

Abstract
Neural stem cell (NSC) transplantation into the hippocampus could offer an alternative therapy to hippocampal resection in patients with drug-resistant chronic epilepsy, which afflicts ~30% of mesial temporal lobe epilepsy (TLE) cases. Multipotent, self-renewing NSCs could be expanded from multiple regions of the developing and adult brain, human embryonic stem cells (hESCs), induced pluripotent stem cells (iPSCs). However, to provide a comprehensive methodology involved in testing the efficacy of transplantation of NSCs in a rat model of chronic TLE, NSCs derived from the embryonic medial ganglionic eminence (MGE) are taken as an example in this article. The topics comprise description of the required materials, reagents and equipment, and protocols for expanding MGE-NSCs in culture, generating chronically epileptic rats, the intrahippocampal grafting, the post-grafting evaluation of the effects of NSC grafts on spontaneous recurrent seizures and cognitive impairments, analyses of the yield and the fate of graft-derived cells, and the effects of NSC grafts on the host hippocampus.
Keywords: Chronic temporal lobe epilepsy, Dentate neurogenesis, Glial-cell line derived neurotrophic factor, Hippocampal neurogenesis, Learning and memory, Medial ganglionic eminence, Neural stem cells, Neural stem cell transplantation, Neurotrophic factors, Phenotypic differentiation of graft-derived cells, Stem cell proliferation, Stem cell differentiation, Spontaneous recurrent seizures
INTRODUCTION
Chronic temporal lobe epilepsy (TLE) is characterized by recurrent partial complex seizures, memory impairments, depression, anxiety and substantially declined hippocampal neurogenesis (Astur et al., 2002; Detour et al., 2005; Hattiangady et al, 2004, 2010; Coras et al., 2010). Antiepileptic drug therapy, though widely used for controlling seizures, has no effect on the course of the disease and fails to restrain seizures in ~30% of TLE patients (Strine et al., 2005, Fisher et al., 1998). Intracerebral transplantation of NSCs is evolving as an attractive therapy for promoting regeneration and repair in various brain disorders including the TLE. This is because NSCs are multipotent, have ability for self-renewal, and could be expanded from multiple regions of the developing and adult brain, human embryonic stem cells (hESCs), and induced pluripotent stem cells (iPSCs) (Shetty and Hattiangady, 2007).
Studies have also shown that NSCs can survive intracerebral grafting, engraft into the injured brain areas, release a multitude of neurotrophic factors and positively influence the survival of host cells and tissues and promote functional recovery. Transplantation of appropriate NSCs in TLE (such as NSCs from the medial ganglionic eminence [MGE] and the subventricular zone [SVZ]) may considerably restrain seizures because of their ability to give rise to a large number of neurons synthesizing the inhibitory neurotransmitter gamma-amino butyric acid (GABA) and astrocytes synthesizing the anticonvulsant protein the glial-cell line derived neurotrophic factor (GDNF) (Waldau et al., 2010). Grafting of NSCs may also improve the cognitive function in chronic TLE because of the ability of NSCs to engraft into the neurogenic regions of the dentate gyrus and thereby influence the extent of hippocampal neurogenesis.
In this unit, to provide a detailed methodology involved in evaluating the usefulness of transplantation of NSCs in a rat model of chronic TLE, we describe the protocol for grafting of NSCs expanded from the MGE of the embryonic day 14 rat fetuses into hippocampi of rats exhibiting chronic TLE. The protocols mainly include description of the required materials, reagents and equipment, and the methodology for expanding MGE-NSCs in culture, generating chronically epileptic rats, the intrahippocampal grafting, the post-grafting evaluation of the effects of NSC grafts on spontaneous recurrent seizures and learning and memory impairments, analyses of the yield and the fate of graft-derived cells, and the effects of NSC grafts on the host hippocampus.
Figure 1 shows schematic representation of major experiments described in this manuscript.
Figure 1.
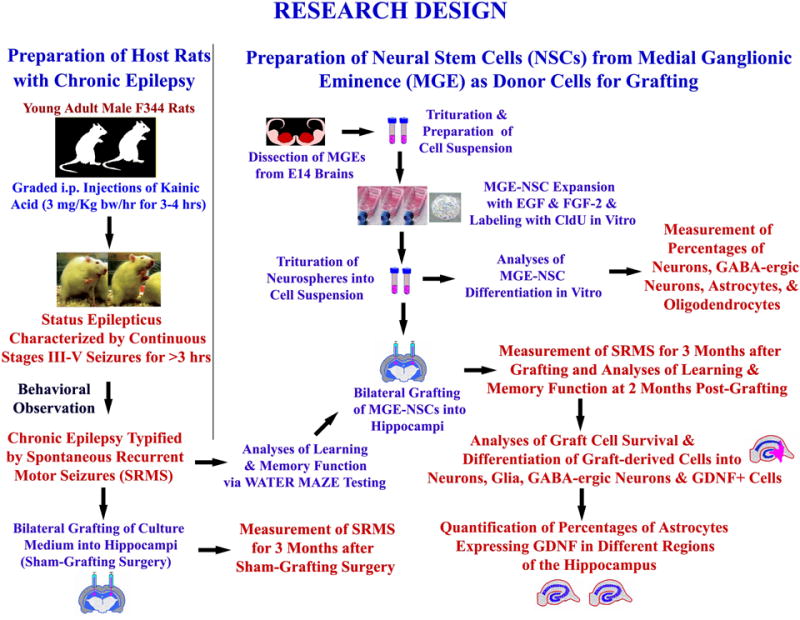
Experimental design of a MGE-NSC grafting study performed in chronically epileptic rats. It shows the creation of host rats with chronic epilepsy for grafting and sham-grafting surgery (on the left side), and the preparation of fresh neural stem cells (NSCs) expanded in vitro from embryonic medial ganglionic eminence (MGE) as donor cells (upper half on right). The various measurements performed after MGE-NSC grafting include frequency, duration and severity of spontaneous seizures, learning and memory function, graft cell survival and differentiation, and effects of grafts on expression of GDNF in hippocampal astrocytes (lower half on right). [Reproduced from: Waldau et al., Stem Cells, 28:1153–1164].
NOTE: All protocols using live animal studies must be first reviewed and approved by the institutional Animal Care and Use committee (IACUC). The experimenter must strictly follow all the guidelines recommended by the IACUC while performing the experiments in animal models.
GENERATION OF THE HOST RATS EXHIBITING CHRONIC TLE
In this protocol, we describe how to generate rats exhibiting chronic temporal lobe epilepsy characterized by spontaneous recurrent motor seizures (SRMS) and cognitive dysfunction using a chemoconvulsant chemical (i.e. kainic acid [KA]) to induce status epilepticus. As generation of rats exhibiting chronic TLE requires a time-frame of 3–5 months, the experiments to be performed on chronically epileptic rats need to be planned well in advance. Furthermore, as the extent of SRMS varies between animals (Rao et al., 2006, 2007; Waldau et al., 2010), having a larger pool of rats exhibiting chronic TLE would help in choosing animals exhibiting a similar extent (frequency and intensity) of SRMS for the transplantation study.
Materials
Experimental animals. Our laboratory employs 4–5 months old male Fischer 344 (F344) rats
Kainic Acid (KA)
Soft foam covered cage tops
A rat cage placed on ice
Saline or Ringer’s lactate solution
1 ml syringes with appropriate needles for intraperitoneal injection of kainic acid
5 or 10 ml syringes with appropriate needles for subcutaneous injection of saline or Ringer’s lactate.
Regular rat chow soaked in water (soft pellets) and transgel.
Protocol for Induction of Status Epilepticus (SE) in Adult Male F344 rats
Establish the animal model
-
1
Order 4–5 months old male F344 rats and allow them to acclimatize to the new animal resources center (or the vivarium) for at least a week.
Other stains of rats such as Sprague-Dawley may also be used but they appear to require higher or additional doses of KA for induction of SE (see Hellier et al., 1998 for details). Acute seizure behavior varies depending on the age and sex of the animal and hence the protocol described here is good only for the 4–5 months old male F344 rats. If induction of SE is planned for female, younger or aged rats, it is important to standardize the required dose and injections of KA for eliciting SE in these models.
-
2
Prepare a desired amount of the KA solution (e.g. 3.0 mg/ml in sterile saline).
As KA can be obtained from multiple sources, it will be important to stick to a single source to avoid confounds in SE induction between different groups of rats. We currently use the KA sold by the Tocris (USA) which has worked well in our experiments.
-
3
Measure the weight of each rat and inject KA intraperitoneally at a dose of 3.0 mg/kg bw/hr. Three-four injections of KA are typically sufficient for inducing SE in most rats for the age group mentioned above.
It is possible that some rats may develop SE with just two injections of KA while some others may need additional (i.e. >4) injections at a full dose (3mg/Kg/bw) or at a half-dose (1.5 mg/Kg bw) for exhibiting SE. Therefore, it is important to closely observe and score the type and intensity of acute seizures after two injections of KA and empirically determine whether or not additional KA injections would be required to induce SE on a rat-by-rat basis.
Furthermore, a small fraction of rats (5–10%) exhibit freezing behavior and fail to show the features of SE even with four injections of KA. Such rats need to be excluded from the study as additional KA injections would typically lead to mortality in these animals.
Scoring of acute seizures induced by KA
It will be difficult to score stages I-II seizures that are characterized by salivation, excessive grooming behavior, mastication, wet dog shakes etc. However, stages III-V seizures are much easier to follow and score them using a modified Racine scale (Racine, 1972; Ben-Ari, 1985; Hellier et al., 1998).
-
4Seizures are scored as follows:
The definition of the onset of SE in experimental animals varies in different studies depending upon the behavior of the animal model employed in the study. Based on our experience in scoring acute seizures in a large number of male F344 rats injected with KA over the last 5 years, our definition of the onset of SE has also evolved. We now identify the onset of SE in male F344 rats as the occurrence of the first stage V seizure followed by continuous stages III-V seizures for over 10 minutes (Type 1 SE) or continuous stage IV seizures for over 10 minutes (Type 2 SE).
Rats exhibiting Type 1 or Type 2 SE typically continue to have intermittent stages III-V seizures for over 3 hrs at a rate of 5–10 seizures per hr. Therefore, once a rat exhibits either Type 1 or Type 2 SE, it is prudent to discontinue KA injections and to score the rat for seizures in the next 3 hours to determine the extent of SE. Virtually all rats that exhibit Type 1 or Type 2 SE and display 5–10 seizures during the 2nd and 3rd hour after SE would develop chronic epilepsy characterized by SRMS by 3–6 moths post-SE. However, it is possible that a small fraction (~5%) of rats exhibiting Type 1 or Type 2 SE may not display additional seizures in the 2nd and 3rd hour after SE. It will be important to exclude these rats from the study, as such rats rarely develop chronic TLE.
Figure 2.
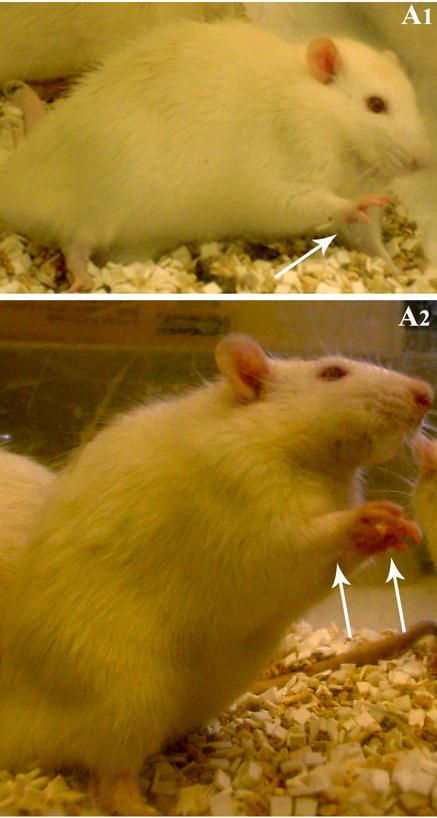
Examples of a Stage III seizure characterized by the unilateral forelimb clonus (A1) and a stage IV seizure typified by a bilateral forelimb clonus (A2).
Treat animals post-SE
-
5
In our earlier studies, we left the rats after SE to recover without any antiepileptic medication (Rao et al., 2006, 2007). While such rats developed robust chronic epilepsy (typified by the increased frequency and intensity of SRMS), they were prone to increased mortality both immediately after SE and also during the chronic phase of epilepsy. Therefore, over the last few years, we have tested the effect of diazepam injection at a dose of 5 mg/Kg body weight once the animal completes 3 hrs of seizure activity. Based on our experience so far, diazepam injection after 3 hrs of seizure activity reduces mortality of animals that undergo SE and also does not interfere with the development of chronic epilepsy.
