

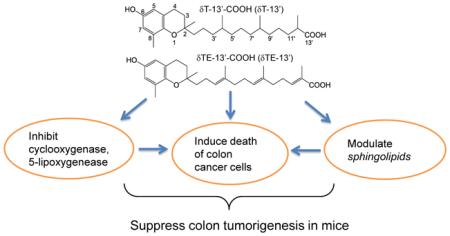
Abstract
Vitamin E forms are substantially metabolized to various carboxychromanols including 13’-carboxychromanols (13’-COOHs) that are found at high levels in feces. However, there is limited knowledge about functions of these metabolites. Here we studied δT-13’-COOH and δTE-13’-COOH, which are metabolites of δ-tocopherol and δ-tocotrienol, respectively. δTE-13’-COOH is also a natural constituent of a traditional medicine Garcinia Kola. Both 13’-COOHs are much stronger than tocopherols in inhibition of pro-inflammatory and cancer promoting cyclooxygenase-2 (COX-2) and 5-lipoxygnease (5-LOX), and in induction of apoptosis and autophagy in colon cancer cells. The anti-cancer effects by 13’-COOHs appeared to be partially independent of inhibition of COX-2/5-LOX. Using liquid chromatography tandem mass spectrometry, we found that 13’-COOHs increased intracellular dihydrosphingosin and dihydroceramides after short-time incubation in HCT-116 cells, and enhanced ceramides while decreased sphingomyelins during prolonged treatment. Modulation of sphingolipids by 13’-COOHs was observed prior to or coinciding with biochemical manifestation of cell death. Pharmaceutically blocking the increase of these sphingolipids partially counteracted 13’-COOH-induced cell death. Further, 13’-COOH inhibited dihydroceramide desaturase without affecting the protein expression. In agreement with these mechanistic findings, δTE-13’-COOH significantly suppressed the growth and multiplicity of colon tumor in mice. Our study demonstrates that 13’-COOHs have anti-inflammatory and anticancer activities, may contribute to in vivo anticancer effect of vitamin E forms and are promising novel cancer prevention agents.
Keywords: cancer, sphingolipid, vitamin E metabolites, autophagy, apoptosis
Graphical abstract
INTRODUCTION
Colorectal cancer is one of the leading causes of cancer death worldwide [1]. There is, however, no effective therapy for the late-stage cancer. Chemoprevention that prevents or delays the onset of late-stage cancer is an important strategy for reducing cancer-related mortality. To this end, specific forms of vitamin E may be potentially good candidates for chemoprevention. The vitamin E family consists of α-, β-, γ- and δ-tocopherol (αT, βT, γT and δT) and α-, β-, γ- and δ-tocotrienol (αTE, βTE, γTE and δTE). αT is the only vitamin E that has been tested in large intervention trials, but failed to show consistent beneficial effects in prevention of cancer in many clinical or preclinical studies [2]. On the other hand, we and others have documented that other vitamin E forms appear to be superior to αT in cancer prevention. For instance, γT, δT and γT/δT-rich mixed tocopherols but not αT inhibit development of colon cancer in preclinical animal model [3-7].
Despite these interesting findings, the anticancer effects of γT or δT may not solely be rooted in tocopherol themselves because these vitamin E forms are readily metabolized in the whole body environment. Except for αT, tocopherols and tocotrienols are substantially metabolized by cytochrome P450-catalyzed ω-hydroxylation and oxidation to generate 13’-carboxychromanols (13’-COOH), which are further catabolized to shorter chain carboxychromanols and sulfated carboxychromanols [3, 8, 9]. Carboxychromanols and their sulfated counterparts have been detected in the plasma and liver of rodents supplemented with γT and γTE [9, 10]. Recently, 13’-COOHs are found at high levels in feces from mice fed diet supplemented with γT, δT or mixed tocopherols [4, 11-13].
Emerging evidence suggests that long-chain carboxychroamols have bioactivities that are relevant to prevention and therapy of inflammation-associated diseases including cancer [3]. Specifically, we have demonstrated that δT-13’-COOH, a long-chain metabolite from δT, competitively inhibits cyclooxygenases (COX-1 and COX-2) and is much stronger than tocopherols or short-chain metabolites in these effects [14]. δT-13’-COOH, but not tocopherols, inhibits 5-lipoxygenase (5-LOX) [15]. It has been well recognized that COXs and 5-LOX are pro-inflammatory enzymes that play significant roles in inflammation and promote development of colon cancer [16, 17]. COX inhibitors have proven effective in decreasing colon cancer incidence in numerous clinical studies [16, 17]. Since high levels of long-chain carboxychromanols are found in feces, we reason that 13’-COOHs may play a role in prevention of colon cancer.
To test this hypothesis, we investigated anti-inflammatory and chemopreventive activities of long-chain carboxychromanols including δT-13’-COOH and δTE-13’-COOH (Figure 1A). δT-13’-COOH used in this study is the synthetic counterpart of δ-tocopherol-derived 13’-COOH [14, 15]. δTE-13’-COOH is a metabolite from δ-tocotrienol and also found in the African Garcinia Kola, a traditional medicine used to treat osteoarthritis in West Africa [18, 19]. In our studies, we examined the effect of 13’-COOHs on COX-2 and 5-LOX, their impact on the growth and survival of various colon cancer cells and the mechanisms underlying observed anticancer actions. To translate mechanistic findings to a whole body environment, we examined the anticancer efficacy of δTE-13’-COOH against colon tumorigenesis induced by AOM/DSS (azoxymethane/dextran sodium sulfate) in mice.
Figure 1. The structures of δT-13’-COOH and δTE-13’-COOH (A); The effect of 13’-COOHs and tocopherols on cell viability of human colon HCT-116 (B) and HT-29 (C) cancer cells; The relative anti-proliferation effect (24-h incubation) of 13’-COOHs on cancer cells vs. human normal CCD841CoN (CCD) colon epithelial cells (D).

Relative cell viability was measured by MTT assay after cells were treated with 13’-COOHs or tocopherols at indicated concentrations and times compared with DMSO controls. IC50s are the concentrations that caused 50% decrease in cell viability after 24-h incubation. The data are mean SD from at least three independent experiments. *p < 0.05 and **p < 0.01 indicate a significant difference between treated and DMSO-control cells. δT-13’-COOH and δTE-13’-COOH are abbreviated as δT-13’ and δTE-13’, respectively.
