

Abstract
The ancestor of Paulinella chromatophora established a symbiotic relationship with cyanobacteria related to the Prochloroccocus/Synechococcus clade. This event has been described as a second primary endosymbiosis leading to a plastid in the making. Based on the rate of pseudogene disintegration in the endosymbiotic bacteria Buchnera aphidicola, it was suggested that the chromatophore in P. chromatophora has a minimum age of ~60 Myr. Here we revisit this estimation by using a lognormal relaxed molecular clock on the 18S rRNA of P. chromatophora. Our time estimates show that depending on the assumptions made to calibrate the molecular clock, P. chromatophora diverged from heterotrophic Paulinella spp. ~ 90 to 140 Myr ago, thus establishing a maximum date for the origin of the chromatophore.
Introduction
Mitochondria and plastids evolved from freeliving bacteria by symbiogenesis more than one billion years ago1. Both events boosted the evolution of eukaryotes by expanding their metabolic abilities. Primary endosymbiosis leading to organelles (i.e., mitochondria and plastids) was thought to be unique in the history of life until the recent discovery of an independent primary endosymbiosis in Paulinella chromatophora 2. This thecate filose amoeba hosts in its cytoplasm photosynthetic organelles of cyanobacterial origin, called chromatophores. Phylogenetic analysis of 16S rRNA showed that the chromatophores originated from marine αcyanobacteria from the Prochlorococcus/Synechococcus clade. In contrast, “classical” plastids of plants and algae evolved from an ancient unknown lineage of cyanobacteria3.
The genome of the cyanobacteria Synechococcus WH5701, one of the closest known free-living relatives of the chromatophore with a sequenced genome, has 2917 protein-coding genes4. In contrast, the chromatophores of P. chromatophora strains FK01 and M0880/a contain between 841 and 867 protein-coding genes respectively5 , 6. Remaining genes in the chromatophore suggest a strong metabolic interdependence with the amoebal nucleocytoplasm. The discovery that proteins of the chromatophore photosynthetic apparatus are encoded in the host genome and imported back into the cyanobacterial-derived compartment, reinforces the suggestion that the chromatophore is a bona fide primary organelle7.
It is likely that the origin of the chromatophore is one or two orders of magnitude more recent than the establishment of the primary plastids of plants and algae. But, how ancient is the chromatophore? Since there is no fossil record of P. chromatophora or its close relatives, it is difficult to have a precise answer to this question. However, an initial guess suggested the chromatophore has a minimum age of 60 Myr. This was based on the proposal that in Buchnera aphidicola (Aphid’s endosymbiotic bacteria) a pseudogene needs between 40-60 Myr to disintegrate completely8. Since the genome of the chromatophore has pseudogenes, the same tempo was extrapolated for the chromatophore6.
Surprisingly, the suggestion that the chromatophore in P. chromatophora has a minimum age of 60 Myr underwent a change in part of the subsequent literature indicating that P. chromatophora has ~60 Myr. Clearly, this is an assertion that requires clarification and further analysis. The recent identification by single cell genomics of non-photosynthetic close relatives of P. chromatophora 9 and novel attempts to calibrate the origin of major eukaryotic groups10 , 11 offer an opportunity to revisit the origin in time of this extraordinary symbiosis.
Methods
Taxon sampling. Based on previous published phylogenetic analyses we retrieved a sample of 18S rRNA sequences from rhizaria and stramenopila from SILVA database12. These included sequences from: i) the phylogenetic analysis whereby close relatives of P. cromatophora were identified9; ii) the phylogenetic analyses describing divergence times of major eukaryotic groups10 , 11; and iii) sequences from phylogenetic analyses of euglyphids13 and diatoms14. We did not include sequences from foraminifera due to their high rate of evolution reflected in extreme long branches. Sequences were aligned with MUSCLE15. The final alignment contained 43 sequences and 1128 aligned positions without gaps. The phylogenetic tree reconstructed from these sequences (see below) is in general terms congruent with published phylogenetic analyses.
Molecular clock analyses. BEAUTi was used to prepare xml files. Time trees were inferred by using a lognormal relaxed molecular clock as implemented in BEAST 216. To select the model of evolution we used jModelTest17. The Tamura and Nei 1993 model (TrN) plus gamma distribution and an estimated proportion of invariant sites were rated best by the bayesian information criterion (BIC). Therefore, all the analyses were conducted under this model of evolution. In addition, we used a Yule tree prior for all analyses.