Animals treated with diazepam after SE display reduced frequency and intensity of SRMS in the chronic phase of epilepsy, in comparison to animals treated with no diazepam after SE. Therefore, it is important to stick to one SE protocol (either with or without diazepam) to avoid confounds in an experiment.
Care for Animals Post-SE
-
6
Observe animals for tiredness and lethargy after SE. Provide moistened standard rat chow and transgel in a small dish placed inside the cage within a range that is easily accessible to the rat. Additionally, to prevent dehydration, give subcutaneous injections of Ringer’s lactate solution (5–10 ml/day) every day for 3–5 days following SE. Rats that undergo SE will exhibit hematuria for a day or two and hence multiple red spots will be seen on cage bedding.
Typically, hematuria stops on its own within two days after SE.
SELECTING RATS EXHIBITING CHRONIC TLE
Measure SRMS in the animals
Following SE, there will be a silent period of 1–2 months during which no or occasional SRMS are observed. Therefore, commencing the measurement of SRMS in the 3rd month after SE is ideal. In most of our studies, we intermittently score the frequency and duration of stages III-V SRMS at 3–6 months post-SE (i.e. 8 hrs/week; 4 hrs/session; 2 sessions/week, total 32 hrs/month) to determine the extent/pattern of chronic epilepsy. From the recorded seizures, calculate the following parameters for every month of observation: the frequency of all (stages III-V) SRMS, the frequency of stage V seizures (the most severe form of SRMS), the average duration of individual SRMS (i.e. the total amount of time spent in seizures/the total number of seizures), and the percentage of time spent in SRMS (i.e. the total amount of time spent in seizures/the total duration of observation × 100).
Intermittent scoring of the frequency and intensity of SRMS for several months (i.e. 8 hrs/week; 4 hrs/session; 2 sessions/week, total 32 hrs/month) has been found to be sufficient for determining the extent/intensity of chronic epilepsy typified by SRMS in male F344 rats (Rao et al., 2006, 2007; Waldau et al., 2010). However, measuring the frequency and intensity of SRMS with additional hours of observation or continuous (24/7) video-monitoring will be superior and especially important if animals mostly exhibit SRMS in clusters.
Select CERs for grafting studies
Select groups of age-matched chronically epileptic animals exhibiting a similar extent of SRMS (in terms of both frequency and intensity) from a larger pool of chronically epileptic animals for transplantation studies.
The extent of SRMS can vary between epileptic animals (Rao et al., 2006, 2007; Waldau et al., 2010). Choosing animals exhibiting a similar extent of SRMS will facilitate the comparison of changes in the seizure frequency and intensity between different groups such as CERs receiving NSC grafts, CERs receiving sham-grafting surgery and CERs receiving neither grafts nor surgery.
ANALYZING COGNITIVE FUNCTION IN CERS CHOSEN FOR GRAFTING STUDIES
In order to facilitate the assessment of improvement in hippocampal-dependent cognitive function with NSC grafting, it is important to examine the extent of cognitive dysfunction prior to NSC grafting in the chosen CERs. While one can assess hippocampal-dependent cognitive function using quite a few tests, we have selected a water maze test (WMT) as an example for assessing spatial learning and memory function in this article. The different aspects of this test are described below.
Equipment
The water maze tank is a circular blue fiberglass pool measuring 170 cm in diameter and 75 cm in height. Pool is filled with the room temperature water to 35 cm height and extra-maze cues need to be placed on the walls of the room. The animals are trained to find the platform submerged in the water in one of the 4 quadrants using spatial cues (Morris 1984). The movement of the rat in the water maze tank is continuously video-tracked and recorded using a computerized video-tracking system. In our laboratory, we have been using the ANY-maze video-tracking system for water maze studies.
Learning paradigm
Although several different training paradigms can be used, we chose the one that is considered ideal for assessing the learning and memory function rats exhibiting hippocampal injury (Moser and Moser, 1998). This paradigm comprises 11 sessions of learning over 6 days (one session on day 1, and 2 sessions on each of the days 2–6. Each rat gets 4 trials in every learning session and each trial lasts 90 seconds with an inter trial interval of 120 seconds. The location of the hidden platform remains constant across all trials and days in this paradigm.
Steps in the learning task
Release the rat into the water maze tank facing the wall of the pool in a predetermined pseudo-random fashion so that each trial commences from a different start location. Allow 90 seconds to explore the platform using spatial cues that are placed on walls around the water maze tank.
If the rat finds the platform submerged in water within the ceiling time of 90 seconds, allow the rat to stay on the platform for 30 seconds to explore the spatial cues. If the rat fails to find the platform within the ceiling period, guide the rat to the platform and allow it to stay there for 30 seconds. Note: In between trials, place the wet rat in an empty cage placed on a heating pad maintained at 37°C. At the end of the fourth trial, wipe the rat’s body thoroughly with dry towels, dry off the fur using a hair dryer and place them back in the home cage.
The video-tracking system (such as the ANY-maze program used in our studies) will document both the latency to reach the platform and the path length in each trial for every rat. Using these parameters, calculate the mean ± S.E.M. for different parameters such as the latency to reach the platform (in seconds), the swim path length and the swim speed for every learning session in different groups of rats. Compare the latency to reach the platform across the eleven learning sessions to assess the learning curve using regression analysis in each group. If swim speed is significantly variable between individual rats within a group or between groups, select the swim path length for assessing the learning curves in different groups. Measure the extent of improvement in the learning over the 11 learning sessions in each group by comparing the values (either the latencies to reach the platform or swim path lengths) between the 1st and 11th learning sessions.
Probe test (or memory retrieval test)
While memory retrieval test can be given at different intervals after the completion of learning sessions, we chose to conduct this test after a delay of 24 hrs, as control animals (i.e. animals without any hippocampal injury) exhibit robust memory retrieval ability at this time-point after 11 sessions of learning. For assessing memory retrieval, subject each rat to a single 45-second trial in the absence of the submerged platform. Release the rat from a quadrant that is opposite to the original position of the submerged platform. Using the video-tracking program (such as ANY-maze used in our laboratory), measure parameters such as the latency to reach the platform area, time spent in platform area, number of platform area crossings, and the dwell time in the platform quadrant.
Interpretation of probe test results
Typically, rats that have the ability to retrieve the learned memory head straight to the platform area after their release, spend most of the trial time searching for the platform in the quadrant (or area) where the platform was originally placed, and exhibit many platform area crossings. Notes: Rats that exhibit shorter latencies to reach the platform area after the release, greater dwell time in the platform area and/or quadrant, greater number of crossings in the platform area are considered to have superior spatial memory function. In contrast, rats exhibiting longer latencies to reach the platform area (or unable to reach the platform area), significantly reduced or no dwell time in the platform area, and fewer or no platform area crossings are considered to have impaired spatial memory function. Virtually all rats exhibiting chronic TLE display impaired spatial learning and memory function in this WMT (Waldau et al., 2010). A comparison of learning and memory function in a WMT between age-matched naïve control rats and chronically epileptic rats is illustrated in Fig. 3.
Figure 3.
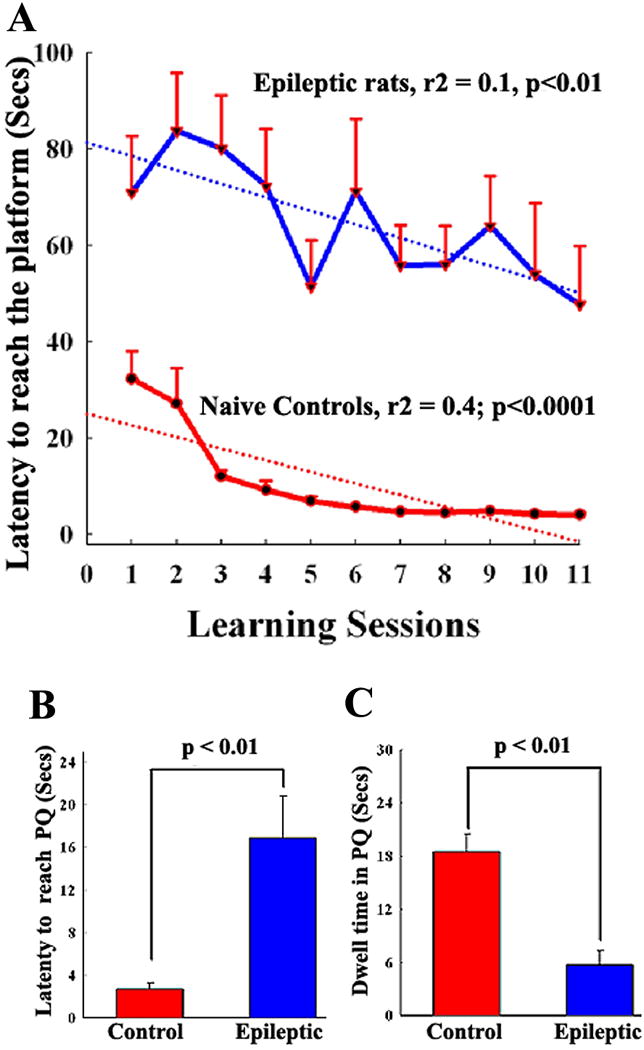
Comparison of the spatial learning ability of chronically epileptic rats at 4-months after status epilepticus (SE) with age-matched naive rats using a WMT. Note that, in comparison to intact rats, the average latency (in seconds) to reach the platform were much greater in rats exhibiting chronic epilepsy at all training sessions (A). Naïve rats learn quickly to locate the hidden platform using spatial cues. Epileptic rats are clearly slow learners and overnight forgetting is common in this group. However, these rats are able to find the platform within the ceiling time (90 sec) in most learning sessions. Thus, learning impairment in chronically epileptic rats is revealed mainly by the greater latency values to reach the platform in all training sessions in comparison to naïve rats. B and C show the results of a probe test conducted one-day after the learning paradigm. Naïve rats appear to exhibit robust memory retention, as they reach the platform quadrant faster (B) and spend most of their probe test time searching for the platform in the platform quadrant (C). In contrast, chronically epileptic rats explore all quadrants of the maze and hence their dwell time in the platform quadrant is much less than that of naïve rats (C). Thus, chronically epileptic rats exhibit memory dysfunction.
HARVESTING & PREPARATION OF MGE-NSC SUSPENSION FOR GRAFTING
Required Culture media, Equipment and Supplies
Proliferation medium (100ml)
Dulbecco’s Modified Eagle Medium (DMEM; GIBCO Cat#11960-044): 74.520ml
F-12 nutrient mixture (GIBCO; (Cat#11765-054): 24.4ml
B-27 without retinoic acid (GIBCO Cat#12587-010): 1ml
Antibiotic-Antimycotic 100X; (GIBCO; Cat#15240-062):80μl
FGF-2 and EGF (Peprotech: Cat#AF-100-18B and AF-100-15): 20ng/ml
Heparin: (Sigma cat#H1027; Prepared in sterile DMEM): 5μM
Differentiation medium (50ml)
Neurobasal Medium (GIBCO Cat#21103-049): 48.335 ml
Antibiotic-Antimycotic 100X; (GIBCO; Cat#15240-062):40μl
B-27 with retinoic acid (GIBCO; Cat#17504-044): 1ml
L-Glutamine: (GIBCO; Cat#25030-081): 625 μl
Equipment and supplies
Dissection microscope
CO2 incubator for cell culture
Centrifuge
Fine surgical instruments for microdissection of the desired region (e.g. MGE) from the embryonic brain. Note: All surgical instruments need to be autoclaved before use.
Sterile media filtration units
Sterile 25 cm2 canted culture flasks
Sterile 10 ml Serological pipettes and stripettor
Sterilized fire-polished Pasteur pipettes
Sterile centrifuge tubes (15 ml and 50 ml capacity)
Different sizes of sterile cell culture dishes (35 mm, 60 mm and 100 mm in diameter)
Sterile cell filters (40 μm pore size)
Hemocytometer for measuring viable cell numbers in a suspension of NSCs
Trypan Blue stain (0.4%) for assessing the viability of cells in a suspension of NSCs
5′-bromodeoxyuridine (BrdU)
Poly-D-Lysine coated 8 well culture slides or 35 mm culture dishes.
Dissection of MGE tissues and plating of MGE cells for expansion of NSCs
-
1
Deeply anesthetize the timed pregnant rat (Gestation day 14) using isoflurane and remove fetuses via cesarean section. Collect them in a sterile culture medium containing Dulbecco’s modified Eagle medium (DMEM) and F12 (Invitrogen).
