

Abstract
Our previous studies revealed that tetraspanin CD151 plays multiple roles in the progression of hepatocellular carcinoma (HCC) by forming a functional complex with integrin α6β1. Herein, we generated a monoclonal antibody (mAb) that dissociates the CD151/integrin α6β1 complex, and we evaluated its bioactivity in HCCs. A murine mAb, tetraspanin CD151 (IgG1, called CD151 mAb 9B), was successfully generated against the CD151-integrin α6β1 binding site of CD151 extracellular domains. Co-immunoprecipitation using CD151 mAb 9B followed by Western blotting detected a 28 kDa protein. Both immunofluorescent and immunohistochemical staining showed a good reactivity of CD151 mAb 9B in the plasma membrane and cytoplasm of HCC cells, as well as in liver cells. In vitro assays demonstrated that CD151 mAb 9B could inhibit neoangiogenesis and both the mobility and the invasiveness of HCC cells. An in vivo assay showed that CD151 mAb 9B inhibited tumor growth potential and HCC cells metastasis. We successfully produced a CD151 mAb 9B targeting the CD151/integrin α6β1-binding domain, which not only can displayed good reactivity to the CD151 antigen but also prevented tumor progression in HCC.
Keywords: hepatocellular carcinoma, tetraspanin CD151, integrin α6β1, monoclonal antibody, therapeutical agent
INTRODUCTION
Hepatocellular carcinoma (HCC) is the fifth most common malignant neoplasm worldwide [1]. Disease outcomes have been attributed to (1) early distant metastasis and high rates of recurrence after intervention [2] and (2) a lack of effective and curative interventions [3, 4]. An increased understanding of the molecular mechanisms underlying this disease may give us insight into potential targets for developing new preventive and therapeutic options for HCC [5].
Tetraspanin CD151, one of the most important members of the tetraspanins, has been identified as an important player in physiological processes [6] and the progression of malignant tumors [7], including skin [8], pancreas [9], lung [10], colon [11] and breast tumors [12, 13]. Consistent with relevant reports, our serial studies implicated CD151 in several pathological processes, including tumor cell mobility [14], tumor neo-angiogenesis [15] and epithelial-mesenchymal transition (EMT) [4] in HCCs. A distinct feature of CD151 is its ability to self-assemble or associate with other transmembrane molecules to form tetraspanin-enriched microdomains (TEM) [6]. Among the associated partners, integrins are the principal group of proteins that interact with tetraspanin CD151 [6]. Previously, we also identified a group of partners that associated with CD151 in the HCC cell line HCCLM3 [16]. Additionally, we validated the role of the CD151/integrin α6β1 complex in the progression of HCC [16]. Therefore, both CD151 and CD151-enriched microdomains appears to be promising targets in the treatment of HCC [17]. However, simple inhibition of CD151 in HCC is evidently inappropriate because CD151 plays essential roles in normal physiological processes, including cell adhesion, motility, activation and proliferation [6, 18–20]. Based on the above evidence, the dissociation of CD151-depedent TEM could be an effective strategy for inhibiting CD151's tumor-promoting abilities without disrupting its physiological functions [17].
Molecular targeted therapies have provided researchers with a broader perspective on the management of cancer, including colorectal and non-small cell lung cancers [21, 22]. Clinical trials have demonstrated that sorafenib, a multikinase inhibitor against Raf-1, B-Raf, VEGFR2, PDGFR and c-Kit receptors, improves progression-free survival in advanced HCC [23]. The benefit derived from sorafenib for advanced HCC patients is significant, but the drug prolongs mean survival by only 3 months compared with placebo [23]. Moreover, adverse events such as diarrhea, fatigue, weight loss, and hand-food skin reactions are frequent and limit the use of sorafenib for HCC patients. Therefore, it is important to develop alternative monoclonal antibodies for HCC [24]. In this study, we generated a CD151 mAb 9B (IgG1, κ) against the CD151/integrin α6β1-binding domain and determined its bioactivity in HCC cells.
