

Propionibacterium acnes is a dominant bacterium residing on skin, and it has been thought to play a causal role in several diseases including acne, a common skin disease affecting more than 80% of people worldwide. While specific strains of P. acnes have been associated with either disease or healthy skin, the mechanisms remain unclear. Recently, we showed that vitamin B12 supplementation increased porphyrin production in P. acnes, leading to acne development (D. Kang, B. Shi, M. C. Erfe, N. Craft, and H. Li, Sci. Transl. Med. 7:293ra103, 2015, doi:10.1126/scitranslmed.aab2009). Here, we reveal that the levels of porphyrin production and vitamin B12 regulation are different between acne- and health-associated strains, suggesting a potential molecular mechanism for disease-associated strains in acne pathogenesis and for health-associated strains in skin health. This study highlights the importance of understanding the strain-level differences of the human microbiota in disease pathogenesis. Our findings also suggest the porphyrin biosynthesis pathway as a candidate drug target and use of health-associated strains as potential probiotics in novel acne therapeutics.
KEYWORDS: porphyrin, vitamin B12, Propionibacterium acnes, strain, acne, 5-aminolevulinic acid, levulinic acid
ABSTRACT
Propionibacterium acnes is an important skin commensal, but it is also considered a pathogenic factor in several diseases including acne vulgaris, the most common skin disease. While previous studies have revealed P. acnes strain-level differences in health and disease associations, the underlying molecular mechanisms remain unknown. Recently, we demonstrated that vitamin B12 supplementation increases P. acnes production of porphyrins, a group of proinflammatory metabolites important in acne development (D. Kang, B. Shi, M. C. Erfe, N. Craft, and H. Li, Sci. Transl. Med. 7:293ra103, 2015, doi:10.1126/scitranslmed.aab2009). In this study, we compared the porphyrin production and regulation of multiple P. acnes strains. We revealed that acne-associated type IA-2 strains inherently produced significantly higher levels of porphyrins, which were further enhanced by vitamin B12 supplementation. On the other hand, health-associated type II strains produced low levels of porphyrins and did not respond to vitamin B12. Using a small-molecule substrate and inhibitor, we demonstrated that porphyrin biosynthesis was modulated at the metabolic level. We identified a repressor gene (deoR) of porphyrin biosynthesis that was carried in all health-associated type II strains, but not in acne-associated type IA-2 strains. The expression of deoR suggests additional regulation of porphyrin production at the transcriptional level in health-associated strains. Our findings provide one potential molecular mechanism for the different contributions of P. acnes strains to skin health and disease and support the role of vitamin B12 in acne pathogenesis. Our study emphasizes the importance of understanding the role of the commensal microbial community in health and disease at the strain level and suggests potential utility of health-associated P. acnes strains in acne treatment.
IMPORTANCE Propionibacterium acnes is a dominant bacterium residing on skin, and it has been thought to play a causal role in several diseases including acne, a common skin disease affecting more than 80% of people worldwide. While specific strains of P. acnes have been associated with either disease or healthy skin, the mechanisms remain unclear. Recently, we showed that vitamin B12 supplementation increased porphyrin production in P. acnes, leading to acne development (D. Kang, B. Shi, M. C. Erfe, N. Craft, and H. Li, Sci. Transl. Med. 7:293ra103, 2015, doi:10.1126/scitranslmed.aab2009). Here, we reveal that the levels of porphyrin production and vitamin B12 regulation are different between acne- and health-associated strains, suggesting a potential molecular mechanism for disease-associated strains in acne pathogenesis and for health-associated strains in skin health. This study highlights the importance of understanding the strain-level differences of the human microbiota in disease pathogenesis. Our findings also suggest the porphyrin biosynthesis pathway as a candidate drug target and use of health-associated strains as potential probiotics in novel acne therapeutics.
INTRODUCTION
Propionibacterium acnes is a major commensal bacterium residing on human skin. It plays important roles in maintaining skin health, but it has also been implicated in the pathogenesis of several diseases and infections, including sarcoidosis, SAPHO (synovitis, acne, pustulosis, hyperostosis, and osteitis) syndrome, endodontic lesions, eye infections, prosthetic joint infections, prostate cancer, and acne vulgaris (commonly called acne) (1–4). Acne is the most common skin disease, affecting more than 80% of adolescents and young adults worldwide (5, 6). Despite the clinical importance of the disease (7, 8), the etiology of acne is not yet clear. P. acnes dominates the skin of both acne patients and healthy individuals, and thus, its role in acne pathogenesis has not been well understood. Previously, culture-based and 16S rRNA metagenomic studies identified P. acnes strains that were associated with either acne or healthy skin (9–12). Genome comparison of a large number of strains revealed key genetic differences among P. acnes lineages (11, 13, 14). Type IA-2 (primarily ribotype 4 [RT4] and RT5) strains have been associated with acne. They harbor extra genomic elements encoding multiple virulence genes. On the other hand, type II strains, in particular RT6 and some RT2 strains, have rarely been found in acneic skin and thus are defined as health-associated strains in the context of acne (10–12, 15). Type II strains carry clustered regularly interspaced short palindromic repeat (CRISPR) elements, which may prevent these strains from acquiring virulence genes from phage or other foreign DNA. The presence of these genetic elements partly explains how different P. acnes strains may play roles in health or disease; however, the molecular mechanisms underlying P. acnes strain-level differences in health and disease associations remain to be elucidated.
To date, limited information exists about P. acnes strain-level differences beyond the genomic variations. Molecules secreted by P. acnes, such as proteases, lipases, hemolysins, and porphyrins, can degrade host tissue and have been suggested as causal factors in acne (16–20). Porphyrins can generate reactive oxygen species and induce inflammation in keratinocytes (21–23). Previous studies have shown correlations between acne severity and the concentrations of bacterium-derived porphyrins in the hair follicle. Increased levels of porphyrins were observed in acneic skin compared to healthy skin, as well as in acne lesions compared to nonlesional sites of acne patients (24–26). Consistently, a reduction in porphyrin levels was observed in acne patients who positively responded to acne treatment, while those who did not respond to acne treatment exhibited unchanged or increased levels of bacterial porphyrins on their skin (27–29).
Vitamin B12 has been suggested to induce acne (18, 30–33). In propionibacteria, the vitamin B12 and porphyrin biosynthesis pathways are inversely correlated (18, 34). Our recent study suggested that vitamin B12 supplementation repressed its own biosynthesis in P. acnes, resulting in increased porphyrin production, and led to acne development in a subset of individuals (18).
In this study, we compared the porphyrin production and regulation between acne-associated type IA-2 strains and health-associated type II strains to investigate a potential molecular link between porphyrin production and disease association of various P. acnes strains.
RESULTS
Acne-associated P. acnes strains produced significantly more porphyrins than health-associated strains.