-
2
Dissect the brain from each fetus and carefully isolate the two cerebral hemispheres from the brain stem using fine micro-scissors under a dissection microscope.
-
3
From each hemisphere, expose the caudal, medial and lateral ganglionic eminences located in the basal forebrain by making a cut through the supero-lateral wall of the primitive cerebral cortex (or the cortical plate). Identify the MGE and carefully scoop out the MGE tissue using fine micro-scissors leaving behind both caudal and lateral ganglionic eminences.
-
4
Mechanically dissociate the collected MGE tissues in the proliferation medium using a fire-polished sterile Pasteur pipette and obtain a single cell suspension.
-
5
Wash the cell suspension twice via reconstitution in a fresh proliferation medium and centrifugation at 800G for 8 minutes. Reconstitute the final pellet in 1 ml of the proliferation medium and assess the viability of MGE cells in the suspension using the trypan blue exclusion test. For this, mix 17μl of the proliferation medium with 10 μl of the trypan blue and 3 μl of the cell suspension, charge 10 μl of this cocktail into each side of the cell counting chamber and count both live cells (unstained cells) and dead cells (cells stained with trypan blue) to estimate both the overall viability of cells in the suspension as well as the total number of viable cells in the suspension.
-
6
Adjust the density of viable cells to 300,000 live cells per 10 ml of the proliferation medium, plate the medium into 25cm2 canted tissue culture flasks (10 ml/flask), and place them in the cell culture incubator maintaining a temperature of 37°C and 5% CO2.
-
7
One day after the plating of cells, label the proliferating cells via addition of 5′bromodeoxyuridine (BrdU; Sigma-Aldrich) to the medium at a final concentration of 2.5 μM. Allow 5–6 days for the neurospheres to grow.
-
8
On day 6 or 7, transfer the media containing NSC-derived neurospheres from flasks into 15 ml centrifuge tubes and let the neurospheres to settle down. Remove the supernatant media in each tube, pool neurospheres from all tubes in a single 15 ml centrifuge tube, and gently triturate using fire polished Pasteur pipette until a clear cell suspension is obtained.
-
9
Wash triturated cells thrice via reconstitution in a fresh proliferation medium and centrifugation to get rid most of the dead cells in the suspension. Reconstitute the final pellet in a 30–50 μl of the differentiation medium and count the viable and dead cells using a trypan blue exclusion test. Adjust the density of viable cells to 80,000–100,000 cells/μl. Note: If viability is <75%, wash cells again to obtain the higher ratio of viable cells in the final cell suspension. Typically, the cell suspension exhibiting >80% viability is ideal for transplantation studies.
-
10
Use an aliquot of the final cell suspension to determine the BrdU labeling index at the time of grafting. For this, plate cells in poly-D-Lysine coated culture slides/dishes, incubate in the differentiation medium for an hr, fix in 4% paraformaldehyde, perform BrdU immunofluorescence, counter stain with DAPI, and calculate the percentage of cells expressing the BrdU. Typically, expanding neurospheres in a medium containing 2.5 μM of BrdU results in labeling of over 90% of neurosphere cells.
Characterization of the Differentiation Potential of NSCs in Vitro
In order to ascertain the differentiation potential of the chosen donor NSCs, it will be useful to examine their differentiation in culture. The protocols we typically use are described below.
-
11
Plate samples of the NSC cell suspension (200,000 cells/dish) into poly-D-lysine-coated 35 mm culture dishes containing the differentiation medium.
-
12
Incubate cells for 6–8 days at 37°C with 5% CO2.
-
13
Terminate cultures by treating with 2% paraformaldehyde for 20 minutes. If GABA immunostaining is planned, fix cultures in 2% paraformaldehyde containing 0.01% glutaraldehyde. Following the above fixation, thoroughly wash cultures with 0.1 M PBS.
-
14
Process cultures for single or dual immunofluorescence for identifying: (i) neurons expressing beta-III tubulin (TuJ-1); (ii) astrocytes expressing the glial fibrillary acidic protein (GFAP); (iii) oligodendrocytes expressing the protein O1; and (iv) inhibitory interneurons expressing the GABA. Table 1 illustrates the combination of primary and secondary antibodies that we typically use in our dual immunofluorescence studies.
-
15
Counter stain the immunostained cultures with DAPI (1:200) to facilitate the visualization of all nuclei with blue fluorescence.
-
16
Using a systematic random sampling scheme, obtain digital illustrations of dual/triple labeled samples (at least 6 fields per each of the four independent samples) using an inverted fluorescence or confocal microscope. Count the total number of cells (i.e. all DAPI+ blue nuclei) as well as cells that are positive for a neural cell marker to measure the percentages of cells expressing different neural markers. Figure 4 illustrates neural phenotypes derived from MGE-NSCs following 4 and 8 days of incubation in a differentiation medium.
Table 2D.7.1.
Combinations of Various Primary and Secondary Antibodies Used in Single and Dual Immunofluorescence Staininga
| Name | Blocking serum | Primary antibody, sources, and dilution | Secondary antibodyb |
|---|---|---|---|
| BrdU staining | NHS | Mouse anti-BrdU (BD Bioscience), 1:200 | Biotinylated anti–mouse IgG (Vector Labs) followed by ABC reagent and DAB |
| DCX staining | NHS | Goat anti-DCX (Santa Cruz Biotechnology), 1:200 | Biotinylated anti–goat IgG (Vector Labs) followed by ABC reagent and VG |
| BrdU-NeuN dual immunofluorescence | NGS and NDS | Rat anti-BrdU (Serotec), 1:200 Mouse anti-NeuN (Millipore), 1:1000 |
Goat anti–rat IgG with Alexa Fluor 594 Donkey anti–mouse IgG with Alexa Fluor 488 |
| BrdU-GABA dual immunofluorescence | NGS | Rat anti-BrdU (Serotec), 1:200 Rabbit anti-GABA (Sigma), 1:5000 |
Goat anti–rat IgG with Alexa Fluor 594 Goat anti–rabbit IgG with Alexa Fluor 488 |
| BrdU-GFAP dual immunofluorescence | NGS | Rat anti-BrdU (Serotec), 1:200 Rabbit anti-GFAP (Sigma), 1:1000 |
Goat anti–rat IgG with Alexa Fluor 594 Goat anti–rabbit IgG with Alexa Fluor 488 |
| BrdU-s100β dual immunofluorescence | NGS | Rat anti-BrdU (Serotec), 1:200 Rabbit anti-s100β (Millipore), 1:1000 |
Goat anti–rat IgG with Alexa Fluor 594 Goat anti–rabbit IgG with Alexa Fluor 488 |
| BrdU-O1 dual immunofluorescence | NDS | Rat anti-BrdU (Serotec), 1:200 Mouse anti-O1 (Millipore), 1:1000 |
Goat anti–rat IgG with Alexa Fluor 594 Donkey anti–mouse IgG with Alexa Fluor 488 |
| BrdU-NG2 dual immunofluorescence | NGS | Rat anti-BrdU (Serotec), 1:200 Rabbit anti-NG2 (Millipore), 1:1000 |
Goat anti–rat IgG with Alexa Fluor 594 Goat anti–rabbit IgG with Alexa Fluor 488 |
| BrdU-Sox2 dual immunofluorescence | NGS | Rat anti-BrdU (Serotec), 1:200 Rabbit anti-Sox2 (Millipore), 1:1000 |
Goat anti–rat IgG with Alexa Fluor 594 Goat anti–rabbit IgG with Alexa Fluor 488 |
| Tuj1-GABA dual immunofluorescence | NGS | Mouse anti-Tuj1 (Covance), 1:1000 Rabbit anti-GABA (Millipore), 1:1000 |
Goat anti–rat IgG with Alexa Fluor 594 Goat anti–rabbit IgG with Alexa Fluor 488 |
| Tuj1-GFAP dual immunofluorescence | NGS | Mouse anti-Tuj1 (Covance), 1:1000 Rabbit anti-GFAP (Millipore), 1:1000 |
Goat anti–rat IgG with Alexa Fluor 594 Goat anti–rabbit IgG with Alexa Fluor 488 |
| O1-S100β dual immunofluorescence | NGS | Mouse anti-O1 (R&D), 1:500 Rabbit anti-S100β (Millipore), 1:1000 | Goat anti–rat IgG with Alexa Fluor 594 Goat anti–rabbit IgG with Alexa Fluor 488 |
| S100β-GDNF dual immunofluorescence | NGS | Mouse anti-S100β (Millipore), 1:1000 Rabbit anti-GDNF (Santa Cruz Biotechnology), 1:1000 |
Goat anti–rat IgG with Alexa Fluor 594 Goat anti–rabbit IgG with Alexa Fluor 488 |
Abbreviations: NHS, normal horse serum; NGS, normal goat serum; NDS normal donkey serum.
Secondary antibodies purchased from Invitrogen were used at 1:200 dilution in all staining described in this unit.
Figure 4.
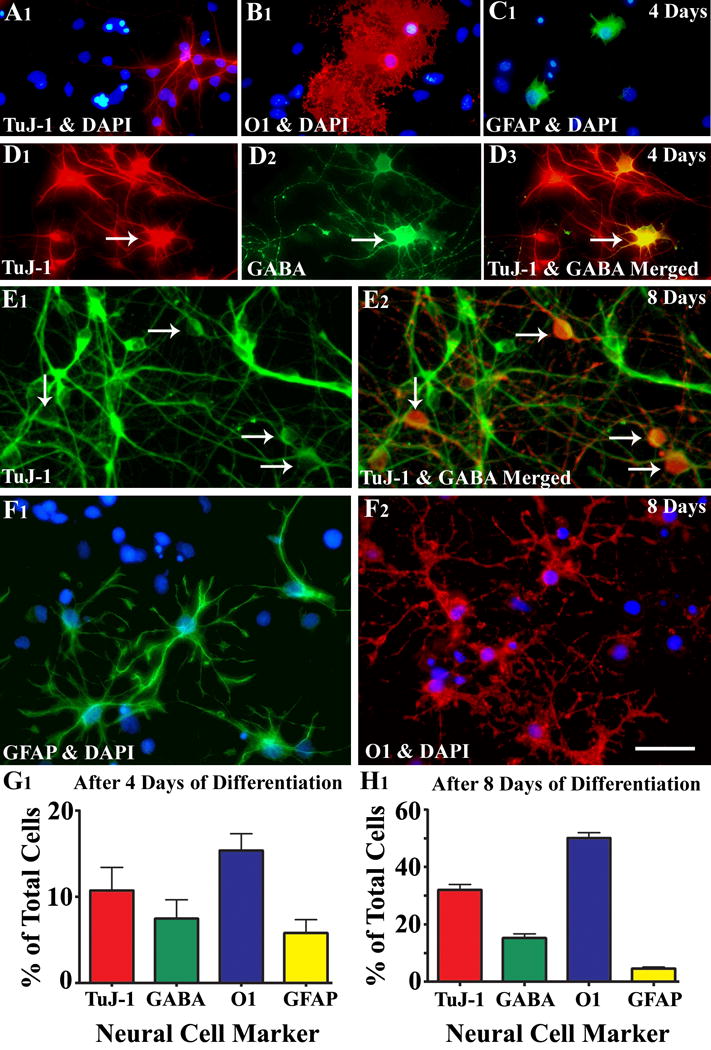
Differentiation of medial ganglionic eminence-neural stem cells (MGE-NSCs) after their dissociation from CldU-labeled neurospheres and incubation in the differentiation medium for 4 days (A1-D3) or eight days (E1-F2). Differentiation of fractions of MGE-NSCs into TuJ-1+ neurons (A1, E1), O1+ oligodendrocytes (B1, F2), and GFAP+ astrocytes (C1, F1) could be seen at both time-points. Furthermore, fractions of MGE-NSCs also differentiate into GABA-ergic neurons (arrows in D1-D3 and E1-E2). Scale bar: 50 μm. The bar charts (G1, H1) illustrate percentages of MGE-NSCs that exhibit differentiation into TuJ-1+ neurons, GABA+ neurons, O1+ oligodendrocytes and GFAP+ astrocytes after incubation in the differentiation medium for 4 days (G1) or 8 days (H1). Note that the expression of GABA among TuJ-1+ neuronal population is ~70% after 4 days and ~50% after 8 days of incubation in the differentiation medium. [Reproduced from: Waldau et al., Stem Cells, 28:1153–1164].
GRAFTING OF NSCs INTO THE HIPPOCAMPI OF CERs
Prior to the surgery, choose CERs exhibiting a similar extent of SRMS as well as cognitive dysfunction. Choosing rats displaying the above characteristics of TLE would facilitate testing of the efficacy of NSC grafts for both restraining seizures and reversing the cognitive dysfunction.
