

Abstract
The microbiome of plants is diverse, and like that of animals, is important for overall health and nutrient acquisition. In legumes and actinorhizal plants, a portion of essential nitrogen (N) is obtained through symbiosis with nodule-inhabiting, N2-fixing microorganisms. However, a variety of non-nodulating plant species can also thrive in natural, low-N settings. Some of these species may rely on endophytes, microorganisms that live within plants, to fix N2 gas into usable forms. Here we report the first direct evidence of N2 fixation in the early successional wild tree, Populus trichocarpa, a non-leguminous tree, from its native riparian habitat. In order to measure N2 fixation, surface-sterilized cuttings of wild poplar were assayed using both 15N2 incorporation and the commonly used acetylene reduction assay. The 15N label was incorporated at high levels in a subset of cuttings, suggesting a high level of N-fixation. Similarly, acetylene was reduced to ethylene in some samples. The microbiota of the cuttings was highly variable, both in numbers of cultured bacteria and in genetic diversity. Our results indicated that associative N2-fixation occurred within wild poplar and that a non-uniformity in the distribution of endophytic bacteria may explain the variability in N-fixation activity. These results point to the need for molecular studies to decipher the required microbial consortia and conditions for effective endophytic N2-fixation in trees.
Introduction
The microbiota of plants can provide a wide range of benefits to the host including increased tolerance to drought, salt, or temperature extremes, the production of phytohormones, resistance to microbial pathogens, detoxification of pollutants, and increased nutrient acquisition [1–4]. In nutrient-limiting environments, plants are most likely to form associations with microorganisms capable of fixing atmospheric dinitrogen (N2) gas into usable forms. Bacteria accomplish this reduction through the action of nitrogenase, a complex multi-subunit enzyme that is inhibited by oxygen. In both aerobic and microaerobic environments, however, diazotrophic (N2-fixing) microorganisms can use a variety of strategies to protect the nitrogenase complex [5,6].
In the well-known symbioses of rhizobia with legumes and Frankia with actinorhizal plants, diazotrophic bacteria fix N2 while within specialized root nodule structures where oxygen levels are regulated. For non-nodulating plants, however, recent evidence points to symbiosis with internal microorganisms, termed endophytes, as a mechanism for these plants to obtain their essential nitrogen (N) [7]. Although N2-fixing endophytes are not in an external structure, the colonized plant tissue itself may be microaerobic [8,9]. The expression of the nitrogenase subunit genes is regulated by oxygen [10], and since nifH expression has been demonstrated in several systems while the bacteria are associated with the plant [11–14], the appropriate conditions must therefore have been met.
N2-fixing endophytes have been isolated from such varied species as kallar grass [15], sugarcane [16], wild rice [17,18], maize [19], Sorghum halepense [20], miscanthus [21], elephant grass [22], rock-dwelling cactus [23], sweet potato [24,25], Boreal mosses [26], dune grasses [27], coffee plants [28], and conifers [29,30], N2-fixation was directly quantified in the Graminaceae such as sugarcane, wheat, and rice [13,15,18,31]. Several field studies utilized the 15N natural abundance technique to demonstrate very substantial inputs of N in sugarcane and elephant grass through biological N2-fixation [32–34]. Despite the decades of research on endophytic N2-fixation, the idea that significant symbiotic N2-fixation can occur in plant tissue without root nodules has remained controversial [35]. In a recent seminal paper by Pankievicz, et al., symbiotic N2-fixation by endophytic and associative bacteria of sugarcane was unequivocally demonstrated in a model C4 grass system [36]. Addition of diazotrophic bacteria to Setaria viridis resulted in N transfer as demonstrated by 13N tracer studies, relief of N-stress symptoms, and restoration of the metabolic profile to that of an N-sufficient state. There is a need for more studies utilizing these technically-challenging and expensive yet direct assays of N2-fixation using isotope tracers.
Although there are numerous reports on biological N2-fixation (BNF) by endophytes in the Graminaceae, especially of tropical grasses, few have reported BNF in temperate plants, especially in trees [29,37]. Members of the Salicaceae family, including poplars (Populus sp.) and willows (Salix sp.), are early successional tree species able to colonize nutrient-poor environments, and are increasingly important for bioenergy, wood products, and environmental services [38,39]. Several studies have assessed the microbiome of P. deltoides and hybrid poplar, and its involvement in poplar growth promotion and remediation of pollutants [40–44]. Populus trichocarpa can thrive in riparian zones where regular flooding from high alpine snow melt deposits cobble and sand, creating new substrate for colonization [45]. Due to its rocky composition, however, this new substrate can be nutrient-limited. The ability to colonize such substrate has often been attributed to N use efficiency; however, in these nutrient-limited areas devoid of organic matter, there is little organic N with which to be efficient. We previously reported the presence of endophytic Rhizobium tropici [46] from hybrid poplar grown in greenhouses. A variety of other diazotrophic endophytic species were subsequently isolated from wild poplar growing in a natural riparian area dominated by cobble [47]. These endophytes were shown to be mutualistic symbionts by inoculation into other plant species, including grasses [48], corn [49], rice [50], and a variety of crop plants including tomato, pepper, squash, and turfgrasses [51], all of which showed improved growth and health under nutrient-limited conditions. The endophytes improved fruit yields in two varieties of tomato by approximately 2-fold. Inoculated perennial rye grass had up to a 26% increase in the foliar N content and up to a 6-fold increase in root N content [51]. Addition of the diazotrophic endophytes from wild poplar to hybrid poplars under greenhouse conditions resulted in an increased chlorophyll and total root N content. BNF in the inoculated poplar plants was estimated through 15N dilution to be 65% N derived from air [52]. Although these studies demonstrated the benefits of inoculation with the endophytes, it was not known if N2-fixation occurs in wild poplar with its natural composition and density of microbiota.
The overall objective of this study was to assess N2-fixation in native riparian black cottonwood (Populus trichocarpa) taken from its natural setting. The 15N incorporation assay is the most direct assay for biological N2 fixation since 15N2 gas is chemically inert such that tissues will only be labeled with 15N if the molecule was biologically reduced into usable forms. Isotopic 15N is present in the atmosphere at such low natural abundance compared to 14N (0.364 atom %15N) that even small increases in 15N label above the unexposed controls are indicative of N2 fixation [53]. The acetylene reduction assay (ARA), an indirect assessment of N2 fixation [54] was also used. We report N2-fixation within some wild poplar samples and demonstrate the presence of diazotrophic bacteria in these plants, helping to explain the biology of poplar as a pioneer plant species.
Materials and Methods
Media used
Cuttings were grown in N-free hydroponic medium (NFM) [47] or in Murashige and Skoog agar [55] modified to be N-free (NFMS; Caisson). Microbial media included the rich mannitol glutamate medium, MG/L [56], the N-limited combined carbon medium, NL-CCM, with sucrose, mannitol, and sodium lactate as C sources [57], and the N-free medium, Nfb, with malic acid as the C source [58].
Research site and plant sampling
Plant samples were collected at the Three Forks Natural Area in King County, WA in the riparian zone of the Snoqualmie River (+47° 31' 14.30", -121° 46' 28.32"). No specific permissions were required since only small clippings were made of some of the trees in this city park; however, a permit from King County was obtained. Although some riparian zones are rich in organic N, this is not the case for this area. The substrate is river cobbles, not rich sediment, and the water is clean, originating from high alpine snow melt. The N level of the river from this site was quantified in 2011 (0.15 mg NO3-N and 0.30 mg NH4-N per liter), in 2014 (0.142 mg/L total N), and in 2015 (average of 8 samples on three sampling dates was 0.33 mg/L total N). Several branch cuttings of 7 black cottonwood (Populus trichocarpa) at a height of approximately 2 meters with a sample length of approximately 25 cm were collected on August 15, 2013 for the 15N2 experiments #1 and #2 and on July 17, 2015 for the ARA experiments. Samples were surface-sterilized with 10% bleach (10 min.) and 1% Iodophor (5 min.), and rinsed three times in sterile DI water. The samples collected on August 15 were cut to fit in 1-L bottles and were allowed to root in in sterile NFM for 11 days. For 15N experiments, samples were then cut into 5–8 cm sections, with samples from five of the seven trees healthy enough for experimentation.
15N2 dosing
Cylinders of compressed 15N2 isotope gas (98 atom %) were obtained from Sigma Aldrich (lot number SZ1670V). Following transfer by water displacement to serum bottles, the gas was treated with 1 ml of HCl (1N) in order to precipitate any possible trace amounts of ammonia.
