

Abstract
Site-directed mutagenesis of CphA indicated that prolines in the P158-P172 loop are essential for the stability and the catalytic activity of subclass B2 metallo-β-lactamases against carbapenems. The sequential substitution of proline led to a decrease of the catalytic efficiency of the variant compared to the wild-type (WT) enzyme but also to a higher affinity for the binding of the second zinc ion.
TEXT
The subclass B2 metallo-β-lactamase (MBL) CphA is a narrow-spectrum metallo-β-lactamase, which hydrolyzes only carbapenems very efficiently and shows poor activity versus penicillins and cephalosporins (1). CphA has two potential metal binding sites, as observed in the B1 and B3 subclass β-lactamases (2, 3). At low zinc ion concentration (≤0.4 μM), CphA binds only a single zinc ion bound in the so-called Zn2 site (D120-C221-H264), whereas the Zn1 site (N116-H118-H149) is occupied by water molecules. Despite the low sequence identity of the three subclass enzymes, the αββα fold is identical in the three subclasses and the active site scaffold is relatively conserved. In contrast to B1 and B3 subclass structures, CphA possesses a long α3 helix (R140-L161) located near the active-site groove, which may impair a strong interaction with the majority of the β-lactam molecules. This α3 helix is followed immediately by an unusual proline-rich loop (Pro158, Pro161, Pro164, and Pro172). The proline motif reduces the flexibility of the loop and may affect the position of the α3 helix and, therefore, modify the accessibility of the catalytic active site. In order to understand the impact of the proline-rich loop on the narrow spectrum of activity of CphA, six mutants with progressive point mutations were sequentially generated, namely, CphAP158A, CphAP161A, CphAP172A, CphAP158A-P161A, CphAP158A-P161A-P164A, and CphAP158A-P161A-P164A-P172A. We choose to replace the proline residues with alanine to preserve the neutral and apolar features of the side chain. The sequence of the CphA enzyme (GenBank accession number X57102) was used as the template to generate the mutants. The synthetic mutant genes and blaCphA with codons optimized for expression in Escherichia coli were generated by GeneArt (Germany).
The genes were cloned in the pBC-SK(+) and pET24(a) vectors. The presence of the stop codon in the different genes prevented the production of the protein with a poly-His sequence. The positive amplicons were sequenced by using the BigDye sequencing reaction kit and an ABI Prism 310 capillary automated sequencer (Life Technologies, Monza, Italy).
The phenotypic profile was carried out on E. coli HB101(pBC-SK), E. coli HB101(pBC-SK/CphA), E. coli HB101(pBC-SK/CphAP158A), E. coli HB101(pBC-SK/CphAP158A-P161A), E. coli HB101(pBC-SK/CphAP158A-P161A-P164A), and E. coli HB101(pBC-SK/CphAP158A-P161A-P164A-P172A) by microdilution method using a bacterial inoculum of 5 × 105 CFU/ml according to Clinical and Laboratory Standards Institute (CLSI) performance standards (4). The antimicrobial agents used in this study were piperacillin, amoxicillin, ceftriaxone, cefotaxime, ceftazidime, cefepime, cefazolin, imipenem, meropenem, ertapenem, biapenem, and aztreonam.
E. coli BL21(DE3)pET24aCphA wild type (WT), BL21(DE3)(pET24aCphAP158A), BL21(DE3)(pET24aCphAP161A), BL21(DE3)(pET24aCphAP172A), BL21(DE3)(pET24aCphAP158A-P161A), BL21(DE3)(pET24aCphAP158A-P161A-P164A), and BL21(DE3)(pET24aCphAP158A-P161A-P164A-P172A) recombinant strains, used to produce the CphA enzymes, were grown in 500 ml of 2xYT medium with 50 μg/ml kanamycin at 37°C in an orbital shaker. The expression of the different enzymes was induced by the addition of isopropyl-β-thiogalactoside (IPTG) at a final concentration of 1 mM when the cultures reached an A600 of 0.7. The cultures were incubated at 18°C for 6 additional hours. Bacteria were harvested by centrifugation (Avanti JE centrifuge; Beckman Coulter Inc., USA) at 10,000 × g for 10 min at 4°C. The pellet was washed twice and was finally resuspended in 25 mM morpholineethanesulfonic acid (MES) buffer, pH 6.5. Cell crude extract was obtained with the help of a cell disrupter (Emulsiflex C3; Avestin GmbH, Germany), which allows