubMed] [Google Scholar]
- [11].T.C. IUPS, Glossary of Terms for Thermal Physiology. Second Edition . Revised by the commission for thermal physiology of the international union of physiological sciences (IUPS thermal commission). Pflugers Arch 1987; 410:567-87; PMID:3324054; http://dx.doi.org/ 10.1007/BF00586542 [DOI] [PubMed] [Google Scholar]
- [12].Taylor NAS. Human heat adaptation. Compr Physiol 2014; 4:325-65; PMID:24692142; http://dx.doi.org/ 10.1002/cphy.c130022 [DOI] [PubMed] [Google Scholar]
- [13].Claessens-Van Ooijen AMJ, Westerterp KR, Wouters L, Schoffelen PFM, Van Steenhoven AA, Van Marken Lichtenbelt WD.. Heat production and body temperature during cooling and rewarming in overweight and lean men. Obesity 2006; 14:1914-20; PMID:17135606; http://dx.doi.org/ 10.1038/oby.2006.223 [DOI] [PubMed] [Google Scholar]
- [14].Benzinger TH. Heat regulation: homeostasis of central temperature in man. Physiol Rev 1969; 49:671-759; PMID:4898601 [DOI] [PubMed] [Google Scholar]
- [15].Vybíral S, Lesná I, Janský L, Zeman V.. Thermoregulation in winter swimmers and physiological significance of human catecholamine thermogenesis. Exp Physiol 2000; 85:321-6; http://dx.doi.org/ 10.1111/j.1469-445X.2000.01909.x [DOI] [PubMed] [Google Scholar]
- [16].Martineau L, Jacobs I.. Muscle glycogen utilization during shivering thermogenesis in humans. J Appl Physiol 1988; 65:2046-50; PMID:3209549 [DOI] [PubMed] [Google Scholar]
- [17].Strydom NB, Wyndham CH, Williams CG, Morrison JF, Bredell GAG, Benade AJS, Von Rahden M.. Acclimatization to humid heat and the role of physical conditioning. J Appl Physiol 1966; 21:636-42; PMID:5934473 [DOI] [PubMed] [Google Scholar]
- [18].Daanen HAM. Finger cold-induced vasodilation: a review. Eur J Appl Physiol 2003; 89:411-26; PMID:12712346; http://dx.doi.org/ 10.1007/s00421-003-0818-2 [DOI] [PubMed] [Google Scholar]
- [19].Daanen HAM, Van Der Struijs NR. Resistance index of frostbite as a predictor of cold injury in arctic operations. Aviat Space Environ Med 2005; 76:1119-22; PMID:16370261 [PubMed] [Google Scholar]
- [20].Stothers JK, Warner RM.. Thermal balance and sleep state in the newborn. Early Hum Dev 1984; 9:313-22; PMID:6745152; http://dx.doi.org/ 10.1016/0378-3782(84)90076-8 [DOI] [PubMed] [Google Scholar]
- [21].Donaldson GC, Rintamäki H, Näyhä S.. Outdoor clothing: its relationship to geography, climate, behaviour and cold-related mortality in Europe. Int J Biometeorol 2001; 45:45-51; PMID:11411415; http://dx.doi.org/ 10.1007/s004840000072 [DOI] [PubMed] [Google Scholar]
- [22].Chatterjee SK. The pattern of indian clothing in relation to tropical climate. J Hum Evol 1978; 7:95-99; http://dx.doi.org/ 10.1016/S0047-2484(78)80040-2 [DOI] [Google Scholar]
- [23].Shkolnik A, Taylor CR, Finch V, Borut A. Why do bedouins wear black robes in hot deserts? Nature 1980; 283:373-5; http://dx.doi.org/ 10.1038/283373a0 [DOI] [Google Scholar]
- [24].Taylor NAS, Allsopp NK, Parkes DG. Preferred room temperature of young vs aged males: the influence of thermal sensation, thermal comfort, and affect. J Gerontol - Series A Biol Sci Med Sci 1995; 50:M216-21; http://dx.doi.org/ 10.1093/gerona/50A.4.M216 [DOI] [PubMed] [Google Scholar]