To investigate whether different P. acnes strains produce the same porphyrin species, we first characterized the types of porphyrins secreted by multiple P. acnes strains using mass spectrometry. Four acne-associated type IA-2 strains, HL053PA1 (RT4), HL045PA1 (RT4), HL043PA1 (RT5), and HL043PA2 (RT5), and three health-associated type II strains, HL001PA1 (RT2), HL103PA1 (RT2), and HL042PA3 (RT6), were examined (Table 1). We found that there was no difference in the porphyrin species produced by these P. acnes strains (see Fig. S1 in the supplemental material). Consistent with previous studies (35–37), coproporphyrin III was the dominant porphyrin isomer produced by all strains ([M+H]+ =655.3). Minimal amounts of coproporphyrin I were also detected.
TABLE 1 .
P. acnes strains used in this study
| Association or type | Lineagea | Phylogroupb | Strain | Ribotypea | Clonal complex (sequence type by Belfast MLST8)b |
Clonal complex (sequence type by Aarhus MLST9)c |
|---|---|---|---|---|---|---|
| Acne-associated | IA-2 | IA1 | HL053PA1 | RT4 | CC3 (ST3) | CC3 (ST3) |
| HL045PA1 | RT4 | CC3 (ST17) | CC3 (ST3) | |||
| HL043PA1 | RT5 | CC3 (ST3) | CC3 (ST58) | |||
| HL043PA2 | RT5 | CC3 (ST3) | CC3 (ST58) | |||
| Health-associated | II | II | HL001PA1 | RT2 | CC72 (ST30) | CC60 (ST60) |
| HL103PA1 | RT2 | CC6 (ST25) | CC60 (ST60) | |||
| HL042PA3 | RT6 | CC6 (ST7) | CC60 (ST73) | |||
| Type I with deoR | I | IA1 | HL025PA1 | RT1 | CC4 (ST4) | CC28 (ST27) |
Coproporphyrin was the dominant porphyrin isoform produced by P. acnes. Porphyrins secreted by P. acnes have a mass spectrum characteristic of the monoisotopic coproporphyrin isoform ([M + H]+ = 655.3). The doubly charged parent ion was also observed ([M + 2H]2+ = 328.2). Download Figure S1, PDF file, 0.1 MB (53.1KB, pdf) .
Copyright © 2016 Johnson et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
To determine whether P. acnes strains produce different amounts of porphyrins, we next quantified the secreted porphyrins in the seven P. acnes strains. The average porphyrin level produced by acne-associated type IA-2 strains was 6.5 µM (5.6 to 8.8 µM), which was significantly greater than that produced by health-associated type II strains, 1.1 µM (0 to 2.1 µM) (P < 0.0001) (Fig. 1). Notably, the RT2 strain HL001PA1 produced no detected porphyrins in all experiments. This strain belongs to clonal complex 72 (CC72) based on the Belfast MLST8 (multilocus sequence typing using eight housekeeping genes) scheme (Table 1). McDowell et al. reported that strains from CC72 were isolated from healthy skin (10). This is consistent with our finding.
FIG 1 .

Acne-associated type IA-2 P. acnes strains produced significantly more porphyrins than health-associated type II strains. Each bar represents the porphyrins produced by a strain normalized to bacterial culture density. The means plus standard errors (error bars) of data obtained from at least three independent experiments with at least three replicates each are shown.
Vitamin B12 supplementation significantly increased porphyrin production in acne-associated strains, but not in health-associated strains.
To investigate whether vitamin B12 modulates P. acnes porphyrin production in a strain-specific manner, we compared the levels of porphyrins produced by different P. acnes strains with and without vitamin B12 supplementation. We found that vitamin B12 supplementation led to increased porphyrin production in all tested acne-associated type IA-2 strains (average porphyrin level increasing from 6.5 µM to 9.2 µM), with statistical significance in three of the four tested strains (P ≤ 0.02) (Fig. 2). In contrast, vitamin B12 supplementation had no significant effect on porphyrin production in health-associated type II strains (average porphyrin level remaining at 1.1 µM) (P ≥ 0.77). The porphyrin levels in strain HL001PA1 remained undetected. To confirm our results, we tested porphyrin production, with and without the addition of vitamin B12, of three additional type II strains, HL110PA3 (RT6), HL106PA1 (RT2), and HL050PA2 (RT1). We found that these strains produced similarly low levels of porphyrins (1.4 ± 1.2 µM) and that the production level was not significantly affected by vitamin B12 supplementation. Our data suggest that vitamin B12 modulates porphyrin production in acne-associated type IA-2 strains, but not in health-associated type II strains, indicating a molecular link between P. acnes strain composition of the skin microbiota and the observation that vitamin B12 induces acne in a subset of individuals.
FIG 2 .

Vitamin B12 supplementation significantly increased porphyrin production in acne-associated type IA-2 strains, but not in health-associated type II strains. P. acnes strains were cultured in medium with or without the addition of 10 µg/ml vitamin B12. Each bar represents the porphyrins produced by a strain normalized to bacterial culture density. The means plus standard errors (error bars) of data obtained from at least three independent experiments with at least three replicates each are shown.
Vitamin B12 supplementation repressed vitamin B12 biosynthesis gene expression.
To determine whether vitamin B12 affects porphyrin production via repression of its own biosynthesis in P. acnes, we performed quantitative reverse transcription-PCR (qRT-PCR) to measure the expression level of cbiL, a gene in the vitamin B12 biosynthesis pathway. The cbiL gene encodes precorrin-2 C-20-methyltransferase, a key enzyme involved in corrin ring formation in the vitamin B12 biosynthesis pathway. We found that the addition of vitamin B12 to P. acnes cultures resulted in the downregulation of cbiL gene expression, with average fold changes of 0.74 in acne-associated type IA-2 strains and 0.24 in health-associated type II strains (Fig. 3). The downregulation of cbiL gene expression is consistent with the previous finding that vitamin B12 supplementation repressed the vitamin B12 biosynthesis pathway (18), leading to increased porphyrin production. The expression of cbiL was downregulated in both type IA-2 and type II strains, despite our above observation that the porphyrin production in type II strains was unaffected by vitamin B12 supplementation. This suggests that additional mechanisms are involved in inhibiting porphyrin production in health-associated type II strains.
FIG 3 .

Vitamin B12 supplementation repressed the expression of a vitamin B12 biosynthesis gene, cbiL. The expression level of cbiL was quantified by qRT-PCR from P. acnes strains cultured with or without the addition of 10 µg/ml vitamin B12. Strains of types IA-2 and II and an RT1 strain carrying deoR, HL025PA1, are shown. Each bar represents the fold change in gene expression of cbiL in cultures with vitamin B12 supplementation compared to cultures without supplementation. The means plus standard deviations (error bars) of data obtained from at least two independent experiments with at least three replicates each are shown.
Addition of 5-aminolevulinic acid increased porphyrin production, which was further enhanced by vitamin B12.