RESULTS
Characterization of CD151 mAb 9B
A mouse anti-human CD151 mAb against the CD151/integrin α6β1 binding site [25] (Figure 1A, QRD194-196 site) was successfully produced and designated as CD151 mAb 9B. SDS-PAGE and Coomassie brilliant blue revealed two bands, including a light chain at 28 kDa and a heavy chain at 52 kDa (Figure 1B). The immunoisotype of CD151 mAb 9B was identified as IgG1 (Figure 1C). Western blotting using CD151 mAb 9B as the first antibody showed a band at 28 kDa in both HCCLM3 cell lysates and recombinant CD151 proteins (Figure 1D). A co-immunoprecipitation (IP) assay also showed that CD151 mAb 9B effectively immunoprecipitated CD151 protein as efficiently as the anti-CD151 antibody purchased from Abcam, which recognizes a different epitope of CD151. Interestingly, the integrin α6 protein cannot be detected by Western blotting from immuno-complex mixtures immunoprecipitated by CD151 mAb 9B, whereas integrin α6 can be detected from immuno-complex mixtures immunoprecipitated by the other anti-CD151 antibody. These results may indicate that CD151 mAb 9B competitively binds to the epitope through which integrin α6 binds to CD151 (Figure 1E). It was confirmed in our previous study that CD151 forms a complex with integrin α6, inducing Akt signaling. The results of the current study showed that treatment with 0.2 mg/ml of CD151 mAb 9B significantly decreased p-Akt expression in HCCLM3 cells (Figure 1F).
Figure 1. Characterization of CD151 mAb 9B.
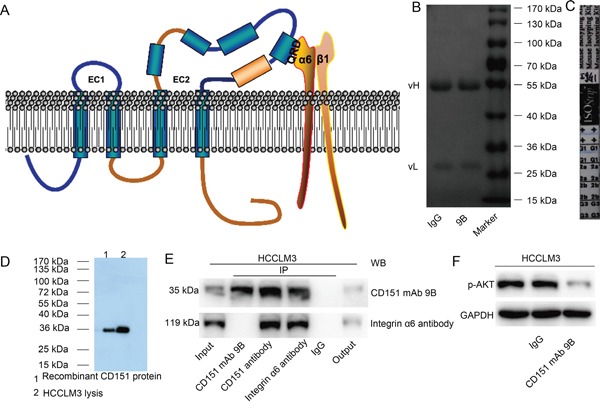
A. Schematic representation of the anti-human CD151 mAb 9B against the CD151/integrin α6β1-binding site. B. SDS-PAGE and Coomassie brilliant blue for the anti-human CD151 mAb 9B. C. The immunotype of CD151 mAb 9B. D. Western blot analysis for CD151 in HCCLM3 cells and recombinant CD151 protein using CD151 mAb 9B. E. Co-IP assay using anti-human CD151 mAb 9B in HCCLM3 cells. F. CD151 mAb 9B inhibits Akt signaling in HCCLM3 cells.
Localization and expression of CD151 protein in HCC cells and tissues
Immunofluorescent staining of HCCLM3 cells using the CD151 mAb 9B revealed that CD151 is expressed in the plasma membrane of tumor cells (Figure 2A). CD151 protein expression was detected in 30 pairs of HCC tumor and nontumor samples using immunohistochemical staining with CD151 mAb 9B. The results also showed that the immunoreactivity of CD151 was localized to the plasma membrane of the tumor cells and that the intensity of CD151 immunoreactivity in the HCC tissues was stronger than that in the paratumoral samples (Figure 2B and C, p<0.01).
Figure 2. Localization and Expression of CD151 Protein in HCCs.
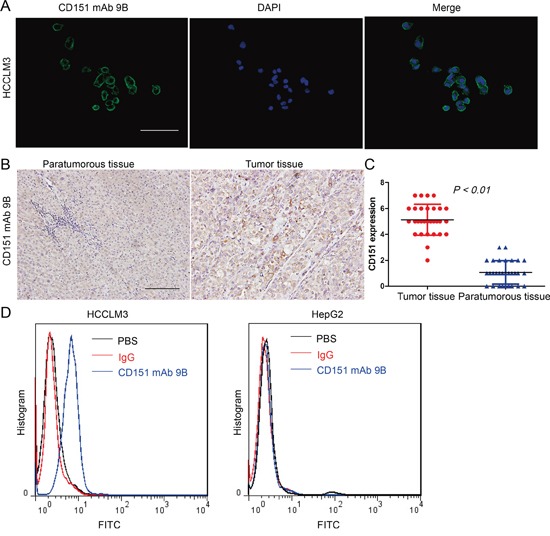
A. Immunofluorescent staining with CD151 mAb 9B in HCCLM3 cells. B. Immunohistochemical staining with CD151 mAb 9B in HCC tissues. C. Statistical analysis of CD151 expression in HCC tissues. D. FACS analysis of CD151 mAb 9B binding to the QRD194-196 conformational epitope of CD151 in HCC cells.