- [25].Flouris AD. Functional architecture of behavioural thermoregulation. Eur J Appl Physiol 2011; 111:1-8; PMID:20711785; http://dx.doi.org/ 10.1007/s00421-010-1602-8 [DOI] [PubMed] [Google Scholar]
- [26].Jacquot CMC, Schellen L, Kingma BR, van Baak MA, Van Marken Lichtenbelt WD, Influence of thermophysiology on thermal behavior: the essentials of categorization. Physiol Behav 2014; 128:180-7; http://dx.doi.org/ 10.1016/j.physbeh.2014.01.025 [DOI] [PubMed] [Google Scholar]
- [27].Lee J, Wakabayashi H, Wijayanto T, Hashiguchi N, Saat M, Tochihara Y.. Ethnic differences in thermoregulatory responses during resting, passive and active heating: application of werner's adaptation model. Eur J Appl Physiol 2011; 111:2895-905; PMID:21437607; http://dx.doi.org/ 10.1007/s00421-011-1912-5 [DOI] [PubMed] [Google Scholar]
- [28].Montagna W, Carlisle K. The architecture of black and white facial skin. J Am Acad Dermatol 1991; 24:929-37; PMID:1714469; http://dx.doi.org/ 10.1016/0190-9622(91)70148-U [DOI] [PubMed] [Google Scholar]
- [29].Kuno Y. Human Perspiration. CT Thomas, 1956 [Google Scholar]
- [30].Raj SM, Pagani L, Gallego Romero I, Kivisild T, Amos W. A general linear model-based approach for inferring selection to climate. BMC Genet 2013; 14:PMID:24053227; http://dx.doi.org/ 10.1186/1471-2156-14-87 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Ito S. Adaptation to cold: general concept of cold adaptation. Hokkaido Igaku Zasshi 1975; 50:123-32; PMID:1238342 [PubMed] [Google Scholar]
- [32].Mäkinen TM. Different types of cold adaptation in humans. Front Biosci - Scholar 2010; 2 S:1047-67; http://dx.doi.org/ 10.2741/S117 [DOI] [PubMed] [Google Scholar]
- [33].Van Marken Lichtenbelt WD, Schrauwen P, Van De Kerckhove S, Westerterp-Plantenga MS. Individual variation in body temperature and energy expenditure in response to mild cold. Am J Physiol Endocrinol Metab 2002; 282:E1077-83; PMID:11934673; http://dx.doi.org/ 10.1152/ajpendo.00020.2001 [DOI] [PubMed] [Google Scholar]
- [34].Young AJ. Homeostatic responses to prolonged cold exposure: human cold Acclimatization. in Section 4 Environmental Physiology, Fregly MJ, Blatteis CM, eds., Oxford University Press, Oxford, UK, 1996. [Google Scholar]
- [35].Bittel J, The different types of general cold adaptation in Man, Int J Sports Med 1992; 13:S172-6; PMID:1483765; http://dx.doi.org/ 10.1055/s-2007-1024630 [DOI] [PubMed] [Google Scholar]
- [36].Adams T, Covino BG.. Racial variations to a standardized cold stress. J Appl Physiol 1958; 12:9-12; PMID:13502249 [DOI] [PubMed] [Google Scholar]
- [37].Janský L. Non-shivering thermogenesis and its thermoregulatory significance. Biol Rev Camb Philos Soc 1973; 48:85-132; PMID:4578360; http://dx.doi.org/ 10.1111/j.1469-185X.1973.tb01115.x [DOI] [PubMed] [Google Scholar]
- [38].Cannon B, Nedergaard J.. Brown adipose tissue: function and physiological significance. Physiol Rev 2004; 84:277-359; PMID:14715917; http://dx.doi.org/ 10.1152/physrev.00015.2003 [DOI] [PubMed] [Google Scholar]