Nitrogen-free hydroponics (15N2 Experiment 1)
Rooted apical cuttings were transferred to 125-ml flasks containing 25 ml NFM and sealed with screw top septum valve caps (Mininert). Three flasks each containing two rooted cuttings were used for each individual tree. Two of these flasks were dosed with 15N2 and one was not dosed. For example, 1–1 and 1–2 shared a dosed flask and 1–3 and 1–4 shared a dosed flask, and two undosed plant samples shared a flask as well. Plants were initially exposed to 1% 15N2 gas by removing 1 ml of air and replacing it with 1 ml 15N2 gas. After 4 days, the flasks were opened inside a sterile hood for fresh air exchange for 15 minutes before being re-dosed with 5 ml of 15N2 isotope gas. A week later the flasks were aired out again and the 25 ml of NFM was replaced with fresh medium. The plants were then dosed a final time with 5 ml of 15N2 isotope gas. After 4 more days of growth, the plants were removed and prepared for mass spectrometry analysis. Overall, the plants were exposed to an experimental atmosphere with a 15N atom percent excess of 6.17% for 2 weeks. To verify that CO2 levels were adequate in the 125-ml flasks for the plant experiments, surface-sterilized cuttings of wild poplar tree 4 were incubated as described above. CO2 concentrations were measured for two time points a week apart at the end of the light cycles (235 and 280 ppm) and at the end of the following dark cycles (325 and 361 ppm). These values indicated that the plants were actively photosynthesizing but that the system was closed such that adequate CO2 levels were maintained.
Nitrogen-free agar (15N2 Experiment 2)
For a longer study, agar rather than hydroponics, was used. Four cuttings of each of the samples from three of the poplar trees (Poplar 1, 5, and 6) were made and were transferred individually into 125 ml flasks containing 50 ml of NFMS agar. This allowed for 3 dosed and 1 undosed control plant from each tree. Plant samples were given an initial dose of 5 ml of 15N2 gas. After 8 days the flasks were opened and aired out in a sterile hood as in Experiment 1. A final dose of 5 ml 15N2 was given, and the plants were allowed to grow for 3 more weeks before being prepared for mass spectrometry as before. Throughout the experiment, the plants were exposed to an experimental atmosphere with a 15N atom percent excess of 6.61% for 1 month. During this one month experiment, several of the plants died including the undosed samples of Poplar1 and Poplar6 as well as one dosed sample each of Poplar1 and Poplar 5.
In vitro-propagated poplar in nitrogen-free hydroponics (15N2 Experiment 3)
After obtaining shipment of Nisqually-1 tissue culture plants (provided by Steve Strauss, Oregon State University), the plants were dosed as in Experiment 1 for the wild poplar. Three rooted in vitro-grown plants were transferred to 125-ml flasks with 25 ml NFM and exposed to the same 15N2 dosing regime as in Experiment 1. As 15N incorporation controls for this experiment, Saccharomyces cerevisiae, Azotobacter vinelandii [59], and Rahnella sp. WP5 [47] cultures were also tested using the same serum bottle of 15N2 gas. These microbial cells had average delta 15N values of 47 ‰, 5075 ‰, and 142 ‰, respectively. Sterility of in vitro-propagated Nisqually-1 plants was tested as follows: one of the rooted cuttings was weighed and homogenized in 5 ml NL-CCM per gram of tissue, and 100 μl of diluted and undiluted extract were plated on three types of media (MG/L, NL-CCM, and Nfb) and incubated at 30°C for three days. There was no visible microbial growth. In addition, genomic DNA was prepared from two aliquots of in vitro grown Nisqually-1 tissue and from soil-grown Nisqually-1 and subjected to PCR using primers that amplify differentially-sized 16S rDNA fragments for mitochondria (1090 bp) and bacteria (735 bp) [60]. While PCR of the soil-grown plant sample resulted in both bands, the two tissue culture plant samples had only the mitochondrial band (data not shown), supporting the hypothesis that these plants did not harbor bacteria.
Data from two hydroponic experiments with comparable conditions (Experiment 1 and 3) were pooled for ANOVA to test the differences in δ15N‰ between wild-dosed, sterile-dosed, and undosed (wild and sterile combined) plants. The δ15N‰ values from different parts (subsamples) within a plant were averaged and the means for individual plants were used to represent each source plant as an experimental unit (replicate). The least squares means (LSMEAN) were used to make pairwise comparisons between treatments using SAS software (ver. 9.4, SAS Institute, Cary, NC).
Isotope-Ratio Mass Spectrometry
Samples were flash frozen using liquid N2 and ground to a fine powder with a mortar and pestle. The samples were then transferred to aluminum weigh boats and dried at 75°C. Tin capsules (Costech) were prepared for analysis with 9–11 mg of tissue, and sent to the Alaska Isotope Lab at the University of Alaska-Fairbanks (http://ine.uaf.edu/werc/asif/) for elemental analysis isotope-ratio mass spectrometry (EA-IRMS).
Analysis of Isotope-Ratio Mass Spectrometry Data
Data from EA-IRMS was received in the form of δ15N‰ (at-air). For calculation:
Where
This can be rearranged to calculate the proportion of 15N to total N:
%Ndfa (Percent Nitrogen derived from atmosphere) is calculated [61]:
Where is the average value of the undosed control plants.
Acetylene Reduction Assay (ARA)
For the ARA, Poplar 6 branch samples were used immediately after the July 17, 2015 collection. Branches were cut into 1 gram (0.9–1.2 g) samples of the stem portions, six from each branch, were weighed and transferred aseptically into sterile, 20 ml round bottom, beveled edge glass vials containing 1 ml filter-sterilized NFM, and sealed with magnetic caps. For nine of the samples (three from each branch), 1.8 ml of headspace gas were removed and replaced with acetylene gas (99.6% purity, Praxair). Samples were incubated for 3d in a growth chamber (Percival CU41Lx) at 25°C with a 12 hour light cycle. Headspace gas was analyzed on a gas chromatograph (TRACE GC ULTRA, Thermo Scientific, Waltham, MA) equipped with a flame ionization detector (FID) and a HayeSep R column (2.6m x 1/8” x 2.0mm). The oven temperature was set at 70°C with a flow rate of 35 ml/min. We used high purity N2 (g) as the carrier gas, H2 (g) as the fuel gas ((H2 generator, Parker Dominic Hunter, Cleveland, OH), and high purity synthetic air as the oxidizing gas.
A standard curve of ethylene concentrations was generated and used to calculate ethylene production. Values were adjusted for small differences in sample mass.
Analysis of nitrogenase gene diversity within wild poplar
In parallel with the 15N2 experiments, the presence of endophytic nitrogenase gene sequences was assessed. Leaf and stem samples of surface-sterilized wild poplar trees 1–7 that had been collected in summer 2013 and grown in NFM were tested by PCR for the presence of putative diazotrophic bacteria. Polygenomic DNA was extracted using the MasterPure Plant Leaf DNA Purification Kit (Epicentre), following the manufacturer’s protocol except that 100 mg of plant tissue was used. PCR was performed using Populus ralf 6 primers [62] in Epicentre Fail-Safe PCR pre-mix G as a positive control for PCR. The nifH b1 primers [63] were used in Epicentre Pre-mix E to detect the presence of the nitrogenase subunit gene. Genomic DNA extracted from Azotobacter vinelandii was used as a positive control for nifH and no-DNA samples were included in both the ralf and nifH PCR experiments. Two additional surface-sterilized plants of wild poplar genotype 4 from the 125-ml flasks of NFM were analyzed for total diazotrophic bacteria diversity. Genomic DNA was extracted from the plants (included leaf, stem, and root together) using the MasterPure Plant Leaf DNA Purification Kit (Epicentre). PCR was performed using nifH b1 primers as described above. The nifH amplicons (351 bp) were cleaned using Exo-SAP-IT (Affimetrix) and tagged with Illumina sequencing primers with a unique ID for each sample following the standard Illumina protocol for amplicon library preparation. The libraries were then sequenced on an Illumina MiSeq sequencer using a 2x300 bp read kit. The resulting reads were processed in Geneious 7.1 (Biomatters Software LTD). The algorithm used was Geneious’ built-in “Trim and Filter” workflow with the following parameters—Error Probability limit 1%, trim the 5’ and 3’ ends, filter to retain sequences >350bp in length. Paired MiSeq reads were overlap aligned using PEAR [64] with the default settings. Based on the FASTQ values, 44,746 assembled reads were then trimmed to remove any base with an estimated error rate of greater than 1% and filtered for reads that were less than or equal to 350 bp post-trimming. This produced 6573 assembled high quality reads that spanned most of the nifH amplicon. A subsample of 192 of these were translated in all 6 reading frames and the correct reading frame for each read was determined by alignment against a nifH consensus sequence that spanned the amplicon region. The consensus sequence was created from the 2012 nifH database [65]. The translated sequences were then subjected to multiple sequence alignments along with nifH protein sequences in GenBank selected from known species and selected to span much of the nifH phylogenetic tree [66]. After alignment, a phylogenetic tree was constructed. Both the multiple sequence alignment and tree construction was done within Geneious using Geneious’ own alignment and tree building algorithms. Multiple sequence alignment was performed in Geneious using the MAFFT algorithm [67]. The default parameters were used (e.g. a BLOSUM62 scoring matrix, with a gap open penalty of 1.53 and an offset value of 0.123). Tree building was performed using the Geneious Tree Builder with the default parameters (e.g. a Juke-Cantor genetic distance model, and neighbor-joining with no outgrip). After tree construction, a representative sequence was selected from regions within the tree for which there was no nearby named reference sequence and searched against GenBank using BLAST [68] to identify additional reference sequences. These were then added to the alignment, and the alignment and tree was rebuilt.