The porphyrin and vitamin B12 biosynthesis pathways in P. acnes share the same initial enzymatic steps and a common precursor, 5-aminolevulinic acid (5-ALA), the substrate of the rate-limiting enzyme, porphobilinogen synthase. To investigate whether porphyrin production can be promoted by increasing the availability of 5-ALA, we compared the porphyrin production levels of P. acnes strains with and without addition of the substrate. Additionally, we examined whether vitamin B12 has an additive effect on porphyrin production when supplemented in combination with 5-ALA. Upon substrate addition only, porphyrin production was significantly increased by average values of 2.2-fold in acne-associated type IA-2 strains (all P < 0.0001) and 3.4-fold in health-associated type II strains (P ≤ 0.06, except for strain HL001PA1) compared to untreated controls (Fig. 4). Strain HL001PA1 consistently produced no detected porphyrins, even upon addition of 5-ALA. Supplementation of 5-ALA in combination with vitamin B12 further enhanced porphyrin production in acne-associated type IA-2 strains (1.2-fold increase compared to 5-ALA only) with statistical significance in three of the four tested strains (P ≤ 0.04), but not in health-associated type II strains (P ≥ 0.94). This is consistent with our earlier finding that type II strains did not respond to vitamin B12 in their porphyrin production (Fig. 2). Our data suggest that the metabolic influx of substrates influences porphyrin production in P. acnes. The modulation of porphyrin production by vitamin B12 supplementation is specific to acne-associated type IA-2 strains, but not health-associated type II strains.
FIG 4 .

5-ALA increased porphyrin production, which was further enhanced by vitamin B12 supplementation in acne-associated type IA-2 strains. P. acnes strains were cultured in medium with substrate 5-ALA (0.1 mg/ml) (grey bars) or without 5-ALA (white bars) or with both 5-ALA and vitamin B12 (10 µg/ml) added (black bars). 5-ALA significantly increased porphyrin production in acne-associated type IA-2 strains (P < 0.0001) and in health-associated type II strains (P ≤ 0.06, except for HL001PA1). Vitamin B12 supplementation further increased porphyrin production in the presence of 5-ALA in acne-associated type IA-2 strains, but not in health-associated type II strains. Each bar represents the porphyrins produced by each strain normalized to the bacterial culture density. The means plus standard errors (error bars) of data obtained from at least three independent experiments with at least three replicates each are shown.
Small-molecule inhibitor reduced porphyrin production in P. acnes, and its inhibition was counteracted by vitamin B12 supplementation.
To further demonstrate that porphyrin production can be modulated at the metabolic level, we investigated the effect of a small-molecule inhibitor, levulinic acid (LA), on porphyrin biosynthesis in P. acnes strains. Additionally, we examined whether vitamin B12 counteracts the effect of LA on porphyrin production when supplemented in combination with LA. LA is an analog of 5-ALA and has been shown in Pseudomonas aeruginosa to inhibit the enzymatic activity of porphobilinogen synthase (38), blocking the metabolic influx to the porphyrin and vitamin B12 biosynthesis pathways. We examined a range of concentrations of LA from 0.1 mg/ml to 1.0 mg/ml and found that at the concentration of 0.1 mg/ml, LA did not significantly affect bacterial growth, thus making the measurements of porphyrin production experimentally feasible. LA (0.1 mg/ml) significantly reduced porphyrin production in all strains except for strain HL001PA1 with an average of 30% reduction compared to untreated cultures (P < 0.0001) (Fig. 5). However, in acne-associated type IA-2 strains, there was no significant reduction of porphyrins when supplemented in combination with 10 µg/ml vitamin B12. Vitamin B12 supplementation counteracted inhibition of porphyrin biosynthesis by LA specifically in acne-associated type IA-2 strains, but not in health-associated type II strains. This is consistent with our above findings (Fig. 2 and 4) and further supports our conclusions that vitamin B12 modulates porphyrin production in acne-associated type IA-2 strains and that porphyrin production can be modulated at the metabolic level. Our data also suggest that LA is an effective inhibitor of porphyrin biosynthesis in P. acnes and that the porphyrin biosynthesis pathway can be a potential therapeutic target for new acne treatments.
FIG 5 .

Small-molecule inhibitor reduced porphyrin production in P. acnes, and its inhibition was counteracted by vitamin B12 supplementation in acne-associated type IA-2 strains. P. acnes strains were cultured in medium with inhibitor LA (0.1 mg/ml) (grey bars) or without LA (white bars) or with both LA and vitamin B12 (10 µg/ml) added (black bars). LA significantly reduced porphyrin biosynthesis in all strains except for HL001PA1 (P < 0.0001). Vitamin B12 supplementation counteracted the inhibition of porphyrin biosynthesis by LA in acne-associated type IA-2 strains, but not in health-associated type II strains. Each bar represents the porphyrins produced by each strain normalized to the bacterial culture density. The means plus standard errors (error bars) of data obtained from at least three independent experiments with at least three replicates each are shown.
Health-associated strains harbor a porphyrin biosynthesis repressor gene, deoR.
To identify potential molecular mechanisms that can explain differences in porphyrin levels produced by acne- and health-associated P. acnes strains, we compared the porphyrin biosynthesis gene operon (hem) of 82 P. acnes strains (13), including the strains tested in this study. We found that all health-associated type II strains and a few type I strains (mainly IB-3 and IC strains) harbor an additional gene in the porphyrin biosynthesis operon, annotated as deoR transcriptional repressor (see Fig. S2 and S3 in the supplemental material). The deoR gene is located 13 bp upstream of the porphyrin biosynthesis gene cluster. This gene is absent in all acne-associated type IA-2 strains (13). To determine whether deoR is functional, we performed gene expression analysis and found that it was expressed in two of the three health-associated type II strains, HL001PA1 and HL042PA3, but not in HL103PA1 (Fig. 6). Characterizations of deoR-like repressors in multiple other species have revealed their roles in regulating cellular processes such as sugar phosphotransferase activity, daptomycin production, and fatty acid beta-oxidation (39–41). The close proximity of deoR to the porphyrin biosynthesis genes in the genome and the presence and expression of deoR in health-associated type II strains suggest that deoR may function as a transcriptional repressor in porphyrin biosynthesis. This may partly explain the low porphyrin levels produced by type II strains and their associations with healthy skin.
FIG 6 .