To calibrate the clock, we relied upon several sources of information (Fig 2). These included organismal as well as chemical fossils. We also used previous published estimates of the time divergence between rhizaria and stramenoplia. These sources of information were organized into four different calibration schemes. All four schemes used information from non-controversial fossil record (evidences: a, b, c and d). However, the schemes differ on the use of soft evidence from previous molecular clock analyses (evidence: e.1 and e.2); and on the use of evidence from vaseshaped microfossils (VSM) described by18. VSM were originally assigned to rhizaria. However, a posterior molecular clock analysis favored a reinterpretation of VSM as members of amoebozoa10. Thus, the four calibration schemes were: (a, b, c, d, e.1); (a, b, c, d, e.1, f); (a, b, c, d, e.2); (a, b, c, d).
Priors on the age of nodes were adjusted to reflect confidence on time divergence. For instance, we assigned a strong prior on the origin of rhizosolenid diatoms because the origin of this lineage has been dated to 91.5 ± 1.5 Myr based on chemical fossils14. The 95% probability distribution on this case is between 91.5 to 97.0 Myr. Other calibrations received less strong priors, as indicated by the larger number of years contained along the 2.5% to 9.5% quantiles of the shape column from Table 1. Priors used for the origin of pennate diatoms and for the origins of earliest diatoms were based on those used by11 and represent minimal divergence times. The prior used for the origin of Eugliphydae was based on the supposition that the group is much older than the fossil evidence as suggested by19. This prior also represents a minimal divergence time. Finally, to assign a prior to the divergence in time of rhizaria from stramenopila we first averaged BEAST time estimates published by11 and then we parametrized a normal distribution to include the variability of these estimates. We used this normal distribution as a prior. We constructed a second prior for the same divergence of rhizaria from stramenopila but now based on the inference published by10. All sequences within each prior were restricted to be monophyletic. The root of the tree was determined by making rhizaria monophyletic.
Calibration constrains.
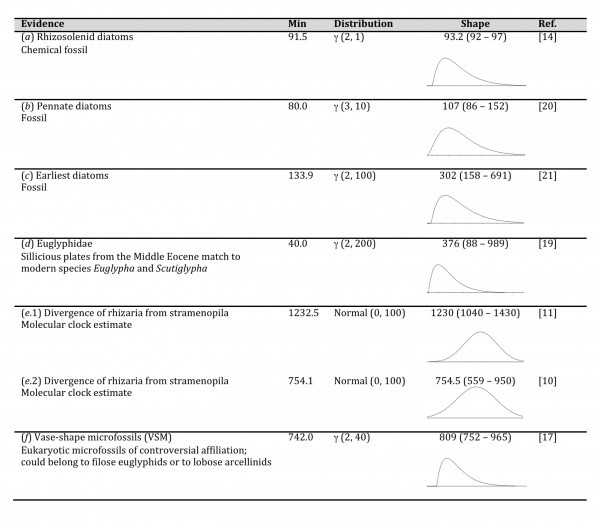
Evidence: used to derive priors for calibration constraints; Min: Minimal divergence time (Offset parameter in BEAUTi) in Myr; Distribution: Used to model each prior together with their respective parameter values (alpha α and beta β for gamma γ and mean and sigma σ for normal distributions); Shape: shows the median for each distribution and the values containing the 2.5% and 97.5% quantiles. Stronger priors span few years between the 2.5% and 97.5% quantiles.
We evaluated each calibration scheme by looking at convergence and ESS > 200 in Tracer after running chains of length 1010 and sampling each 104 generations. We discarded 10% of generations as burnin. Finally, we conducted all analyses without sequences (i.e., sampling from priors) to determine the impact of priors and to test whether sequences are informative on estimated divergence dates. Trees were obtained with TreeAnnotator and visualised with FigTree. We provide BEAUTi xml files of the four calibration schemes for BEAST 2 analyses here.
Results and discussion
In Fig. 1 we show an estimate of the divergence in time of P. chromatophora by using a lognormal relaxed clock on 18S rRNA. To calibrate this tree we relied on: a) the origin of rhizosolenid diatoms, which is known with high confidence (91.5 ± 1.5 Myr)14; b) a minimal time divergence of pennate diatoms (80 Myr)11 , 20; c) a minimal time divergence for diatoms (133.9 Myr)11 , 21; d) a minimal time divergence of Euglyphidae 40 Myr19; and e.1) a time estimation of the divergence of rhizaria from stramenopila ~1232 Myr11. This gave us an estimation for the origin of P. chromatophora of 93.6 Myr (38.1 – 138.2).