Assessment of culturable endophytic population variability
A stem cutting harvested from wild Poplar 4 in August 2014 was surface-sterilized and maintained in NFM in a 1L glass vessel. A rooted shoot that grew from the original stem cutting was harvested in 25–50 mg sections and homogenized in 5 ml NL-CCM per gm tissue for one minute. A 1:1000 dilution of each extract was prepared and 100 μl were plated onto three types of medium with varied N levels and C sources to assess microbial populations: MG/L, NL-CCM, and Nfb. Colonies were counted after 2d and 4d at 30°C. In addition, the poplar samples used in the ARA in 2015 were examined similarly for culturable endophytic populations.
Results
15N2 Incorporation
A site along the Snoqualmie River, a natural river system in Western Washington State, was chosen for the collection of wild poplar cuttings because the water carries a very low nutrient load, and the cobble substrate is also nutrient-limited (Fig 1), yet poplar and willow plants thrive in this environment. Isotopic analysis of the poplar tissues from the two independent 15N2 labeling experiments demonstrated high levels of N2 fixation in some of the cuttings from wild plants, with several having δ 15N values over a 100 ‰ (Fig 2) and one as high as 284 ‰ (Fig 3). Overall, the 15N incorporation was higher in 15N2-dosed wild plants compared to undosed controls with a statistically highly significant difference (p < 0.01; pairwise comparison for H0: 15N dosed = undosed) (Fig 4a). In vitro propagated P. trichocarpa (clone Nisqually-1) plants were tested to verify that the 15N incorporation correlated with the presence of microbiota (Experiment 3). This clone was verified to be internally-sterile by microbiological and molecular tests (data not shown). The average δ 15N value was only 13.7 for these plants. Even when all the wild poplar hydroponic experiment data were combined, the values for 15N incorporation by wild poplar plants was significantly greater than that of these empirically sterile Nisqually-1 plants as well as that of un-dosed control plants (p < 0.05) (Fig 4a).
Fig 1. Study site along the Snoqualmie River in western Washington State.

Poplar and willow are the dominant plant species in this N-limited, cobble-dominated floodplain. Photographs of the site were taken in 2002, 2006, and 2015.
Fig 2. 15N incorporation in cuttings from wild poplar plants grown in hydroponics.

Cuttings from five independent plants were collected during the peak growing season in 2013. Rooted cuttings were exposed to 6.17% atom excess of 15N2 gas in N-free hydroponic medium for two weeks. Data from samples of the same plant were not averaged since the diazotrophic endophytes are not equally distributed throughout the plants.
Fig 3. 15N2 incorporation in cuttings from wild poplar plants grown in N-free agar.
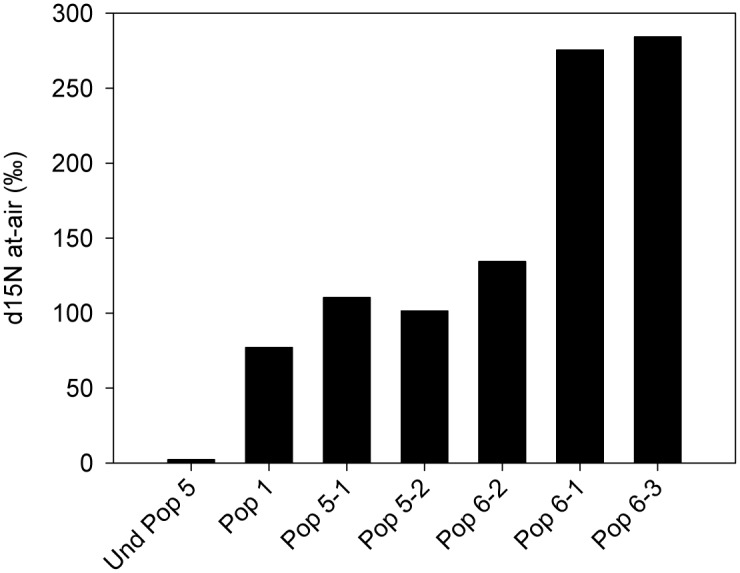
Cuttings from three independent wild plants (Poplar 1, 5, and 6) were collected during the peak growing season in 2013. Rooted cuttings were exposed to 6.61% atom excess of 15N2 gas for 4 weeks.
Fig 4.

(a) Combined data from the hydroponic experiments (Experiment 1 and Experiment 3) illustrate an overall significantly greater 15N incorporation by dosed wild poplar plants compared to undosed plants (p < 0.001). In vitro propagated Nisqually-1 plants showed an intermediate response that was significantly lower than dosed wild plants (p < 0.05) while only marginally greater than undosed wild plants (p = 0.08). In these experiments, rooted cuttings were exposed to 15N2 gas in N-free hydroponic medium (NFM) for two weeks. Bars annotated with the same letters are not significantly different from each other based on pairwise comparisons of least squares means at alpha = 0.05. Error bars represent one standard error. (b and c) Distributions of δ15N‰ data from dosed wild poplar cuttings tested in Experiment 1. The percent frequency plot (b) shows that about a half of dosed wild cuttings incorporated 15N at similarly low levels as the in vitro grown Nisqually-1 cuttings from Experiment 3 (vertical dashed line). The box plot of δ15N‰ grouped by source trees (c) suggests that the capacity to harbor N2 fixing endophytes is likely to vary not only between trees but also between locations within a single tree.
Given the variability of N2-fixation within the wild poplar plants, it is not possible to accurately extrapolate from the data to whole-tree estimates of N acquired through biological N2-fixation by the diazotrophic endophytes, but general estimates may be calculated. Levels of 15N in tissue suggest that during the two week period of exposure, up to 0.8% of total plant N was derived from atmospheric N2-fixation (Table 1, Poplar sample 4–3). As in other studies of endophytic N2-fixation, such rates of N accumulation per unit time are generally lower than those fixed in nodules associated with leguminous plants [7]. The poplar plants exposed to 15N2 for two weeks had a rate of N2-fixation of up to an average of 20.6 mg N/kg/day, if it is assumed that N2-fixation occurred uniformly throughout the 2-week period. We did not replace all the natural air with 15N2 and helium or argon as in other studies [18,31,69] but only spiked the headspace with the labeled gas. Given that the plant sizes were small, there was no visible growth of the plant sample during the assay, and as there may have been an enzymatic preference for the more abundant 14N in the system over 15N, the observed rate is likely an underestimate of the N2 fixation rate considering that under controlled greenhouse conditions, BNF in inoculated poplar was estimated to be 65% N [52].
Table 1. N2-fixation (%Ndfa) in poplar plants from five wild poplar trees.
| Sample | % Ndfa | Rate of N2-Fixation (mg/kg/day) |
|---|---|---|
| 1–1 | 0.087 | 2.70 |
| 1–2 | 0.089 | 3.01 |
| 1–3 | 0.031 | 0.75 |
| 1–4 | 0.102 | 2.21 |
| 1–5 UNDOSED | BDLa | BDL |
| 2–1 | 0.292 | 13.2 |
| 2–2 | 0.101 | 4.55 |
| 2–3 | 0.081 | 3.04 |
| 2–4 | 0.040 | 1.48 |
| 2–5 UNDOSED | BDL | BDL |
| 4–1 | 0.103 | 5.74 |
| 4–2 | 0.077 | 2.91 |
| 4–3 | 0.761 | 20.6 |
| 4–4 | 0.529 | 14.3 |
| 4–5 UNDOSED | BDL | BDL |
| 5–1 | 0.168 | 5.54 |
| 5–2 | 0.689 | 14.6 |
| 5–3 | 0.241 | 11.4 |
| 5–4 | 0.276 | 11.1 |
| 5–5 UNDOSED | BDL | BDL |
| 6–1 | 0.041 | 2.08 |
| 6–2 | 0.038 | 1.21 |
| 6–3 | 0.218 | 10.0 |
| 6–4 | 0.474 | 16.9 |
| 6–5 UNDOSED | BDL | BDL |
Nitrogen derived from a 15N-enriched atmosphere, %Ndfa represents the proportion of total N content originating from atmospheric N-fixation, calculated using the average of the undosed samples as a reference value. Rate of N2 fixation is measured as mg dry weight of newly fixed N per kg of total plant tissue per day. Data were from Expt. 1.
a BDL, Below detection limits
Assessment of the diazotrophic endophytic population
Variable degrees of N2 fixation within cuttings from the same plant could be due to N2-fixing endophytes being unevenly distributed throughout the plant. Although the hypothesis that specific microorganisms were required for effective N2-fixation could not be tested directly at this time, we confirmed the general presence of diazotrophic endophytes in all the wild poplar trees used in the studies by conducting PCR of the genomic DNA to amplify the nitrogenase subunit gene, nifH. All of the wild poplar plant samples were positive for the presence of the nifH gene (S1 Fig) indicating that endophytic or closely associated diazotrophic microorganisms were present. To assess the culturable endophytic population within the wild poplar cuttings, we homogenized three surface-sterilized samples that were of similar size to those used in the 15N2 assays and then plated the extracts on N-limited medium. The total population density ranged from 5.1 x 106 to 1.9 x 107 CFU/g. Sequencing of the nifH amplicons from one of the wild poplar trees revealed a diversity of diazotrophic taxa (S2 Fig). These included nifH sequences from genera we had previously isolated and studied including Burkholderia and Sphingomonas but also others such as Azospirillum brasilense and Bradyrhizobium sp., photosynthetic diazotrophs such as Rhodospirillum rubrum, Rhodobacter capsulatus and Nostoc punctiforme, and Archaeal species including Methanococcus maripaludis and Methanosarcina acetivoran. Many of the other sequences most closely matched nifH sequences of uncultured bacteria in the database. A detailed assessment of the microbiome of these poplar trees is underway (U.S. Department of Energy Joint Genome Institute).