A FACS assay was performed to test the binding efficiency of CD151 mAb 9B with the conformational epitope of CD151 QRD194-196 in HCC cells. The results indicated that CD151 mAb 9B bound to CD151 in the HCCLM3 cells efficiently; however CD151 mAb 9B did not bind to any proteins in the extracts from the HepG2 cells, which express low levels of CD151 (Figure 2D).
CD151 mAb 9B Inhibited mobility, invasiveness and vascular remodeling in HCCs
To determine the bioactivity of CD151 mAb 9B, we performed an in vitro cell migration assay to assess its role in the mobility of tumor cells. The result showed an apparent decrease in the migratory ability of HCC cells treated with 0.2 mg/ml of CD151 mAb 9B. Representative photography indicated accelerated wound closure in the control cells (Figure 3A). This indicated that the migratory ability of the HCC cells was markedly suppressed after the administration of CD151 mAb 9B. Next, a transwell assay was used to investigate the role of CD151 mAb 9B in the invasiveness of tumor cells. The result showed that the average number of invaded cells significantly decreased after treatment with 0.2 mg/ml of CD151 mAb 9B compared with that of the control cells (Figure 3B, p<0.05). Third, a matrigel angiogenesis assay demonstrated that significantly more integrated capillary-like structures were found in the HUVECs cultured with DMEM and 10% fetal bovine serum than in the HUVECs supplemented with 0.2 mg/ml of CD151 mAb 9B (Figure 3C). However, we further determined the effect of the CD151 mAb 9B on the growth of the HCC cells and found that CD151 mAb 9B does not affect the proliferation of HCC cells (Figure 3D, p>0.05).
Figure 3. The effect of CD151 mAb 9B on HCCs.
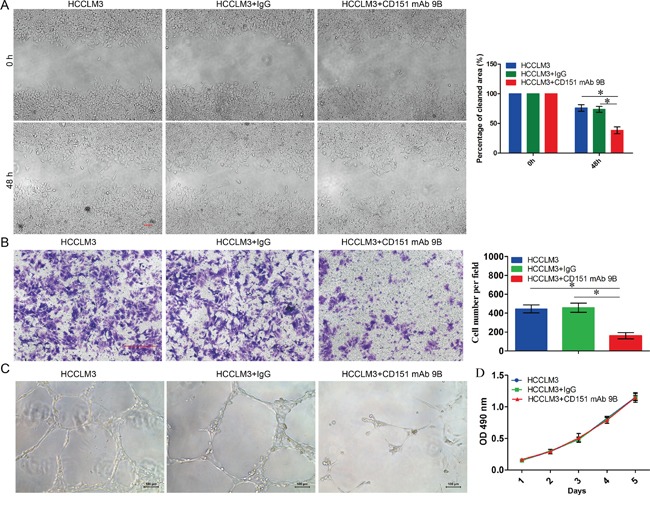
A. Wound healing assay for the effect of CD151 mAb 9B on the mobility of tumor cells. B. Transwell assay for the effect of CD151 mAb 9B on the invasiveness of tumor cells. C. Matrigel angiogenesis to assay the effect of CD151 mAb 9B on the invasiveness of tumor cells. D. MTT assay for the effect of CD151 mAb 9B on the invasiveness of tumor cells. *P < 0.05.
CD151 mAb 9B inhibited neoangiogenesis and tumor growth and attenuated lung metastasis of HCC cells
To further investigate the role of CD151 mAb 9B in vivo, we used a subcutaneous metastasis model to assay its impact on the efficiency of tumor growth and progression. The results revealed that the tumor volume and weight of the HCCLM3 cell-derived xenografts treated with 25 mg/kg CD151 mAb 9B was much lower than that in of the HCCLM3 cell-derived xenografts treated with PBS (Figure 4A, B and C;741.3 ± 191.4 mm3 vs. 1195.2 ± 202.5 mm3, 3.42 ± 0.88 g vs. 5.51 ± 0.93 g).
Figure 4. CD151 mAb 9B Inhibited the progression of HCCs in vivo.