MATERIALS AND METHODS
Materials and reagents
δT-13’-COOH and δTE-13’-COOH (>95% pure) were synthesized as previously described [20]. δTE-13’-COOH (> 92%) used in animal studies was isolated from the Garcinia Kola seeds based on a published procedure [18]. γT (≥96%) and δT (93-97%) were purchased from Sigma (St Louis, MO) and Suppleco (Bellefonte, PA). Arachidonic acid (AA), human recombinant COX-2 and 5-LOX were from Cayman Chemical (Ann Arbor, MI). C8-cyclopropenylceramide (C8-CPPC) was purchased from Matreya LLC (Pleasant Gap, PA). All sphingolipid standards were obtained from Avanti Polar Lipids (Alabaster, AL). Myriocin from Mycelia Sterilia, dimethyl sulfoxide (DMSO), [3-(4,5)-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide] (MTT) and all other chemicals were from Sigma.
Effects on COX-2 and 5-LOX activity
COX-2 enzymatic reactions were performed in 0.1 M Tris (pH 8.0) in the presence of 5 mM EDTA, 2 mM phenol and 1 µM hematin. After preincubation of 13’-COOHs with human COX-2, AA at a final concentration of 5 µM was added for 2 min and the reaction was stopped by addition of 0.1 M HCl. Stannous chloride in 0.1 M HCl was added to reduce PGG2 and PGH2 to PGF2α, which was quantified using ELISA assays [14]. Potential effects on the activity of 5-LOX were evaluated using the ferrous oxidation-xylenol orange assay (FOX assay) as previously described [15].
Cell culture and MTT assay
Human colon cancer (HCT-116, HT-29, Caco-2) and normal epithelial (CCD841CoN) cells were obtained from American Type Culture Collection (Manassas, VA) and routinely cultured in growth media containing 10% fetal bovine serum (FBS) at 37 °C in 5% CO2. HT-29 cells and HCT-116 cells were cultured in Dulbecco’s modified eagle medium (DMEM) and McCoy’s 5A modified medium, respectively. Caco-2 cells were grown in DMEM supplemented with 1% non-essential amino acids (GIBCO, Grand Island, NY), and CCD841CoN cells were cultured in Eagle’s minimum essential medium. Cell viability was examined by the MTT assay for estimating mitochondrial dehydrogenase activity as previously described [21].
Detection of apoptosis with Annexin V and Propidium Iodide Staining
Both floating and attached cells were collected and stained with Annexin-V-Flous staining kit (Roche Applied Science, Indianapolis, IN). Apoptosis (Annexin V: Ex = 488 nm; Em = 518 nm) and necrosis (Propidium Iodide: Ex = 488-540 nm; Em = 617 nm) were evaluated by Beckman Coulter FC500 (Beckman Coulter, Miami, FL) or BD FACS Aria III cell sorter (BD Biosciences, San Jose, CA) with FlowJo software system.
Western Blotting
Cells were lysed in a buffer containing Tris-EDTA, 1% SDS, 2 mM Na3VO4 and protease inhibitor cocktails (Sigma). Total proteins were quantified by bicinchoninic acid (BCA) protein assay kit (Pierce, Rockford, IL). Equal amount of proteins (15-30 g) were separated on acrylamide gels by SDS-electrophoresis and probed by antibodies as described [22]. The antibodies used in the study were as follows: membrane bound microtubule-associated protein light chain 3 (LC3; MBL international, Woburn, MA), Caspase-9, poly (ADP-ribose) polymerase-1 (PARP-1) and Actin (Santa Cruz Biotechnology, Santa Cruz, CA), and DEGS-1 (Novus Biologicals, Littleton, CO).
Lipid extraction
Lipid was extracted as previously described [23]. Briefly, cell pellets were resuspended in 500 µL of methanol, 250 µL of chloroform and 50 µL of water after the addition of 20 µL of internal standard mixture containing 25 µM of C12:0-ceramide, C25:0-ceramide, C17-sphingosine, C17-sphinganine, and C12:0-sphingomyelin (Avanti Polar Lipids). The suspension was dispersed fully by tip sonication for 20 sec and then incubated overnight at 48 °C. 100 µL of solution was used to determine the amount of total choline-containing phospholipids (PCs) by an enzymatic colorimetric assay (Wako chemicals, Osaka, Japan) [21]. 75 µL of 1 M potassium hydroxide in methanol was added to the rest of the solution and sonicated for 30 min. Samples were then incubated at 37 °C for 2 h and evaporated under N2.
Measurement of sphingolipids using liquid chromatography tandem mass spectrometry (LC-MS/MS)
The effect of 13’-COOHs on sphingolipids was evaluated by LC-MS/MS as previously described [22, 23]. Briefly, the analyses were performed using the Agilent 6460 triple quadrupole mass spectrometer coupled with the Agilent 1200 Rapid Resolution HPLC (Agilent Technologies, Santa Clara, CA). Sphingolipids were detected in positive mode by multiple reaction monitoring (MRM) technique [23]. Cers and sphingoid bases were separated on an Agilent XDB-C18 column (1.8 m, 4.6 χ 50 mm), respectively, by an isocratic (100% B - methanol-formic acid 99:1, v/v) or gradient run at 20% B for 0-1 min, 100% B for 10-13 min and 20% B for 15-20 min with solvent A containing methanol-H2O-formic acid (74:25:1, v/v/v). SMs were separated on an Agilent Zorbax XDB-C8 (3.5 m, 2.1 χ 50 mm) with gradient of 20% B for 0-1 min, 100% B for 10-20 min, and 20% B for 22-30 min. The MS/MS parameters were as follows: gas temperature at 325-350 °C, gas flow rate at 7-10 L/min, nebulizer pressure at 45-50 psim capillary voltage at 3500 V, the fragmentor voltage at 100 V and collision energy at 12-20 V. Precursor-to-product ion transitions for each sphingolipid were used according to the method by Merrill et al. [23].
Dihydroceramide desaturase (DEGS) assay
For the in vitro assay of DEGS, HCT-116 cells were treated with DMSO (controls), δT-13’-COOH (20 μM) or C8-CPPC (1 μM) for 1 or 2 h. Cells were collected and homogenized in a buffer (5 mM Hepes, pH 7.4, containing 50 mM sucrose) and kept on ice for 10 min. The cell homogenate was centrifuged at 250 χ g for 5 min at 4 °C to remove unbroken cells. In some studies, microsomal fraction from rats’ liver were isolated and used as previously described [24]. In cell homogenates, reactions were initiated by addition of NADH (2 mM) and 10 μM of C8:0-dhCer as a non-physiological substrate for DEGS for 1 h at 37 °C. Lipids were extracted and the products including C8:0-Cer and C8:0-SM were quantified by LC-MS/MS.