The origin of P. chromatophora in time according to SSU rRNA lognormal molecular clock.
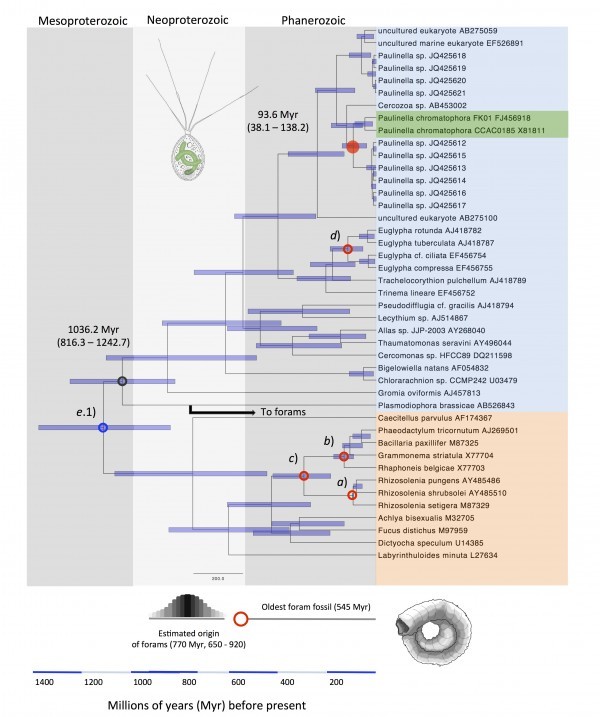
Tree calibrated under the scheme (a, b, c, d, e.1). Small open red circles indicate calibrations based on fossil record (a, b, c, and d); small open blue circle indicate calibration based on previous estimation (e.1); red full circle, estimated age of P. chromatophora. Notice that the divergence in time of Plasmodiophora brassicae (black open circle) is consistent with the proposed origin in time of forams. Species names in orange: stramenopila; species name in blue and green: rhizaria.
However other calibration schemes are possible. For instance, we can use the report of vase-shaped microfossils (VSM) of ~742 to 770 Myr (calibration f) that were described tentatively as members of Euglyphida18 together with calibrations a, b, c, d and e.1, to get an estimate of 141.4 Myr (48.7 – 210.4) for the origin of P. chromatophora. Alternatively, we can follow the suggestions that VSMs belong to amoebozoa and that rhizaria diverged from stramenopila 754.1 Myr (639.8 – 903.3)10 together with calibration points a, b, c, d and e.2 to get an estimate of 55.8 Myr (25.4 – 78.6) for the origin of P. chromatophora. Finally, we can estimate the divergence of P. chromatophora based only on calibration points a, b, c and d; thus avoiding controversial VSM fossils (calibration f) and the soft information provided from previous estimates of the divergence between rhizaria and stramenopila (calibrations e.1 and e.2). By this, we get an estimate of 47.9 Myr (28.8 – 64.9).
Which of the four time estimates is closer to the true divergence time of P. chromatophora? Foraminifera offer a clue. Foraminifera belong to rhizaria and as well as diatoms have a detailed fossil record. However, their 18S sequences are too divergent to include them in our analysis. The first appearance of forams in the fossil record is dated at 545 Myr22. Recent molecular time estimates of forams suggest this group originated 770 Myr (650 – 920)23. A phylogenetic analysis based on 15 concatenated genes shows that forams are a sister group of Plasmodiophora brassicae with the exclusion of Gromia oviformis as an outgroup11. Therefore, the origin of the lineage leading to P. brassicae in our tree has to be at least as old as the proposed origin of forams. By looking at our time estimates, it happens that only the two proposals that are based on a time of ~1232 Myr for the origin of rhizaria (i.e., that uses calibration e.1) result in a date for the origin of P. brassicae that is congruent with the estimated origin of forams (Fig. 3).
Concordance between different time divergence inferences and earliest forams in the fossil record.