cell lysis at a pressure of 5,500 to 6,500 kPa. The lysate was centrifuged at 105,000 × g for 30 min. The cleared supernatant was filtered in a 45-μm filter and was then loaded onto a Sepharose SP-HP 5-ml column equilibrated in 25 mM MES buffer, pH 6.5 (buffer A). The β-lactamase was eluted with a gradient of 25 mM MES plus 0.5 M NaCl, pH 6.5 (buffer B). The fractions containing β-lactamase activity were eluted at 45% to 55% buffer B, pooled, and then dialyzed overnight in 25 mM HEPES, pH 7, with the addition of protease inhibitor cocktail tablets (EDTA free) (Roche Diagnostics GmBH, Germany). The active fractions were loaded onto a column containing pentadentate chelator (PDC) for binding Zn2+, equilibrated with 25 mM HEPES, pH 7.0 (buffer A), for the second chromatographic step. The pure fractions were pooled and dialyzed in 20 mM HEPES buffer, pH 7.0, to remove the excess NaCl. At the end of each purification step, β-lactamase activity was routinely measured spectrophotometrically following the hydrolysis of 100 μM imipenem as the substrate. Kinetic experiments were performed following the hydrolysis of each substrate at 30°C in 25 mM HEPES buffer (pH 7.0). The data were collected with a Specord 50 spectrophotometer (Analytik Jena, Germany). Each kinetic value is the mean of five different measurements; error was below 5%. Kinetic parameters were determined under initial rate conditions. The dependence of zinc content on activity was determined by a hydrolysis rate of 150 μM imipenem as the reporter substrate using a concentration of ZnCl2 ranging from 0 to 100 μM.
The results of the antimicrobial susceptibility test are shown in Table 1. The presence of the CphA enzyme confers resistance to E. coli HB101 versus all carbapenems. However, low MIC values were observed for cephalosporins, penicillins, and aztreonam. The same pattern was reported for E. coli HB101(pBC-SK/CphAP158A). A slight difference was observed for the second mutant [E. coli HB101(pBC-SK/CphAP158A-P161A)]. Indeed, the MIC values versus carbapenems decrease, but E. coli HB101(pBC-SK/CphAP158A-P161A) remains resistant. The triple mutant showed a drastic reduction of MIC values toward carbapenems. The MIC values observed for the fourth mutant [E. coli HB101(pBC-SK/CphAP158A-P161A-P164A-P172A)] are very similar to those of E. coli HB101(pBC-SK), which was used as the control strain. The β-lactamase activity of the crude extracts indicated that, with the exception of CphAP158A-P161A-P164A-P172A, the reduction of the MIC values is correlated to the reduced catalytic efficiency and to the reduced production of active enzymes (Table 1). Therefore, our data indicated that the addition of the point mutation on the P158-P172 loop restored the susceptibility of the host bacteria to carbapenems.
TABLE 1.
Antimicrobial susceptibility mediated by CphA, CphAP158A, CphAP158A-P161A, CphAP158A-P161A-P164A, and CphAP158A-P161A-P164A-P172A in E. coli HB101a
| Antibiotic | MIC value (μg/ml) according to CphA type |
|||||
|---|---|---|---|---|---|---|
| WT | P158A | P158A-P161A | P158A-P161A-P164A | P158A-P161A-P164A-P172A | pBC-SK | |
| Amoxicillin | 8 | 8 | 8 | 8 | 8 | 4 |
| Piperacillin | 2 | 2 | 2 | 1 | 0.5 | 0.5 |
| Cefepime | 2 | 2 | 2 | 2 | 0.5 | 0.25 |
| Ceftriaxone | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.25 |
| Cefazolin | 1 | 1 | 1 | 1 | 1 | 0.25 |
| Cefotaxime | 1 | 1 | 1 | 1 | 0.5 | 0.25 |
| Ceftazidime | 2 | 2 | 1 | 1 | 0.5 | 0.5 |
| Imipenem | >64 | >64 | 32 | 2 | 0.25 | 0.25 |
| Meropenem | >64 | >64 | 32 | 4 | ≤0.0312 | ≤0.0312 |
| Ertapenem | >64 | >64 | 32 | 2 | ≤0.0312 | ≤0.0312 |
| Biapenem | >64 | >64 | 4 | 2 | 0.25 | 0.25 |
| Aztreonam | 4 | 4 | 4 | 2 | 2 | 2 |
| Activity | 12 | 14 | 5 | 0.6 | 0.012 | <0.001 |
MIC experiments were performed on E. coli HB101(pBC-SK), E. coli HB101(pBC-SK/CphA), E. coli HB101(pBC-SK/CphAP158A), E. coli HB101(pBC-SK/CphAP158A-P161A), E. coli HB101(pBC-SK/CphAP158A-P161A-P164A), and E. coli HB101(pBC-SK/CphAP158A-P161A-P164A-P172A). The inoculum used for MIC testing was 5 × 105 CFU/ml. The activity of the crude extracts, expressed in micromoles per minute per milligram, was measured using 100 μM imipenem.