Animal Study
The animal use protocol was approved by the Animal Care and Use Committee at Purdue University. δTE-13’-COOH (0.022% in diet) was prepared in the AIN93G-based diet (Dyets Inc, Bethlehem, PA). This dose is equivalent to ~200 mg daily intake for a 70 kg person and represents a medium supplement dose. After one week adaptation, male Balb/c mice (5-6 weeks old from Harlan, Indianapolis, IN) were i.p. injected with azoxymethane (AOM; Sigma) at 9.5 mg/kg body weight. A week later, AOM-injected mice were randomized into AIN-93G (control) or δTE-13’-COOH-supplemented group. Meanwhile, mice were given 1.5% dextran sodium sulfate (DSS; molecular weight of 36,000-50,000, MP Biochemicals, Solon, OH) in drinking water for 1 week. The DSS cycle was repeated after a two-week interval (design outlined in Figure 6A). Animals were observed and weighed daily and food intake was measured once a week. During tissue harvest, colons were removed, rinsed with cold saline, cut open longitudinally from rectum to cecum, and tumors were examined and counted. Colon was fixed flat in 4% formaldehyde at 4C overnight for histologic evaluation.
Figure 6. δTE-13’-COOH significantly suppressed colon tumorigenesis induced by AOM with two cycles of DSS (1.5%) in Balb/c mice.
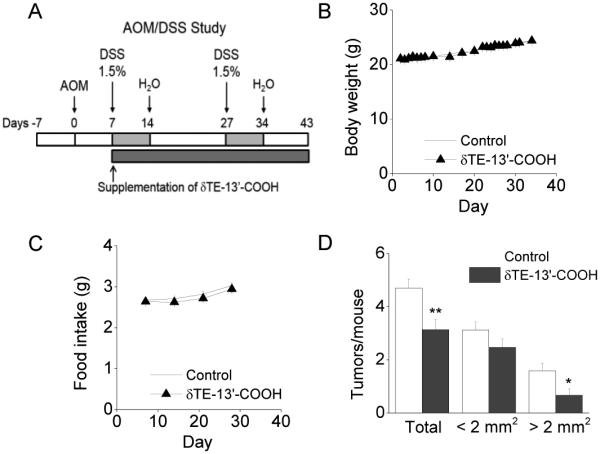
A: The study design. B and C: Body weight and food intake of each group during the days after supplementation started. D: The effect of δTE-13’-COOH on colon tumor multiplicity and polyps with sizes of < 2 mm2 or > 2 mm2 in the AOM/DSS study (mean SEM, n = 15-17). *p < 0.05 and **p < 0.01 differences between the AIN-93G control and δTE-13’-COOH supplement group.
Statistics
The Student’s t-test was used to compare two groups. P < 0.05 was considered statistically significant.
RESULTS
The effect of 13’-COOHs on the COX-2 and 5-LOX activity
We have previously shown that δT-13’COOH, which was isolated from conditioned media from cells incubated with δT, is an inhibitor of COX-1/COX-2 and 5-LOX [14, 15]. Here we found that chemically synthesized δT-13’-COOH showed similar inhibitory potency toward COX-2 and 5-LOX compared to that generated from δT metabolism in cells [14, 15] (Table 1). δTE-13’-COOH, an analog of δT-13’-COOH with three double bonds in the side chain (Figure 1A), appeared to be slightly stronger than δT-13’-COOH in inhibition of 5-LOX, but less potent in inhibition of COX-2 (Table 1). Like our previous observations [14, 15], neither γT nor δT at 50 μM inhibited COX-2 or 5-LOX in these enzyme assays.
TABLE 1.
Inhibitory effects of 13’-COOHs and tocopherols on COX-2 and 5-LOX
| IC50 (μM) | δT-13’-COOH | δTE-13’-COOH | γT | δT |
|---|---|---|---|---|
| COX-2 | 4 ± 1 | 9.8 ± 2 | > 50 | > 50 |
| 5-LOX | 2 ± 0.5 | 1 ± 0.5 | > 50 | > 50 |
The effect of 13’-COOHs or tocopherols on COX-2 and 5-LOX were evaluated with purified human enzymes as described in Materials and Methods. Results are mean ± SEM based on four or more independent experiments.
13’-COOHs inhibited proliferation of human colon cancer cells
Birringer et al. [25] showed that 13’-COOHs derived from γT or δT induced apoptosis in human HepG2 cells, but it is not clear whether these compounds have any impact on other types of cancer cells. Here we investigated the effect of 13’-COOHs on proliferation of colon cancer cells using MTT assays. δT-13’-COOH and δTE-13’-COOH decreased the viability of HCT-116 and HT-29 cells in a time- and dose-dependent manner with indicated IC50 values (estimated after 24 h incubation) (Figure 1B and 1C). In these activities, δT-13’-COOH was more potent than δTE-13’-COOH, and both 13’-COOHs were much stronger than tocopherols. Importantly, normal human colon epithelial (CCD) cells appeared to be less sensitive to 13’-COOHs than cancer cells (Figure 1D).
13’-COOHs induced apoptosis and autophagy in cancer cells
Like dual COX and 5-LOX inhibitor Licofelon [26], both δT-13’-COOH and δTE-13’-COOH induced early and late-stage apoptosis in HCT-116 cells compared with DMSO-treated controls, as indicate by enhanced annexin V staining that is associated with externalization of phosphatidylserine to the cytoplasmic membrane (Figure 2A). Consistently, 13’-COOHs caused PARP cleavage and caspase-9 activation (Figure 2B). In addition to apoptosis, δT- and δTE-13’-COOH treatment led to an increase of LC3-II, a marker of autophagy (Figure 2C). Similar to the results from MTT assays, δT-13’-COOH appeared to be stronger than δTE-13’-COOH in the induction of apoptosis and autophagy.
Figure 2. 13’-COOHs induced apoptosis (A, B) and autophagy (C) in HCT116 cells and AA partially reversed anti-proliferative effects of 13’-COOHs in Caco-2 and HCT-116 cells (D).

HCT-116 cells were treated with δT-13’-COOH or δTE-13’-COOH at 20 μM for 24 h. Apoptosis and necrosis were evaluated by annexin V and PI staining using flow cytometry (A). Western blots were conducted to monitor apoptosis and autophagy as indicated by caspase-9 activation and PARP cleavage (B) and LC3-II (C), respectively. Results of flow cytometry and Western blots are representative of three or more independent experiments. Caco-2 or HCT116 cells were treated with δT-13’-COOH or δTE-13’-COOH with or without 1 μM of AA for 72 h and relative cell viability was measured by MTT assays (D). The data are mean SEM for at least three independent experiments. *p < 0.05 and **p < 0.01 indicate a significant difference between cells treated with 13’-COOHs in the presence vs. absence of AA. δT-13’-COOH and δTE-13’-COOH are abbreviated as δT-13’ and δTE-13’, respectively.