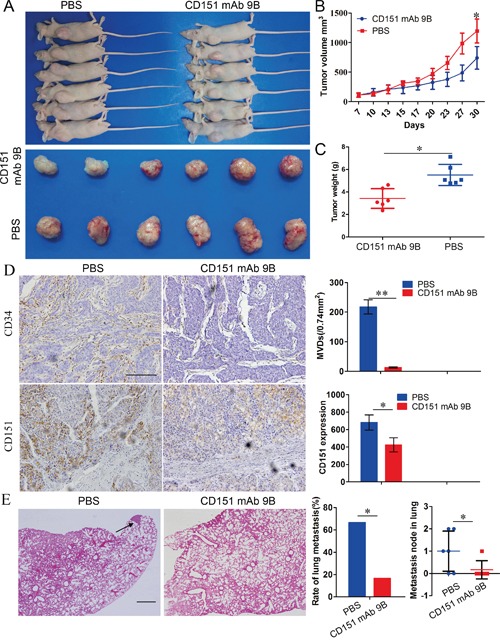
A, B and C. Subcutaneous metastasis model to assay the effect of CD151 mAb 9B on the growth of tumor cells. D. IHC staining for CD34 and CD151 to assay the effect of CD151 mAb 9B on neoangiogenesis. E. Lung metastasis model to assay the effect of CD151 mAb 9B on the metastasis. *P< 0.05, **P< 0.01.
The microvessel density (MVDs) of HCCLM3 cell-derived xenografts treated with PBS were 217.69 ± 24.13/0.74 mm2 and were larger than those of the HCCLM3 cell-derived xenografts treated with 25 mg/kg CD151 mAb 9B (12.68 ± 2.47/0.74 mm2) (Figure 4D, p<0.01). CD151 expression in the HCCLM3 cell-derived xenografts treated with PBS was stronger than in the HCCLM3 cell-derived xenografts treated with 25 mg/kg CD151 mAb 9B (Figure 4D, p<0.05). These results indicate that CD151 mAb 9B inhibits neoangiogenesis through blockage of CD151-dependent TEM in HCC tissues.
Pulmonary metastases occurred in 66.7% (4/6) of the PBS group. However, a lower rate was observed in the CD151 mAb 9B group (16.7%, 1/6; Figure 4E). These data indicate that CD151 mAb 9B can inhibit tumor growth and the progression of HCC cells in vivo.
DISCUSSION
Previously, our studies showed that CD151 is an important player in several processes during the progression of HCC [4, 14–16] and is considered an ideal therapeutic target for HCC [4, 14–17]. In the present study, we successfully produced a CD151 mAb 9B targeting tetraspanin CD151.
Intriguingly, the expression of CD151 was significantly reduced after CD151 mAb 9B treatment in vivo, which would be expected to result in reductions of both p-Akt and CD34 expression, as well as a reduced rate of lung metastasis. The alteration of these downstream molecular signals and phenotypes was similar to that observed in CD151 knockdown HCC cells [26]. It should be noted that a similar assay was applied in vitro as well. The above similarity suggests that CD151 mAb 9B exerts its antitumor effect though competitive binding to the tetraspanin CD151/integrin α6β1-binding domain.
In a previous study, we identified a set of proteins associated with CD151 in HCCLM3 cells (Established in Liver Cancer Institute, Zhongshan Hospital) and identified an important role for the CD151/integrin α6β1 complex in the progression of HCC [16]. Therefore, CD151-dependent TEM appears to be promising therapeutic targets for HCC [17]. Given that CD151 implicates in physiological processes, such as cell adhesion, motility, activation and proliferation [6, 18–20], simple blockage of CD151 in HCC is evidently inappropriate. Based on the above evidence, the dissociation of CD151-depedent TEM could be an effective strategy for inhibiting CD151's tumor-promoting abilities without disrupting its physiological functions [17]. Recent studies have shown that the QRD194–196 site of CD151 was required for binding with integrin α6β1 and its epitope [25]. In the present study, we chemically synthesized peptides of the CD151/integrin α6β1-binding domain (GQRDHASNIYKVEGGC) and then successfully produced a CD151 mAb 9B with a molecular weight of 28kDa. Second, CD151 mAb 9B displayed good reactivity to the CD151 antigen in HCCs. The newly synthesized antibody not only accurately reflected the intensity of the CD151 antigen by Western blotting but also correctly displayed the localization of the CD151 antigen by immunofluorescent and immunohistochemical staining, which indicates that it can be used in detection of the expression and localization of CD151 antigen in basic research. Third, CD151 mAb 9B showed good bioactivity for HCCs. On one hand, the newly synthesized antibody significantly inhibited the mobility and invasiveness of HCC cells in vitro. On the other hand, CD151 mAb 9B also inhibited growth and lung metastasis in vivo. Interestingly, CD151 mAb 9B had a minimal effect on the proliferation of HCC cells in vitro. These data indicate that CD151 mAb 9B plays an important role in the progression of HCC by targeting the CD151/integrin α6β1-binding; therefore, CD151 mAb 9B could overcome the limitation of CD151 blockage in HCC.