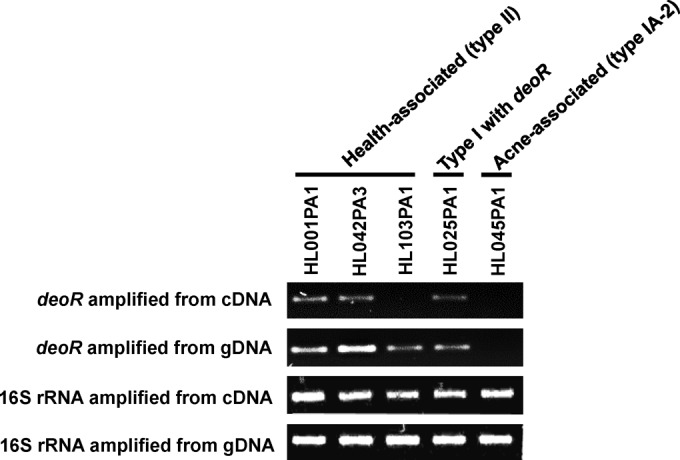
Health-associated strains carried and expressed deoR, a repressor gene in the porphyrin biosynthesis operon. deoR amplification from the cDNA and genomic DNA (gDNA) samples of multiple strains is shown in the gel image. 16S rRNA gene was used as a positive control. Strain HL045PA1, which is an acne-associated type IA-2 strain and does not carry deoR, is shown as a negative control.
Acne-associated type IA-2 strains and health-associated type II strains belong to distinct P. acnes lineages (11). To further investigate whether the low levels of porphyrins produced by health-associated type II strains were potentially due to repression by deoR and not due to other lineage differences, we examined the porphyrin levels produced by a type I P. acnes strain, HL025PA1. Strain HL025PA1 represents a distinct lineage within type I and is phylogenetically more similar to type IA-2 strains than to type II strains (see Fig. S3 in the supplemental material). Unlike most other type I strains, HL025PA1 carries and expresses deoR in its porphyrin biosynthesis operon (Fig. 6). We found that HL025PA1 produced porphyrins at a level similar to that of type II strains (2.7 µM; P = 0.31) (Fig. S4), significantly lower than type IA-2 strains (P = 0.0006). Consistent with the trend observed in health-associated type II strains, vitamin B12 supplementation resulted in transcriptional repression of the vitamin B12 biosynthesis pathway gene in HL025PA1 (Fig. 3), while its porphyrin production was unaffected by vitamin B12 supplementation (Fig. S4). This finding further supports an inhibitory role for deoR in porphyrin biosynthesis and may potentially explain the health association of type II strains.
Porphyrin (hem) gene cluster in P. acnes strains. All health-associated type II strains harbor a deoR transcription repressor (PPA0299, in green) in the porphyrin gene cluster. The number below each box represents the GeneID based on the gene annotation in strain KPA171202. The letters in black boxes indicate the names of the hem genes, which are porphyrin biosynthesis genes. PPA0300 and PPA0310 were not assigned a hem gene name. Download Figure S2, PDF file, 0.01 MB (13.2KB, pdf) .
Copyright © 2016 Johnson et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Presence of deoR in P. acnes lineages. P. acnes strains carrying deoR are shown in green in a phylogenetic tree constructed based on 82 P. acnes genomes. All type II strains harbor the deoR repressor. Asterisks denote the strains tested in this study. Acne index indicates the strain association with health and disease, as described previously by Fitz-Gibbon et al. (11). Download Figure S3, PDF file, 0.1 MB (76.7KB, pdf) .
Copyright © 2016 Johnson et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Similar to health-associated type II strains, strain HL025PA1 produced a low level of porphyrins and did not respond to vitamin B12 supplementation. Strain HL025PA1 was cultured in the medium with (black bars) or without (white bars) vitamin B12 (10 µg/ml). Health-associated type II strains, HL103PA1 and HL042PA3, are shown for comparison. Each bar represents the porphyrins produced by each strain normalized to the bacterial culture density. The means plus standard errors (error bars) of data obtained from at least three independent experiments with at least three replicates each are shown. Download Figure S4, PDF file, 0.01 MB (15.6KB, pdf) .
Copyright © 2016 Johnson et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
DISCUSSION
The role of the human microbiota in disease and health is not yet fully understood. In particular, the molecular mechanisms for different strains of the same commensal species with distinct functions in maintaining health or triggering disease have yet to be elucidated. While the dominant skin commensal, P. acnes, is thought to provide protection for skin from colonization of pathogens such as Staphylococcus aureus (42, 43), multiple studies have suggested that P. acnes can act as an opportunistic pathogen in various diseases, including sarcoidosis, SAPHO syndrome, endodontic lesions, eye infections, prosthetic joint infections, prostate cancer, and acne (1–4). In acne, the dominance of P. acnes on both acneic and healthy skin has long been a concern in defining the role of this bacterium in disease pathogenesis (3, 11, 44). Microbiome studies of the skin follicle have revealed strain-level differences of P. acnes in health and disease associations (11, 13, 14). However, the molecular mechanisms explaining the strain differences are not well understood.
In P. acnes, strain-level differences have been reported at the genomic level (13, 14) and proteomic level (17, 45, 46); however, their molecular link to health and disease remains to be defined. In this study, we investigated the strain-level differences in P. acnes at the metabolic level. We discovered that compared to health-associated type II strains, acne-associated type IA-2 strains produced significantly more porphyrins, a group of bacterial metabolites that induce inflammation in acne (Fig. 1). This finding is consistent with previous observations that porphyrin levels were higher in acneic skin than in healthy skin (24–26) and provides one potential molecular mechanism for the role of acne-associated type IA-2 strains in the disease pathogenesis.
This study confirmed our earlier finding that vitamin B12 modulates gene expression and metabolic activities of P. acnes in acne development (18) and further revealed that acne-associated type IA-2 strains, but not health-associated type II strains, responded to vitamin B12 supplementation with increased porphyrin production (Fig. 2). It has been documented that vitamin B12 supplementation leads to acne development in a subset of populations (18, 30–33). However, the determinants in individuals who respond to vitamin B12 supplementation and develop acne have not yet been identified. Our data show that vitamin B12 modulation of porphyrin production is strain specific and suggest that the P. acnes strain composition of an individual’s skin microbiota may contribute to vitamin B12-induced acne. Individuals harboring acne-associated type IA-2 strains are likely to be at increased risk for developing acne in response to high vitamin B12 levels due to the ability of their skin bacteria to produce more porphyrins. On the other hand, individuals whose skin is dominated by health-associated type II strains may have lower porphyrin levels produced by the bacteria, leading to a reduced risk for acne development when supplemented with vitamin B12.
This study also revealed a potential transcriptional repression mechanism of porphyrin biosynthesis in health-associated strains through deoR regulation (Fig. 6 and see Fig. S2 in the supplemental material). While the function and regulation of deoR in P. acnes require further investigation, the presence and expression of this gene suggest that deoR may play a role in the inhibition of porphyrin production in health-associated type II strains. P. acnes strain HL025PA1, although belonging to type I, represents a distinct lineage. It carries and expresses deoR. Lomholt and Kilian previously reported that only 1 of their 13 studied P. acnes isolates from this lineage (sequence type 27 [ST27], as designated by the Aarhus MLST9 scheme) was from severe acne and suggested an association of this lineage with healthy skin (12). This observation supports our theory of a role for deoR in skin health. As deoR expression was not detected in all tested health-associated type II strains, additional mechanisms regulating porphyrin production likely exist, warranting future investigation. The low levels of porphyrins produced in health-associated strains, especially strain HL001PA1, make these strains candidates for topical probiotics to potentially modify an acne-prone skin microbial community and return the skin to a healthy state. This could be a new strategy in future acne therapeutics.