- [39].Nedergaard J, Bengtsson T, Cannon B. Unexpected evidence for active brown adipose tissue in adult humans. Am J Physiol Endocrinol Metab 2007; 293:E444-52; PMID:17473055; http://dx.doi.org/ 10.1152/ajpendo.00691.2006 [DOI] [PubMed] [Google Scholar]
- [40].Saito M, Okamatsu-Ogura Y, Matsushita M, Watanabe K, Yoneshiro T, Nio-Kobayashi J, Iwanaga T, Miyagawa M, Kameya T, Nakada K, Kawai Y, Tsujisaki M. High incidence of metabolically active brown adipose tissue in healthy adult humans: effects of cold exposure and adiposity. Diabetes 2009; 58:1526-31; PMID:19401428; http://dx.doi.org/ 10.2337/db09-0530 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [41].Virtanen KA, Lidell ME, Orava J, Heglind M, Westergren R, Niemi T, Taittonen M, Laine J, Savisto N, Enerbäck S, Nuutila P. Functional brown adipose tissue in healthy adults. New Engl J Med 2009; 360:1518-25; http://dx.doi.org/ 10.1056/NEJMoa0808949 [DOI] [PubMed] [Google Scholar]
- [42].Van Marken Lichtenbelt WD, Vanhommerig JW, Smulders NM, Drossaerts JMAFL, Kemerink GJ, Bouvy ND, Schrauwen P, Teule GJJ.. Cold-activated brown adipose tissue in healthy Men. New Engl J Med 2009; 360:1500-8; http://dx.doi.org/ 10.1056/NEJMoa0808718 [DOI] [PubMed] [Google Scholar]
- [43].Cypess AM, Lehman S, Williams G, Tal I, Rodman D, Goldfine AB, Kuo FC, Palmer EL, Tseng Y, Doria A, et al.. Identification and importance of brown adipose tissue in adult humans. New Engl J Med 2009; 360:1509-17; http://dx.doi.org/ 10.1056/NEJMoa0810780 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].van Marken Lichtenbelt WD, Schrauwen P. Implications of nonshivering thermogenesis for energy balance regulation in humans. Am J Physiol Regul Integr Comp Physiol 2011; 301:R285-R296; PMID:21490370; http://dx.doi.org/ 10.1152/ajpregu.00652.2010 [DOI] [PubMed] [Google Scholar]
- [45].Van Der Lans AAJJ, Hoeks J, Brans B, Vijgen GHEJ, Visser MGW, Vosselman MJ, Hansen J, Jörgensen JA, Wu J, Mottaghy FM, et al.. Cold acclimation recruits human brown fat and increases nonshivering thermogenesis. J Clin Invest 2013; 123:3395-403; PMID:23867626; http://dx.doi.org/ 10.1172/JCI68993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Nnodim JO. The occurrence of brown adipose in man inhabiting the tropics. Z Mikrosk -Anat Forsch Abt 2 1990; 104:721-8 [PubMed] [Google Scholar]
- [47].Perkins AC, Mshelia DS, Symonds ME, Sathekge M. Prevalence and pattern of brown adipose tissue distribution of 18F-FDG in patients undergoing PET-CT in a subtropical climatic zone. Nucl Med Commun 2013; 34:168-74; PMID:23196673; http://dx.doi.org/ 10.1097/MNM.0b013e32835bbbf0 [DOI] [PubMed] [Google Scholar]
- [48].Bakker LEH, Boon MR, van der Linden RAD, Arias-Bouda LP, van Klinken JB, Smit F, Verberne HJ, Jukema JW, Tamsma JT, Havekes LM, et al.. Brown adipose tissue volume in healthy lean south asian adults compared with white caucasians: a prospective, case-controlled observational study. Lancet Diabetes Endocrinol 2014; 2:210-7; PMID:24622751; http://dx.doi.org/ 10.1016/S2213-8587(13)70156-6 [DOI] [PubMed] [Google Scholar]