In conclusion, we successfully produced a CD151 mAb 9B that competitively binds to the tetraspanin CD151/integrin α6β1-binding domain. This mAb may be widely used for the detection of the expression and localization of the CD151 antigen in HCCs. More importantly, CD151 mAb 9B may be an efficient method for inhibiting of the progression of HCCs.
MATERIALS AND METHODS
Cell lines and animals
HCC cell lines HCCLM3, MHCC97-L (established in Liver Cancer Institute, Zhongshan Hospital [27]), HepG2 (American Type Culture Collection) and SP2/0 myeloma cells (American Type Culture Collection) were used in this study and routinely raised. Male BALB/c mice (8 weeks old) and male athymic BALB/c nude mice (4 weeks old) were purchased from the Shanghai Institute of Material Medicine and raised in specific pathogen-free conditions. Animal care was provided according to the guidelines established by the Zhongshan Hospital Experimental Animal Care Commission.
Sample collection
Thirty pairs of fresh tumor and corresponding nontumor samples were collected from consecutive HCC patients who underwent curative resection at Zhongshan Hospital. All HCC samples were identified by histopathological examination. Of these 30 patients, 26 patients had a history of hepatitis B. No patients received preoperative anticancer treatment. Ethical approval was provided by the research ethics committee of Zhongshan Hospital, and written informed consent was obtained from each patient.
Preparation and purification of anti-CD151 monoclonal antibodies
The monoclonal antibody was produced as previously described [28]. Briefly, small peptides were designed according to the CD151/integrin α6β1-binding domain (GQRDHASNIYKVEGGC), chemically synthesized and coupled with keyhole limpet hemocyanin (KLH). BALB/c mice were immunized with CD151/integrin α6β1 binding domain recombinant peptides. Splenocyte fusion was performed with SP2/0 myeloma cells, from which hybridomas of the desired characteristics were subsequently selected. The antibody was purified by protein G. The immunoglobulin subtype of the antibody was determined using an antibody isotyping kit (Roche Molecular Biochemicals, Indianapolis, IN, USA). The purified mAb was named CD151 mAb 9B and used for subsequent experiments.
Western blotting, co-immunoprecipitation (co-IP), immunofluorescent (IF) and immunohistochemical (IHC) staining
Western blotting was performed as previously described [15]. CD151 mAb 9B (1:2000) was used to detect the expression of CD151 in three HCC cell lines and tumor samples from 30 patients. β-actin (1:5,000; Chemicon, USA) was used as an internal control. Co-IP was conducted as previously described [16]. Five milligrams of CD151 mAb 9B was used to extract CD151 from the HCCLM3 lysates (250 mg). The gel was stained using the PlusOne Coomassie brilliant blue (Amersham Biosciences). Immunofluorescent staining was performed as previously described [14]. The expression and localization of CD151 protein in HCCLM3 cells were assayed using the CD151 mAb 9B (1:200). The immunohistochemistry protocols are described elsewhere [14]. CD151 mAb 9B (1:200) detected the expression of CD151 in the HCC samples. Positive staining was measured as described in our previous report [14].
FACS analysis
A total of 1 × 106 cells were collected by centrifugation and incubated with 20 μg/ml of either CD151 mAb 9B or IgG in PBS containing 1% newborn calf serum for 45 min at 4°C. After being washed with cold PBS three times, the cells were incubated for an additional 45 min at 4°C with a FITC-conjugated secondary antibody (Kang-Chen Bio-tech, Shanghai, China) in the dark. The cells were then analyzed by FACS cytometry (Beckman Coulter Epics Altra, Miami, FL).
Wound healing, invasion assay and matrigel angiogenesis assay
A wound-healing assay was used to evaluate the effect of CD151 mAb 9B on cell migration [29]. These cells were raised in serum-free medium supplemented with or without 0.2 mg/ml CD151 mAb 9B. The invasion assay was performed as described elsewhere [14]. The supernatant of NIH3T3 in DMEM supplemented with 10% fetal bovine serum with or without 0.2 mg/ml CD151 mAb 9B, which served as a chemoattractant, was placed in the lower chamber. All experiments were performed in triplicate. The Matrigel angiogenesis assay was performed as described previously [15]. The human umbilical vein endothelial cells (HUVECs) were cultured with DMEM supplemented with 10% fetal bovine serum with or without 0.2 mg/ml CD151 mAb 9B.