Equipment, Materials and Supplies
Stereotaxic equipment: Any stereotaxic device made for rat neurosurgery can be used. In our laboratory, we use digital stereotaxic equipment purchased from MyNeurolab.
Heating pads to maintain the body temperature of the rat during and after the surgery
Hand or foot controlled electrical driller with different sizes of drill bits to make small burr holes in the skull using the desired stereotaxic coordinates.
Glass bead sterilizer to sterilize the stainless steel surgical instruments during the surgery of multiple animals in one surgery session.
Surgical shaver to remove the rat’s fur from the head.
Sterile 10μl syringes with needle for loading the stem cell suspension. Our laboratory utilizes 10μl Hamilton syringes.
Sterile scalpel blades and sterile applicators with cotton tips.
Autoclaved surgical instruments: This comprises the scalpel blade holders, skin retractors, blunt forceps of different sizes, autoclipping device loaded with 9 mm wound clips (for stapling the skin incision after the grafting surgery) and autoclip remover.
Anesthetics that are approved by the IACUC for the survival rodent surgery: In our laboratory, we use an anesthetic cocktail comprising ketamine (50 mg/ml), xylazine (4.5 mg/ml) and acepromazine (0.4 mg/ml) at a dose of 0.7ml/kg bw.
Ice bucket filled with ice for storing the NSC suspension and anesthesia during the grafting session.
Betadine, isopropyl alcohol pads, 3% hydrogen peroxide, and bone wax for sealing the burr holes after the surgery.
Other drugs and fluids: Artificial tear for application into eyes during the surgery, bupivicaine for application into the sutured skin after the surgery for relieving pain, buprenorphine for post-operative pain management, and sterile saline for subcutaneous administration after the surgery.
Personnel protective equipment (PPE): sterile surgical latex gloves, disposable lab coats, head cover, booties, and surgical masks.
Grafting Procedure
Prepare the aseptic surgical area, arrange all autoclaved surgical instruments and other required supplies around the stereotaxic device, and brightly illuminate the area of the stereotaxic device that holds the head of the rat.
Intramuscularly inject the anesthetic cocktail to the rat chosen for transplantation surgery at a dose of 0.7 ml/Kg bw.
Confirm the depth of anesthesia at 5–10 minutes after the administration of anesthesia via tail and toe pinches. Following confirmation of deep anesthesia, remove the fur from rat’s head using an electrical shaver in a corner of the room that is at a reasonable distance from the surgery table. Wipe off the cut hair from the shaved area with 70% alcohol and carefully drape the rat with a sterile draper exposing only the head area.
Fix the rat’s head securely into the stereotaxic device using ear bars etc.
Wipe the surgical site sequentially with betadine, isopropyl alcohol and betadine.
Make a midline incision on the rat’s head using a sterile scalpel blade, retract the skin flaps with surgical skin retractors, clean the fascia on top of the cranium using 3% hydrogen peroxide and identify the anatomical landmarks on the cranium such as the bregma (meeting point of the frontal bone and the two parietal bones) and the lambda (meeting point of the two parietal bones and the occipital bone).
-
Using the stereotaxic device and coordinates and the bregma as a reference point, mark the chosen grafting sites on the skull. In our NSC grafting studies in CERs, we place NSC grafts into 4 sites in each hippocampus using the following coordinates. (i) antero-posterior (AP) – 3 mm posterior to the bregma, lateral (L) – 1.8 mm lateral to the midline, ventral (V) – 3.5 mm from the brain surface; (ii) AP – 3.6 mm, L – 2.5 mm, V – 3.5 mm; (iii) AP – 4.2 mm, L – 3.2 mm, V – 3.5 mm; (iv) AP – 4.8 mm, L – 4 mm, V – 3.5 mm).
These coordinates were chosen using the rat brain atlas (George Paxinos and Charles Watson; fourth edition) to particularly place the grafts at the end of the hippocampal fissure (i.e. just above the CA3 pyramidal cell layer and the adjoining the lateral ventricle) in CERs. If placement in other areas of the hippocampus are desired (such as CA1 pyramidal cell layer), adjust the coordinates using the rat brain atlas. Furthermore, as hippocampal shrinkage can vary between different models, determine the appropriate coordinates for the desired location empirically using injections of the dye in a set of pilot studies.
-
Load a small volume of the NSC suspension (3–4 μl) into a 10 μl Hamilton syringe, fix it to the stereotaxic instrument, and make sure that no air bubble is trapped between the needle and the cell suspension by ejecting out ~1 μl of the cell suspension.
Load fresh NSC suspension for each injection to minimize inconsistency between grafts placed at different injection sites in a rat or between rats in a group.
Injection of the NSC suspension into the hippocampus: Lower the Hamilton syringe needle first to the surface of the brain through the first burr hole. Then, using the ventral stereotaxic coordinate, gently lower the needle through the meninges and cortex and reach the desired depth and dispense 1.0μl of the NSC suspension in spurts of ~0.2µl/min over a period of 5 minutes.
-
After injecting each spurt of 0.2μl of the NSC suspension, gently withdraw the needle by ~0.05 mm to create room for the injected NSC suspension.
This enhances the probability of placing the graft in the chosen location and minimizes the flow of injected cells mostly into the adjoining lateral ventricle.
-
After dispensing 1.0μl volume of the NSC suspension, allow the needle to stay in place for another 10 minutes and then slowly withdraw the needle.
This ensures that the injected cells disperse in the hippocampus and minimizes the backflow of the injected fluid along the needle track into the corpus callosum area or the overlying cortex.
Repeat the above procedure for each of the remaining 7 graft sites (i.e. 4 sites/hippocampus in every CER).
Following NSC graft injections to all sites, close the top of the burr holes with a small amount of the bone wax, staple the skin flaps using a autoclipping device loaded with 9 mm wound clips, apply bupivicaine solution on the stapled skin. Betadine may also be applied at the wound site.
-
For post-operative pain management, intramuscularly administer buprenorphine (at 0.1 mg/kg bw every 12 hrs for 48 hours). To prevent dehydration after the surgery, subcutaneously administer 5 ml of the sterile saline to each rat and place them back in the home cage that was kept warm on a heating pad maintained at 37°C. The cages need to be kept on a heating pad until the rat recovers from the anesthesia.
The CERs typically recover from the grafting surgery within 2–3 days. It is important to carefully observe these rats during this period and ensure that they eat and drink well. If they are not actively eating or drinking, it will be necessary to subcutaneously administer saline (5 ml/day) and place soft rat chow and/or transgel inside the cage.
Animals chosen for the sham-grafting surgery need to undergo a surgical procedure that is identical to what is described above for the transplantation of MGE-NSCs. The only difference is that these animals will receive a sterile culture medium (1μl/site) in place of the cell suspension.
ANALYSES OF NSC GRAFTING—MEDIATED CHANGES IN SEIZURES
Measurement of SRMS
Measure changes in the extent of SRMS rigorously beginning from a week after the transplantation surgery continuing for at least 3 months after the grafting/sham grafting procedure.
Quantification of SRMS after grafting can be done as described earlier for quantification of the pre-grafting SRMS (i.e. a minimum of 32 hrs of observation/month in 8 intermittent sessions or continuous (24/7) video monitoring). Notes: Monitoring of SRMS for a longer duration after the grafting (i.e. for 6–8 months post-grafting) will be helpful to determine whether any beneficial effects observed on seizure control in CERs at early time-points after the grafting persist at extended periods after the grafting or are just transient.
From the above measurement of SRMS, calculate the average frequencies of all SRMS and stage V seizures, the average duration of individual SRMS, and the total time spent in seizures for every month.
Compare the post-grafting seizure scores (for different months post-grafting) with the pre-grafting seizure scores using repeated measures ANOVA and ascertain the extent of seizure suppression mediated by NSC grafting. Fig. 5 illustrates the efficacy of MGE-NSC grafts for restraining spontaneous recurrent motor seizures (SRMS) in chronically epileptic rats. For CERs undergoing the sham-grafting surgery, compare the post-sham grafting seizure scores with the pre-sham grafting seizure scores using repeated measures ANOVA and ascertain the sham-grafting mediated changes in the SRMS. Notes: Different parameters of SRMS can also be compared statistically across the three age-matched groups (CERs receiving NSC grafts, CERs receiving sham-grafting surgery, and CERs receiving neither grafts nor surgery) using one-way ANOVA, if all three groups of rats had similar extent of SRMS at the time of their classification into different groups (i.e. prior to the grafting procedure).
Figure 5.
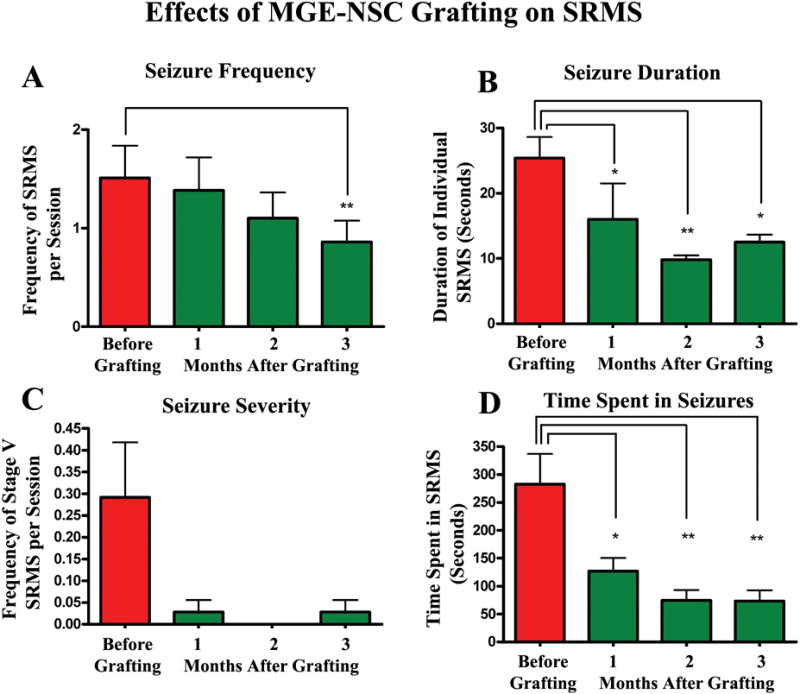
Efficacy of MGE-NSC grafts for restraining spontaneous recurrent motor seizures (SRMS) in chronically epileptic rats. The Y-axis in bar charts A and C denotes the average numbers of seizures per session (4-hr block) of observation. Note that MGE-NSC grafting considerably decreases the seizure frequency (A), the duration of individual seizures (B), the severity of seizures (C) and the total time spent in seizures (D). * = p < 0.05; ** = p <0.01. [Reproduced from: Waldau et al., Stem Cells, 28:1153–1164].
Measurement of Spontaneous Recurrent Seizures (SRS) via Electroencephalographic (EEG) Recordings
In addition to the quantification of SRMS as described above, one can also quantify the long-term changes in all SRS (i.e. electrographic seizures with or without a behavioral component) after the grafting using continuous electroencephalographic recordings at an extended time-point after grafting. Note: It is ideal to do EEG recordings following completion of all behavioral measures of SRMS and cognitive function.
In our laboratory, we use a time-locked video-digital EEG monitoring system (AS40 from Grass Telefactor) for measuring SRS from CERs.
For EEG recordings from CERs (with or without grafts), we implant sterile metal EEG recording electrodes with mounting screws (Plastics one) epidurally, one over the right fronto-parietal cortex for recording EEG from the cortex, and another over the left cerebellum as a reference electrode. To record EEG directly from the hippocampus, an intracranial stainless steel (Teflon coated except the tip) electrode with socket (Plastics One) is also placed into the right DG (Rao et al., 2006). We also implant a couple of anchoring screws on the skull to secure the EEG electrodes with dental cement. The screws and electrodes are cemented in place, and electrode leads are attached to a microplug, which is then cemented to the animal’s head. The above implantation procedures are done in one surgery session (as detailed in Rao et al., 2006). In grafted studies, this is typically done at ~2–3 months after the grafting.
Two weeks after the implantation surgery, each rat is placed in a Plexiglas cage, the connector cable of the video-EEG system is fixed into the electrode pedestal on the rat’s head. The video-EEG system monitors simultaneously occurring behavior and EEG activity in awake, freely behaving rats with ad libitum access to food & water.
In our laboratory, the EEG recordings are done with low frequency filter (LF) set at 0.3Hz, high frequency (HF) set at 35 Hz, and data rate at 200 Hz. Furthermore, Szac Detector component of AS40 is turned on to quantify the numbers of all high amplitude spikes (first half-wave amplitude ratio set at ≥1.5 and 2nd half-wave amplitude ratio set at ≥2.5), and seizure events (high frequency multispike complexes [at 3–20 Hz & 50% faster than background] and/or high voltage synchronized spike or wave activity (≥45% variation in amplitude, ≥36% variation in duration, amplitude ratio of 3, and lasting for ≥ 6 seconds).