- [49].Kuhnlein HV, Receveur O, Soueida R, Egeland GM.. Arctic indigenous peoples experience the nutrition transition with changing dietary patterns and obesity. J Nutr 2004; 134:1447-53; PMID:15173410 [DOI] [PubMed] [Google Scholar]
- [50].Hopping BN, Erber E, Mead E, Roache C, Sharma S. High levels of physical activity and obesity co-exist amongst inuit adults in arctic Canada. J Hum Nutr Diet 2010; 23:110-4; PMID:21158969; http://dx.doi.org/ 10.1111/j.1365-277X.2010.01096.x [DOI] [PubMed] [Google Scholar]
- [51].Charbonneau-Roberts G, Saudny-Unterberger H, Kuhnlein HV, Egeland GM. Body mass index may overestimate the prevalence of overweight and obesity among the inuit. Int J Circumpolar Health 2005; 64:163-9; PMID:15945286; http://dx.doi.org/ 10.3402/ijch.v64i2.17969 [DOI] [PubMed] [Google Scholar]
- [52].Vijgen GHEJ, Bouvy ND, Teule GJJ, Brans B, Hoeks J, Schrauwen P, Van Marken Lichtenbelt WD. Increase in brown adipose tissue activity after weight loss in morbidly obese subjects. J Clin Endocrinol Metab 2012; 97:E1229-33; PMID:22535970; http://dx.doi.org/ 10.1210/jc.2012-1289 [DOI] [PubMed] [Google Scholar]
- [53].Rodahl K. Basal metabolism of the eskimo. J Nutrit 1952; 48:359-68. [DOI] [PubMed] [Google Scholar]
- [54].Leonard WR, Sorensen MV, Galloway VA, Spencer GJ, Mosher MJ, Osipova L, Spitsyn VA. Climatic influences on basal metabolic rates among circumpolar populations. Am J Human Biol 2002; 14:609-20; http://dx.doi.org/ 10.1002/ajhb.10072 [DOI] [PubMed] [Google Scholar]
- [55].Wulan SN, Westerterp KR, Plasqui G. Ethnic differences in body composition and the associated metabolic profile: a comparative study between asians and caucasians. Maturitas 2010; 65:315-9; PMID:20079586; http://dx.doi.org/ 10.1016/j.maturitas.2009.12.012 [DOI] [PubMed] [Google Scholar]
- [56].Luke A, Durazo-Arvizu RA, Rotimi CN, Iams H, Schoeller DA, Adeyemo AA, Forrester TE, Wilks R, Cooper RS. Activity energy expenditure and adiposity among black adults in nigeria and the united states. Am J Clin Nutr 2002; 75:1045-50; PMID:12036811 [DOI] [PubMed] [Google Scholar]
- [57].Luke A, Rotimi CN, Adeyemo AA, Durazo-Arvizu RA, Prewitt TE, Moragne-Kayser L, Marders R, Cooper RS. Comparability of resting energy expenditure in Nigerians and U.S. blacks. Obes Res 2000; 8:351-9; PMID:10968726; http://dx.doi.org/ 10.1038/oby.2000.42 [DOI] [PubMed] [Google Scholar]
- [58].Scholander PF, Hammel HT, Hart JS, Lemessurier DH, Steen J. Cold adaptation in australian aborigines. J Appl Physiol 1958; 13:211-8; PMID:13575330 [DOI] [PubMed] [Google Scholar]
- [59].Wyndham CH, Morrison JF. Adjustment to cold of bushmen in the kalahari desert. J Appl Physiol 1958; 13:219-25; PMID:13575331 [DOI] [PubMed] [Google Scholar]
- [60].Foley RA, Lewin R. Principles of Human Evolution. ISBN 978-0-632-04704-8 ed., Blackwell, 2003. [Google Scholar]
- [61].Ruff CB. Morphological adaptation to climate in modern and fossil hominids. Am J Phys Anthropol 1994; 95:65-107; http://dx.doi.org/ 10.1002/ajpa.1330370605 [DOI] [Google Scholar]