Cell proliferation assay
Cell proliferation was analyzed using a 3-(4,5)- dimethylthiahiazo (-z-y1)-3,5-di- phenytetrazoliumromide (MTT) assay as previously described [14]. These cells were raised with DMEM supplemented with 10% fetal bovine serum and 0.2 mg/ml CD151 mAb 9B or IgG.
In vivo metastasis assays and immunohistochemical analysis
A total of 6.0×106 HCCLM3 cells were used for subcutaneous xenografts in a spontaneous metastasis assay as previously described [15]. When the tumors reached a mean tumor volume of 100 mm3, the mice were randomly allocated into two groups (n=6); 25 mg/kg of either CD151 mAb 9B or phosphate buffer saline (PBS) was administered intraperitoneally three times per week for two weeks, and the diameter of the xenografts was monitored twice a week. The xenografts and visceral organs, including the lungs, were examined histopathologically. Tumor volume, weight, and the total number of lung metastases were assayed as previously described [30, 31].
Mouse anti-human CD34 antibodies (1:100; DakoCytomation, Denmark) and CD151 mAb 9B were used to measure microvessel density (MVD) and CD151 expression. MVD and CD151 were evaluated as described elsewhere [15].
Statistical analysis
The statistical analysis was performed with SPSS 16.0 (SPSS, Chicago, IL). Values are expressed as the mean ± standard deviation. Quantitative data between groups were compared using Student's t test. Categorical data were analyzed using the χ2 test or Fisher's exact test. P<0.05 was considered statistically significant.
Acknowledgments
This study was supported by the National Key Sci-Tech Project (2012ZX10002011-002), the National Natural Science Foundation of China (81472840, 81160062, 81272295, 81172023 and 81071741) and the Shanghai Municipal Natural Science Foundation (14ZR1405800, 11ZR1428300, 114119a5000, and 12ZR1402200).
Abbreviations
- HCC
hepatocellular carcinoma
- TEM
tetraspanin-enriched microdomains
- TNM
tumor node metastasis
- mAb
monoclonal antibody
- EMT
epithelial-mesenchymal transition
Footnotes
CONFLICTS OF INTEREST
The authors have no conflicts of interest.
REFERENCES
- 1.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA: a cancer journal for clinicians. 2011;61:69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 2.Clavien PA, Petrowsky H, DeOliveira ML, Graf R. Strategies for safer liver surgery and partial liver transplantation. The New England journal of medicine. 2007;356:1545–1559. doi: 10.1056/NEJMra065156. [DOI] [PubMed] [Google Scholar]
- 3.Sherman M. Recurrence of hepatocellular carcinoma. The New England journal of medicine. 2008;359:2045–2047. doi: 10.1056/NEJMe0807581. [DOI] [PubMed] [Google Scholar]
- 4.Shi GM, Xu Y, Fan J, Zhou J, Yang XR, Qiu SJ, Liao Y, Wu WZ, Ji Y, Ke AW, Ding ZB, He YZ, Wu B, et al. Identification of side population cells in human hepatocellular carcinoma cell lines with stepwise metastatic potentials. Journal of cancer research and clinical oncology. 2008;134:1155–1163. doi: 10.1007/s00432-008-0407-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ji J, Shi J, Budhu A, Yu Z, Forgues M, Roessler S, Ambs S, Chen Y, Meltzer PS, Croce CM, Qin LX, Man K, Lo CM, et al. MicroRNA expression, survival, and response to interferon in liver cancer. The New England journal of medicine. 2009;361:1437–1447. doi: 10.1056/NEJMoa0901282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hemler ME. Tetraspanin functions and associated microdomains. Nat Rev Mol Cell Biol. 2005;6:801–811. doi: 10.1038/nrm1736. [DOI] [PubMed] [Google Scholar]
- 7.Zoller M. Tetraspanins: push and pull in suppressing and promoting metastasis. Nature reviews. 2009;9:40–55. doi: 10.1038/nrc2543. [DOI] [PubMed] [Google Scholar]