Our current analysis is focused on two major P. acnes lineages, types IA-2 and II, which have been associated with acne and healthy skin, respectively. Further investigations of metabolic activities and their regulations including porphyrin production in strains from other lineages will shed light on additional bacterial factors contributing to disease or health.
Our study has implications in the development of novel acne therapies. Current acne treatments, such as antibiotics and retinoids, are often ineffective and can have adverse side effects. Moreover, the use of antibiotics has led to the emergence of antibiotic-resistant strains and an increase in treatment failure (47, 48). Although new acne therapeutics have been in demand for a long time, the unclear etiology of the disease has crippled the design of new and effective treatments over the past 3 decades. We demonstrated that LA at the concentration of 0.1 mg/ml effectively inhibited porphyrin biosynthesis in P. acnes strains (Fig. 5). This inhibition is consistent with the decreased porphobilinogen synthase activity observed in Pseudomonas aeruginosa upon LA treatment in vitro (38). LA is also known to inhibit bacterial growth; thus, a suboptimal concentration was used in this study to demonstrate the ability of this molecule to effectively reduce the porphyrin levels produced by P. acnes without significant inhibition of bacterial growth. In our study, we aimed to model a treatment strategy in which the virulence of acne-associated strains is targeted without disrupting the growth of health-associated strains. Other compounds, such as 4,6-dioxoheptanoic acid and isonicotinic acid hydrazide, also inhibit porphobilinogen synthase (49, 50) and can potentially be used to inhibit porphyrin biosynthesis. LA and other related small molecules that inhibit porphyrin biosynthesis in P. acnes are attractive drug candidates for the treatment of acne.
The benefit of the microbiota to human health is increasingly recognized. There is a need to improve our current approaches in treating microorganism-associated diseases. Most of the available approaches nonspecifically target the microbiota using broad-spectrum antibiotics and antimicrobials, potentially leading to a disruption in the colonization of beneficial microorganisms. This study presents an advance toward a better understanding of the beneficial microbiota and targeted therapeutics. By investigating the molecular mechanisms underlying the differences between health- and disease-associated strains, we suggest that inhibiting disease-associated strains with specific targets while maintaining or supplementing health-associated strains can potentially be a new strategy in the future for treating microorganism-associated diseases.
MATERIALS AND METHODS
P. acnes strains and cultures.
P. acnes strains used in this study (Table 1) were isolated previously as described by Fitz-Gibbon et al. (11). Briefly, each strain was isolated from the contents collected from multiple hair follicles on the nose of an acne patient or healthy individual. The sampled skin site of acne patients may or may not have visible acne lesions; therefore, the strains do not necessarily correspond to the diseased or healthy state. Four RT4 and RT5 P. acnes strains, HL053PA1, HL045PA1, HL043PA1, and HL043PA2, were selected to represent type IA-2 strains, which were associated with the disease based on a 16S rRNA metagenomic study (11). Like most of the RT4 and RT5 strains, these strains harbor mutations in their 16S and 23S rRNA genes, which confer antibiotic resistance. Three RT2 and RT6 strains, HL001PA1, HL103PA1, and HL042PA3, were selected to represent type II strains that were associated with healthy skin (10–12, 15). To confirm the findings, three other type II strains, HL110PA3, HL106PA1, and HL050PA2, were also tested. The genome sequences of these 10 strains were reported previously (11, 13). For each experiment, 5 ml of reinforced clostridial broth was inoculated with 5 × 105 P. acnes cells per ml of culture. Cultures were grown to stationary phase anaerobically at 37°C in a light-protected box. Cultures were supplemented on day 0 with vitamin B12 (10 µg/ml), and/or 5-ALA (0.1 mg/ml), and/or LA (0.1 mg/ml). For controls, P. acnes strains were also cultured without supplementation. Three to five independent experiments with at least three replicates per experiment were performed for each strain, except for strains HL110PA3, HL106PA1, and HL050PA2, which were tested in only one experiment with three replicates.
Extraction, identification, and quantification of extracellular porphyrins.
For each strain, porphyrins were extracted by the method of Kang et al. (18). Briefly, 500 µl of bacterial culture was extracted in ethyl acetate and acetic acid (4:1, vol/vol) and solubilized in 1.5 M HCl. The absorbance at 405 nm was measured from 200 µl of the soluble phase using a Tecan Genios spectrophotometer M1000 (Tecan U.S. Inc., Morrisville, NC). The standard curve to convert absorbance to concentration was generated using coproporphyrin III standards of known concentration (catalog no. C654-3; Frontier SCI). We measured the porphyrin level in P. acnes strain KPA171202 using our porphyrin extraction method and the quantification method of Wollenberg et al. (36). Our measurement, which was 206 pmol/mg, is consistent with the reported value of 220 pmol/mg by Wollenberg et al. (36). This indicates that our method is comparable to the methods in previous studies. Porphyrins were extracted from cultures at stationary phase. The results from cultures at mid-log phase had a consistent trend found in cultures at stationary phase. Bacterial culture density was measured at an optical density at 595 nm (OD595) for normalization of porphyrin levels. For mass spectrometry experiments, extracted porphyrins were directly injected onto an Agilent 6460 triple quadrupole liquid chromatography-mass spectrometry (LC/MS) system, and each strain’s m/z in the negative-ion mode was identified.
Statistical analysis for porphyrin production comparisons.
The average amount of porphyrins produced by each strain under each culture condition was calculated based on the data from at least three independent experiments, with at least three replicates for each. The porphyrin levels between strains, groups (acne-associated versus health-associated), and culture conditions (with treatment versus without treatment) were estimated in a linear mixed-effect model, with random intercepts by trial to account for trial effects. P values for specific comparisons among groups were corrected using Tukey’s method. All statistical analysis was performed using R software (version 3.1.3).
Identification of deoR transcription repressor.
The sequences of the porphyrin gene clusters from 82 P. acnes genomes (13) were aligned using the multiple sequence alignment software (Clustal W2) (51). The deoR transcription repressor (PPA0299) was identified as an extra genomic element found in all health-associated type II strains and a few type I strains.
RNA extraction and cDNA synthesis.
For cbiL and deoR gene expression analysis, cells were lysed with bead beating. Total RNA was extracted using the standard phenol-chloroform method and purified using RNeasy kit (Qiagen). DNA was removed using the Turbo DNA-free kit (Life Technologies). RNA quality was assessed using gel electrophoresis. Single-stranded cDNA was synthesized using SuperScript III first-strand synthesis supermix (Life Technologies).