Arachidonic acid (AA) partially reversed anti-cancer effect of 13’-COOHs
Eicosanoids generated from AA via COX- or 5-LOX-catalyzed reactions are known to promote colon cancer development [16, 17]. To evaluate whether 13’-COOH-induced anticancer effect is rooted in inhibition of COX or 5-LOX, we examined potential impact of AA on 13’-COOH’s anti-proliferative effects. AA appeared to stimulate the growth of Caco-2 cells that are known to express COX-2 [27], but only slightly promoted the growth of HCT-116 cells that are COX-2 negative [28] (Figure 2D). We observed that AA partially counteracted 13’-COOH’s anti-proliferative effects in the MTT assays, suggesting that the anticancer effect may in part stem from inhibition of COX-2 and 5-LOX. On the other hand, the fact that 13’-COOHs decreased the viability of COX-2 negative cells indicates that additional mechanisms independent of suppression of COX or 5-LOX are likely involved in 13’-COOHs’ anticancer action.
13’-COOHs modulated sphingolipids in a dose- and time-dependent manner
We have previously demonstrated that modulation of sphingolipid metabolism plays a role in γT and γTE-induced death in human prostate and breast cancer cells [21, 29, 30] or in their inhibition of NF-κB in macrophages [22]. Here we investigated potential effects of 13’-COOHs on sphingolipids using LC-MS/MS. After 1-2 h treatment, δT-13’-COOH dose-dependently increased total dihydroceramides (dhCers), individual dhCers including C16:0-, C18:0-, C24:1- and C24:0-dhCers and dihydrosphingosine (dhSph) compared with controls, and the elevation of these sphingolipids accentuated during prolonged incubation (Figure 3A-C) (Table S1). In contrast to dhCer that was enhanced after 2 h incubation, the effect of 13’-COOHs on ceramides (Cer) varied with treatment time, concentrations and individual Cer species. Specifically, compared with solvent controls, δT-13’-COOH at 20 μM induced a significant decrease of C16:0-Cer after 2 h incubation, but enhanced this sphingolipid during longer treatment (8 h or 16 h incubation) and increased C18:0-Cer after 4 h treatment (Figure 3D). δT-13’-COOH at 10 and 20 μM caused opposite effects on C24-Cers (Table S1). For sphingomyelin (SM), δT-13’-COOH led to persistent decrease in all types of SM species and total SMs (Figure 3E) (Table S1). Furthermore, δTE-13’-COOH induced similar modulation of sphingolipids to that by δT-13’-COOH (Table S2). In summary, 13’-COOH-treated cells accumulated higher levels of dhCers, dhSph, total Cers but lower SMs than controls after 8-16 h incubation when autophagy and apoptosis were observed.
Figure 3. δT-13’-COOH (δT-13’) profoundly modulated sphingolipid metabolism in HCT-116 cells in time- and dose-dependent manners.

HCT-116 cells were treated with 10 or 20 μM of δT-13’-COOH for 1, 2, 4, 8, or 16 h. LC-MS/MS was used to measure the sphingolipid levels including total dhCers (A), C16:0-dhCer (B), dhSph (C), total Cers, C16:0-Cer, C18:0-Cer (D) and total SMs (E). Results are shown as mean SEM for at least three independent experiments. *p < 0.05 and **p < 0.01 indicate a significant difference between treated and DMSO control cells at each indicated time.
13’-COOHs inhibited DEGS1 activity without affecting its protein expression
Since δT-13’-COOH increased dhCers but decreased Cers after its 2-h incubation with cells, we reason that δT-13’-COOH likely inhibits the activity or protein expression of DEGS1, which is responsible for converting dhCer to Cer (Figure 4A). The Western blotting data revealed that δT-13’-COOH had no effect on DEGS-1 protein expression in HCT-116 cells (Figure 4B). In an in vitro assay with cell homogenates, δT-13’-COOH, after 1 and 2 h pre-incubation, showed ~38% and ~47% inhibition of DEGS1 enzyme activity, respectively. C8-CPPC, a known competitive inhibitor of DEGS1 [31], inhibited the enzyme activity by ~95% (Figure 4C). On the other hand, despite inhibition of DEGS1 in the cell homogenate, δT-13’-COOH failed to inhibit the enzyme activity when rat liver microsomal fraction was used as the source of DEGS (data not shown).
Figure 4. δT-13’-COOH (δT-13’) inhibited DEGS activity but not protein expression.

A: The de novo biosynthesis pathway of sphingolipids (SMS: sphingomyelin synthase; SMase: sphingomyelinase). B: HCT-116 cells were treated with δT-13’-COOH at 10 or 20 μM for 8 or 16 h and the effect on DEGS-1 expression was detected by Western blotting. C: HCT-116 cells were treated with DMSO (control), δT-13’-COOH at 20 μM or C8-CPPC at 1 μM for 1 or 2 h. Cells were collected and homogenized. The homogenates were added with NADH and C8:0-dhCer as a substrate for DEGS. After 1 h incubation at 37 C, the production of C8:0-sphingolipids were analyzed by LC-MS/MS. The data are mean SEM of three independent experiments. *p < 0.05 and **p < 0.01 indicate significant differences between DMSO-control cells and 13’-COOH or CPPC-treated cells.
The role of sphingolipid modulation in 13’-COOH-induced cell death
Because 13’-COOHs induced accumulation of intracellular dhCer and dhSph, which are known to induce cell stress and/or death [32], we used myriocin, a specific inhibitor of serine palmitoyltransferase (Fig 4A), to block the increase of these sphingoid bases [22]. Interestingly, co-treatment of cells with myriocin resulted in partial counteraction of 13’-COOH-induced LC3-II expression (Figure 5A), but had no effect on PARP-1 cleavage (data not shown). These data suggest that elevation of dhCer and dhSph may play a role in 13’-COOHs-induced autophagy.
Figure 5. Myriocin and GW48695 partially counteracted 13’-COOHs-induced anticancer activities.