- 8.Sachs N, Secades P, van Hulst L, Song JY, Sonnenberg A. Reduced Susceptibility to Two-Stage Skin Carcinogenesis in Mice with Epidermis-Specific Deletion of Cd151. The Journal of investigative dermatology. 2014;134:221–228. doi: 10.1038/jid.2013.280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gesierich S, Paret C, Hildebrand D, Weitz J, Zgraggen K, Schmitz-Winnenthal FH, Horejsi V, Yoshie O, Herlyn D, Ashman LK, Zoller M. Colocalization of the tetraspanins, CO-029 and CD151, with integrins in human pancreatic adenocarcinoma: impact on cell motility. Clin Cancer Res. 2005;11:2840–2852. doi: 10.1158/1078-0432.CCR-04-1935. [DOI] [PubMed] [Google Scholar]
- 10.Tokuhara T, Hasegawa H, Hattori N, Ishida H, Taki T, Tachibana S, Sasaki S, Miyake M. Clinical significance of CD151 gene expression in non-small cell lung cancer. Clin Cancer Res. 2001;7:4109–4114. [PubMed] [Google Scholar]
- 11.Chien CW, Lin SC, Lai YY, Lin BW, Lin SC, Lee JC, Tsai SJ. Regulation of CD151 by hypoxia controls cell adhesion and metastasis in colorectal cancer. Clin Cancer Res. 2008;14:8043–8051. doi: 10.1158/1078-0432.CCR-08-1651. [DOI] [PubMed] [Google Scholar]
- 12.Yang XH, Richardson AL, Torres-Arzayus MI, Zhou P, Sharma C, Kazarov AR, Andzelm MM, Strominger JL, Brown M, Hemler ME. CD151 accelerates breast cancer by regulating alpha 6 integrin function, signaling, and molecular organization. Cancer research. 2008;68:3204–3213. doi: 10.1158/0008-5472.CAN-07-2949. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zijlstra A, Lewis J, Degryse B, Stuhlmann H, Quigley JP. The inhibition of tumor cell intravasation and subsequent metastasis via regulation of in vivo tumor cell motility by the tetraspanin CD151. Cancer cell. 2008;13:221–234. doi: 10.1016/j.ccr.2008.01.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ke AW, Shi GM, Zhou J, Wu FZ, Ding ZB, Hu MY, Xu Y, Song ZJ, Wang ZJ, Wu JC, Bai DS, Li JC, Liu KD, et al. Role of overexpression of CD151 and/or c-Met in predicting prognosis of hepatocellular carcinoma. Hepatology. 2009;49:491–503. doi: 10.1002/hep.22639. [DOI] [PubMed] [Google Scholar]
- 15.Wang ZC, Gao Q, Shi JY, Guo WJ, Yang LX, Liu XY, Liu LZ, Ma LJ, Duan M, Zhao YJ, Wu YN, Gao DM, Wang XY, et al. PTPRS Acts as A Metastatic Suppressor in Hepatocellular Carcinoma by Control of EGFR Induced Epithelial-Mesenchymal Transition. Hepatology. 2015;62:1201–1214. doi: 10.1002/hep.27911. [DOI] [PubMed] [Google Scholar]
- 16.Devbhandari RP, Shi GM, Ke AW, Wu FZ, Huang XY, Wang XY, Shi YH, Ding ZB, Xu Y, Dai Z, Fan J, Zhou J. Profiling of the tetraspanin CD151 web and conspiracy of CD151/integrin beta1 complex in the progression of hepatocellular carcinoma. PloS one. 2011;6:e24901. doi: 10.1371/journal.pone.0024901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hemler ME. Targeting of tetraspanin proteins—potential benefits and strategies. Nat Rev Drug Discov. 2008;7:747–758. doi: 10.1038/nrd2659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Liu L, He B, Liu WM, Zhou D, Cox JV, Zhang XA. Tetraspanin CD151 promotes cell migration by regulating integrin trafficking. The Journal of biological chemistry. 2007;282:31631–31642. doi: 10.1074/jbc.M701165200. [DOI] [PubMed] [Google Scholar]
- 19.Sachs N, Claessen N, Aten J, Kreft M, Teske GJ, Koeman A, Zuurbier CJ, Janssen H, Sonnenberg A. Blood pressure influences end-stage renal disease of Cd151 knockout mice. The Journal of clinical investigation. 2012;122:348–358. doi: 10.1172/JCI58878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Karamatic Crew V, Burton N, Kagan A, Green CA, Levene C, Flinter F, Brady RL, Daniels G, Anstee DJ. CD151, the first member of the tetraspanin (TM4) superfamily detected on erythrocytes, is essential for the correct assembly of human basement membranes in kidney and skin. Blood. 2004;104:2217–2223. doi: 10.1182/blood-2004-04-1512. [DOI] [PubMed] [Google Scholar]