Analysis of cbiL gene expression.
qRT-PCR was performed using the LightCycler 480 high-resolution melting master mix (Roche) on a LightCycler 480 instrument (Roche) with the following primers: cbiL-forward, 5′ GCGCGAGGCAGACGTGATCC 3′, and cbiL-reverse, 5′ GACACCGGACCTCTCCCGCA 3′. The following qRT-PCR protocol was used: initial denaturation at 95°C for 5 min, followed by 50 cycles, with 1 cycle consisting of 95°C for 10 s, 62°C for 30 s, and 72°C for 30 s. The fold change in cbiL gene expression between cultures with vitamin B12 supplementation and cultures without supplementation was calculated. The gene expression level of cbiL in each sample was normalized against the 16S rRNA transcript level. Melting curve analysis was performed using the LightCycler 480 software version 1.5 (Roche) to verify the specificity of the amplified products based on their melting temperatures. All reactions were run in triplicate.
Analysis of deoR gene expression.
deoR was amplified from the cDNA and genomic DNA of several P. acnes strains using a C1000 thermal cycler (BioRad). The following primers were used in the PCR: deoR-forward, 5′ CTGGCACGAGAAGGAACAA 3′, and deoR-reverse, 5′ GAATCGAGCAGAACTAGGTCAC 3′. The following PCR protocol was used: initial denaturation at 95°C for 5 min, followed by 35 cycles, with 1 cycle consisting of 95°C for 10 s, 62°C for 30 s, and 72°C for 30 s, followed by one cycle at 72°C for 5 min. Amplified deoR products were visualized on a 2% agarose gel. The amplification of 16S rRNA was included as a positive control. Acne-associated strain HL045PA1 was used as a negative control for deoR expression.
ACKNOWLEDGMENTS
We thank Kym Faull in the UCLA Pasarow Mass Spectrometry Laboratory as well as the UCLA-DOE Biochemistry Instrument Facility for support on experimental instruments. We thank Andy Lin in the UCLA Institute for Digital Research and Education (IDRE) for statistics consultation. We also thank Emily Curd and Jared Liu for technical help.
This research was funded by NIH grant R01GM099530 from the National Institute of General Medical Sciences. T.J. is supported by Ruth L. Kirschstein National Research Service Award AI007323.
T.J. performed the experiments, analyzed the data, and wrote the manuscript. D.K. analyzed the data and revised the manuscript. E.B. performed the experiments and revised the manuscript. H.L. conceived and directed the project and wrote the manuscript.
The Regents of the University of California are filing a patent application related to this study, which names T.J., D.K., E.B., and H.L. as inventors. H.L. is a cofounder and consultant of Skinomics, Inc.
REFERENCES
- 1.Nagy I, Pivarcsi A, Kis K, Koreck A, Bodai L, McDowell A, Seltmann H, Patrick S, Zouboulis CC, Kemény L. 2006. Propionibacterium acnes and lipopolysaccharide induce the expression of antimicrobial peptides and proinflammatory cytokines/chemokines in human sebocytes. Microbes Infect 8:2195–2205. doi: 10.1016/j.micinf.2006.04.001. [DOI] [PubMed] [Google Scholar]
- 2.Bojar RA, Holland KT. 2004. Acne and Propionibacterium acnes. Clin Dermatol 22:375–379. doi: 10.1016/j.clindermatol.2004.03.005. [DOI] [PubMed] [Google Scholar]
- 3.Leeming JP, Holland KT, Cuncliffe WJ. 1988. The microbial colonization of inflamed acne vulgaris lesions. Br J Dermatol 118:203–208. doi: 10.1111/j.1365-2133.1988.tb01775.x. [DOI] [PubMed] [Google Scholar]
- 4.Perry A, Lambert P. 2011. Propionibacterium acnes: infection beyond the skin. Expert Rev Anti Infect Ther 9:1149–1156. doi: 10.1586/eri.11.137. [DOI] [PubMed] [Google Scholar]
- 5.Cunliffe WJ, Gould DJ. 1979. Prevalence of facial acne vulgaris in late adolescence and in adults. Br Med J 1:1109–1110. doi: 10.1136/bmj.1.6171.1109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.White GM. 1998. Recent findings in the epidemiologic evidence, classification, and subtypes of acne vulgaris. J Am Acad Dermatol 39:S34–S37. doi: 10.1016/S0190-9622(98)70442-6. [DOI] [PubMed] [Google Scholar]
- 7.Gupta MA, Gupta AK. 1998. Depression and suicidal ideation in dermatology patients with acne, alopecia areata, atopic dermatitis and psoriasis. Br J Dermatol 139:846–850. doi: 10.1046/j.1365-2133.1998.02511.x. [DOI] [PubMed] [Google Scholar]
- 8.Mallon E, Newton JN, Klassen A, Stewart-Brown SL, Ryan TJ, Finlay AY. 1999. The quality of life in acne: a comparison with general medical conditions using generic questionnaires. Br J Dermatol 140:672–676. doi: 10.1046/j.1365-2133.1999.02768.x. [DOI] [PubMed] [Google Scholar]
- 9.McDowell A, Gao A, Barnard E, Fink C, Murray PI, Dowson CG, Nagy I, Lambert PA, Patrick S. 2011. A novel multilocus sequence typing scheme for the opportunistic pathogen Propionibacterium acnes and characterization of type I cell surface-associated antigens. Microbiology 157:1990–2003. doi: 10.1099/mic.0.049676-0. [DOI] [PubMed] [Google Scholar]
- 10.McDowell A, Barnard E, Nagy I, Gao A, Tomida S, Li H, Eady A, Cove J, Nord CE, Patrick S. 2012. An expanded multilocus sequence typing scheme for Propionibacterium acnes: investigation of ‘pathogenic’, ‘commensal’ and antibiotic resistant strains. PLoS One 7:e41480. doi: 10.1371/journal.pone.0041480. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fitz-Gibbon S, Tomida S, Chiu BH, Nguyen L, Du C, Liu M, Elashoff D, Erfe MC, Loncaric A, Kim J, Modlin RL, Miller JF, Sodergren E, Craft N, Weinstock GM, Li H. 2013. Propionibacterium acnes strain populations in the human skin microbiome associated with acne. J Invest Dermatol 133:2152–2160. doi: 10.1038/jid.2013.