Having confirmed that the wild poplar trees harbored diazotrophic microbial species, we assessed microbial diversity within different sections of the same tree. Wild Poplar 4 was chosen for this study since, in the previous growing season, some of the cuttings of this tree had exhibited high 15N incorporation (samples 4–3 and 4–4) while others had exhibited no significant incorporation (samples 4–1 and 4–2). Both the cultured endophytic population density (Table 2) and composition (Fig 5) varied greatly in different sections of the same plant. For example, the colony forming units (CFU) on N-limited medium differed by 2 orders of magnitude in closely adjacent stem samples (samples 3 and 4). The colony morphology differed considerably (Fig 5) suggesting that species diversity was different even in adjacent samples of the same tissue.
Table 2. Variability of the culturable endophytic population of Wild Poplar 4.
| Sample | Tissue | CFU on MG/L | CFU on NL-CCM | CFU on Nfb |
|---|---|---|---|---|
| 1 | Root | 2 x 107 | 1.7 x 107 | 8.2 x 106 |
| 2 | Root | 3.4 x 107 | TNTC | 2.0 x 107 |
| 3 | Stem | 5.7 x 106 | 1.3 x 107 | 2.9 x 106 |
| 4 | Stem | 4.0 x 105 | 7.5 x 105 | 4.5 x 105 |
| 5 | Stem | 3.0 x 105 | 7.0 x 105 | 3.5 x 105 |
| 6 | Stem | 2.9 x 106 | 5.5 x 105 | 1.1 x 106 |
| 7 | Leaf | 6.4 x 106 | 4.3 x 106 | 1.1 x 106 |
| 8 | Leaf | 1.5 x 107 | 1.7 x 107 | 5.7 x 106 |
| 9 | Leaf | 1.0 x 106 | 7.0 x 105 | 2.0 x 105 |
| 10 | Leaf | TNTC | 1.4 x 106 | 4.5 x 105 |
Fig 5. Variability in endophytic population and composition in four stem sections of wild poplar genotype 4, each extracted with 5 ml/gm.

Extracts were diluted 1:1000 and 100 μl plated on N-limited combined carbon medium (NL-CCM). Plates were photographed after 4 days at 30°C.
Culturable endophytes were extracted in 5 ml NL-CCM per gm of tissue. Extracts were diluted 1:1000 and 100 μl were plated on three types of medium. The number of colony forming units (CFU) was assessed after 4 days. TNTC, too numerous to count.
Acetylene reduction assay
As a further test of N2 fixation in wild poplar, we used the acetylene reduction assay [54]. This commonly used indirect measure of N2 fixation has been used to study endophytic and associative N2-fixation, and relies on the ability of the nitrogenase enzyme to reduce acetylene gas to ethylene which can be measured by gas chromatography [18,25,70–72]. We chose Wild Poplar 6 for this study as it had the highest level of N2-fixation (Fig 3) although this level was variable (Fig 2). Branch samples from Wild Poplar 6 were collected in the following growing season, surface-sterilized, and transferred to GC vials. Half of the samples were dosed with acetylene, and incubated for three days. The undosed stem samples produced an average of only 5 mmol ethylene per gm (Table 3), indicating the background levels of ethylene production by the plants. Of the nine dosed samples, three had levels of ethylene production that were 6, 12, and 21-fold higher than the averaged undosed samples. The other six samples had values comparable to the undosed samples.
Table 3. Acetylene reduction assay results of Wild Poplar 6 stem samples.
| Plant Sample | mmol ethylene per gram |
|---|---|
| Undosed 1 | 4.84 |
| Undosed 2 | 3.38 |
| Undosed 3 | 4.50 |
| Undosed 7 | 4.52 |
| Undosed 8 | 5.01 |
| Undosed 9 | 3.52 |
| Undosed 13 | 4.42 |
| Undosed 14 | 7.34 |
| Undosed 15 | 8.46 |
| Dosed 21 | 20.63 |
| Dosed 22 | 61.42 |
| Dosed 23 | 6.19 |
| Dosed 27 | 8.52 |
| Dosed 28 | 5.76 |
| Dosed 29 | 9.05 |
| Dosed 33 | 5.69 |
| Dosed 34 | 4.24 |
| Dosed 35 | 106.9 |
Samples were incubated for 3d before headspace analysis by gas chromatography. Ethylene was calculated using a standard curve and adjusted for plant mass.
The high variability in N2-fixation in wild poplar, even within the same tree, was therefore determined using two independent assays in two different growing seasons. The samples used in the ARA were tested for culturable endophyte populations at the end of the experiment. The population densities were highly variable in the different stem sections (Fig 6). There was no correlation between overall number of culturable bacteria and ARA values.
Fig 6. Variability of the culturable endophytic population of wild poplar genotype 6 stem samples used in the acetylene reduction assay experiment.

CFU, colony forming units.
Discussion
The data presented here represent the first direct demonstration that N2-fixation is possible within wild poplar trees and the first demonstration that a varied diazotrophic microbial population exists within individual poplar sections. Further studies are needed to localize in planta active diazotrophic endophytes and the dynamics of N transfer between these potentially symbiotic partners. The variation in N2-fixation amongst different cuttings of the same tree alludes to the intriguing possibility that there may be microbial social requirements that limit effective N2 fixation to particular groupings within the plant that have achieved a threshold of density [73] or diazotrophic species composition. Since the ARA results did not correlate with the overall numbers of cultured bacteria, it is likely that there are either specific diazotrophic strains that are required for effective in planta N2-fixation or that these strains were not culturable on the media tested. This population of diazotrophs would be of great importance but would not be seen in our CFU counts. Using nifH gene expression analysis in sugarcane, it was shown that rhizobial species rather than the dominant cultured endophytic species were likely the primary contributors of N [74,75]. Likewise, a metagenomic study of the endophytic community in rice revealed that there was a high apparent density of N-fixing endophytes but that the dominant nifH expressed was only that of Rhizobium [8]. Molecular studies are required to identify the active diazotrophic species and their density requirements for effective N2-fixation in vivo in Populus. Such studies including fluorescent in situ nifH hybridization and nanoscale secondary ion mass spectrometry (NanoSIMS) were recently approved by the U.S. Department of Energy Joint Genome Institute and will begin in 2016.
While there have been several studies quantifying N2-fixation in grasses, the variability in N2-fixation such as we have encountered in Populus has not been reported. The structure of poplar trees, and eudicots in general, is much more complex than that of monocots. The trunk and branches, internodes and nodes, leaves and petioles of eudicots—all provide unique habitats that may lead to a highly variable microbiota occupying the different niches. Each of these compartments is likely influenced within trees by its location in the canopy, the sun exposure, and the age of the leaf or branch. There are multiple factors involved in microbial community assembly including plant genotype, environment, microbe-microbe interactions [76] and the season [77]. Plants seem to have some influence over the microbial population of the rhizosphere [78] as well as the endosphere [44]. There may also be a general selection for particular functional traits [79]. In the N-limited site used in this study, diazotrophy is likely a selected trait. The culturable endophytic population of the poplar and willow trees is dominated by diazotrophic bacteria of many different species [47]. A culture-independent assessment of the microbiome of the leaves of one poplar tree revealed a high percentage of Burkholderia (24%) and Sphingomonas (16%) (unpublished data), genera that include many N-fixing species. In the nifH analysis presented here, a broad range of diazotrophic species was represented including the well-studied plant-associated diazotrophs, Azospirillum brasilense and Bradyrhizobium sp. Where the different strains ultimately reside within the available niches of an entire tree, and in what microbial communities and densities, are likely not homogeneous throughout the plant. Therefore, unlike the legume-rhizobium symbiosis in which the N2-fixing bacteria are housed in specific nodule structures, diazotrophic endophytic bacteria are unevenly distributed, and could lead to variation in N2-fixation in different areas of trees.