- [62].Katzmarzyk PT, Leonard WR. Climatic influences on human body size and proportions: ecological adaptations and secular trends, Am J Phys Anthropol 1998; 106:483-503; PMID:9712477; http://dx.doi.org/ [DOI] [PubMed] [Google Scholar]
- [63].Roberts DF. Body weight, race and climate. Am J Phys Anthropol 1953; 11:533-58; PMID:13124471; http://dx.doi.org/ 10.1002/ajpa.1330110404 [DOI] [PubMed] [Google Scholar]
- [64].Steegmann AT Jr, Cerny FJ, Holliday TW. Neandertal cold adaptation: physiological and energetic factors. Am J Hum Biol 2002; 14:566-83; PMID:12203812; http://dx.doi.org/ 10.1002/ajhb.10070 [DOI] [PubMed] [Google Scholar]
- [65].Wales N. Modeling neanderthal clothing using ethnographic analogues. J Hum Evol 2012; 63:781-95; PMID:23084621; http://dx.doi.org/ 10.1016/j.jhevol.2012.08.006 [DOI] [PubMed] [Google Scholar]
- [66].Finlayson C, Giles Pacheco F, Rodríguez-Vidal J, Fa DA, María Gutierrez López J, Santiago Pérez A, Finlayson G, Allue E, Baena Preysler J, Cáceres I, et al.. Late survival of neanderthals at the southernmost extreme of Europe. Nature 2006; 443:850-3; PMID:16971951; http://dx.doi.org/ 10.1038/nature05195 [DOI] [PubMed] [Google Scholar]
- [67].Plagnol V, Wall JD.. Possible ancestral structure in human populations. PLoS Genet 2006; 2:0972-9; http://dx.doi.org/ 10.1371/journal.pgen.0020105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [68].Iampietro PF, Goldman RF, Buskirk ER, Bass DE. Response of Negro and white males to cold. J Appl Physiol 1959; 14:798-800; PMID:14405789 [DOI] [PubMed] [Google Scholar]
- [69].Jackson RL, Roberts DE, Cote R, McNeal P, Sharp MW, Fay JT, Kraus E, Rahman SA, Hamlet MP. Psychological and Physiological Responses of Blacks and Caucasians to Hand Cooling, Report T20-89. Natick, MA: US Army Research Institute of Environmental Medicine, 1989. [Google Scholar]
- [70].Meehan JP. Individual and racial variations in a vascular response to a cold stimulus, Military Med 1955; 116:330-4; PMID:14369742 [PubMed] [Google Scholar]
- [71].Purkayastha SS, Majumdar D, Selvamurthy W.. Cold acclimatization of tropical men during short- & long-term sojourn to polar environment. Def Sci J 1997; 47:149-58; http://dx.doi.org/ 10.14429/dsj.47.3987 [DOI] [Google Scholar]
- [72].Purkayastha SS, Selvamurthy W, Ilavazhagan G. Peripheral vascular response to local cold stress of tropical men during sojourn in the arctic cold region. Jpn J Physiol 1992; 42:877-89; PMID:1297856 [DOI] [PubMed] [Google Scholar]
- [73].Taylor NAS, Cotter JD. Heat adaptation: guidelines for the optimisation of human performance. Int Sport Med J 2006; 7:33-57; [Google Scholar]
- [74].Adams JM, Fox RH, Grimby G, Kidd DJ, Wolff HS. Acclimatization to heat and its rate of decay in Man. J Physiol (London) 1960; 152:26-7 [Google Scholar]
- [75].Pandolf KB, Time course of heat acclimation and its decay, Int J Sports Med 1998; 19:S157-60; PMID:9694426; http://dx.doi.org/ 10.1055/s-2007-971985 [DOI] [PubMed] [Google Scholar]
- [76].Sawka MN, Wenger CB, Pandolf KB. Thermoregulatory responses to acute exercise - heat stress and heat acclimation, In: Handbook of Physiology. Environmental Physiology. Bethesda, MD: Am.Physiol.Soc. Sect. 4, Vol. I, Chapter 9; 1996; 157-86. [Google Scholar]