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lomholt HB, Kilian M. 2010. Population genetic analysis of Propionibacterium acnes identifies a subpopulation and epidemic clones associated with acne. PLoS One 5:e12277. doi: 10.1371/journal.pone.0012277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tomida S, Nguyen L, Chiu BH, Liu J, Sodergren E, Weinstock GM, Li H. 2013. Pan-genome and comparative genome analyses of Propionibacterium acnes reveal its genomic diversity in the healthy and diseased human skin microbiome. mBio 4:e00003-13. doi: 10.1128/mBio.00003-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kasimatis G, Fitz-Gibbon S, Tomida S, Wong M, Li H. 2013. Analysis of complete genomes of Propionibacterium acnes reveals a novel plasmid and increased pseudogenes in an acne associated strain. BioMed Res Int 2013:918320. doi: 10.1155/2013/918320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.McDowell A, Nagy I, Magyari M, Barnard E, Patrick S. 2013. The opportunistic pathogen Propionibacterium acnes: insights into typing, human disease, clonal diversification and CAMP factor evolution. PLoS One 8:e70897. doi: 10.1371/journal.pone.0070897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Brüggemann H, Henne A, Hoster F, Liesegang H, Wiezer A, Strittmatter A, Hujer S, Dürre P, Gottschalk G. 2004. The complete genome sequence of Propionibacterium acnes, a commensal of human skin. Science 305:671–673. doi: 10.1126/science.1100330. [DOI] [PubMed] [Google Scholar]
- 17.Holland C, Mak TN, Zimny-Arndt U, Schmid M, Meyer TF, Jungblut PR, Brüggemann H. 2010. Proteomic identification of secreted proteins of Propionibacterium acnes. BMC Microbiol 10:230. doi: 10.1186/1471-2180-10-230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Kang D, Shi B, Erfe MC, Craft N, Li H. 2015. Vitamin B12 modulates the transcriptome of the skin microbiota in acne pathogenesis. Sci Transl Med 7:293ra103. doi: 10.1126/scitranslmed.aab2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Valanne S, McDowell A, Ramage G, Tunney MM, Einarsson GG, O’Hagan S, Wisdom GB, Fairley D, Bhatia A, Maisonneuve JF, Lodes M, Persing DH, Patrick S. 2005. CAMP factor homologues in Propionibacterium acnes: a new protein family differentially expressed by types I and II. Microbiology 151:1369–1379. doi: 10.1099/mic.0.27788-0. [DOI] [PubMed] [Google Scholar]
- 20.Lodes MJ, Secrist H, Benson DR, Jen S, Shanebeck KD, Guderian J, Maisonneuve JF, Bhatia A, Persing D, Patrick S, Skeiky YA. 2006. Variable expression of immunoreactive surface proteins of Propionibacterium acnes. Microbiology 152:3667–3681. doi: 10.1099/mic.0.29219-0. [DOI] [PubMed] [Google Scholar]
- 21.Fanta D, Formanek I, Poitschek C, Thurner J. 1978. Porphyrin synthesis of Propionibacterium acnes in acne and seborrhea. Arch Dermatol Res 261:175–179. (In German.) doi: 10.1007/BF00447162. [DOI] [PubMed] [Google Scholar]
- 22.Schaller M, Loewenstein M, Borelli C, Jacob K, Vogeser M, Burgdorf WH, Plewig G. 2005. Induction of a chemoattractive proinflammatory cytokine response after stimulation of keratinocytes with Propionibacterium acnes and coproporphyrin III. Br J Dermatol 153:66–71. doi: 10.1111/j.1365-2133.2005.06530.x. [DOI] [PubMed] [Google Scholar]
- 23.Saint-Leger D, Bague A, Cohen E, Chivot M. 1986. A possible role for squalene in the pathogenesis of acne. I. In vitro study of squalene oxidation. Br J Dermatol 114:535–542. doi: 10.1111/j.1365-2133.1986.tb04060.x. [DOI] [PubMed] [Google Scholar]
- 24.Nacht S, Gans EH, McGinley KJ, Kligman AM. 1983. Comparative activity of benzoyl peroxide and hexachlorophene. In vivo studies against Propionibacterium acnes in humans. Arch Dermatol 119:577–579. doi: 10.1001/archderm.119.7.577. [DOI] [PubMed] [Google Scholar]
- 25.Meffert H, Gaunitz K, Gutewort T, Amlong UJ. 1990. Therapy of acne with visible light. Decreased irradiation time by using a blue-light high-energy lamp. Dermatol Monatsschr 176:597–603. (In German.). [PubMed] [Google Scholar]
- 26.Meyer K, Pappas A, Dunn K, Cula GO, Seo I, Ruvolo E, Batchvarova N. 2015. Evaluation of seasonal changes in facial skin with and without acne. J Drugs Dermatol 14:593–601. [PubMed] [Google Scholar]
- 27.Martin R, Kahn G, Gooding J, Brown G. 1973. Cutaneous porphyrin fluorescence as an indicator of antibiotic absorption and effectiveness. Cutis 12:758–764. [Google Scholar]
- 28.Mills OH, Kligman AM, Pochi P, Comite H. 1986. Comparing 2.5%, 5%, and 10% benzoyl peroxide on inflammatory acne vulgaris. Int J Dermatol 25:664–667. [DOI] [PubMed] [Google Scholar]
- 29.Borelli C, Merk K, Schaller M, Jacob K, Vogeser M, Weindl G, Berger U, Plewig G. 2006. In vivo porphyrin production by P. acnes in untreated acne patients and its modulation by acne treatment. Acta Derm Venereol 86:316–319. doi: 10.2340/00015555-0088. [DOI] [PubMed] [Google Scholar]
- 30.Jadassohn W, Paillard R, Hofer R, Golaz M. 1958. Vitamin e B12 et poussée aenéïforme. Dermatologica 116:349 (In French.) [Google Scholar]
- 31.Puissant A, Vanbremeersch F, Monfort J, Lamberton JN. 1967. A new iatrogenic dermatosis: acne caused by vitamin B 12. Bull Soc Fr Dermatol Syphiligr 74:813–815. (In French.) [PubMed] [Google Scholar]
- 32.Dugois P, Amblard P, Imbert R, de Bignicourt B. 1969. Acne due to vitamin B 12. Bull Soc Fr Dermatol Syphiligr 76:382–383. (In French.) [PubMed] [Google Scholar]
- 33.Balta I, Ozuguz P. 2014. Vitamin B12-induced acneiform eruption. Cutan Ocul Toxicol 33:94–95. doi: 10.3109/15569527.2013.808657. [DOI] [PubMed] [Google Scholar]
- 34.Bykhovskii V, Zaitseva NI, Bukin VN. 1968. Possible competition in the use of delta-aminolevulinic acid for the biosynthesis of vitamin B12 and porphyrin by resting suspensions of Propionibacterium shermanii. Dokl Akad Nauk SSSR 180:232–234. (In Russian.) [PubMed] [Google Scholar]
- 35.Lee WL, Shalita AR, Poh-Fitzpatrick MB. 1978. Comparative studies of porphyrin production in Propionibacterium acnes and Propionibacterium granulosum. J Bacteriol 133:811–815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wollenberg MS, Claesen J, Escapa IF, Aldridge KL, Fischbach MA, Lemon KP. 2014. Propionibacterium-produced coproporphyrin III induces Staphylococcus aureus aggregation and biofilm formation. mBio 5:e01286-14. doi: 10.1128/mBio.01286-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Cornelius CE III, Ludwig GD. 1967. Red fluorescence of comedones: production of porphyrins by Corynebacterium acnes. J Invest Dermatol 49:368–370. [PubMed] [Google Scholar]