Both frequency & severity of EEG seizures with or without behavioral manifestation is continuously measured for a period of time. An “EEG seizure with behavior” is a generalized electrographic seizure with accompanying motor seizure activity (e.g. unilateral or bilateral forelimb clonus, and rearing and falling [Stages III–V seizures). The various parameters for comparison across the three age-matched groups (CERs receiving NSC grafts, CERs receiving sham-grafting surgery, and CERs receiving neither grafts nor surgery) using one-way ANOVA the following. (1) Frequency of all SRS. (2) EEG-SRS with behavior. (3) EEG-SRS without behavior (electrographic SRS). (4) Average duration of individual SRS. (5) The total time spent in SRS. (6) Numbers of high-amplitude spikes. Notes: The EEG studies are labor intensive and clearly need personnel with expertise for appropriate implantation of electrodes, ability for connecting the electrodes on the rat’s head to the video-EEG system through a connector cable without anesthetizing the animal. One pitfall of this procedure is that the number of rats initially implanted with electrodes may be reduced at the time of recordings due to malfunction of electrodes. Alternatively, electrodes may fall off due to accidents in the cage. To circumvent these difficulties, it is important to include a higher number of CERs than required for statistics in each group. Furthermore, it will be necessary to single house rats in a cage with flat tops, which minimizes the loss/malfunction of electrodes due to accidents in the cage.
ANALYSES OF THE NSC GRAFTING MEDIATED EFFECTS ON COGNITIVE FUNCTION
Since CERs that received NSC grafts or sham-grafting surgery would undergone a WMT prior to the grafting or sham-grafting surgery (as described in an earlier section of this article), their pre-grafting/pre-sham grafting spatial learning and memory scores are available.
In order to assess the changes in the spatial learning and memory function with NSC grafting or sham grafting surgery, a second WMT is typically given at 2–4 months after the grafting.
The entire learning paradigm and the protocol for memory retrieval test will be similar to that described earlier for the pre-grafting WMT. However, it is important to change the location of submerged platform to a quadrant which is different from the one used for the pre-grafting WMT.
Following the completion of the second WMT, the pre-transplant and post-transplant scores for the extent of learning & memory function are compared within the grafted group using a two-tailed, unpaired t-test. The parameters include: (i) average latencies to reach the platform in different learning sessions; (ii) the latency to reach the platform area, dwell time in the platform area and the platform quadrant, and platform area crossings in the memory retention (Probe) test. These analyses are efficient for ascertaining whether or not grafting of NSCs improves cognitive function, in comparison to other control groups (CERs receiving sham-grafting surgery and CERs receiving neither grafts nor surgery).
ANALYSES OF THE YIELD & DIFFERENTIATION OF NSC-GRAFT DERIVED CELLS
Following the completion of all behavioral measures described above, animals are perfused with 4% paraformaldehyde for analyses of the yield and differentiation of NSC-graft derived cells. This comprises tissue fixation, tissue processing for cryostat sectioning, immunostaining and/or immunofluorescence procedures, quantification of the yield of graft-derived cells using stereology, and measurement of the percentages of the graft-derived cells that differentiate into NeuN+ neurons, GABA-ergic interneurons and subclasses of GABA-ergic neurons expressing various markers, astrocytes, astrocytes expressing neurotrophic factors such as the GDNF, oligodendrocyte progenitors and mature oligodendrocytes.
Required Equipment, Materials and Reagents
Perfusion Pump: Our laboratory uses a perfusion pump purchased from MyNeurolab.
Cryostat: Our laboratory uses a cryocut purchased from Leica.
Refrigerator and Freezer for storage of free-floating cryostat sections.
Light and Fluorescent Microscopes and a Confocal Microscope: Our laboratory uses microscopes purchased from Nikon and Olympus.
Imaging system for stereological counting of graft-derived cells: Our laboratory utilizes the StereoInvestigator system purchased from MicroBrightfield.
Dissection instruments to open the thoracic cavity and to remove the perfusion-fixed brain from the skull.
Smaller bottles for storing brain tissues.
Solutions such as 4% paraformaldehyde for perfusion fixation, 0.9% saline, 0.1M phosphate buffer (PB) with 0.01% sodium azide (storage buffer), 10, 20 and 30% sucrose solutions; 0.01% Triton X-100 (for increasing tissue permeability during immunostaining).
Twenty-four well plates for storing the cut tissue sections and for the immunostaining and immunofluorescence procedure.
Anti-freeze buffer for long term storage of tissue sections. Immunostaining buffers (phosphate buffered saline at pH 7.4 and Tris-buffered saline at pH-7.5).
Normal goat serum, normal horse serum, and normal rabbit serum for the blocking step in the immunostaining procedure.
Multiple primary and secondary antibodies (see Table 1 for the source); Avidin-biotin-complex (ABC), diaminobenzidine (DAB), Vector gray kits purchased form Vector Labs.
Artist’s brushes for transferring sections during the free-floating immunostaining procedure and mounting sections on the slides. Glass slides, cover slips, and a mounting medium such as permount and slow-fade-antifade reagent (Molecular Probes).
Various grades of alcohol (50%, 70%, 95% 100%), xylene, distilled water, hematoxylin, neutral red and DAPI for counterstaining immunostained sections.
Tissue Harvesting and Section Cutting
-
1
Deeply anaesthetize rats using a lethal dose of isoflurane, secure all four limbs on a flat fixation board using thumbnails.
-
2
Cut open the thoracic cavity, open the pericardium, turn on the perfusion pump and carefully insert a 20–25G syringe needle (attached to a long plastic perfusion tube immersed in 200 ml of normal saline solution containing 1% heparin) into the left ventricle. Immediately following this, make a nick in the right atrium for the blood and the perfusion fluid to flow out of the heart and adjust the speed of the perfusion pump so that 200 ml of saline perfused in ~10–12 minutes.
-
3
Immerse the perfusion tube in a 4% paraformaldehyde solution and perfuse ~500 ml of 4% paraformaldehyde in 30–40 minutes.
-
4
Cut open the skull using bone cutters and blunt seizures, carefully remove the entire brain from the skull by cutting the meninges and cranial nerves, immerse the brain in 4% paraformaldehyde solution overnight at 4°C, and transfer it to 0.1M PB solution.
-
5
Detach the hindbrain and process the rest of the brain for cryo-protection by successively treating the brain tissue with different percentages (10, 20 and 30%) of the sucrose solution at 4°C.
-
6
Fix the brain tissue to the cryostat chuck using the cryo-embedding gel and let it stay for ~30 minutes. Following this, fix the cryostat chuck containing the frozen brain tissue into the object holder of the cryostat, adjust the cryostat knife settings and section thickness to desired values and cut coronal sections through the brain. Notes: We routinely collect 30 μm-thick serial coronal sections through the entire hippocampus in 24 well plates filled with PB containing 0.01% sodium azide. For long term storage, we store sections in the antifreeze solution at −20°C.
BrdU Immunostaining
-
7
Choose every 10th section through the entire hippocampus for analyses of the yield of graft-derived cells. Process these serial sections for BrdU immunostaining in 24 well plates.
-
8
Etching: Treat sections with 0.1 M Tris-buffered saline (TBS) containing 20% methanol and 3% hydrogen peroxide to remove the endogenous peroxidase.
-
9
Wash sections thoroughly in the TBS, incubate sections in a 50% formamide solution prepared in 2X saline sodium citrate buffer (2X SSC buffer) for 2 hrs at 65°C using an oven.
-
10
Wash sections in a cold 2X SSC buffer twice and incubate in 2N HCl at 37 °C for 60 minutes using an oven. Following this, treat sections with a 0.1M borate buffer (pH = 8.5) and wash 3 times in TBS.
-
11
Incubate sections in a blocking TBS solution comprising 10% normal horse serum (NHS) and 0.01% TritonX-100 for 30 minutes at room temperature.
-
12
Incubate sections overnight at 4 °C in the primary antibody solution comprising the mouse anti-BrdU (BD Bioscience, 1:500), 3% NHS, and 0.01% TritonX-100.
-
13
Wash sections thrice in the TBS and incubate for an hr in a secondary antibody solution comprising the biotinylated anti Mouse IgG (Vector Labs, 1:200) and 2% normal horse serum.
-
14
Rinse sections thrice in TBS and incubate for an hour in the ABC reagent (Elite ABC kit, Vector Labs).
-
15
Wash sections thrice in TBS and then incubate for 5–10 minutes in a DAB solution prepared as per manufacturer’s instructions (DAB kit, Vector Labs). Give three washes in TBS before mounting on slides for dehydration, clearing and cover slipping using standard histological methods.
It is important to standardize the incubation time required for an optimal color reaction using a couple of sections. Terminate the DAB reaction by transferring sections into the distilled water.
-
16
An example of the distribution of BrdU+ NSC graft-derived cells in the chronically epileptic hippocampus is illustrated in Fig. 6.
Figure 6.
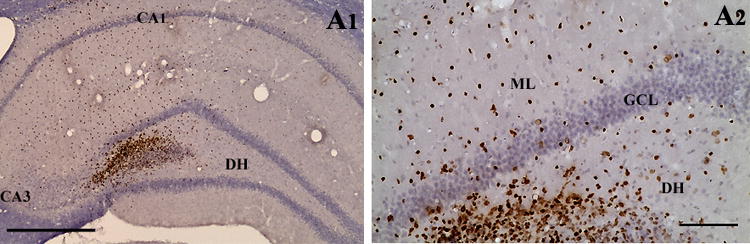
An example showing the distribution of BrdU+ NSC graft-derived cells in the hippocampus of a chronically epileptic rat (A1). A2 is a magnified view of a region from A1 showing the migration of cells into different regions of the host dentate gyrus. Scale bar, A1, 400 μm; A2, 100 μm.
Quantification of the Yield of Graft-Derived Cells (BrdU+ cells) using Stereology
For quantifying the yield of graft-derived cells, we typically perform counting of all BrdU+ cells in every 10th section through the entire anterior-posterior extent of the hippocampus using the optical fractionator counting method in the StereoInvestigator system (Microbrightfield Inc., Williston, VT). The overall yield of graft-derived cells may be expressed as the total number of engrafted cells per hippocampus or as the percentage of the total injected cells per hippocampus (i.e. the total number of BrdU+ cells placed at 4 sites in each hippocampus). Notes: It is important to note that the yield of graft-derived cells does not represent the absolute survival of cells that are originally implanted. However, assessment of the yield does give an idea about the numbers of graft-derived cells that engraft for prolonged periods when a given number of NSCs are implanted. Gauging the yield is important for NSC grafts, as quantification of the absolute graft cell survival is difficult due to the likelihood that both cell proliferation and cell death occur in these grafts. Thus, one can conclude that the overall engraftment is robust if the yield of graft-derived cells is closer to or greater than the number of cells originally implanted. A yield greater than the number of cells initially injected can happen if the NSCs divide a couple of times after the grafting. The major aspects of the optical fractionator cell counting methodology are described below.
The StereoInvestigator system consisted of a color digital video camera (Optronics Inc., Muskogee, OK) interfaced with a Nikon E600 microscope. In each hippocampus, the BrdU+ cells are counted from randomly and systematically selected frames (e.g. each measuring 150 × 150 μm, 0.0225 mm2 area) in every 10th section using the 100× oil immersion objective lens. The numbers and densities of frames are determined by entering the parameter grid size (e.g. 60 × 60 μm) in the optical fractionator component of the StereoInvestigator system (Rao and Shetty, 2004). Although 30 μm thick sections through the hippocampus are cut using a cryostat, the section thickness at the time of counting reduces to 40–50% of the initial thickness because of the shrinkage of the tissue with the BrdU immunostaining. Hence, at the time of data collection, the thickness of sections varies from 12–15 μm. Note: It is very important to check the thickness of each section used for counting before entering the section thickness, as shrinkage can vary between sections.
For commencing cell counting in the each chosen section, open the serial section manager dialog box and enter different parameters such as the number of sections, section evaluation interval, section cut thickness, mounted section thickness, and the starting Z-level.
Mark the contour of the transplant area (i.e. the area of the hippocampus containing the graft-derived [i.e. BrdU+] cells) using the 4× or 10× objective lens and the tracing function of the StereoInvestigator.
