

Abstract
Tumor necrosis factor (TNF) is a potent promoter of carcinogenesis and potentially important target for cancer prevention. TNF is produced as functionally distinct transmembrane and soluble molecules (tmTNF and sTNF, respectively), but their individual roles in carcinogenesis are unexplored. We investigated the participation of tmTNF and sTNF in chemically induced carcinogenesis in mice. We found that injection of XPro1595, a dominant-negative TNF biologic (DN-TNF) and specific antagonist of sTNF, decreased tumor incidence and growth, and prolonged survival of 3-methylcholanthrene (MCA)-injected mice. Similar results were obtained following the exclusion of both TNF forms by either TNF-receptor 2–Fc fusion protein (TNFR2-Fc) treatment or TNF-gene deletion. In addition, gene-deletion of TNFR1, which is preferentially triggered by sTNF, temporarily blocked, whereas gene-deletion of TNFR2, which is preferentially triggered by tmTNF, enhanced MCA-induced carcinogenesis. Concomitantly with carcinogenesis induction, MCA increased circulating IL1α, accumulation of myeloid-derived suppressor-cells (MDSCs), STAT3 phosphorylation, and immunosuppression in the spleen. In sharp contrast, DN-TNF treatment dramatically decreased IL1α and increased the essential immunoregulatory cytokines IL1β, IL12p70, and IL17 in the peripheral blood of MCA-injected mice. In addition, MDSC accumulation, STAT3 phosphorylation, and immunosuppression in MCA-injected mice were prevented by DN-TNF treatment, TNFR2-Fc treatment, and/or gene deletion of TNF or TNFR1, but not deletion of TNFR2. These findings reveal that sTNF is both an essential promoter of carcinogenesis and a pivotal regulator of MDSCs; and indicate that sTNF could be a significant target for cancer prevention and therapy.
Keywords: 3-methylcholanthrene, carcinogenesis, soluble TNF, transmembrane TNF, myeloid-derived suppressor cells, immunosuppression
Introduction
Causal links of chronic inflammation and carcinogenesis have been increasingly documented (1,2). The first indications were provided by epidemiological findings that virtually all cancers arise in tissues undergoing chronic inflammation (2). The increased secretion of pro-inflammatory cytokines associated with gene polymorphisms was also found to be linked to increased risks to various inflammatory diseases and cancers (3,4).
More pertinent evidence has been obtained in mouse models that have been modified to have either activated or suppressed nuclear factor-κB (NF-κB) in specific tissues (5). Specifically, Mdr2-knockout mice spontaneously develop cholestatic hepatitis, a form of chronic inflammation that triggers via TNF NF-κB signaling in hepatocytes and leads to hepatocellular carcinoma. In contrast, these mice fail to develop cancer if activation of NF-κB is suppressed by neutralization of TNF or when hepatocyte-specific inducible IκB-super-repressor transgene is introduced (6). Similarly, mice lacking NF-κB–activator IKKβ in intestinal epithelial or myeloid cells show decreased colon-tumor incidence or size, respectively, after being fed with azoxymethane (AOM)/dextran-sulfate-sodium (DSS), a colitis-associated cancer inducer (7).
An inflammation-carcinogenesis link has been demonstrated in mice having overexpressed or deleted genes encoding pro-inflammatory cytokines or their receptors. Gastric-mucosa overexpression of IL1β leads to local inflammation, recruitment of MDSCs and cancer development (8). In contrast, mice lacking TNF-gene are resistant to chemical carcinogenesis in the skin (9,10). Similarly, TNFR1-gene deficient mice show reduced AOM/DSS-caused colon inflammation and tumor incidence (11). Extending these findings and supporting the central role for TNF in inflammation, carcinogenesis and targeted cancer prevention, TNF neutralization by TNFR2-Fc (ENBREL) leads to reduction of carcinogen-induced inflammation and tumorigenesis (10).
The mechanism of TNF involvement in carcinogenesis has been partially elucidated. The combination of TNF, IFNγ, and IL1β activates inducible nitric-oxide synthase (iNOS) leading to excessive production of NO, which causes DNA damage and inhibits DNA-repair proteins (12). More importantly, TNF directly induces transformation of carcinogen-initiated fibroblasts (13), inhibits carcinogen-induced cell death, and promotes cell proliferation via NF-κB activation (14).