- 38.Frère F, Schubert WD, Stauffer F, Frankenberg N, Neier R, Jahn D, Heinz DW. 2002. Structure of porphobilinogen synthase from Pseudomonas aeruginosa in complex with 5-fluorolevulinic acid suggests a double Schiff base mechanism. J Mol Biol 320:237–247. doi: 10.1016/S0022-2836(02)00472-2. [DOI] [PubMed] [Google Scholar]
- 39.Gaigalat L, Schlüter JP, Hartmann M, Mormann S, Tauch A, Pühler A, Kalinowski J. 2007. The DeoR-type transcriptional regulator SugR acts as a repressor for genes encoding the phosphoenolpyruvate:sugar phosphotransferase system (PTS) in Corynebacterium glutamicum. BMC Mol Biol 8:104. doi: 10.1186/1471-2199-8-104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wang F, Ren NN, Luo S, Chen XX, Mao XM, Li YQ. 2014. DptR2, a DeoR-type autoregulator, is required for daptomycin production in Streptomyces roseosporus. Gene 544:208–215. doi: 10.1016/j.gene.2014.04.044. [DOI] [PubMed] [Google Scholar]
- 41.Elgrably-Weiss M, Schlosser-Silverman E, Rosenshine I, Altuvia S. 2006. DeoT, a DeoR-type transcriptional regulator of multiple target genes. FEMS Microbiol Lett 254:141–148. doi: 10.1111/j.1574-6968.2005.00020.x. [DOI] [PubMed] [Google Scholar]
- 42.Shu M, Wang Y, Yu J, Kuo S, Coda A, Jiang Y, Gallo RL, Huang CM. 2013. Fermentation of Propionibacterium acnes, a commensal bacterium in the human skin microbiome, as skin probiotics against methicillin-resistant Staphylococcus aureus. PLoS One 8:e55380. doi: 10.1371/journal.pone.0055380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Sikorska H, Smoragiewicz W. 2013. Role of probiotics in the prevention and treatment of meticillin-resistant Staphylococcus aureus infections. Int J Antimicrob Agents 42:475–481. doi: 10.1016/j.ijantimicag.2013.08.003. [DOI] [PubMed] [Google Scholar]
- 44.Jahns AC, Lundskog B, Ganceviciene R, Palmer RH, Golovleva I, Zouboulis CC, McDowell A, Patrick S, Alexeyev OA. 2012. An increased incidence of Propionibacterium acnes biofilms in acne vulgaris: a case-control study. Br J Dermatol 167:50–58. doi: 10.1111/j.1365-2133.2012.10897.x. [DOI] [PubMed] [Google Scholar]
- 45.Dekio I, Culak R, Fang M, Ball G, Gharbia S, Shah HN. 2013. Correlation between phylogroups and intracellular proteomes of Propionibacterium acnes and differences in the protein expression profiles between anaerobically and aerobically grown cells. Biomed Res Int 2013:151797. doi: 10.1155/2013/151797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nagy E, Urbán E, Becker S, Kostrzewa M, Vörös A, Hunyadkürti J, Nagy I. 2013. MALDI-TOF MS fingerprinting facilitates rapid discrimination of phylotypes I, II and III of Propionibacterium acnes. Anaerobe 20:20–26. doi: 10.1016/j.anaerobe.2013.01.007. [DOI] [PubMed] [Google Scholar]
- 47.Leyden JJ, McGinley KJ, Cavalieri S, Webster GF, Mills OH, Kligman AM. 1983. Propionibacterium acnes resistance to antibiotics in acne patients. J Am Acad Dermatol 8:41–45. doi: 10.1016/S0190-9622(83)70005-8. [DOI] [PubMed] [Google Scholar]
- 48.Eady EA, Cove JH, Holland KT, Cunliffe WJ. 1989. Erythromycin resistant propionibacteria in antibiotic treated acne patients: association with therapeutic failure. Br J Dermatol 121:51–57. doi: 10.1111/j.1365-2133.1989.tb01399.x. [DOI] [PubMed] [Google Scholar]
- 49.Schuster A, Harel E. 1985. A low molecular weight polypeptide which accumulates upon inhibition of porphyrin biosynthesis in maize. Plant Physiol 77:648–652. doi: 10.1104/pp.77.3.648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Kotal P, Klepácek I, Jirsa M, Kordac V. 1989. Inhibition of porphyrin synthesis by isonicotinic acid hydrazide in models simulating porphyria. Sb Lek 91:161–168. (In Czech.). [PubMed] [Google Scholar]
- 51.Thompson JD, Higgins DG, Gibson TJ. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res 22:4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Coproporphyrin was the dominant porphyrin isoform produced by P. acnes. Porphyrins secreted by P. acnes have a mass spectrum characteristic of the monoisotopic coproporphyrin isoform ([M + H]+ = 655.3). The doubly charged parent ion was also observed ([M + 2H]2+ = 328.2). Download Figure S1, PDF file, 0.1 MB (53.1KB, pdf) .
Copyright © 2016 Johnson et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Porphyrin (hem) gene cluster in P. acnes strains. All health-associated type II strains harbor a deoR transcription repressor (PPA0299, in green) in the porphyrin gene cluster. The number below each box represents the GeneID based on the gene annotation in strain KPA171202. The letters in black boxes indicate the names of the hem genes, which are porphyrin biosynthesis genes. PPA0300 and PPA0310 were not assigned a hem gene name. Download Figure S2, PDF file, 0.01 MB (13.2KB, pdf) .
Copyright © 2016 Johnson et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Presence of deoR in P. acnes lineages. P. acnes strains carrying deoR are shown in green in a phylogenetic tree constructed based on 82 P. acnes genomes. All type II strains harbor the deoR repressor. Asterisks denote the strains tested in this study. Acne index indicates the strain association with health and disease, as described previously by Fitz-Gibbon et al. (11). Download Figure S3, PDF file, 0.1 MB (76.7KB, pdf) .
Copyright © 2016 Johnson et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.
Similar to health-associated type II strains, strain HL025PA1 produced a low level of porphyrins and did not respond to vitamin B12 supplementation. Strain HL025PA1 was cultured in the medium with (black bars) or without (white bars) vitamin B12 (10 µg/ml). Health-associated type II strains, HL103PA1 and HL042PA3, are shown for comparison. Each bar represents the porphyrins produced by each strain normalized to the bacterial culture density. The means plus standard errors (error bars) of data obtained from at least three independent experiments with at least three replicates each are shown. Download Figure S4, PDF file, 0.01 MB (15.6KB, pdf) .
Copyright © 2016 Johnson et al.
This content is distributed under the terms of the Creative Commons Attribution 4.0 International license.