Activate the optical fractionator component of the program and select the numbers and locations of the counting frames (using a systematic random sampling scheme) and the counting depth by entering parameters such as the grid size (e.g. 60 × 60 μm), the thickness of top guard zone (e.g. 4 μm) and the optical dissector height (e.g. 6 μm). Note: The number of frames per section counted varies as the overall area comprising the graft-derived cells varies from section to section. This sampling scheme is consistent with the principle of the optical fractionator counting method that the sample concentration must remain constant for each section, implying that the number of unit volumes to be counted per volume of the structure on a given section needs to be a constant ratio from section to section. Thus, counting of cells from randomly and systematically chosen frames in every 10th section through the hippocampus guarantees that effectively every BrdU+ cell within the transplant area has equal odds of being counted. This is imperative because the scattering of BrdU cells within the graft area is visibly heterogeneous.
A computer driven motorized stage then allows the section to be analyzed at each of the counting frame locations. In every counting frame location, identify the top of the section, move the plane of the focus to a depth of 4 μm (guard zone) to get rid of the problem of uneven section surface. This plane serves as the first point of the counting process. Count all BrdU+ cells that come into focus in the next 6-μm section thickness if they are entirely within the counting frame or touching the upper or right side of the counting frame.
Repeat the above protocol for every chosen section in each hippocampus. Following collection of data from all sections, select “display probe run list” to obtain the total number of graft-derived (BrdU+) cells. The extent of migration of graft-derived cells can be assessed indirectly by obtaining the total volume of the hippocampal tissue comprising the graft-derived cells from “display probe run list”. Based on the different parameters that are entered at the beginning of the counting and the number of cells sample at different counting locations in all sections, the StereoInvestigator program calculates the total number of BrdU+ cells per hippocampus by utilizing the optical fractionator formula, N = 1/ssf.1/asf.1/hsf.EQ− (Dorph-Petersen et al., 2001). The abbreviation ssf stands for the section sampling fraction, which is 10 in this example; asf symbolizes the area sampling fraction, which is calculated by dividing the total areas sampled with the total area of the transplant (i.e. the sum of transplant areas in every 10th section); hsf stands for the height sampling fraction, which is calculated by dividing the height sampled (which is 6 μm in this example) with the section thickness at the time of analysis (i.e. 12–15 μm); EQ− denotes the total count of particles (i.e. BrdU+ cells) sampled for the entire hippocampus. Figure 7 illustrates the distribution of cells derived from NSC grafts along the septo-temporal axis of the hippocampus of a chronically epileptic rat (Waldau et al., 2010).
Following collection of data (such as the total number of BrdU+ cells per hippocampus), check the Gundersen coefficient of error (CE) for each sample. The CE values that are <0.05 suggest that the counts obtained are valid. Notes: If larger CE values are seen for some samples, repeat the counting process by modifying the counting parameters, which may include increasing the number of sections (e.g. every 5th section instead of the every 10th section), altering the grid size to increase the number of sites per section, and changing the counting frame dimensions to increase the probability of counting more cells at each counting location.
Figure 7.
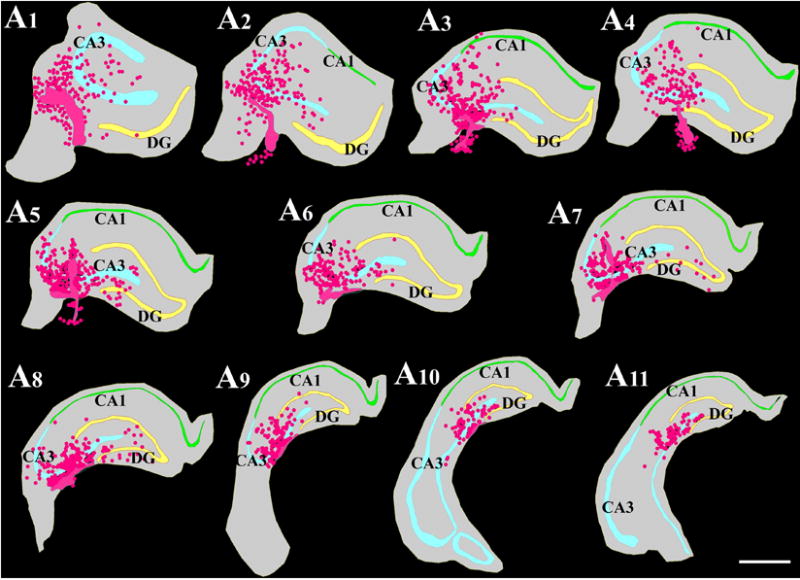
Location of NSC grafts and NSC graft-derived cells (shown in pink color based on Chlorodeoxyuridine+ [CldU+]) graft-derived cells) with respect to hippocampal cell layers and subfields in a chronically epileptic rat. These tracings, performed using the Neurolucida software (Microbrightfield Inc), represent every tenth 30-μm thick section through a chronically epileptic hippocampus that received four MGE-NSC grafts. Scale bar, 1000 μm. [Reproduced from: Waldau et al., Stem Cells, 28:1153–1164].
Phenotypic Analyses of Graft-Derived Cells in the Host Brain
Cells derived from the NSCs are heterogeneous and each type of cell derived from NSCs has unique function. Therefore, it is vital to quantify the fraction of graft-derived cells that become mature neurons (e.g. NeuN+ cells), specialized neurons (e.g. GABA-ergic neurons), mature astrocytes (e.g. S-100β+ cells), oligodendrocyte progenitors (e.g. NG2+ cells), and mature oligodendrocytes (e.g. O1+ and Rip+ cells). In addition, as NSC differentiation might depend on the factors in the host tissue or the location of engraftment, it is also important to assess the fraction of graft-derived cells that persist as NSCs (e.g. Sox-2+ or nestin+ cells) in the host brain. This will provide clues as to whether cells derived from NSC grafts facilitate repair and regeneration of the host brain area via replacement of lost cells or through mechanisms such as the release of neurotrophic or other beneficial factors. The differentiation of graft-derived cells is typically assessed through a standard dual immunofluorescence method and Z-section analyses in a confocal microscope, by identifying both the graft cell marker and the chosen neural cell marker (such as NeuN, GABA, S-100β+, NG2, O1, Rip, Sox-2, nestin etc.). Notes: As dual immunofluorescence approach can be done using different protocols, we restrict our description mainly to the listing of primary and secondary antibody combinations that we typically use in our studies (see Table 1 for details). We employ free-floating sections in 24 well tissue culture plates and sequentially visualize the two antigens. The first antigen is typically the graft cell marker (i.e. BrdU) where as the second antigen is the chosen neural cell/NSC marker. The overall procedure takes ~3 days to complete for each combination. Following dual immunofluorescence staining, we perform Z-section analyses in a confocal microscope within a couple of weeks to prevent the fading of fluorescence over time. We obtain Z-sections at 1 μm intervals and perform orthogonal analysis to confirm the presence of dual antigens in all graft-derived cells that are selected for quantification. This will facilitate the assessment of the percentages of graft-derived cells that express different neural antigens. Examples of the differentiation of cells derived from MGE-NSC grafts into different neural phenotypes in the chronically epileptic hippocampus are illustrated in Fig. 8.
Figure 8.
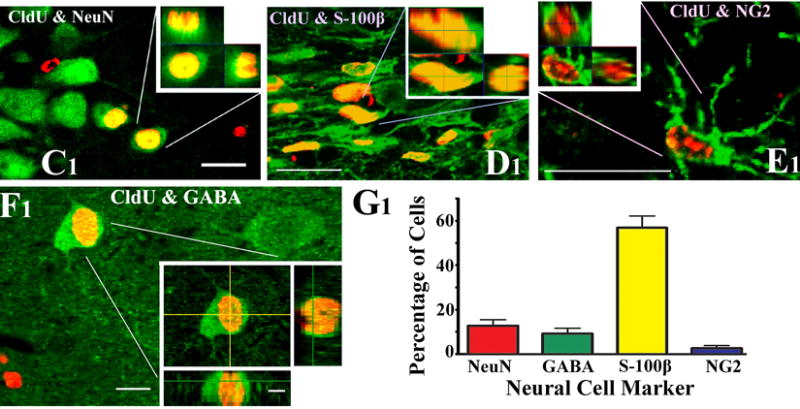
Examples of the differentiation of MGE-NSC graft-derived CldU+ cells into NeuN+ mature neurons (C1), S-100β+ mature astrocytes (D1), NG2+ oligodendrocyte precursors (E1) and GABA+ neurons (F1), visualized through dual immunolabeling for CldU (red) and markers of neurons/glia (green) and Z-section analyses in a confocal microscope. Scale bar, C1-E1, 20 μm; F1, 10 μm; orthogonal inset of F1, 5 μm. The bar chart in G1 depicts the percentages of graft-derived cells that differentiate into NeuN+ neurons, GABA+ neurons, S-100β+ astrocytes and NG2+ oligodendrocyte precursors. [Reproduced from: Waldau et al., Stem Cells, 28:1153–1164].
ANALYSES OF EFFECT OF NSC GRAFTING ON THE HOST HIPPOCAMPUS
In addition to contributing new neurons synthesizing the inhibitory neurotransmitter GABA and new astrocytes synthesizing the anticonvulsant protein GDNF (Waldau et al., 2010), NSC grafting into the epileptic hippocampus might also influence the host cells and neurons, as observed in other disease models. For example, NSCs placed into the substantia nigra and caudate nuclei of monkeys exhibiting Parkinson’s disease can induce behavioral recovery with differentiation of only a small fraction of grafted NSCs into dopaminergic neurons (Redmond et al., 2007). As large numbers of grafted NSCs differentiated into astrocytes and expressed multiple neurotrophic factors including GDNF, it appeared that NSC grafting mediated changes to the host milieu enhanced the function of the existing dopaminergic neurons (Redmond et al., 2007; Sanberg, 2007). Considering this, it is important to examine the effects of NSC grafts on the host microenvironment and host cells, in addition to examining the differentiation of graft-derived cells into desired neuronal or glial phenotypes. In our recent study, we utilized a standard dual immunofluorescence approach to assess the changes in the function of S-100β+ host astrocytes with NSC grafting into the hippocampus of CERs (Waldau et al., 2010). The results showed that NSC grafting into the hippocampus of CERs considerably restores the GDNF expression in the S-100β+ host hippocampal astrocytes (Waldau et al., 2010). However, it is likely helpful to use a high-throughput genomic (such as microarray and qRT-PCR arrays) and proteomic approaches in the future to understand the effects of NSC grafting on multiple pathways in the host hippocampus of CERs. The methods for these approaches are not described here as they are beyond the scope of this article.
COMMENTARY
Background Information for TLE
Epilepsy, typified by spontaneous recurrent seizures (SRS) due to hyperexcitability and synchronization of activity within populations of neurons, affects >50 million people (Strine et al., 2005). The TLE, characterized by progressive development of complex partial seizures and hippocampal neurodegeneration, is seen in ~30% of epileptic patients (Manford et al., 1992). While the etiology of TLE is unknown in most cases (McNamara, 1999), it is typically seen after an initial precipitating injury such as SE, brain injury, tumors, meningitis, encephalitis and febrile seizures in other cases (French et al., 1993; Mathern et al., 1995; Lewis, 2005). Seizures in TLE originate from the temporal lobe foci, which are associated with learning and memory impairments, reduced dentate neurogenesis and depression (Devinsky, 2004; Hattiangady et al., 2004; Detour et al., 2005; Pirttila et al., 2005; Hattiangady and Shetty, 2010). Nearly 35% of patients with TLE exhibit seizures that cannot be controlled by antiepileptic drugs (Litt et al., 2001) and memory difficulties are a frequent cognitive complaint in patients with chronic epilepsy (Vannest et al., 2008). Although surgical resection of the epileptic hippocampus gives better seizure control, this option is often associated with significant cognitive impairments (Helmstaedter et al., 2008). Hence, there is a pressing need to develop alternative therapeutic approaches that greatly diminish both frequency & intensity of SRS in patients with chronic TLE on a long-term basis.
Features of KA Model of TLE
Pharmacological SE animal models using KA or pilocarpine are popular for studying TLE, as they replicate several features of TLE. Kainic acid is a widely used excitotoxin in studies of the hippocampus (Nadler et al., 1978; Sperk, 1994; Shetty and Turner, 1996). Intraperitoneal (IP) administration of KA in rat induces degeneration of dentate hilar neurons and fractions of CA1 and CA3 pyramidal neurons, deafferentation of dentate granule cells due to loss of hilar mossy cells, and reductions in numbers of GABA-ergic interneurons in the hippocampus. These changes are followed by aberrant sprouting of dentate mossy fibers into the dentate supragranular layer (DSGL), hyperexcitability in the hippocampus, and a chronic epileptic condition, typified by SRMS, learning and memory deficits and depression (Ben-Ari, 1985; Letty et al., 1995; Hellier et al., 1998, 1999; Buckmaster and Dudek, 1999; Wuarin and Dudek, 2001; Rao et al., 2006). Since the above changes in the adult hippocampus closely resemble human mesial TLE, IP KA model has been widely used for studying TLE. This model has also been found to be useful for studying the effects of cell grafts on chronic seizures (Rao et al., 2007; Waldau et al., 2010). Thus, the IP KA model is suitable for testing the efficacy of NSC grafts for alleviating chronic epilepsy characterized by spontaneous seizures and learning & memory impairments.