The 15N incorporation assay is viewed as the most direct method for assessing N2-fixation compared to the 15N dilution assay or isotopic differences in natural abundance. A criticism of the technique, however, is that there is possible contamination of 15N-labeled ammonia or nitrate in the 15N2 gas [80]. Treatment of the gas with HCl precipitates out any contaminating ammonia, and since only the gas is delivered to the samples, any contaminating nitrate would not be present. Since the sterile plants had the lowest 15N incorporation and since plants in the same flask (Expt 1) had different 15N incorporation values, the HCl treatment was effective in removing any possible contamination. The ARA is commonly used to qualitatively assess endophytic and epiphytic N2-fixation but can have limitations [81]. One concern is that ethylene, the indicator of N2-fixation in this assay, is produced by the plant. However, by testing equal numbers of undosed and acetylene-dosed samples, a baseline can be established. Another method for testing for BNF is through the 15N natural abundance assay. This assay relies upon a comparison to substrate and to “non-fixing” controls. In our low-nutrient, cobble-dominated study site, however, the substrate is rocky and the only plants present are the poplar and willow, making this assay untenable.
The possibility of biological N2-fixation by endophytes has faced criticism because the nitrogenase enzyme is inhibited by oxygen and is therefore thought to require the specialized structure of root nodules. However, since free-living diazotrophic bacteria evolved long before legumes or actinorhizal plants [6], this criticism cannot be justified. N2-fixing bacteria can utilize a range of mechanisms to protect the nitrogenase enzyme complex [5,6]. Trichodesmium, for example, is an ancient marine diazotrophic cyanobacterium that fixes N2 effectively without the specialized heterocyst cells common to some other cyanobacteria species [82]. In this bacterium, oxygenic photosynthesis and N2-fixation operate concurrently during the day through temporal and spatial segregation and a reduction of photosynthetically-evolved oxygen [83] to solve the so-called “oxygen paradox” [5,84]. While the nitrogenase enzyme is inhibited by oxygen, the exceptionally high energy demands of N2-fixation often require oxidative phosphorylation for maximum ATP generation. By rapidly utilizing sugars, intracellular oxygen can be depleted or at least sufficiently reduced. The free-living, rhizospheric diazotroph, Azotobacter vinelandii, for example, can employ this respiratory protection mechanism as well as conformational protection, avoidance and spatial separation [59]. Endophytes may use a variety of strategies to achieve in planta N2-fixation. One method could be through migrating to a microhabitat that has optimal oxygen levels [85]. nifH gene expression is regulated by oxygen, and has been shown to be expressed in planta by endophytes and associated bacteria, thus demonstrating that permissive microhabitats are available or that metabolic conditions have otherwise been met [11–14]. In addition, since some of the nifH sequences of wild poplar matched those of anaerobic Archaeal species, there are presumably niches within the plant tissue that are anaerobic. Exopolysaccharide production is a common trait in endophytes [86–89], and could possibly provide the necessary microaerobic environment for N2-fixation inside biofilms. More research is necessary to elucidate the specific mechanisms used by diazotrophic endophytes to protect the nitrogenase complex.
Populus species have long been known as early-successional trees able to colonize highly disturbed sites. We have previously shown that poplar and willow at this study site host a variety of microorganisms within branch tissues, some of which are capable of fixing N2 in vitro. Inoculation of cultivated poplar plants with these wild poplar endophytes increased N2 fixation [52]. Addition of the wild poplar endophytes to grasses and crop species (eudicots as well as monocots) increased plant growth in N-limited conditions [48–51]. The host range also encompassed gymnosperms, providing increased biomass under N-limited conditions to the commercially important forest tree, Douglas-fir (Pseudotsuga menziesii) [90]. These cross-host studies strongly support the hypothesis that the diazotrophic endophytes of wild poplar can provide significant amounts of N to the plant host. Other research has also given indirect evidence of N2 fixation in poplar. Metabolic profiling of hybrid poplar inoculated with a Paenibacillus strain from within micropropagated poplar suggested N2 fixation [91]. In comparison to uninoculated controls, the inoculated poplar had increased levels of asparagine and urea, which suggests increased N assimilation. Other research in the early 1980’s indicated that N2 fixation may be occurring within poplar. For example, wetwood samples of eastern cottonwood (Populus deltoides) tested positive with the acetylene reduction assay (ARA), and the degree of acetylene reduction increased when glucose was added and decreased when NH4Cl was added, demonstrating that this activity was regulated as expected [92]. Wood samples of black cottonwood (P. trichocarpa) from four felled trees also tested positive with the ARA [93]. Our results provide the first direct evidence of N2-fixation within wild poplar. The variability in both microbial composition and concentration points to the need for a better understanding of endophytic colonization and its relation to effective N2-fixation. As poplar was the first tree genome to be sequenced [94] and given that genomic sequencing of the first diazotrophic endophytes isolated from poplar is now underway, the poplar-endophyte symbiosis could serve as a model system for studying N2-fixation in trees at the molecular level.
One of the next great challenges will be maximizing food and biomass production in a sustainable way, and innovative new technologies must be developed. While one approach has been to attempt engineering of nodulation into crop plants, a simpler approach is evident [95,96]. Tailoring of the microbiome of plants can increase plant growth with reduced inputs. Although inoculation of crop plants with specific diazotrophic strains can contribute significant levels of N, it is often still necessary to apply some N fertilizer [97]. Further research is required to identify the best inoculants, especially for specific plant genotypes [98]. Considering that long-lived trees in natural, nutrient-limited environments have had the longest time and the greatest need to select beneficial microbial strains from their environment, such pioneer tree species may therefore be the best source for effective inoculants. The microbiota of wild poplar trees helps an exceptionally broad range of plant species, from grasses [48,51] to conifers [90], to overcome nutrient deficiencies. A deeper understanding of the microbiome of this non-nodulated early successional pioneer plant species may not only provide insight on how to optimize this species' biomass production, but also provide critical insights into how microbial endophytes can increase production of other bioenergy plant species and agricultural crops while at the same time reducing reliance on chemical fertilizers.
Supporting Information
Leaf and stem samples of surface-sterilized wild poplar genotypes 1–7 that had been collected in summer 2013 and grown in NFM were tested by PCR for the presence of putative diazotrophic bacteria. PCR was performed using Populus ralf 6 primers as a positive control (upper panel). The nifH b1 primers [63] were used to detect the presence of nitrogenase (lower panel). Genomic DNA extracted from Azotobacter vinelandii was used as a positive control for nifH. The arrow indicates the nifH gene product. S, stem; L, leaf. Lanes 1, 1Kb Plus DNA ladder; Lanes 2, no-DNA controls; Lane 3, Pop1S; Lanes 4, Pop1L; Lanes 5, Pop2S; Lanes 6, Pop2L; Lanes 7, Pop3S; Lanes 8, Pop3S.2; Lanes 9, Pop3L; Lanes 10, Pop4S; Lanes 11, Pop4L; Lanes 12, Pop5S; Lanes 13, Pop5L; Lanes 14, Pop6S; Lanes 15, Pop6L; Lanes 16, Pop7S; Lanes 17, Pop7L; Lanes 18, empty; Lanes 19, Azotobacter vinelandii.
(TIF)
Polygenomic (plant and endophytic) DNA was isolated from two rooted cuttings of Wild Poplar 4. A subset of the branches were labeled of those with similarity 90% or above in GenBank. Unlabeled branches indicate sequences with closest matches to nifH genes of uncultured bacteria in Gene Bank. The three colored groups represent Group 1 (black), Group II (red) and Group III (blue) nifH sequences as per [66].