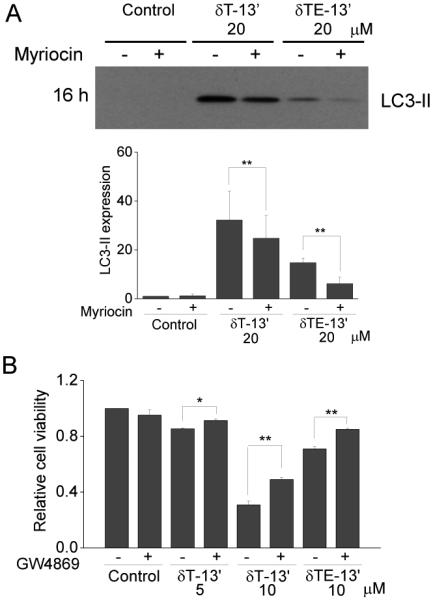
A: HCT-116 cells were treated with 13’-COOH at 20 μM with or without myriocin (3 μM), a specific inhibitor of serine palmitoyltransferase to block the de novo sphingolipid synthesis. After 16-h treatment, cells were collected and analyzed for LC3-II by Western blotting. The data are mean SEM of more than 4 independent experiments. *p < 0.05 and **p < 0.01 indicate a significant difference between 13’-COOHs in the presence vs. absence of myriocin. B: HCT-116 cells were treated with δT-13’-COOH (5 or 10 μM) or δTE-13’-COOH (10 μM) with or without GW48695 (5 μM, an inhibitor of neutral SMase) for 24 h. Relative cell viability was measured by MTT assays. The data are mean SEM of three independent experiments. *p < 0.05 and **p < 0.01 indicate a significant difference between 13’-COOHs with vs. without GW48695. δT-13’-COOH and δTE-13’-COOH are abbreviated as δT-13’ and δTE-13’, respectively.
Ceramides are known to be potent inducers of apoptosis [32]. The inhibition of DEGS by 13’-COOHs is consistent with decreased C16:0-Cer after 2 h incubation (Figure 3D). However, Cers were elevated after 4 h or longer treatment (Figure 3D), which, we reason, may be caused by enhanced hydrolysis of SM via sphingomyelinases (SMases). To establish the role of ceramides in 13’-COOH-induced cancer cell death, we used desipramine and GW4869 to inhibit acid or neutral SMases, respectively. Co-treatment of GW4869 but not desipramine (data not shown) partially reversed 13’-COOH-caused anti-proliferation (Figure 5B). These data suggest that SM hydrolysis through the neutral SMase activation may in part contribute to 13’-COOHs-induced anticancer effects.
δTE-13’-COOH inhibited colon tumorigenesis in a preclinical model in mice
To translate the mechanistic findings to a whole body environment, we investigated the effectiveness of δTE-13’-COOH against colon tumorigenesis induced by AOM and DSS in male Balb/c mice, as designed in Figure 6A. This preclinical model is commonly used for testing chemoprevention agents and believed to resemble inflammation-promoted colon carcinogenesis. Supplementation of δTE-13’-COOH at 0.022% in diet had no effect on body weight or food intake compared with the control diet (Figure 6B and C), indicating lack of obvious adverse effects. As observed in our previous studies in this model [4], AOM/DSS treatment resulted in formation of adenomatous polyps in the middle to the rectal part of the distal colon. Mice fed control diet had 4.7 ± 0.3 tumors per mouse, and δTE-13’-COOH supplemented group had significantly lowered colon tumors (3.1 ± 0.3; p < 0.01; Fig. 6D). More importantly, when the tumors were categorized as small (< 2 mm2) and large sizes, supplementation of δTE-13’-COOH significantly decreased the number of large tumors (> 2 mm2) by 58% (p < 0.05), compared with controls (Fig. 6D). These results suggest that δTE-13’-COOH likely suppressed tumor promotion in addition to decreasing tumor multiplicity.
DISCUSSION
We demonstrate that vitamin E long-chain metabolites δT-13’-COOH and δTE-13’-COOH are dual inhibitors of COX-2 and 5-LOX and decrease viability of human colon cancer cells by induction of apoptosis and autophagy. For the anti-proliferative effect, δT-13’-COOH appears to be more potent than δTE-13’-COOH, and both 13’-COOHs are much stronger than tocopherols. Using a lipidomic approach with LC-MS/MS, we show for the first time that 13’-COOHs profoundly modulate sphingolipid metabolism. Specifically, 13’-COOH treatment quickly increased dhCers and dhSph, and subsequently enhanced Cers including C16- and C18-Cer while decreased SMs. The importance of sphingolipid modulation in 13’-COOH-induced anticancer effects is supported by the fact that dhCers, dhSph and Cers, which are known to induce antiproliferation, cell stress and death in different types of cancer cells [32], are elevated by 13’-COOHs prior to or coinciding with induction of apoptosis or autophagy. Further, chemically blocking the increase of these sphingolipids by myriocin or a neutral SMase inhibitor partially reversed 13’-COOH-mediated cell death. Consistent with these mechanistic findings, δTE-13’-COOH significantly suppressed colon tumorigenesis induced by AOM/DSS in mice.
Eicosanoids generated by COX-2- or 5-LOX-catalyzed reactions are known to accelerate cancer development by promoting inflammation in tumor micro-environments and by fostering proliferation and resistance to apoptosis of cancer cells [16, 17]. Inhibition of COXs is a well-recognized chemopreventive strategy and has especially proven effective against colorectal cancer [16, 17]. Dual inhibition of COX and 5-LOX is deemed to be better than blocking either pathway alone because inhibition of both pathways may not only deliver stronger anti-inflammatory and anticancer effects but also potentially reduce adverse effects associated with COX inhibitors [3, 33]. Like our previous observation with naturally generated δT-13’-COOH [14, 15], its synthetic counterpart tested in the present study similarly inhibits COX-2 and 5-LOX. δTE-13’-COOH, which has three double bonds in the side chain and is a component in Garcinia Kola, showed stronger inhibitory potency of 5-LOX but weaker inhibition of COX-2 than δT-13’-COOH. We have previously shown that δT-13’-COOH is a competitive inhibitor of COXs [14], while the nature underlying 13’-COOHs’ inhibition of 5-LOX is currently under investigation. Like licofelone (a COX/5-LOX inhibitor) [26], 13’-COOHs are capable of induction of apoptosis in colon cancer cells. Further, we observe that 13’-COOHs decreased cancer cell viability in COX-2-positive and negative cells and that arachidonic acid only partially counteracted 13’-COOH’s anticancer effect. We therefore conclude that COX- and/or 5-LOX independent mechanism(s) is likely involved in their anticancer effects observed in cancer cells.
Based on the temporal modulation of sphingolipids in HCT-116 cells, we identify DEGS as another inhibitory target by 13’-COOHs. In particular, 13’-COOHs caused rapid increase of dhCers but decrease of Cers including C16:0-Cer during short 2-h incubation, indicating that DEGS-catalyzed conversion of dhCers to Cers is compromised. The similar pattern of sphingolipid changes was reported in DEGS knockout model [34, 35]. Consistently, δT-13’-COOH inhibited DEGS enzyme activity in an in vitro assay despite having no impact on its protein expression. DEGS was first proposed as a potential target for vitamin E γT that induced apoptosis of prostate cancer cells via causing intracellular accumulation of dhCers and dhSph without changing total ceramides [21]. Similar modulatory effects on sphingolipids were subsequently observed by other anticancer agents including vitamin E γ-tocotrienol [29, 30], resveratrol [36] and fenretinide (or 4-HPR) [24]. Interestingly, like 13’-COOHs, celecoxib (a selective COX-2 inhibitor) has recently been demonstrated to inhibit the DEGS activity and cause similar sphingolipid alterations in colon cancer cells [28]. It is noteworthy that downregulation of DEGS has been shown to cause profound changes of cell signaling, lipid homeostasis and gene expression [34, 35], which may contribute to the treatment effects (e.g. anti-proliferation) on cancer cells by these compounds.