Tumor necrosis factor has both transmembrane (tmTNF) and soluble (sTNF) forms (15). Transmembrane TNF is expressed on cell membrane as functional homotrimers of 26-kDa type-II protomers. The soluble form is also a homotrimer, but consists of the 17-kDa extracellular domain of tmTNF that was cleaved by TNFα-converting enzyme. Discovered as an endotoxin-induced serum factor causing hemorrhagic necrosis of tumors, and applied with limited use in treatment of limb sarcoma, TNF has been subsequently defined as a pleiotropic cytokine that mediates a number of vital functions, including structural organization of secondary lymphoid organs, apoptosis, cell-proliferation/survival, inhibition of intracellular pathogens, immunoregulation, and inflammation (15). Additionally, TNF deregulation plays important roles in pathogenesis of granuloma formation, autoimmune diseases, acute-phase reaction, septic shock, fever, and cachexia. These diverse functions are induced via cognate interactions of the two forms of TNF with two transmembrane receptors, TNFR1 and TNFR2.
Although TNF is mainly made by activated macrophages, TNFR1 is ubiquitously expressed by almost all mammalian cells, and TNFR2 is only expressed by immune and endothelial cells (15). Different immune cells express different levels of the two receptors. Both TNF receptors bind both TNF forms, but sTNF and tmTNF preferentially trigger TNFR1 and TNFR2, respectively (15,16). The receptors for TNF have structurally distinct intracellular domains and transduce different signaling pathways. However, although only TNFR1 is associated with the signaling molecules that transduce apoptosis, both TNF receptors can bind molecules that transduce inflammation and signal cell survival. Due to the shared signaling molecules, both TNF receptors can cross-transduce signals to one another (15,16). Because of this complexity, the specificity of sTNF and tmTNF stimulation of their preferred receptors is variable.
The individual roles of tmTNF and sTNF in immunity and inflammation have started to be elucidated. Mouse and human dendritic cell (DC)/natural killer (NK)-cell crosstalk, a central immunoregulatory mechanism that induces skewing and increases of Th1-type cytokines and antitumor immunity, is a non-redundant function of tmTNF-TNFR2 interaction, not involving the sTNF-TNFR1 axis (17-19). In contrast, inflammation-driven autoimmune diseases can be prevented by selective sequestration of sTNF, or genetic-deletion or antibody-blockade of TNFR1, showing that inflammation is mediated preferentially by sTNF-TNFR1 interaction (20,21). However, the individual roles of sTNF and tmTNF in carcinogenesis are still unexplored.
In the present study, we demonstrate a pivotal role of sTNF in expansion of MDSCs, development of immunosuppression and promotion of carcinogenesis and tumorigenesis in a murine model of chemically induced carcinogenesis. These findings suggest that sTNF could be an important target for cancer prevention.
Materials and Methods
Mice
Eight-week-old wild-type C57BL/6 (B6), T-cell/B-cell–deficient SCID (B6.129S7-Rag1tm1mom/J), TNF-deficient (TNFko, B6.129S-Tnftm1Gkl/J), TNFR1-deficient (TNFR1ko, B6.129S2-Tnfrsf1atm1Mak/J) and TNFR2-deficient (TNFR2ko, B6.129S2-Tnfrsf1btm1Mwm/J) female mice were purchased from Jackson Laboratory (Bar Harbor, ME). All mice were homozygous, and bread by standard brother-sister mating at Jackson Laboratory. Mice were housed at the University of Pittsburgh Cancer Institute's Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC) internationally accredited animal facility. The animal studies were approved by the University of Pittsburgh Institutional Animal Care and Use Committee.