(TIF)
Acknowledgments
This research was funded by the USDA-AFRI grant # 2011-68005-30407 and the Byron and Alice Lockwood Endowed Professorship. We are grateful to Steve Strauss (Oregon State University) for providing the sterile Nisqually-1 plants from tissue culture, to Alex Batson for assistance in plant sample processing, and to Amanda Bidwell for assistance with the acetylene reduction assay. We thank Prof. Emeritus Thomas Hinckley for helpful editorial comments.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was funded by United States Department of Agriculture-National Institute of Food and Agriculture-Agriculture and Food Research Initiative grant # 2011-68005-30407 to SLD ZK; and a Byron and Alice Lockwood Endowed Professorship to SLD AWS NDF. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Bulgarelli D, Schlaeppi K, Spaepen S, Ver Loren van TE, Schulze-Lefert P (2013) Structure and functions of the bacterial microbiota of plants. Annu Rev Plant Biol 64: 807–838. 10.1146/annurev-arplant-050312-120106 [DOI] [PubMed] [Google Scholar]
- 2.Khan Z, Doty SL (2011) Endophyte-assisted phytoremediation. Current Topics in Plant Biology 12: 97–105. [Google Scholar]
- 3.Rodriguez RJ, White JF Jr, Arnold AE, Redman RS (2009) Tansley Review: Fungal endophytes: diversity and functional roles. New Phytol 182: 314–330. [DOI] [PubMed] [Google Scholar]
- 4.Vandenkoornhuyse P, Quaiser A, Duhamel M, Le VA, Dufresne A (2015) The importance of the microbiome of the plant holobiont. New Phytol 206: 1196–1206. 10.1111/nph.13312 [DOI] [PubMed] [Google Scholar]
- 5.Marchal K, Vanderleyden J (2000) The "oxygen paradox" of dinitrogen-fixing bacteria. Biol Fertil Soils 30: 363–373. [Google Scholar]
- 6.Boyd ES, Costas AM, Hamilton TL, Mus F, Peters JW (2015) Evolution of molybdenum nitrogenase during the transition from anaerobic to aerobic metabolism. J Bacteriol 197: 1690–1699. JB.02611-14 [pii]; 10.1128/JB.02611-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Santi C, Bogusz D, Franche C (2013) Biological nitrogen fixation in non-legume plants. Ann Bot 111: 743–767. mct048 [pii]; 10.1093/aob/mct048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sessitsch A, Hardoim P, Doring J, Weilharter A, Krause A, Woyke T et al. (2012) Functional characteristics of an endophyte community colonizing rice roots as revealed by metagenomic analysis. Mol Plant Microbe Interact 25: 28–36. 10.1094/MPMI-08-11-0204 [DOI] [PubMed] [Google Scholar]
- 9.James EK, Olivares FL (1998) Infection and colonization of sugarcane and other graminacious plants by endophytic diazotrophs. Crit Rev Plant Sci 17: 77–119. [Google Scholar]
- 10.Gussin GN, Ronson CW, Ausubel FM (1986) Regulation of nitrogen fixation genes. Annu Rev Genet 20: 567–591. 10.1146/annurev.ge.20.120186.003031 [DOI] [PubMed] [Google Scholar]
- 11.Olivares FL, James EK, Baldani JI, Dobereiner J (1997) Infection of Mottled Stripe Disease-Susceptible and Resistant Sugar Cane Varieties by the Endophytic Diazotroph Herbaspirillum. New Phytologist 135: 723–737. [Google Scholar]
- 12.Hurek T, Egener T, Reinhold-Hurek B (1997) Divergence in nitrogenases of Azoarcus spp., Proteobacteria of the beta subclass. J Bacteriol 179: 4172–4178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Iniguez AL, Dong Y, Triplett EW (2004) Nitrogen fixation in wheat provided by Klebsiella pneumoniae 342. Mol Plant Microbe Interact 17: 1078–1085. [DOI] [PubMed] [Google Scholar]
- 14.Chelius MK, Triplett EW (2000) Immunolocalization of dinitrogenase reductase produced by Klebsiella pneumoniae in association with Zea mays L. Appl Environ Microbiol 66: 783–787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hurek T, Handley LL, Reinhold-Hurek B, Piche Y (2002) Azoarcus grass endophytes contribute fixed nitrogen to the plant in an unculturable state. Mol Plant Microbe Interact 15: 233–242. [DOI] [PubMed] [Google Scholar]
- 16.Welbaum GE, Sturz AV, Dong Z, Nowak J (2003) Endophytic nitrogen fixation in sugarcane: present knowledge and future applications. Plant Soil 252: 139–149. [Google Scholar]
- 17.Peng G, Zhang W, Luo H, Xie H, Lai W, Tan Z (2009) Enterobacter oryzae sp. nov., a nitrogen-fixing bacterium isolated from the wild rice species Oryza latifolia. Int J Syst Evol Microbiol 59: 1650–1655. 10.1099/ijs.0.65484-0 [DOI] [PubMed] [Google Scholar]
- 18.Elbeltagy A, Nishioka K, Sato T, Suzuki H, Ye B, Hamada T et al. (2001) Endophytic colonization and in planta nitrogen fixation by a Herbaspirillum sp. isolated from wild rice species. Appl Environ Microbiol 67: 5285–5293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Montanez A, Abreu C, Gill PR, Hardarson G, Sicardi M (2009) Biological nitrogen fixation in maize (Zea mays L.) by N-15 isotope-dilution and identification of associated culturable diazotrophs. Biology and Fertility of Soils 45: 253–263. [Google Scholar]
- 20.Rout ME, Chrzanowski TH (2009) The invasive Sorghum halepense harbors endophytic N2-fixing bacteria and alters soil biogeochemistry. Plant and Soil 315: 163–172. [Google Scholar]
- 21.Davis SC, Parton WJ, Dohleman FG, Smith CM, Del Grosso S, Kent AD et al. (2010) Comparitive biogeochemical cycles of bioenergy crops reveal nitrogen-fixation and low greenhouse gas emissions in a Miscanthus x giganteus agro-ecosystem. Ecosystems 13: 144–156. [Google Scholar]
- 22.Videira SS, Pereira e Silva MC, Galisa PS, Dias ACF, Nissinen R, Divan VLB et al. (2013) Culture-independent molecular approaches reveal a mostly unknown high diversity of active nitrogen-fixing bacteria associated with Pennisetum purpureum—a bioenergy crop. Plant Soil 373: 737–754. [Google Scholar]
- 23.Lopez BR, Tinoco-Ojanguren C, Bacilio M, Mendoza A, Bashan Y (2012) Endophytic bacteria of the rock-dwelling cactus Mammillaria fraileana affect plant growth and mobilization of elements from rocks. Environ Exper Botany 81: 26–36. [Google Scholar]
- 24.Reiter B, Burgmann H, Burg K, Sessitsch A (2003) Endophytic nifH gene diversity in African sweet potato. Canadian Journal of Microbiology 49: 549–555. [DOI] [PubMed] [Google Scholar]
- 25.Terakado-Tonooka J, Fujihara S, Ohwaki Y (2012) Possible contribution of Bradyrhizobium on nitrogen fixation in sweet potatoes. Plant and Soil 367: 639–650. [Google Scholar]
- 26.DeLuca TH, Zackrisson O, Gundale MJ, Nilsson MC (2008) Ecosystem feedbacks and nitrogen fixation in boreal forests. Science 320: 1181 320/5880/1181 [pii]; 10.1126/science.1154836 [DOI] [PubMed] [Google Scholar]
- 27.Dalton DA, Kramer S, Azios N, Fusaro S, Cahill E, Kennedy C (2004) Endophytic nitrogen fixation in dune grasses (Ammophila arenaria and Elymus mollis) from Oregon. FEMS Microbiol Ecol 49: 469–479. 10.1016/j.femsec.2004.04.010 [DOI] [PubMed] [Google Scholar]
- 28.Jimenez-Salgado T, Fuentes-Ramirez LE, Tapia-Hernandez A, Mascarua-Esparza MA, Martinez-Romero E, Caballero-Mellado J (1997) Coffea arabica L., a new host plant for Acetobacter diazotrophicus, and isolation of other nitrogen-fixing acetobacteria. Applied and Environmental Microbiology 63: 3676–3683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bal A, Chanway CP (2012) Evidence of nitrogen fixation in lodgepole pine inoculated with diazotrophic Paenibacillus polymyxa. Botany 90: 891–896. [Google Scholar]
- 30.Carrell AA, Frank AC (2014) Pinus flexilis and Picea engelmannii share a simple and consistent needle endophyte microbiota with a potential role in nitrogen fixation. Front Microbiol 5: 333 10.3389/fmicb.2014.00333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Sevilla M, Burris RH, Gunapala N, Kennedy C (2001) Comparison of benefit to sugarcane plant growth and 15N2 incorporation following inoculation of sterile plants with Acetobacter diazotrophicus wild-type and Nif- mutant strains. Molecular Plant Microbe Interactions 14: 358–366. [DOI] [PubMed] [Google Scholar]
- 32.Boddey RM, Dobereiner J (1988) Nitrogen fixation associated with grasses and cereals: Recent results and perspectives for future research. Plant and Soil 108: 53–65. [Google Scholar]