Besides initial inhibition of DEGS, other enzymes in sphingolipid metabolism are likely affected by 13’-COOHs because these compounds enhanced ceramides esp. C16:0- and C18:0-Cer during prolonged treatment. Cellular levels of Cers and SMs are determined by the balance between de novo synthesis and SM hydrolysis by SMases (Figure 4A). Cers are de novo synthesized in the ER and then converted to SMs in the Golgi. SMs can be hydrolyzed to regenerate Cers by acid or neutral SMases [37]. Hydrolysis of SM by either enzymes is known to be activated by TNFα, oxidants, UV radiation [37-41], vitamin D [42] and curcumin [43]. Since 13’-COOHs inhibited DEGS and consequently decreased de novo synthesis of Cers, the increase of Cers in the prolonged treatment was likely caused by SMase-mediated SM hydrolysis, which resulted in further decrease of SMs. Consistently, we found that an inhibitor of neutral but not acid SMase significantly counteracted 13’-COOHs-induced cancer cell death, suggesting that neutral SMase-catalyzed SM hydrolysis may be enhanced during prolonged 13’-COOH treatment. In addition, we observe that 13’-COOHs appeared to have distinct effect on different Cers, e.g. increasing C16:0- and C18:0-species while decreasing C24:0-Cer. This observation suggests that 13’-COOHs may have differential impacts on ceramide synthases (CerS). Further investigation should be conducted to characterize potential effect of 13’-COOH on SMases and Cer synthases.
In agreement with mechanistic findings, δTE-13’-COOH significantly suppressed tumor development in the AOM/DSS-induced colon cancer in mice. The anticancer effect likely stems from multiple factors as observed in mechanistic studies including inhibition of COX-2/5-LOX and anti-proliferative activities. Similarly, an established COX/5-LOX inhibitor licofelone has recently been shown to inhibit tumor development in another preclinical colon cancer model [44]. Previous studies have demonstrated that γT and mixed tocopherols are effective in suppression of colon tumorigenesis in the AOM- or AOM/DSS-induced cancer model [3-7]. Our present data indicate that δTE-13’-COOH at 0.022%, a much lower dose than γT (0.1%) or mixed tocopherols (0.17 or 0.3%), appeared to be more effective than tocopherols in blocking tumorigenesis. This is consistent with the mechanistic observations that 13’-COOHs are much stronger than γT or δT in inhibition of COX/5-LOX and in induction of cancer cell death. Interestingly, 13’-COOHs have been detected as a major fecal-excretion vitamin E metabolite at relatively high levels in fecal samples from mice fed γT or δT supplementation [4, 11-13]. Therefore, 13’-COOHs likely contribute to the anticancer effects of tocopherol supplements against colon cancer in vivo.
In summary, we have demonstrated that δT-13’-COOHs and δTE-13’-COOH are effective dual inhibitors of COX-2 and 5-LOX, and induce apoptosis and autophagy in colon cancer cells by modulating sphingolipids through targeting DEGS and possibly activating SM hydrolysis. We have, for the first time, shown that δTE-13’-COOH , a metabolite from δ-tocotrienol and an active component of Garcinia Kola, significantly suppressed colon cancer development in a preclinical cancer model. Our study indicates that 13’-COOHs may contribute to the chemopreventive effect by vitamin E forms and δTE-13’-COOH may play a role in treatment effects of Garcinia Kola. These long-chain carboxychromanols may be novel preventative and therapeutic agents against inflammation and cancer.
Supplementary Material
Highlights.
13’-Carboxychromanols are dual inhibitors of cyclooxygenase and 5-lipoxygenase
13’-Carboxychromanols induced cancer cell death via modulation of sphingolipids.
δTE-13’-carboxychromanol, a δ-tocotrienol derivative and found in Garcinia Kola, suppressed colon tumorigenesis in mice
ACKNOWLEDGEMENTS
The authors would like to thank Amber S Jannasch for helps with LC-MS/MS analyses of sphingolipids. This study was in part supported by grants R21CA152588 and R01AT006882 (to QJ) from National Institutes of Health. This project was also partially supported by NIH grant P30CA023168 via Purdue Center for Cancer Research.
ABBREVIATIONS
- 13’-COOH
13’-carboxychromanol
- αT, γT, or δT
α, γ, or δ-tocopherol
- γTE or δTE
γ, δ-tocotrienol
- δT-13’-COOH or δT-13’
13’-COOH derived from δT
- δTE-13’-COOH or δTE-13’
13’-COOH derived from δTE
- COX
cyclooxygenase
- 5-LOX
5-lipoxygenase
- Cer
ceramide
- dhCer
dihydroceramide
- Sph
sphingosine
- dhSph
dihydrosphingosine
- SM
sphingomyelin
- LC3
microtubule-associated protein light chain 3
- LC-MS/MS
liquid chromatography tandem mass spectrometry
- DEGS
dihydroceramide desaturase
- SMS
sphingomyelin synthase
- SMase
sphingomyelinase
- AOM
azoxymethane
- DSS
Dextran Sodium Sulfate
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
CONFLICT OF INTEREST: There is no conflict of interest in the study.