GABA-ergic Cell Therapy for TLE
It is believed that an increased excitatory neurotransmission found in the epileptic hippocampus is partly due to a reduced number of GABA-ergic interneurons, loss of functional inhibition and diminished numbers of GABA-ergic terminals (Ribak et al., 1986; Cornish and Wheal, 1989; During et al., 1995; Shetty and Turner, 2000, 2001; Shetty et al., 2009). From this perspective, the idea of restraining SRS in the epileptic hippocampus via grafting of cells that just release the inhibitory neurotransmitter GABA at the seizure focus has received considerable attention (Löscher et al., 2008). For instance, grafting of GABA-soaked beads, immortalized GABA-ergic cells, cells that are engineered to produce GABA, and fetal GABA-ergic cells into the epileptic foci have been shown to transiently reduce seizures in a variety of animal models (Löscher et al., 1998; Gernert et al., 2002; Thompson, 2005; Castillo et al., 2006; Hattiangady et al., 2008). Thus, grafting of GABA-producing cells into the epileptic brain has considerable promise for restraining seizures. However, a routine clinical application of human fetal cells for TLE may not be feasible because of the difficulty in obtaining the required quantity of human fetal cells and ethical issues (Turner and Shetty, 2003). With reference to the other cell types, it is unknown whether immortalized GABA-producing cells have the ability for long-term survival in the chronically epileptic brain, as they are expected to perform as GABA pumps rather than undergoing synaptic integration with the host neurons (Loscher et al., 2008). Therefore, there is a need to find types of cells that are capable of providing an unlimited source of donor cells for grafting and have the ability to give rise to large numbers of GABA-ergic interneurons that survive for prolonged periods after grafting into the epileptic brain.
From the above perspectives, it appears that NSCs expanded from the embryonic MGE (Waldau et al., 2010) and the postnatal & adult subventricular zone (SVZ) are attractive candidates as donor cells for grafting therapy in TLE. This is because, these cells can: (i) be expanded for prolonged periods in culture without losing their multipotential property; (ii) give rise substantial numbers of GABA-ergic interneurons; and (iii) release a multitude of neurotrophic factors (Shetty and Hattiangady, 2007; Waldau et al., 2010). Additionally, NSCs obtained from sources such as hESCs and iPSCs may also be used once their potential to give rise to full-fledged GABA-ergic neurons and to release a multiple neurotrophic factors is validated. However, prior to the clinical application of NSCs obtained from any source for TLE, it will be necessary to rigorously analyze the ability of the chosen NSCs to restrain SRS on a long-term basis using both behavioral and EEG analyses in animal models of TLE. In addition, the efficacy of different NSC grafts for improving cognitive and mood function in TLE need to be examined. Particularly, detailed anatomical, electrophysiological and molecular biological analyses of the efficacy of grafting of NSCs for easing SRS and cognitive and mood dysfunction in chronic TLE on a long-term basis are needed before considering the clinical application of NSC therapy for TLE.
CRITICAL PARAMETERS & TROUBLESHOOTING
Observation and Care of Animals During and Shortly After SE
Generating CERs via SE induction is a laborious process that needs patience and close attention to every step. Some of the issues and precautions to be taken are described here. (1) The sensitivity to KA may vary among F344 rats purchased from different sources. Hence, it is ideal to use rats obtained from the same source for all experiments in a particular study. (2) It is important to use KA from a single source for the entire study, as KA from different vendors seems to vary in terms of their potency to induce acute seizures or SE. It is recommended that the potency of KA from a particular source be checked in a pilot experiment and determine the appropriate dose for inducing SE prior to initiating SE studies in a large number of animals. (3) The use of cages having flat tops fitted with foam lining on the inner side is useful for preventing injury to rats in the event of bouncing seizures after the onset of SE. (4) If a rat is having very intense seizures such as bouncing seizures after the onset of SE (which can happen suddenly), carefully handling the rat and inducing hypothermia (e.g. by placing the rat in an empty cage that was kept on a container filled with ice) for a few minutes stops such intense seizures and prevents mortality. (5) Injecting diazepam (5 mg/kg b.w) after 3 hrs of seizure activity considerably reduces the mortality of rats and does not interfere with the occurrences of SRMS in the chronic phase after SE. Without diazepam administration, the overnight mortality after SE will be significantly higher though the rats that survive might exhibit robust chronic epilepsy (characterized by greater frequency and intensity of SRMS). On the other hand, if diazepam is administered very early after the onset of SE (e.g. immediately after the first stage V seizure or before the occurrence of stage V seizure), only a small fraction of rats would develop chronic epilepsy at 4–6 months post-SE. months.
Appropriate animal care after SE is another requirement for reducing SE related mortality. After SE animals appear weak, lethargic and anorexic and do not seem to eat hard pellets or drink from the water bottle. Therefore, preventing dehydration via subcutaneous administration of fluids (such as saline or Ringer’s lactate solution) and providing soft rat chow or transgel within their cage in a dish clearly helps in minimizing mortality in the recovery phase after SE.
Preparation and Injection of NSC Suspension
The methods such as dissection of MGE tissues from the embryonic brain, expansion of NSCs in culture and preparation of NSC suspension for grafting can be performed only by personnel with specific expertise in these aspects. Trituration of neurospheres is a highly skilled technique, as too gentle trituration does not dissociate the neurospheres well and too harsh trituration would kill a large number of cells. Triturating neurospheres in a proliferation medium minimizes cell death, in comparison to triturating in any other medium. It is important to select the NSC suspension with a good viability index (e.g. >80%) for grafting studies because an excellent viability of donor cells in the suspension at the time of grafting appears to be an important pre-requisite for successful engraftment into the host brain and for obtaining higher yields of graft-derived cells at extended time-points after grafting. The cell suspension with a poor overall viability (e.g. <70%) should not be used for grafting, as greater numbers of dead cells in the graft might initiate an inflammatory reaction in the host brain. Higher volume injections in a single site should be avoided into the hippocampus, as it typically causes tissue damage, opening of the tissue cleavage planes (e.g. the hippocampal fissure) and the flow of the injected fluid mostly into the lateral ventricle. Based on our experience, injection of 1μl volume comprising 80,000 to 100,000 live cells (per site) into the hippocampus results in good engraftment of graft-derived cells and does not seem to cause tissue damage at the grafted site. Therefore, preparation of NSC suspension with a high density of cells is required, which clearly requires expansion of NSCs in a large number of flasks.
Water Maze Studies in CERs
Typically, CERs do not exhibit SRS while handling. However, occasionally, it is possible that a CER can have a seizure in the middle of a learning trial. In such cases, it is necessary to remove the rat from the water quickly to prevent its drowning. Such rats should be observed closely for a while and new trials should not be given in the immediate post-SRS confusion period. However, new set of trials can be given to such rats after a delay of 20–30 minutes depending upon their overall behavior. Furthermore, some CERs might show a tendency to drown in the pre-trials. Such rats should be excluded from the WMT. Another potential problem is that some of the CERs may develop eyesight/motor impairments, which could confound results. To avoid this, test the visual acuity & motor functions of all rats after each WMT. This may consist of 4 swim trials with a randomly positioned visible platform and no extra-maze cues.
ANTICIPATED RESULTS
When all guidelines and protocols described in this article are faithfully followed, success of the experiment (i.e. testing the efficacy of NSC grafting for restraining SRS and improving cognitive function in chronic TLE) in terms of drawing reliable conclusions is ensured. Additionally, the experimental results can be reproduced with such a stringent adherence to the guidelines. However, errors such as inclusion of rats from different sources, the use of KA from different sources, an inappropriate selection of CERs (e.g. a group of rats with widely differing frequency and intensity of SRMS) for grafting or other groups, grafting the donor NSC suspension with a poor viability of cells, larger volume cell suspension injections into the hippocampus, and withdrawing needle immediately after the injection of the cell suspension would introduce significant confounds and the results obtained from such experiments will be unreliable for meaningful conclusions and non-reproducible.
TIME CONSIDERATIONS
The experiment described here for testing the efficacy of NSC grafting for restraining SRS and improving cognitive function using an animal model of chronic TLE is clearly a long-term experiment. From our experience, animal aspects of the work such as induction of SE, characterization of SRMS and cognitive dysfunction in CERs, Expansion and grafting of NSCs, post-grafting analyses of SRMS (via direct observations/video recordings), SRS (through video-EEG recordings), cognitive function (via a WMT) take approximately an year to complete. Following this, processing of tissues via perfusion, serial section cutting of brain tissues, histology, immunostaining, stereological counting of graft-derived cells, dual immunofluorescence & confocal microscopic analyses of the phenotype of graft derived cells, and analyzing the effects of grafts on host cells and microenvironment would need significant additional amounts of time. The overall time required for histological processing, immunostaining and quantification depends on the number of animals in different groups, numbers of experimental groups, and the type of analysis undertaken. Stereotaxic surgery can be laborious, as injections are made into multiple sites in the hippocampus. However, with practice, transplantation can be performed to six animals in one surgery session if two stereotactic devices are available. Measuring SRMS either via direct observation or from the recorded video requires strong observation skills and an ability to distinguish SRMS from the normal grooming behavior of rats throughout the observation period. Furthermore, enthusiastic research personnel/scientists who are committed to this type of long-running and labor-intensive (but very interesting and clinically relevant) experiment are needed for successfully completing these studies.
Acknowledgments
Authors greatly acknowledge the funding from the National Institute of Neurological Disorders and Stroke (R01 NS054780, R01 NS043507 & R01 NS36742 to A.K.S.) and the Department of Veterans Affairs (Merit Review Awards to A.K.S.) for their studies on animal models of temporal lobe epilepsy over the past 13 years. Authors wish to thank Vandana Zaman, Muddanna Rao, Ben Waldau, Ramkumar Kuruba, and Bing Shuai for their outstanding contributions to original articles on neural cell grafting in animal models of temporal lobe epilepsy published from SHETTY LAB at the Duke University Medical Center and Durham VA Medical Center.