- [77].Daanen HAM, Jonkman AG, Layden JD, Linnane DM, Weller AS. Optimising the acquisition and retention of heat acclimation. Int J Sports Med 2011; 32:822-8; PMID:22052034; http://dx.doi.org/ 10.1055/s-0031-1279767 [DOI] [PubMed] [Google Scholar]
- [78].Weller AS, Linnane DM, Jonkman AG, Daanen HAM. Quantification of the decay and re-induction of heat acclimation in dry-heat following 12 and 26 days without exposure to heat stress. Eur J Appl Physiol 2007; 102:57-66; PMID:17891541; http://dx.doi.org/ 10.1007/s00421-007-0563-z [DOI] [PubMed] [Google Scholar]
- [79].Brazaitis M, Eimantas N, Daniuseviciute L, Baranauskiene N, Skrodeniene E, Skurvydas A. Time course of physiological and psychological responses in humans during a 20-day severe-cold-acclimation programme. PLoS One 2014; 9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [80].Cheung SS, Daanen HA. Dynamic adaptation of the peripheral circulation to cold exposure. Microcirculation 2012; 19:65-77; PMID:21851473; http://dx.doi.org/ 10.1111/j.1549-8719.2011.00126.x [DOI] [PubMed] [Google Scholar]
- [81].Tipton MJ, Wakabayashi H, Barwood MJ, Eglin CM, Mekjavic IB, Taylor NAS. Habituation of the metabolic and ventilatory responses to cold-water immersion in humans. J Therm Biol 2013; 38:24-31; PMID:24229801; http://dx.doi.org/ 10.1016/j.jtherbio.2012.10.002 [DOI] [PubMed] [Google Scholar]
- [82].Wakabayashi H, Wijayanto T, Kuroki H, Lee J-, Tochihara Y. The effect of repeated mild cold water immersions on the adaptation of the vasomotor responses. Int J Biometeorol 2012; 56:631-7; PMID:21695574; http://dx.doi.org/ 10.1007/s00484-011-0462-1 [DOI] [PubMed] [Google Scholar]
- [83].O'Brien C, Young AJ, Lee DT, Shitzer A, Sawka MN, Pandolf KB. Role of core temperature as a stimulus for cold acclimation during repeated immersion in 20°C water. J Appl Physiol 2000; 89:242-50; PMID:10904058 [DOI] [PubMed] [Google Scholar]
- [84].Young AJ, Muza SR, Sawka MN, Gonzalez RR, Pandolf KB. Human thermoregulatory responses to cold air are altered by repeated cold water immersion. J Appl Physiol 1986; 60:1542-8; PMID:3710973; http://dx.doi.org/ 10.1063/1.337289 [DOI] [PubMed] [Google Scholar]
- [85].Budd GM, Brotherhood JR, Beasley FA, Hendrie AL, Jeffery SE, Lincoln GJ, Solaga AT. Effects of acclimatization to cold baths on men's responses to whole-body cooling in air. Eur J Appl Physiol Occup Physiol 1993; 67:438-49; PMID:8299616; http://dx.doi.org/ 10.1007/BF00376461 [DOI] [PubMed] [Google Scholar]
- [86].Janský L, Janáková H, Ulicný B, Šrámek P, Hošek V, Heller J, Parízková J. Changes in thermal homeostasis in humans due to repeated cold water immersions. Pflugers Archiv Euro J Physiol 1996; 432:368-72; PMID:8765994; http://dx.doi.org/ 10.1007/s004240050146 [DOI] [PubMed] [Google Scholar]
- [87].Golden FSC, Tipton MJ. Human adaptation to repeated cold immersions. J Physiol (Lond) 1988; 396:349-63; PMID:3411500; http://dx.doi.org/ 10.1113/jphysiol.1988.sp016965 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [88].Stocks JM, Patterson MJ, Hyde DE, Mittleman KD, Taylor NAS. Metabolic habituation following repeated resting cold-water immersion is not apparent during low-intensity cold-water exercise. J Physiol Anthropol Appl Human Sci 2001; 20:263-7; PMID:11759264; http://dx.doi.org/ 10.2114/jpa.20.263 [DOI] [PubMed] [Google Scholar]