REFERENCES
- [1].Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. CA Cancer J Clin. United States: 2011. Global cancer statistics; pp. 69–90. [DOI] [PubMed] [Google Scholar]
- [2].Moya-Camarena SY, Jiang Q. The role of vitamin E forms in cancer prevention and therapy - Studies in human intervention trials and animal models. In: Sarkar FHE, editor. Nutraceuticals and Cancer. Springer Science Business Media B.V.; 2011. pp. 323–354. http://dx.doi.org/310.1007/1978-1094-1007-2630-1007_1015. [Google Scholar]
- [3].Jiang Q. Natural forms of vitamin E: metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic Biol Med. 2014;72:76–90. doi: 10.1016/j.freeradbiomed.2014.03.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Jiang Q, Jiang Z, Hall YJ, Jang Y, Snyder PW, Bain C, Huang J, Jannasch A, Cooper B, Wang Y, Moreland M. Gamma-tocopherol attenuates moderate but not severe colitis and suppresses moderate colitis-promoted colon tumorigenesis in mice. Free Radic Biol Med. 2013;65:1069–1077. doi: 10.1016/j.freeradbiomed.2013.08.187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [5].Guan F, Li G, Liu AB, Lee MJ, Yang Z, Chen YK, Lin Y, Shih W, Yang CS. delta- and gamma-tocopherols, but not alpha-tocopherol, inhibit colon carcinogenesis in azoxymethane-treated F344 rats. Cancer Prev Res (Phila) 2012;5:644–654. doi: 10.1158/1940-6207.CAPR-11-0521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Ju J, Hao X, Lee MJ, Lambert JD, Lu G, Xiao H, Newmark HL, Yang CS. A gamma-tocopherol-rich mixture of tocopherols inhibits colon inflammation and carcinogenesis in azoxymethane and dextran sulfate sodium-treated mice. Cancer Prev Res (Phila) 2009;2:143–152. doi: 10.1158/1940-6207.CAPR-08-0099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Newmark HL, Huang MT, Reddy BS. Mixed tocopherols inhibit azoxymethane-induced aberrant crypt foci in rats. Nutr Cancer. 2006;56:82–85. doi: 10.1207/s15327914nc5601_11. [DOI] [PubMed] [Google Scholar]
- [8].Sontag TJ, Parker RS. Cytochrome P450 omega-hydroxylase pathway of tocopherol catabolism. Novel mechanism of regulation of vitamin E status. J Biol Chem. 2002;277:25290–25296. doi: 10.1074/jbc.M201466200. [DOI] [PubMed] [Google Scholar]
- [9].Jiang Q, Freiser H, Wood KV, Yin X. Identification and quantitation of novel vitamin E metabolites, sulfated long-chain carboxychromanols, in human A549 cells and in rats. J Lipid Res. 2007;48:1221–1230. doi: 10.1194/jlr.D700001-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].Freiser H, Jiang Q. Gamma-tocotrienol and gamma-tocopherol are primarily metabolized to conjugated 2-(beta-carboxyethyl)-6-hydroxy-2,7,8-trimethylchroman and sulfated long-chain carboxychromanols in rats. J Nutr. 2009;139:884–889. doi: 10.3945/jn.108.103309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Bardowell SA, Ding X, Parker RS. Disruption of P450-mediated vitamin E hydroxylase activities alters vitamin E status in tocopherol supplemented mice and reveals extra-hepatic vitamin E metabolism. J Lipid Res. 2012;53:2667–2676. doi: 10.1194/jlr.M030734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Bardowell SA, Duan F, Manor D, Swanson JE, Parker RS. Disruption of mouse cytochrome p450 4f14 (Cyp4f14 gene) causes severe perturbations in vitamin E metabolism. J Biol Chem. 2012;287:26077–26086. doi: 10.1074/jbc.M112.373597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [13].Jiang Q, Xu T, Huang J, Jannasch AS, Cooper B, Yang C. Analysis of vitamin E metabolites including carboxychromanols and sulfated derivatives using liquid chromatography tandem mass spectrometry. J Lipid Res. 2015;56:2217–2225. doi: 10.1194/jlr.D061663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Jiang Q, Yin X, Lill MA, Danielson ML, Freiser H, Huang J. Long-chain carboxychromanols, metabolites of vitamin E, are potent inhibitors of cyclooxygenases. Proc Natl Acad Sci U S A. 2008;105:20464–20469. doi: 10.1073/pnas.0810962106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Jiang Z, Yin X, Jiang Q. Natural forms of vitamin E and 13'-carboxychromanol, a long-chain vitamin E metabolite, inhibit leukotriene generation from stimulated neutrophils by blocking calcium influx and suppressing 5-lipoxygenase activity, respectively. J Immunol. 2011;186:1173–1179. doi: 10.4049/jimmunol.1002342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Wang D, Dubois RN. Eicosanoids and cancer. Nat Rev Cancer. 2010;10:181–193. doi: 10.1038/nrc2809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Wang D, Dubois RN. The role of COX-2 in intestinal inflammation and colorectal cancer. Oncogene. 2010;29:781–788. doi: 10.1038/onc.2009.421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Terashima K, Shimamura T, Tanabayashi M, Aqui l. M., Akinniyi J, Niwa M. Constituents of the seeds of Garcinia kola: Two new antioxidants, Garcinoic acid and Garcinal. Heterocycles. 1997;45:1559–1566. [Google Scholar]
- [19].Adegbehingbe OO, Adesanya SA, Idowu TO, Okimi OC, Oyelami OA, Iwalewa EO. Clinical effects of Garcinia kola in knee osteoarthritis. J Orthop Surg Res. 2008;3:34. doi: 10.1186/1749-799X-3-34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Maloney DJ, Hecht SM. A stereocontrolled synthesis of delta-trans-tocotrienoloic acid. Org Lett. 2005;7:4297–4300. doi: 10.1021/ol051849t. [DOI] [PubMed] [Google Scholar]
- [21].Jiang Q, Wong J, Fyrst H, Saba JD, Ames BN. gamma-Tocopherol or combinations of vitamin E forms induce cell death in human prostate cancer cells by interrupting sphingolipid synthesis. Proc Natl Acad Sci U S A. 2004;101:17825–17830. doi: 10.1073/pnas.0408340102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].Wang Y, Park NY, Jang Y, Ma A, Jiang Q. Vitamin E gamma-Tocotrienol Inhibits Cytokine-Stimulated NF-kappaB Activation by Induction of Anti-Inflammatory A20 via Stress Adaptive Response Due to Modulation of Sphingolipids. J Immunol. 2015;195:126–133. doi: 10.4049/jimmunol.1403149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Merrill AH, Jr., Sullards MC, Allegood JC, Kelly S, Wang E. Sphingolipidomics: high-throughput, structure-specific, and quantitative analysis of sphingolipids by liquid chromatography tandem mass spectrometry. Methods. 2005;36:207–224. doi: 10.1016/j.ymeth.2005.01.009. [DOI] [PubMed] [Google Scholar]