Literature Cited
- Astur RS, Taylor LB, Mamelak AN, Philpott L, Sutherland RJ. Humans with hippocampus damage display severe spatial memory impairments in a virtual Morris water task Behav. Brain Res. 1995;132:77–84. doi: 10.1016/s0166-4328(01)00399-0. [DOI] [PubMed] [Google Scholar]
- Ben-Ari Y. Limbic seizures and brain damage produced by kainic acid: Mechanisms and relevance to human temporal lobe epilepsy. Neurosci. 1985;14:375–403. doi: 10.1016/0306-4522(85)90299-4. [DOI] [PubMed] [Google Scholar]
- Buckmaster PS, Dudek FE. In vivo intracellular analysis of granule cell axon reorganization in epileptic rats. J Neurophysiol. 1999;81:712–721. doi: 10.1152/jn.1999.81.2.712. [DOI] [PubMed] [Google Scholar]
- Castillo CG, Mendoza S, Freed WJ, Giordano M. Intranigral transplants of immortalized GABAergic cells decrease the expression of kainic acid-induced seizures in the rat. Behav Brain Res. 2006;17:109–115. doi: 10.1016/j.bbr.2006.03.025. [DOI] [PubMed] [Google Scholar]
- Coras R, Siebzehnrubl FA, Pauli E, Huttner HB, Njunting M, Kobow K, Villmann C, Hahnen E, Neuhuber W, Weigel D, Buchfelder M, Stefan H, Beck H, Steindler DA, Blümcke I. Low proliferation and differentiation capacities of adult hippocampal stem cells correlate with memory dysfunction in humans. Brain. 2010 Aug 18; doi: 10.1093/brain/awq215. [Epub ahead of print] [DOI] [PubMed] [Google Scholar]
- Cornish SM, Wheal HV. Long-term loss of paired pulse inhibition in the kainic acid-lesioned hippocampus of the rat. Neurosci. 1989;28:563–571. doi: 10.1016/0306-4522(89)90005-5. [DOI] [PubMed] [Google Scholar]
- Detour J, Schroeder H, Desor D, Nehlig A. A 5-month period of epilepsy impairs spatial memory, decreases anxiety, but spares object recognition in the lithium-pilocarpine model in adult rats. Epilepsia. 2005;46:499–508. doi: 10.1111/j.0013-9580.2005.38704.x. [DOI] [PubMed] [Google Scholar]
- Devinsky O. Therapy for neurobehavioral disorders in epilepsy. Epilepsia. 2004;45(Suppl 2):34–40. doi: 10.1111/j.0013-9580.2004.452003.x. [DOI] [PubMed] [Google Scholar]
- Dorph-Petersen KA, Nyengaard JR, Gundersen HJ. Tissue shrinkage and unbiased stereological estimation of particle number and size. J Microsc. 2001;204:232–246. doi: 10.1046/j.1365-2818.2001.00958.x. [DOI] [PubMed] [Google Scholar]
- During MJ, Ryder KM, Spencer DD. Hippocampal GABA transporter function in temporal-lobe epilepsy. Nature. 1995;376:174–177. doi: 10.1038/376174a0. [DOI] [PubMed] [Google Scholar]
- Fisher PD, Sperber EF, Moshe SL. Hippocampal sclerosis revisited. Brain Dev. 1998;20:563–573. doi: 10.1016/s0387-7604(98)00069-2. [DOI] [PubMed] [Google Scholar]
- French JA, Williamson PD, Thadani VM, Darcey TM, Mattson RH, Spencer SS, Spencer DD. Characteristics of medial temporal lobe epilepsy: I. Results of history and physical examination. Ann Neurol. 1993;34:774–780. doi: 10.1002/ana.410340604. [DOI] [PubMed] [Google Scholar]
- Gernert M, Thompson KW, Löscher W, Tobin AJ. Genetically engineered GABA-producing cells demonstrate anticonvulsant effects and long-term transgene expression when transplanted into the central piriform cortex of rats. Exp Neurol. 2002;176:183–192. doi: 10.1006/exnr.2002.7914. [DOI] [PubMed] [Google Scholar]
- Hattiangady B, Rao MS, Shetty AK. Chronic temporal lobe epilepsy is associated with severely diminished dentate neurogenesis in the adult hippocampus. Neurobiol Dis. 2004;17:473–490. doi: 10.1016/j.nbd.2004.08.008. [DOI] [PubMed] [Google Scholar]
- Hattiangady B, Rao MS, Shetty AK. Grafting of striatal precursor cells into hippocampus shortly after status epilepticus restrains temporal lobe epilepsy. Exp Neurol. 2008;212:468–481. doi: 10.1016/j.expneurol.2008.04.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hattiangady B, Shetty AK. Decreased neuronal differentiation of newly generated cells underlies diminished neurogenesis in chronic temporal lobe epilepsy. Hippocampus. 2010;20:97–112. doi: 10.1002/hipo.20594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hellier JL, Patrylo PR, Buckmaster PS, Dudek FE. Recurrent spontaneous motor seizures after repeated low-dose systemic treatment with kainate: assessment of a rat model of temporal lobe epilepsy. Epilepsy Res. 1998;31:73–84. doi: 10.1016/s0920-1211(98)00017-5. [DOI] [PubMed] [Google Scholar]
- Hellier JL, Patrylo PR, Dou P, Nett M, Rose GM, Dudek FE. Assessment of inhibition and epileptiform activity in the septal dentate gyrus of freely behaving rats during the first week after kainate treatment. J Neurosci. 1999;19:10053–10064. doi: 10.1523/JNEUROSCI.19-22-10053.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helmstaedter C, Loer B, Wohlfahrt R, Hammen A, Saar J, Steinhoff BJ, Quiske A, Schulze-Bonhage A. The effects of cognitive rehabilitation on memory outcome after temporal lobe epilepsy surgery. Epilepsy Behav. 2008;12:402–409. doi: 10.1016/j.yebeh.2007.11.010. [DOI] [PubMed] [Google Scholar]
- Letty S, Lerner-Natoli M, Rondouin G. Differential impairments of spatial memory and social behavior in two models of limbic epilepsy. Epilepsia. 1995;36:973–982. doi: 10.1111/j.1528-1157.1995.tb00955.x. [DOI] [PubMed] [Google Scholar]
- Lewis DV. Losing neurons: selective vulnerability and mesial temporal sclerosis. Epilepsia. 2005;46(Suppl 7):39–44. doi: 10.1111/j.1528-1167.2005.00306.x. [DOI] [PubMed] [Google Scholar]
- Litt B, Esteller R, Echauz J, D’Alessandro M, Shor R, Henry T, Pennell P, Epstein C, Bakay R, Dichter M, Vachtsevanos G. Epileptic seizures may begin hours in advance of clinical onset: A report of five patients. Neuron. 2001;30:51–64. doi: 10.1016/s0896-6273(01)00262-8. [DOI] [PubMed] [Google Scholar]
- Löscher W, Ebert U, Lehmann H, Rosenthal C, Nikkhah G. Seizure suppression in kindling epilepsy by grafts of fetal GABAergic neurons in rat substantia nigra. J Neurosci Res. 1998;51:196–209. doi: 10.1002/(SICI)1097-4547(19980115)51:2<196::AID-JNR8>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- Löscher W, Gernert M, Heinemann U. Cell and gene therapies in epilepsy – promising avenues or blind alleys? Trends Neurosci. 2008;31:62–73. doi: 10.1016/j.tins.2007.11.012. [DOI] [PubMed] [Google Scholar]
- Manford M, Hart YM, Sander JW, Shorvon SD. National General Practice Study of Epilepsy (NGPSE): partial seizure patterns in a general population. Neurology. 1992;42:1911–1917. doi: 10.1212/wnl.42.10.1911. [DOI] [PubMed] [Google Scholar]
- Mathern GW, Pretorius JK, Babb TL. Influence of the type of initial precipitating injury and at what age it occurs on course and outcome in patients with temporal lobe seizures. J Neurosurg. 1995;82:220–227. doi: 10.3171/jns.1995.82.2.0220. [DOI] [PubMed] [Google Scholar]
- McNamara JO. Emerging insights into the genesis of epilepsy. Nature. 1999;399(6738 Suppl):A15–22. doi: 10.1038/399a015. 4. [DOI] [PubMed] [Google Scholar]
- Moser MB, Moser EI. Distributed encoding and retrieval of spatial memory in the hippocampus. J Neurosci. 1998;18:7535–7542. doi: 10.1523/JNEUROSCI.18-18-07535.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nadler JV, Perry BW, Cotman CW. Intraventricular kainic acid preferentially destroys hippocampal pyramidal cells. Nature. 1978;271:676–677. doi: 10.1038/271676a0. [DOI] [PubMed] [Google Scholar]
- Pirttilä TJ, Manninen A, Jutila L, Nissinen J, Kälviäinen R, Vapalahti M, Immonen A, Paljärvi L, Karkola K, Alafuzoff I, Mervaala E, Pitkänen A. Cystatin C expression is associated with granule cell dispersion in epilepsy. Ann Neurol. 2005;58:211–223. doi: 10.1002/ana.20545. [DOI] [PubMed] [Google Scholar]
- Racine RJ. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr Clin Neurophysiol. 1972;32:281–294. doi: 10.1016/0013-4694(72)90177-0. [DOI] [PubMed] [Google Scholar]
- Rao MS, Hattiangady B, Reddy DS, Shetty AK. Hippocampal neurodegeneration, spontaneous seizures, and mossy fiber sprouting in the F344 rat model of temporal lobe epilepsy. J Neurosci Res. 2006;83:1088–1105. doi: 10.1002/jnr.20802. [DOI] [PubMed] [Google Scholar]
- Rao MS, Hattiangady B, Rai KS, Shetty AK. Strategies for promoting anti-seizure effects of hippocampal fetal cells grafted into the hippocampus of rats exhibiting chronic temporal lobe epilepsy. Neurobiol Dis. 2007;27:117–132. doi: 10.1016/j.nbd.2007.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rao MS, Shetty AK. Efficacy of doublecortin as a marker to analyze the absolute number and dendritic growth of newly generated neurons in the adult dentate gyrus. Eur J Neurosci. 2004;19:234–246. doi: 10.1111/j.0953-816x.2003.03123.x. [DOI] [PubMed] [Google Scholar]
- Redmond DE, Jr, Bjugstad KB, Teng YD, Ourednik V, Ourednik J, Wakeman DR, Parsons XH, Gonzalez R, Blanchard BC, Kim SU, Gu Z, Lipton SA, Markakis EA, Roth RH, Elsworth JD, Sladek JR, Jr, Sidman RL, Snyder EY. From the Cover: Behavioral improvement in a primate Parkinson’s model is associated with multiple homeostatic effects of human neural stem cells. Proc Natl Acad Sci USA. 2007;104:12175–12180. doi: 10.1073/pnas.0704091104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ribak CE, Hunt CA, Bakay RA, Oertel WH. A decrease in the number of GABAergic somata is associated with the preferential loss of GABAergic terminals at epileptic foci. Brain Res. 1986;363:78–90. doi: 10.1016/0006-8993(86)90660-8. [DOI] [PubMed] [Google Scholar]
- Sanberg PR. Neural stem cells for Parkinson’s disease: To protect and repair. Proc Natl Acad Sci USA. 2007;104:11869–11870. doi: 10.1073/pnas.0704704104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shetty AK, Hattiangady B. Prospects of stem cell therapy for temporal lobe epilepsy. Stem Cells. 2007;25:2396–407. doi: 10.1634/stemcells.2007-0313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shetty AK, Turner DA. Development of fetal hippocampal grafts in intact and lesioned hippocampus. Prog Neurobiol. 1996;50:597–653. doi: 10.1016/s0301-0082(96)00048-2. [DOI] [PubMed] [Google Scholar]
- Shetty AK, Turner DA. Fetal hippocampal CA3 cell transplants restore host hippocampal GAD-positive interneuron numbers in a rat model of TLE. J Neurosci. 2000;20:8788–8801. doi: 10.1523/JNEUROSCI.20-23-08788.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shetty AK, Turner DA. Glutamic acid decarboxylase positive hippocampal interneurons undergo a permanent reduction in number following kainic acid induced destruction of CA3 pyramidal neurons. Exp Neurol. 2001;176:276–297. doi: 10.1006/exnr.2001.7668. [DOI] [PubMed] [Google Scholar]
- Shetty AK, Hattiangady B, Rao MS. Vulnerability of hippocampal GABA-ergic interneurons to kainate induced excitotoxic injury during old age. J Cell Mol Med. 2009;13(8B):2408–2423. doi: 10.1111/j.1582-4934.2008.00675.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sperk G, Wieser R, Widmann R, Singer EA. Kainic acid induced seizures: changes in somatostatin, substance P and neurotensin. Neurosci. 1994;17:1117–1126. doi: 10.1016/0306-4522(86)90081-3. [DOI] [PubMed] [Google Scholar]
- Strine TW, Kobau R, Chapman DP, Thurman DJ, Price P, Balluz LS. Psychological distress, comorbidities, and health behaviors among U.S. adults with seizures: results from the 2002 National Health Interview Survey. Epilepsia. 2005;46:1133–1139. doi: 10.1111/j.1528-1167.2005.01605.x. [DOI] [PubMed] [Google Scholar]
- Thompson KW. Genetically engineered cells with regulatable GABA production can affect afterdischarges and behavioral seizures after transplantation into the dentate gyrus. Neurosci. 2005;133:1029–1037. doi: 10.1016/j.neuroscience.2005.03.003. [DOI] [PubMed] [Google Scholar]
- Turner DA, Shetty AK. Clinical prospects for neural grafting therapy for hippocampal lesions and epilepsy. Neurosurgery. 2003;52:632–644. doi: 10.1227/01.neu.0000047825.91205.e6. [DOI] [PubMed] [Google Scholar]
- Vannest J, Szaflarski JP, Privitera MD, Schefft BK, Holland SK. Medial temporal fMRI activation reflects memory lateralization and memory performance in patients with epilepsy. Epilepsy Behav. 2008;12:410–418. doi: 10.1016/j.yebeh.2007.11.012. [DOI] [PubMed] [Google Scholar]
- Waldau B, Hattiangady B, Kuruba R, Shetty AK. Medial Ganglionic Eminence-Derived Neural Stem Cell Grafts Ease Spontaneous Seizures and Restore GDNF Expression in a Rat Model of Chronic Temporal Lobe Epilepsy. Stem Cells. 2010;28:1153–64. doi: 10.1002/stem.446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wuarin J-P, Dudek FE. Excitatory synaptic input to granule cells increases with time after kainate treatment. J Neurophysiol. 2001:1067–1077. doi: 10.1152/jn.2001.85.3.1067. [DOI] [PubMed] [Google Scholar]