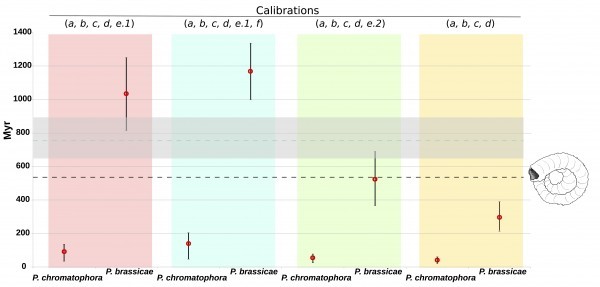
Only calibration schemes assuming a time divergence of rhizaria from stramenopila of ~1232 Myr (e.1) are consistent with the fossil record and proposed time divergence of forams. The horizontal black broken line represents the date of the oldest foram fossil at 545 Myr. The blue broken line represents the estimated origin of forams 770 Myr (650 – 920).
Although the 95% confidence intervals are rater large, the estimates of 93.6 and 141.4 Myr are consistent with the original suggestion that the chromatophore in P. chromatophora has a minimum age of 60 Myr6; and with a more recent proposal of a maximum age of 200 Myr for the divergence of P. chromatophora from Euglypha rotunda24.
If our estimates are correct, the two strains of P. chromatophora diverged from each other about (45.7 – 64.7 Myr). The genomes of the chromatophores in these two strains differ in about 33 genes5. This give us a rate of ~ 0.25 to 0.36 gene inactivations per million year since the divergence of the two strains. These show that genome reduction in the chromatophore is a process that continues slowly however stately. Whether these genes are been lost by genetic drift or natural selection is still a mater of research. Further refinements on the time of origin of P. chromatophora and their chromatophores are expected as our understanding of the evolution of rhizaria and the eukaryotic tree of life improves.
Acknowledgments
We thank Professor Adrian Reyes-Prieto, Mtra. Diana Rossetti and an anonymous reviewer for useful suggestions to the manuscript. We also thank Sofía Delaye for providing drawings of protists.
Biography
Biologist. I did my PhD under the advice of Professor Antonio Lazcano, working on the nature of the last universal common ancestor. Then, I moved to Universidad de Valencia to do a postdoc working on the evolution of symbiotic bacteria. Nowadays, I work at CINVESTAV Irapuato were I lead a group on evolutionary genomics.
Funding Statement
BPZ wishes to thank CINVESTAV Irapuato for funding and support. The authors also declare that no competing interests exist.
Contributor Information
Luis Delaye, CINVESTAV Irapuato.
Bernardo Pérez-Zamorano, Ingeniería Genética, CINVESTAV - Irapuato; Irapuato, Guanajuato, Mexico.
References
- 1.Sagan, L. (1967). On the origin of mitosing cells. J Theor Biol. 14: 255-274. [DOI] [PubMed]
- 2.Marin, B., Nowack, E.C.M, and Melkonian, M. (2005). A plastid in the making: evidence for a second primary endosymbiosis. Protist. 156: 425-432. [DOI] [PubMed]
- 3.Criscuolo, A., Gribaldo, S. (2011) Large-scale phylogenomic analyses indicate a deep origin of primary plastids within cyanobacteria. Mol Biol Evol. 28:3019-3032. [DOI] [PubMed]
- 4.Qiu, H., Yang, E.C., Bhattacharya, D., Yoon, H.S. (2012) Ancient Gene Paralogy May Mislead Inference of Plastid Phylogeny. Mol Biol Evol. 29:3333-3343. [DOI] [PubMed]
- 5.Reyes-Prieto, A., Yoon, H.S., Moustafa, A., Yang, E.C., Andersen, R.A., Boo, S.M., Nakayama, T., Ishida, K., Bhattacharya, D. (2010) Differential gene retention in plastids of common recent origin. Mol Biol Evol. 27:1530-1537. [DOI] [PMC free article] [PubMed]
- 6.Nowack, E.C.M., Melkonian, M. and Glöckner G. (2008) Chromatophore genome sequence of Paulinella sheds light on acquisition of photosynthesis by eukaryotes. Curr. Biol. 18: 410-418. [DOI] [PubMed]
- 7.Nowack, E.C., Grossman, A.R. (2012). Trafficking of protein into the recently established photosynthetic organelles of Paulinella chromatophora. Proc Natl Acad Sci U S A. 109: 5340–5345. doi: 10.1073/pnas.1118800109. [DOI] [PMC free article] [PubMed]