- 21.Maemondo M, Inoue A, Kobayashi K, Sugawara S, Oizumi S, Isobe H, Gemma A, Harada M, Yoshizawa H, Kinoshita I, Fujita Y, Okinaga S, Hirano H, et al. Gefitinib or chemotherapy for non-small-cell lung cancer with mutated EGFR. The New England journal of medicine. 2010;362:2380–2388. doi: 10.1056/NEJMoa0909530. [DOI] [PubMed] [Google Scholar]
- 22.Ciardiello F, Tortora G. EGFR antagonists in cancer treatment. The New England journal of medicine. 2008;358:1160–1174. doi: 10.1056/NEJMra0707704. [DOI] [PubMed] [Google Scholar]
- 23.Llovet JM, Ricci S, Mazzaferro V, Hilgard P, Gane E, Blanc JF, de Oliveira AC, Santoro A, Raoul JL, Forner A, Schwartz M, Porta C, Zeuzem S, Bolondi L, Greten TF, Galle PR, et al. Sorafenib in advanced hepatocellular carcinoma. The New England journal of medicine. 2008;359:378–390. doi: 10.1056/NEJMoa0708857. [DOI] [PubMed] [Google Scholar]
- 24.Luk JM, Wong KF. Monoclonal antibodies as targeting and therapeutic agents: prospects for liver transplantation, hepatitis and hepatocellular carcinoma. Clinical and experimental pharmacology & physiology. 2006;33:482–488. doi: 10.1111/j.1440-1681.2006.04396.x. [DOI] [PubMed] [Google Scholar]
- 25.Kazarov AR, Yang X, Stipp CS, Sehgal B, Hemler ME. An extracellular site on tetraspanin CD151 determines alpha 3 and alpha 6 integrin-dependent cellular morphology. The Journal of cell biology. 2002;158:1299–1309. doi: 10.1083/jcb.200204056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ke AW, Shi GM, Zhou J, Huang XY, Shi YH, Ding ZB, Wang XY, Devbhandari RP, Fan J. CD151 amplifies signaling by integrin alpha6beta1 to PI3K and induces the epithelial-mesenchymal transition in HCC cells. Gastroenterology. 2011;140:1629–1641 e1615. doi: 10.1053/j.gastro.2011.02.008. [DOI] [PubMed] [Google Scholar]
- 27.Ye QH, Qin LX, Forgues M, He P, Kim JW, Peng AC, Simon R, Li Y, Robles AI, Chen Y, Ma ZC, Wu ZQ, Ye SL, et al. Predicting hepatitis B virus-positive metastatic hepatocellular carcinomas using gene expression profiling and supervised machine learning. Nature medicine. 2003;9:416–423. doi: 10.1038/nm843. [DOI] [PubMed] [Google Scholar]
- 28.Luk JM, Su YC, Lam SC, Lee CK, Hu MY, He QY, Lau GK, Wong FW, Fan ST. Proteomic identification of Ku70/Ku80 autoantigen recognized by monoclonal antibody against hepatocellular carcinoma. Proteomics. 2005;5:1980–1986. doi: 10.1002/pmic.200401084. [DOI] [PubMed] [Google Scholar]
- 29.Zhang C, Bai DS, Huang XY, Shi GM, Ke AW, Yang LX, Yang XR, Zhou J, Fan J. Prognostic significance of Capn4 overexpression in intrahepatic cholangiocarcinoma. PloS one. 2013;8:e54619. doi: 10.1371/journal.pone.0054619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tian J, Tang ZY, Ye SL, Liu YK, Lin ZY, Chen J, Xue Q. New human hepatocellular carcinoma (HCC) cell line with highly metastatic potential (MHCC97) and its expressions of the factors associated with metastasis. British journal of cancer. 1999;81:814–821. doi: 10.1038/sj.bjc.6690769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhou ZJ, Dai Z, Zhou SL, Hu ZQ, Chen Q, Zhao YM, Shi YH, Gao Q, Wu WZ, Qiu SJ, Zhou J, Fan J. HNRNPAB induces epithelial-mesenchymal transition and promotes metastasis of hepatocellular carcinoma by transcriptionally activating SNAIL. Cancer research. 2014;74:2750–2762. doi: 10.1158/0008-5472.CAN-13-2509. [DOI] [PubMed] [Google Scholar]