- 33.Urquiaga S, Xavier RP, de Morais RF, Batista RB, Schultz N, Leite JM et al. (2012) Evidence from field nitrogen balance and 15N natural abundance data for the contribution of biological N2 fixation to Brazilian sugarcane varieties. Plant and Soil 356: 5–21. [Google Scholar]
- 34.de Morais RF, Quesada DM, Reis VM, Urquiaga S, Alves BJR, Boddey RM (2012) Contribution of biological nitrogen fixation to Elephant grass (Pennisetum purpureum Schum.). Plant and Soil 356: 23–34. [Google Scholar]
- 35.Beatty PH, Good AG (2011) Plant science. Future prospects for cereals that fix nitrogen. Science 333: 416–417. 333/6041/416 [pii]; [DOI] [PubMed] [Google Scholar]
- 36.Pankievicz VC, do Amaral FP, Santos KF, Agtuca B, Xu Y, Schueller MJ et al. (2015) Robust biological nitrogen fixation in a model grass-bacterial association. Plant J 81: 907–919. [DOI] [PubMed] [Google Scholar]
- 37.Moyes AB, Kueppers LM, Pett-Ridge J, Carper DL, Vandehey N, O'Neil J et al. (2016) Evidence for foliar endophytic nitrogen fixation in a widely distributed subalpine conifer. New Phytologist in press. [DOI] [PubMed] [Google Scholar]
- 38.Isebrands JG, Aronsson P, Carlson M, Ceulemans R, Coleman M, et al. (2014) Environmental Applications of Poplars and Willows In: Isebrands JG, Richardson J, editors. Poplars and Willows. Rome: The Food and Agriculture Organization of the United Nations; pp. 258–336. [Google Scholar]
- 39.Stettler RF, Bradshaw HD, Heilman PE, Hinckley TM (1996) Biology of Populus and its Implications for Management and Conservation. Ottawa: NRC Research Press. [Google Scholar]
- 40.Weyens N, Truyens S, Dupae J, Newman L, Taghavi S, van der Lelie D et al. (2010) Potential of the TCE-degrading endophyte Pseudomonas putida W619-TCE to improve plant growth and reduce TCE phytotoxicity and evapotranspiration in poplar cuttings. Environ Pollut 158: 2915–2919. S0269-7491(10)00223-X [pii]; 10.1016/j.envpol.2010.06.004 [DOI] [PubMed] [Google Scholar]
- 41.Kang JW, Khan Z, Doty SL (2012) Biodegradation of trichloroethylene (TCE) by an endophyte of hybrid poplar. Appl Environ Microbiol 78: 3504–3507. 10.1128/AEM.06852-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Khan Z, Roman D, Kintz T, delas Alas M, Yap R, Doty SL (2014) Degradation, phytoprotection, and phytoremediation of phenanthrene by endophyte Pseudomonas putida PD1. Appl Environ Microbiol 48: 12221–12228. [DOI] [PubMed] [Google Scholar]
- 43.Ulrich K, Ulrich A, Ewald D (2008) Diversity of endophytic bacterial communities in poplar grown under field conditions. FEMS Microbiol Ecol 63: 169–180. 10.1111/j.1574-6941.2007.00419.x [DOI] [PubMed] [Google Scholar]
- 44.Gottel NR, Castro HF, Kerley M, Yang Z, Pelletier DA, Podar M et al. (2011) Distinct microbial communities within the endosphere and rhizosphere of Populus deltoides roots across contrasting soil types. Appl Environ Microbiol 77: 5934–5944. AEM.05255-11 [pii]; 10.1128/AEM.05255-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Stettler R. F. (2009) Cottonwood and the River of Time: on Trees, Evolution, and Society. Seattle: University of Washington Press. [Google Scholar]
- 46.Doty SL, Dosher MR, Singleton GL, Moore AL, van Aken B, Stettler RF et al. (2005) Identification of an endophytic Rhizobium in stems of Populus. Symbiosis 39: 27–36. [Google Scholar]
- 47.Doty SL, Oakely B, Xin G, Kang JW, Singleton G, Khan Z et al. (2009) Diazotrophic endophytes of native black cottonwood and willow. Symbiosis 47: 23–33. [Google Scholar]
- 48.Xin G, Zhang G, Kang JW, Staley JT, Doty SL (2009) A diazotrophic, indole-3-acetic acid-producing endophyte from wild cottonwood. Biology and Fertility of Soils 45: 669–674. [Google Scholar]
- 49.Knoth J, Kim S-H, Ettl G, Doty SL (2013) Effects of cross host species inoculation of nitrogen-fixing endophytes on growth and leaf physiology of maize. GCB Bioenergy 5: 408–418. [Google Scholar]
- 50.Kandel S, Herschberger N, Kim S-H, Doty SL (2015) Diazotrophic endophytes of poplar and willow promote growth of rice plants in nutrient-limited conditions. Crop Science 55: 1765–1772. [Google Scholar]
- 51.Khan Z, Guelich G, Phan H, Redman RS, Doty SL (2012) Bacterial and yeast endophytes from poplar and willow promote growth in crop plants and grasses. ISRN Agronomy 10.5402/2012/890280 [DOI] [Google Scholar]
- 52.Knoth JL, Kim SH, Ettl GJ, Doty SL (2014) Biological nitrogen fixation and biomass accumulation within poplar clones as a result of inoculations with diazotrophic endophyte consortia. New Phytol 201: 599–609. 10.1111/nph.12536 [DOI] [PubMed] [Google Scholar]
- 53.Pinto-Tomas AA, Anderson MA, Suen G, Stevenson DM, Chu FS, Cleland WW et al. (2009) Symbiotic nitrogen fixation in the fungus gardens of leaf-cutter ants. Science 326: 1120–1123. 326/5956/1120 [pii]; 10.1126/science.1173036 [DOI] [PubMed] [Google Scholar]
- 54.Schollhorn R, Burris RH (1967) Acetylene as a competitive inhibitor of N-2 fixation. Proc Natl Acad Sci U S A 58: 213–216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiology of the Plant 15: 473–497. [Google Scholar]
- 56.Cangelosi GA, Best EA, Martinetti G, Nester EW (1991) Genetic analysis of Agrobacterium. Meth Enzymology 204: 384–397. [DOI] [PubMed] [Google Scholar]
- 57.Rennie RJ (1981) A single medium for the isolation of acetylene-reducing (dinitrogen-fixing) bacteria from soils. Can J Microbiol 27: 8–14. [DOI] [PubMed] [Google Scholar]
- 58.Baldani VLD, Dober (1980) Host plant specificity in the infection of cereals with Azospirillum sp.p. Soil Biol Biochem 12: 433–439. [Google Scholar]
- 59.Flores-Encarnacion M, Contreras-Zentella M, Soto-Urzua L, Aguilar GR, Baca BE, Escamilla JE (1999) The respiratory system and diazotropic activity of Acetobacter diazotrophicus PAL5. Journal of Bacteriology 181: 6987–6995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Chelius MK, Triplett EW (2001) The diversity of archaea and bacteria in association with the roots of Zea mays L. Microb Ecol 41: 252–263. [DOI] [PubMed] [Google Scholar]
- 61.Busse MD, Jurgensen MF, Page-Dumroese DS, Powers RF (2007) Contribution of actinorhizal shrubs to site fertility in a Northern California mixed pine forest. Forest Ecol Management 244: 68–75. [Google Scholar]
- 62.Haruta M, Constabel CP (2003) Rapid alkalinization factors in poplar cell cultures. Peptide isolation, cDNA cloning, and differential expression in leaves and methyl jasmonate-treated cells. Plant Physiol 131: 814–823. 10.1104/pp.014597 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Burgmann H, Widmer F, Sigler WV, Zeyer J (2004) New molecular screening tools for the analysis of free-living diazotrophs in soil. Appl Environ Microbiol 70: 240–247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Zhang J, Kobert K, Flouri T, Stamatakis A (2014) PEAR: a fast and accurate Illumina paired-end reAd mergeR. Bioinformatics 30: 614–620. 10.1093/bioinformatics/btt593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Gaby JC, Buckley DH (2012) A comprehensive evaluation of PCR primers to amplify the nifH gene of nitrogenase. PLoS One 7: e42149 10.1371/journal.pone.0042149; PONE-D-11-08927 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Raymond J, Siefert JL, Staples CR, Blankenship RE (2004) The natural history of nitrogen fixation. Mol Biol Evol 21: 541–554. 10.1093/molbev/msh047; msh047 [pii]. [DOI] [PubMed] [Google Scholar]
- 67.Katoh K, Misawa K, Kuma K, Miyata T (2002) MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res 30: 3059–3066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215: 403–410. 10.1016/S0022-2836(05)80360-2; S0022-2836(05)80360-2 [pii]. [DOI] [PubMed] [Google Scholar]
- 69.O'Donohue MJ, Moriarty DJW, MacRae IC (1991) Nitrogen fixation in sediments and the rhizosphere of the seagrass Zostera capricorni. Microbiol Ecol 22: 53–64. [DOI] [PubMed] [Google Scholar]
- 70.Boddey RM (1995) Biological nitrogen fixation in sugarcane; A key to energetically viable biofuel production. CRC Crit Rev Plant Sci 14: 263–279. [Google Scholar]