The production assays of the different proteins indicated that CphAP158A-P161A-P164A-P172A was insoluble and that all of the methods applied to solubilize it failed. Kinetics studies (Table 2) showed that the kcat values for the hydrolysis of imipenem and ertapenem are similar to those of the wild-type CphA, but the Km values increase as a function of the number of mutations introduced. We noted a reduction of 4- to 7-fold for the catalytic efficiency of mutant CphAP158A-P161A-P164A compared to that of the WT. Interestingly, in the cases of meropenem and biapenem, we noted a progressive decrease of Km, kcat, and kcat/Km in the five mutants. We did not detect any significant hydrolysis of penicillins, indicating that the narrow activity spectrum of CphA is not modified by the replacement of prolines.
TABLE 2.
Kinetics study on CphA WT and mutants CphAP158A, CphAP158A-P161A, CphAP158A-P161A-P164A
| CphA | Imipenem |
Meropenem |
Ertapenem |
Biapenem |
Kd (μM) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Km (μM) | kcat (s−1) | kcat/Km (μM−1 s−1) | Km (μM) | kcat (s−1) | kcat/Km (μM−1 s−1) | Km (μM) | kcat (s−1) | kcat/Km (μM−1 s−1) | Km (μM) | kcat (s−1) | kcat/Km (μM−1 s−1) | ||
| WT | 110 | 460 | 4.18 | 1,600 | 3,100 | 1.94 | 230 | 700 | 3.04 | 300 | 150 | 0.5 | 50 |
| P158A | 210 | 500 | 2.38 | 720 | 1,700 | 2.36 | 260 | 770 | 2.96 | 400 | 160 | 0.4 | 45 |
| P161A | 130 | 476 | 3.66 | 312 | 970 | 3.11 | 400 | 1,350 | 3.37 | 210 | 80 | 0.72 | 15 |
| P172A | 320 | 200 | 0.62 | 190 | 210 | 1.1 | 220 | 280 | 1.27 | 200 | 24 | 0.12 | 10 |
| P158A-P161A | 430 | 570 | 1.33 | 510 | 700 | 1.36 | 260 | 550 | 2.12 | 160 | 8 | 0.05 | 20 |
| P158A-P161A-P164A | 700 | 400 | 0.57 | 200 | 160 | 0.8 | 530 | 410 | 0.77 | 190 | 4.4 | 0.023 | 5 |
We also tested the variation of the β-lactamase activity toward zinc concentration. As observed for the WT enzyme (5), we noted a decrease of the activity in the presence of zinc ions (Fig. 1). From these data, we estimated an apparent dissociation constant for the different enzymes. We noted that, compared to the WT enzyme, P161A, P172A, and the triple mutant displayed a higher apparent affinity for the second zinc ion (Kd = 5 to 15 μM). Finally, the thermostability of the triple mutant at 45°C was affected compared to those of the three other proteins. The mutant was inhibited completely after 15 min of incubation.
FIG 1.
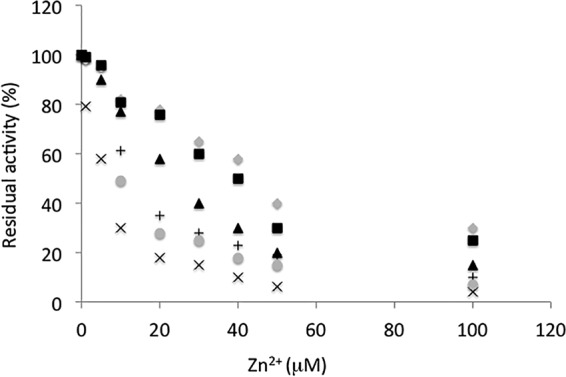
Influence of the zinc concentrations on the CphA activity. Gray diamond, WT; black square, P158A; plus sign, P161A; gray circle, P172A; black triangle, P158A161A; times sign, P158A161A164A.