- 8.Gómez-Valero, L., Latorre, A., Silva, F.J. (2004). The evolutionary fate of nonfunctional DNA in the bacterial endosymbiont Buchnera aphidicola. Mol Biol Evol. 21:2172-2181. [DOI] [PubMed]
- 9.Bhattacharya, D., Price, D.C., Yoon, H.S., Yang, E.C., Poulton, N.J., Andersen, R.A., Das, S.P. (2012). Single cell genome analysis supports a link between phagotrophy and primary plastid endosymbiosis. Sci. Rep. 2:356. doi: 10.1038/srep00356. [DOI] [PMC free article] [PubMed]
- 10.Berney, C. and Pawlowski, J. (2006). A molecular time-scale for eukaryote evolution recalibrated with the continuous microfossil record. Proc. R. Soc. B. 273: 1867-1872. [DOI] [PMC free article] [PubMed]
- 11.Parfrey, L.W., Lahr, D.J., Knoll, A.H., Katz, L.A. (2011). Estimating the timing of early eukaryotic diversification with multigene molecular clocks. Proc. Natl. Acad. Sci. U S A. 108: 13624-13629. doi: 10.1073/pnas.1110633108. [DOI] [PMC free article] [PubMed]
- 12.Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., Peplies, J., Glöckner, F.O. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucl. Acids Res. 41 (D1): D590-D596. [DOI] [PMC free article] [PubMed]
- 13.Heger, T.J., Mitchell, E.A., Todorov, M., Golemansky, V., Lara, E., Leander, B.S., Pawlowski, J. (2010). Molecular phylogeny of euglyphid testate amoebae (Cercozoa: Euglyphida) suggests transitions between marine supralittoral and freshwater/terrestrial environments are infrequent. Mol Phylogenet Evol. 55: 113-122. [DOI] [PubMed]
- 14.Damsté, J.S., Muyzer, G., Abbas, B., Rampen, S.W., Massé, G., Allard, W.G., Belt, S.T., Robert, J.M., Rowland, S.J., Moldowan, J.M., Barbanti, S.M., Fago, F.J., Denisevich, P., Dahl, J., Trindade, L.A., Schouten, S. (2004). The rise of the rhizosolenid diatoms. Science. 304: 584-587. [DOI] [PubMed]
- 15.Edgar, R.C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32: 1792-1797. [DOI] [PMC free article] [PubMed]
- 16.Bouckaert, R., Heled, J., Kühnert, D., Vaughan, T., Wu, C-H, et al. (2014). BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput Biol 10(4): e1003537. doi:10.1371/journal.pcbi.1003537. [DOI] [PMC free article] [PubMed]
- 17.Darriba, D., Taboada, G.L., Doallo, R., Posada, D. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nature Methods. 9: 772. [DOI] [PMC free article] [PubMed]
- 18.Porter, S.M., Knoll, A.H. (2000). Testate amoebae in the Neoproterozoic Era: evidence from vase-shaped microfossils in the Chuar Group, Grand Canyon. Paleobiology. 26: 360-385.
- 19.Barber, A., Siver, P.A., Karis, W. (2013). Euglyphid testate amoebae (rhizaria: euglyphida) from an arctic eocene waterbody: evidence of evolutionary stasis in plate morphology for over 40 million years. Protist. 164: 541-555. [DOI] [PubMed]
- 20.Kooistra, W., Gersonde, R., Medlin, L., Mann, D.G. (2007). In The Evolution of Primary Producers in the Sea, The origin and evolution of the diatoms: Their adaptation to a planktonic existence, eds Falkowski PG, Knoll AH (Elsevier, Burlington, MA), pp 201–249.
- 21.Harwood, D.M., Nikolaev, V.A., Winter, D.M. (2007). Cretaceous records of diatom evolution, radiation, and expansion. Paleontol Soc Papers. 13:33–59.
- 22.Culver, S.J. (1991). Early Cambrian Foraminifera from West Africa. Science. 254:689-691. [DOI] [PubMed]
- 23.Groussin, M., Pawlowski, J., Yang, Z. (2011). Bayesian relaxed clock estimation of divergence times in foraminifera. Mol Phylogenet Evol. 61: 157-166. [DOI] [PubMed]
- 24.Nowack, E.C. (2014). Paulinella chromatophora – rethinking the transition from endosymbiont to organelle. Acta Soc Bot Pol. 83: 387-397.