- 71.Hamisi M, Diez B, Lyimo T, Ininbergs K, Bergman B (2013) Epiphytic cyanobacteria of the seagrass Cymodocea rotundata: diversity, diel nifH expression and nitrogenase activity. Environ Microbiol Rep 5: 367–376. 10.1111/1758-2229.12031 [DOI] [PubMed] [Google Scholar]
- 72.Gentili F, Nilsson MC, Zackrisson O, DeLuca TH, Sellstedt A (2005) Physiological and molecular diversity of feather moss associative N2-fixing cyanobacteria. J Exp Bot 56: 3121–3127. eri309 [pii]; 10.1093/jxb/eri309 [DOI] [PubMed] [Google Scholar]
- 73.Fuqua C, Greenberg EP (2002) Listening in on bacteria: acyl-homoserine lactone signalling. Nat Rev Mol Cell Biol 3: 685–695. 10.1038/nrm907; nrm907 [pii]. [DOI] [PubMed] [Google Scholar]
- 74.Thaweenut N, Hachisuka Y, Ando S, Yanagisawa S, Yoneyama T (2011) Two seasons' study on nifH gene expression and nitrogen fixation by diazotrophic endophytes in sugarcane (Saccharum spp. hybrids): expression of nifH genes similar to those of rhizobia. Plant Soil 338: 435–449. [Google Scholar]
- 75.Burbano CS, Liu Y, Rosner KL, Reis VM, Caballero-Mellado J, Reinhold-Hurek B et al. (2011) Predominant nifH transcript phylotypes related to Rhizobium rosettiformans in field-grown sugarcane plants and in Norway spruce. Environ Microbiol Rep 3: 383–389. 10.1111/j.1758-2229.2010.00238.x [DOI] [PubMed] [Google Scholar]
- 76.Bodenhausen N, Bortfeld-Miller M, Ackermann M, Vorholt JA (2014) A synthetic community approach reveals plant genotypes affecting the phyllosphere microbiota. PLoS Genet 10: e1004283 10.1371/journal.pgen.1004283; PGENETICS-D-13-02228 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Shen SY, Fulthorpe R (2015) Seasonal variation of bacterial endophytes in urban trees. Front Microbiol 6: 427 10.3389/fmicb.2015.00427 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Berendsen RL, Pieterse CM, Bakker PA (2012) The rhizosphere microbiome and plant health. Trends Plant Sci 17: 478–486. S1360-1385(12)00079-9 [pii]; 10.1016/j.tplants.2012.04.001 [DOI] [PubMed] [Google Scholar]
- 79.Burke C, Steinberg P, Rusch D, Kjelleberg S, Thomas T (2011) Bacterial community assembly based on functional genes rather than species. Proc Natl Acad Sci U S A 108: 14288–14293. 1101591108 [pii]; 10.1073/pnas.1101591108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Dabundo R, Lehmann MF, Treibergs L, Tobias CR, Altabet MA, Moisander PH et al. (2014) The Contamination of Commercial 15N2 Gas Stocks with 15N-Labeled Nitrate and Ammonium and Consequences for Nitrogen Fixation Measurements. PLoS One 9: e110335 10.1371/journal.pone.0110335; PONE-D-14-11473 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Giller KE, Merckx R (2003) Exploring the boundaries of N2 fixation in cereals and grasses: An hypothetical and experimental framework. Symbiosis 35: 3–16. [Google Scholar]
- 82.Bergman B, Sandh G, Lin S, Larsson J, Carpenter EJ (2013) Trichodesmium—a widespread marine cyanobacterium with unusual nitrogen fixation properties. FEMS Microbiol Rev 37: 286–302. 10.1111/j.1574-6976.2012.00352.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Berman-Frank I, Lundgren P, Chen YB, Kupper H, Kolber Z, Bergman B et al. (2001) Segregation of nitrogen fixation and oxygenic photosynthesis in the marine cyanobacterium Trichodesmium. Science 294: 1534–1537. 10.1126/science.1064082; 294/5546/1534 [pii]. [DOI] [PubMed] [Google Scholar]
- 84.Gallon JR (1992) Reconciling the Incompatible—N-2 Fixation and O-2. New Phytologist 122: 571–609. [Google Scholar]
- 85.Zhulin IB, Bespalov VA, Johnson MS, Taylor BL (1996) Oxygen taxis and proton motive force in Azospirillum brasilense. J Bacteriol 178: 5199–5204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Krause A, Ramakumar A, Bartels D, Battistoni F, Bekel T, Boch J et al. (2006) Complete genome of the mutualistic, N2-fixing grass endophyte Azoarcus sp. strain BH72. Nat Biotechnol 24: 1385–1391. nbt1243 [pii]; 10.1038/nbt1243 [DOI] [PubMed] [Google Scholar]
- 87.Balsanelli E, de Baura VA, Pedrosa FO, De Souza EM, Monteiro RA (2014) Exopolysaccharide biosynthesis enables mature biofilm formation on abiotic surfaces by Herbaspirillum seropedicae. PLoS One 9: e110392 10.1371/journal.pone.0110392; PONE-D-14-20679 [pii]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Mitter B, Petric A, Shin MW, Chain PS, Hauberg-Lotte L, Reinhold-Hurek B et al. (2013) Comparative genome analysis of Burkholderia phytofirmans PsJN reveals a wide spectrum of endophytic lifestyles based on interaction strategies with host plants. Front Plant Sci 4: 120 10.3389/fpls.2013.00120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Bertalan M, Albano R, de P V, Rouws L, Rojas C, Hemerly A et al. (2009) Complete genome sequence of the sugarcane nitrogen-fixing endophyte Gluconacetobacter diazotrophicus Pal5. BMC Genomics 10: 450 1471-2164-10-450 [pii]; 10.1186/1471-2164-10-450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Khan Z, Kandel S, Ramos D, Ettl GJ, Kim S-H, Doty SL (2015) Increased biomass of nursery-grown Douglas-fir seedlings upon inoculation with diazotrophic endophytic consortia. Forests 6: 3582–3593. [Google Scholar]
- 91.Scherling C, Ulrich K, Ewald D, Weckwerth W (2009) A metabolic signature of the beneficial interaction of the endophyte paenibacillus sp. isolate and in vitro-grown poplar plants revealed by metabolomics. Mol Plant Microbe Interact 22: 1032–1037. 10.1094/MPMI-22-8-1032 [DOI] [PubMed] [Google Scholar]
- 92.Schink B, Ward JC, Zeikus JG (1981) Microbiology of wetwood: role of anaerobic bacterial populations in living trees. J Gen Microbiol 123: 313–322. [Google Scholar]
- 93.van der Kamp BJ (1986) Nitrogen fixation in cottonwood wetwood. Can J For Res 16: 1118–1120. [Google Scholar]
- 94.Tuskan GA, Difazio S, Jansson S, Bohlmann J, Grigoriev I, Hellsten U et al. (2006) The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 313: 1596–1604. [DOI] [PubMed] [Google Scholar]
- 95.Cocking EC (2005) Intracellular colonization of cereals and other crop plants by nitrogen-fixing bacteria for reduced inputs of synthetic nitrogen fertilizers. In vitro Cell Dev Biol-Plant 41: 369–373. [Google Scholar]
- 96.Olivares J, Bedmar EJ, Sanjuan J (2013) Biological nitrogen fixation in the context of global change. Mol Plant Microbe Interact 26: 486–494. 10.1094/MPMI-12-12-0293-CR [DOI] [PubMed] [Google Scholar]
- 97.Carvalho TL, Balsemao-Pires E, Saraiva RM, Ferreira PC, Hemerly AS (2014) Nitrogen signalling in plant interactions with associative and endophytic diazotrophic bacteria. J Exp Bot 65: 5631–5642. eru319 [pii]; 10.1093/jxb/eru319 [DOI] [PubMed] [Google Scholar]
- 98.James EK, Baldani JI (2012) The role of biological nitrogen fixation by non-legumes in the sustainable production of food and biofuels. Plant and Soil 356: 1–3. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Leaf and stem samples of surface-sterilized wild poplar genotypes 1–7 that had been collected in summer 2013 and grown in NFM were tested by PCR for the presence of putative diazotrophic bacteria. PCR was performed using Populus ralf 6 primers as a positive control (upper panel). The nifH b1 primers [63] were used to detect the presence of nitrogenase (lower panel). Genomic DNA extracted from Azotobacter vinelandii was used as a positive control for nifH. The arrow indicates the nifH gene product. S, stem; L, leaf. Lanes 1, 1Kb Plus DNA ladder; Lanes 2, no-DNA controls; Lane 3, Pop1S; Lanes 4, Pop1L; Lanes 5, Pop2S; Lanes 6, Pop2L; Lanes 7, Pop3S; Lanes 8, Pop3S.2; Lanes 9, Pop3L; Lanes 10, Pop4S; Lanes 11, Pop4L; Lanes 12, Pop5S; Lanes 13, Pop5L; Lanes 14, Pop6S; Lanes 15, Pop6L; Lanes 16, Pop7S; Lanes 17, Pop7L; Lanes 18, empty; Lanes 19, Azotobacter vinelandii.
(TIF)
Polygenomic (plant and endophytic) DNA was isolated from two rooted cuttings of Wild Poplar 4. A subset of the branches were labeled of those with similarity 90% or above in GenBank. Unlabeled branches indicate sequences with closest matches to nifH genes of uncultured bacteria in Gene Bank. The three colored groups represent Group 1 (black), Group II (red) and Group III (blue) nifH sequences as per [66].
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.