- [24].Rahmaniyan M, Curley RW, Jr., Obeid LM, Hannun YA, Kraveka JM. Identification of dihydroceramide desaturase as a direct in vitro target for fenretinide. J Biol Chem. 2011;286:24754–24764. doi: 10.1074/jbc.M111.250779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Birringer M, Lington D, Vertuani S, Manfredini S, Scharlau D, Glei M, Ristow M. Proapoptotic effects of long-chain vitamin E metabolites in HepG2 cells are mediated by oxidative stress. Free Radic Biol Med. 2010;49:1315–1322. doi: 10.1016/j.freeradbiomed.2010.07.024. [DOI] [PubMed] [Google Scholar]
- [26].Tavolari S, Bonafe M, Marini M, Ferreri C, Bartolini G, Brighenti E, Manara S, Tomasi V, Laufer S, Guarnieri T. Licofelone, a dual COX/5-LOX inhibitor, induces apoptosis in HCA-7 colon cancer cells through the mitochondrial pathway independently from its ability to affect the arachidonic acid cascade. Carcinogenesis. 2008;29:371–380. doi: 10.1093/carcin/bgm265. [DOI] [PubMed] [Google Scholar]
- [27].Entezari Heravi R, Hadizadeh F, Sankian M, Tavakol Afshari J, Taghdisi SM, Jafarian H, Behravan J. Novel selective Cox-2 inhibitors induce apoptosis in Caco-2 colorectal carcinoma cell line. Eur J Pharm Sci. 2011;44:479–486. doi: 10.1016/j.ejps.2011.09.005. [DOI] [PubMed] [Google Scholar]
- [28].Schiffmann S, Sandner J, Schmidt R, Birod K, Wobst I, Schmidt H, Angioni C, Geisslinger G, Grosch S. The selective COX-2 inhibitor celecoxib modulates sphingolipid synthesis. J Lipid Res. 2009;50:32–40. doi: 10.1194/jlr.M800122-JLR200. [DOI] [PubMed] [Google Scholar]
- [29].Jiang Q, Rao X, Kim CY, Freiser H, Zhang Q, Jiang Z, Li G. Gamma-tocotrienol induces apoptosis and autophagy in prostate cancer cells by increasing intracellular dihydrosphingosine and dihydroceramide. Int J Cancer. 2012;130:685–693. doi: 10.1002/ijc.26054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Gopalan A, Yu W, Jiang Q, Jang Y, Sanders BG, Kline K. Involvement of de novo ceramide synthesis in gamma-tocopherol and gamma-tocotrienol-induced apoptosis in human breast cancer cells. Mol Nutr Food Res. 2012;56:1803–1811. doi: 10.1002/mnfr.201200350. [DOI] [PubMed] [Google Scholar]
- [31].Triola G, Fabrias G, Casas J, Llebaria A. Synthesis of cyclopropene analogues of ceramide and their effect on dihydroceramide desaturase. J Org Chem. 2003;68:9924–9932. doi: 10.1021/jo030141u. [DOI] [PubMed] [Google Scholar]
- [32].Hannun YA, Obeid LM. Principles of bioactive lipid signalling: lessons from sphingolipids. Nat Rev Mol Cell Biol. 2008;9:139–150. doi: 10.1038/nrm2329. [DOI] [PubMed] [Google Scholar]
- [33].Rainsford KD. The ever-emerging anti-inflammatories. Have there been any real advances? J Physiol Paris. 2001;95:11–19. doi: 10.1016/s0928-4257(01)00004-3. [DOI] [PubMed] [Google Scholar]
- [34].Siddique MM, Li Y, Wang L, Ching J, Mal M, Ilkayeva O, Wu YJ, Bay BH, Summers SA. Ablation of dihydroceramide desaturase 1, a therapeutic target for the treatment of metabolic diseases, simultaneously stimulates anabolic and catabolic signaling. Mol Cell Biol. 2013;33:2353–2369. doi: 10.1128/MCB.00226-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Ruangsiriluk W, Grosskurth SE, Ziemek D, Kuhn M, des Etages SG, Francone OL. Silencing of enzymes involved in ceramide biosynthesis causes distinct global alterations of lipid homeostasis and gene expression. J Lipid Res. 2012;53:1459–1471. doi: 10.1194/jlr.M020941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [36].Signorelli P, Munoz-Olaya JM, Gagliostro V, Casas J, Ghidoni R, Fabrias G. Dihydroceramide intracellular increase in response to resveratrol treatment mediates autophagy in gastric cancer cells. Cancer Lett. 2009;282:238–243. doi: 10.1016/j.canlet.2009.03.020. [DOI] [PubMed] [Google Scholar]
- [37].Marchesini N, Hannun YA. Acid and neutral sphingomyelinases: roles and mechanisms of regulation. Biochem Cell Biol. 2004;82:27–44. doi: 10.1139/o03-091. [DOI] [PubMed] [Google Scholar]
- [38].Henry B, Ziobro R, Becker KA, Kolesnick R, Gulbins E. Acid sphingomyelinase. Handb Exp Pharmacol. 2013:77–88. doi: 10.1007/978-3-7091-1368-4_4. [DOI] [PubMed] [Google Scholar]
- [39].Zhang Y, Mattjus P, Schmid PC, Dong Z, Zhong S, Ma WY, Brown RE, Bode AM, Schmid HH, Dong Z. Involvement of the acid sphingomyelinase pathway in uva-induced apoptosis. J Biol Chem. 2001;276:11775–11782. doi: 10.1074/jbc.M006000200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Jayadev S, Liu B, Bielawska AE, Lee JY, Nazaire F, Pushkareva M, Obeid LM, Hannun YA. Role for ceramide in cell cycle arrest. J Biol Chem. 1995;270:2047–2052. doi: 10.1074/jbc.270.5.2047. [DOI] [PubMed] [Google Scholar]
- [41].Luberto C, Hassler DF, Signorelli P, Okamoto Y, Sawai H, Boros E, Hazen-Martin DJ, Obeid LM, Hannun YA, Smith GK. Inhibition of tumor necrosis factor-induced cell death in MCF7 by a novel inhibitor of neutral sphingomyelinase. J Biol Chem. 2002;277:41128–41139. doi: 10.1074/jbc.M206747200. [DOI] [PubMed] [Google Scholar]
- [42].Okazaki T, Bielawska A, Domae N, Bell RM, Hannun YA. Characteristics and partial purification of a novel cytosolic, magnesium-independent, neutral sphingomyelinase activated in the early signal transduction of 1 alpha,25-dihydroxyvitamin D3-induced HL-60 cell differentiation. J Biol Chem. 1994;269:4070–4077. [PubMed] [Google Scholar]
- [43].Abdel Shakor AB, Atia M, Ismail IA, Alshehri A, El-Refaey H, Kwiatkowska K, Sobota A. Curcumin induces apoptosis of multidrug-resistant human leukemia HL60 cells by complex pathways leading to ceramide accumulation. Biochim Biophys Acta. 2014;1841:1672–1682. doi: 10.1016/j.bbalip.2014.09.006. [DOI] [PubMed] [Google Scholar]
- [44].Mohammed A, Janakiram NB, Li Q, Choi CI, Zhang Y, Steele VE, Rao CV. Chemoprevention of colon and small intestinal tumorigenesis in APC(Min/+) mice by licofelone, a novel dual 5-LOX/COX inhibitor: potential implications for human colon cancer prevention. Cancer Prev Res (Phila) 2011;4:2015–2026. doi: 10.1158/1940-6207.CAPR-11-0233. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.