In conclusion, our studies indicated that the progressive replacement of prolines from the P158-P172 loop showed that the prolines are crucial for the activity and the stability of CphA. The replacement of single prolines at P158 and P161 led to a strong modification of CphA activities toward carbapenems. A major impact on catalytic efficiencies of the enzyme was observed when the P158A P161A double mutant was generated. The most affected single mutant was P172A, whose catalytic efficiency toward imipenem was decreased by a factor of 4. The replacement of the four prolines led to a highly unstable protein that could not be produced in E. coli. The comparison of the two three-dimensional structures of known subclass B2 (CphA and Sfh-I) (6–8) indicated that the prolines in positions 161 and 172 are conserved (Fig. 2A and B). The search for new subclass B2 MBLs was done using the CphA sequence as the template. We identified five new enzymes. They share at least 56.5% identity compared to CphA. All of the structural features that characterized a subclass B2 MBL are highly conserved in all of the proteins and suggest that they will act as strict carbapenemases. We also noted that P161 and P172 are strictly conserved, while P158 and P164 are replaced by hydrophilic amino acids, which will reinforce their interaction with water. Our experiments showed that P158 does not play an important function in CphA. The replacement of P161 or P172 yielded an enzyme whose apparent affinity for the second zinc ion increases. This effect was amplified when two or three proline residues were replaced. Therefore, we can conclude that the proline motif plays a structural role in the subclass B2 MBLs. It allows a more rigid and stable structure. It also influences the architecture of the active site and affects the affinity of the second metal binding site.
FIG 2.

(A) ClustalW alignment of subclass B2 metallo-β-lactamases using Clustal 2.1 multiple-sequence alignment. C.violaceum, Chromobacterium violaceum ATCC 12472; P.ferrooxidans, Pseudogulbenkiania ferrooxidans; C.piscinae, Chromobacterium piscinae; A.hydrophila CphA, Aeromonas hydrophila CphA; P.chlororaphis, Pseudomonas chlororaphis; S.Fonticola Sfh-I, Serratia fonticola Sfh-I; Y.mollaretii, Yersinia mollaretii ATCC 43969. (B) Comparison of the structures of Sfh-I and CphA. The lateral chains of the amino acids of the P158-P172 loop are shown.
ACKNOWLEDGMENT
This work was supported by a research grant for foreign doctoral students (2013 to 2014) from the University of Liège, Belgium.
REFERENCES
- 1.Segatore B, Massidda O, Satta G, Setacci D, Amicosante G. 1993. High specificity of CphA-encoded metallo-β-lactamase from Aeromonas hydrophila AE036 for carbapenems and its contribution to β-lactamase resistance. Antimicrob Agents Chemother 37:1324–1328. doi: 10.1128/AAC.37.6.1324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Carfi A, Pares S, Duée E, Galleni M, Duez C, Frère JM, Dideberg O. 1995. The 3-D structure of a zinc metallo-β-lactamase from Bacillus cereus reveals a new type of protein fold. EMBO J 14:4914–4921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Garau G, Garcia-Sàez I, Bebrone C, Anne C, Mercuri P, Galleni M, Frère JM, Dideberg O. 2004. Update of the standard numbering scheme for class B β-lactamases. Antimicrob Agents Chemother 48:2347–2349. doi: 10.1128/AAC.48.7.2347-2349.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Clinical and Laboratory Standards Institute. 2012. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; approved standard—9th ed. CLSI document M09-A7 Clinical and Laboratory Standards Institute, Wayne, PA. [Google Scholar]
- 5.Hernandez Valladares M, Felici A, Weber G, Adolph HW, Zeppezauer M, Rossolini GM, Amicosante G, Frère JM, Galleni M. 1997. Zn(II) dependence of the Aeromonas hydrophila AE036 metallo-β-lactamase activity and stability. Biochemistry 36:11534–11541. doi: 10.1021/bi971056h. [DOI] [PubMed] [Google Scholar]
- 6.Fonseca F, Arthur CJ, Bromley EH, Samyn B, Moerman P, Saavedra MJ, Correia A, Spencer J. 2011. Biochemical characterization of Sfh-I, a subclass B2 metallo-beta-lactamase from Serratia fonticola UTAD54. Antimicrob Agents Chemother 55:5392–5395. doi: 10.1128/AAC.00429-11.21876065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Fonseca F, Bromley EH, Saavedra MJ, Correia A, Spencer J. 2011. Crystal structure of Serratia fonticola Sfh-I: activation of the nucleophile in mono-zinc metallo-β-lactamases. J Mol Biol 411:951–959. doi: 10.1016/j.jmb.2011.06.043. [DOI] [PubMed] [Google Scholar]
- 8.Bebrone C, Delbrück H, Kupper MB, Schlömer P, Willmann C, Frère JM, Fischer R, Galleni M, Hoffmann KM. 2009. The structure of the di-zinc subclass B2 metallo-β-lactamase CphA reveals that the second inhibitory zinc ion binds in the histidine site. Antimicrob Agents Chemother 53:4464–4471. doi: 10.1128/AAC.00288-09. [DOI] [PMC free article] [PubMed] [Google Scholar]