- [89].Clapham JC. Central control of thermogenesis. Neuropharmacology 2012; 63:111-23; PMID:22063719; http://dx.doi.org/ 10.1016/j.neuropharm.2011.10.014 [DOI] [PubMed] [Google Scholar]
- [90].Shephard RJ. Metabolic adaptations to exercise in the cold. Sports Med 1993; 16:266-89; PMID:8248684; http://dx.doi.org/ 10.2165/00007256-199316040-00005 [DOI] [PubMed] [Google Scholar]
- [91].Imamura R, Funatsu M, Kawachi H, Tokura H. Effects of wearing long- and mini-skirts for a year on subcutaneous fat thickness and body circumference In: Werner J, Hexamer M (Red).Aachen: Environmental Ergonomics IX.Shaker Verlag, 2000; 315-8. [Google Scholar]
- [92].Sonna LA, Fujita J, Gaffin SL, Lilly CM. Invited review: effects of heat and cold stress on mammalian gene expression. J Appl Physiol 2002; 92:1725-42; PMID:11896043; http://dx.doi.org/ 10.1152/japplphysiol.01143.2001 [DOI] [PubMed] [Google Scholar]
- [93].Fujita J. Cold shock response in mammalian cells. J Mol Microbiol Biotechnol 1999; 1:243-55; PMID:10943555 [PubMed] [Google Scholar]
- [94].Matz JM, Lavoi KP, Moen RJ, Blake MJ. Cold-induced heat shock protein expression in rat aorta and brown adipose tissue. Physiol Behav 1996; 60:1369-74; PMID:8916196; http://dx.doi.org/ 10.1016/S0031-9384(96)00220-X [DOI] [PubMed] [Google Scholar]
- [95].Eglin DM, Tipton MJ. Repeated cold showers as a method of habituating humans to the initial responses to cold water immersion. Euro J Appl Physiol 2005; 93:624-9; PMID:15778892; http://dx.doi.org/ 10.1007/s00421-004-1239-6 [DOI] [PubMed] [Google Scholar]
- [96].Davis TR, Johnston DR, Bell FC, Cremer BJ. Regulation of shivering and non-shivering heat production during acclimation of rats. Am J Physiol 1960; 198:471-5; PMID:13814518 [DOI] [PubMed] [Google Scholar]
- [97].Davis TR. Chamber cold acclimatization in Man. J Appl Physiol 1961; 16:1011-5; PMID:13883973 [DOI] [PubMed] [Google Scholar]
- [98].Van AMJ, Ooijen Van Marken Lichtenbelt WD, Van Steenhoven AA, Westerterp KR. Seasonal changes in metabolic and temperature responses to cold air in humans. Physiol Behav 2004; 82:545-53; PMID:15276821; http://dx.doi.org/ 10.1016/j.physbeh.2004.05.001 [DOI] [PubMed] [Google Scholar]
- [99].Vosselman MJ, Vijgen GHEJ, Kingma BRM, Brans B, Van Marken Lichtenbelt WD. Frequent extreme cold exposure and brown fat and cold-induced thermogenesis: a study in a monozygotic twin. PLoS One 2014; 9:PMID:25014028; http://dx.doi.org/ 10.1371/journal.pone.0101653 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [100].Hanssen MJW, Hoeks J, Brans B, Van Der Lans AAJJ, Schaart G, Van Den Driessche JJ, Jörgensen JA, Boekschoten MV, Hesselink MKC, Havekes B, et al.. Short-term cold acclimation improves insulin sensitivity in patients with type 2 diabetes mellitus. Nat Med 2015; 21:863-5; PMID:26147760; http://dx.doi.org/ 10.1038/nm.3891 [DOI] [PubMed] [Google Scholar]
- [101].Yoneshiro T, Aita S, Matsushita M, Kayahara T, Kameya T, Kawai Y, Iwanaga T, Saito M. Recruited brown adipose tissue as an antiobesity agent in humans. J Clin Invest 2013; 123:3404-8; PMID:23867622; http://dx.doi.org/ 10.1172/JCI67803 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [102].Blondin DP, Labbé SM, Tingelstad HC, Noll C, Kunach M, Phoenix S, Guérin B, Turcotte ÉE, Carpentier AC, Richard D, et al.. Increased brown adipose tissue oxidative capacity in cold-acclimated humans. J Clin Endocrinol Metab 2014; 99:E438-46; PMID:24423363 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [103].Lee P, Smith S, Linderman J, Courville AB, Brychta RJ, Dieckmann W, Werner CD, Chen KY, Celi FS. Temperature-acclimated brown adipose tissue modulates insulin sensitivity in humans. Diabetes 2014; 63:3686-98; PMID:24954193; http://dx.doi.org/ 10.2337/db14-0513 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [104].Van Ooijen AMJ, Van Marken Lichtenbelt WD, Westerterp KR. Individual differences in body temperature and the relation to energy expenditure: the influence of mild cold. J Therm Biol 2001; 26:455-9; http://dx.doi.org/ 10.1016/S0306-4565(01)00060-2 [DOI] [Google Scholar]
- [105].Scholander PF, Hammel HT, Andersen KL, Loyning Y. Metabolic acclimation to cold in Man. J Appl Physiol 1958; 12:1-8; PMID:13502248 [DOI] [PubMed] [Google Scholar]
- [106].Lorenzo S, Halliwill JR, Sawka MN, Minson CT. Heat acclimation improves exercise performance. J Appl Physiol 2010; 109:1140-7; PMID:20724560; http://dx.doi.org/ 10.1152/japplphysiol.00495.2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [107].Pilcher JJ, Nadler E, Busch C. Effects of hot and cold temperature exposure on performance: a meta-analytic review. Ergonomics 2002; 45:682-98; PMID:12437852; http://dx.doi.org/ 10.1080/00140130210158419 [DOI] [PubMed] [Google Scholar]
- [108].Enander A. Effects of moderate cold on performance of psychomotor and cognitive tasks. Ergonomics 1987; 30:1431-45; PMID:3428250; http://dx.doi.org/ 10.1080/00140138708966037 [DOI] [PubMed] [Google Scholar]
- [109].Daanen HAM, Koedam J, Cheung SS. Trainability of cold induced vasodilatation in fingers and toes. Eur J Appl Physiol 2012; 112:2595-601; PMID:22081047; http://dx.doi.org/ 10.1007/s00421-011-2233-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [110].Black AE, Coward WA, Cole TJ, Prentice AM. Human energy expenditure in affluent societies: an analysis of 574 doubly-labelled water measurements. Eur J Clin Nutr 1996; 50:72-92; PMID:8641250 [PubMed] [Google Scholar]
- [111].Van Ooijen AMJ, Van Marken Lichtenbelt WD, Van Steenhoven AA, Westerterp KR. Cold-induced heat production preceding shivering. Br J Nutr 2005; 93:387-391; PMID:15877879; http://dx.doi.org/ 10.1079/BJN20041362 [DOI] [PubMed] [Google Scholar]
- [112].Forslind B, Engström S, Engblom J, Norlén L. A novel approach to the understanding of human skin barrier function. J Dermatol Sci 1997; 14:115-125; PMID:9039975; http://dx.doi.org/ 10.1016/S0923-1811(96)00559-2 [DOI] [PubMed] [Google Scholar]