Reagents
The following reagents were used in the present study: human-recombinant interleukin-2 (IL2) (Chiron Corp., Emeryville, CA); TLR4 ligand, Escherichia-coli lipopolysaccharide (LPS) (Lonza, Walkersville, MD); human TNFR2-Fc fusion protein (Amgen, Thousand Oaks, CA); clinical-grade pegylated DN-TNF XPro1595 biologic (20, Xencor, Monrovia, CA); fluorescein isothiocyanate (FITC)-conjugated anti-CD3, anti-CD4, anti-CD11b and anti-CD45R monoclonal antibodies (mAbs); phycoerythrin (PE)-conjugated anti-CD3, anti-Gr1, anti-CD25, anti-NK1.1, anti-CD11c and anti-F4/80 mAbs; allophycocyanin (APC)-conjugated anti-Ly6C, anti-FoxP3 and anti-CD8 mAbs (all anti-mouse CDs); and corresponding fluorochrome-conjugated isotype-control mAbs (eBioscience, San Diego, CA); mouse Quantikine IFN-γ enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN); and Millipore mouse 32-plex cytokine kit (Billerica, MA).
Induction, treatment, and measurement of carcinogenesis in vivo
Wild-type and TNF-, TNFR1- and TNFR2-deficient B6 mice were injected subcutaneously (s.c.) with 0.1 mg MCA dissolved in 0.1 mL sesame oil (Sigma, St. Louis, MO). Wild-type mice were then randomly divided into groups of 10-15 mice. PBS and its XPro1595-DN-TNF and ENBREL-TNFR2-Fc solutions (200 μg/0.5 mL/mouse) were injected intraperitoneally (i.p.) twice a week, for 12 weeks, beginning on the day of MCA injection. Appearance of 2-mm diameter tumors was detected by palpation twice weekly. Tumor growth was determined by measuring two perpendicular tumor-diameters with calipers twice weekly. Data are presented as multiplications of two tumor diameters of individual tumors and their mean values. Survival was scored daily.
In vitro suppression of DC/NK-cell crosstalk
Endogenous DC/NK-cell crosstalk was assessed as follows. Splenocytes (2 × 106/mL) of untreated and treated wild-type mice were resuspended in complete cell-culture medium (CM: RPMI-1640 medium, 0.1 mM nonessential amino acids, 2 mM sodium pyruvate, 1 mM L-glutamine, 100 μg/ml streptomycin, 100 U/mL penicillin, 10% fetal calf serum [FCS], Life Technologies, Grand Island, NY; 50 μM 2-mercaptoethanol, Bio-Rad, Hercules, CA) supplemented with 1 μg/mL LPS and 6,000 IU/mL IL2. The cell suspensions were seeded in 96-well round-bottom plates (BD-Biosciences, San Jose, CA) and incubated at 37°C, for 24 h.
Cell-mediated suppression of DC/NK-cell crosstalk was assessed as follows. SCID-mouse bulk splenocytes (composed of 50% NK1.1+CD3− NK cells, 30% CD11c+ iDCs and 20% F4/80+monocytes/macrophages) served as NK cell/DC-crosstalk responders (22). Bulk splenocytes of untreated or MCA-injected (MCA/PBS-, MCA/DN-TNF-, or MCA/ENBREL-treated) wild-type mice served as stimulators/suppressors of SCID-mouse DC/NK-cell responders. The responders and stimulators/suppressors were suspended in CM, supplemented with LPS (1 μg/mL) and IL2 (6,000 IU/mL), seeded either alone or mixed in 1:1 and 1:3 cell-to-cell ratios in 96-well round-bottom plates, and incubated at 37°C, for 24 h.
The assays were performed in triplicates. Following the incubations, cell-free supernatants were examined for the presence of IFN-γ using ELISA.
Flow cytometry
Standard cell-surface flow cytometry with fluorochrome-conjugated antibodies was performed as described (17). B cells (CD45R+F4/80−), macrophages (CD45R−F4/80+), NK cells (CD3−NK.1.1+) and DCs (CD11b+CD11c+), CD4 (CD3+CD4+CD8−) and CD8 (CD3+CD4−CD8+) T cells, monocytic (CD11b+Gr1lo/−Ly6C+/hi) and granulocytic (CD11b+Gr1hiLy6C+) MDSCs were examined using direct multi-color cell surface staining. Treg (CD4+Foxp3+CD25hi) were examined per manufacturer protocol (Biolegend, San Diego, CA). Phosphorylated STAT3 was investigated using PE-conjugated anti-phosphorylated STAT3-tyrosine-705 (pSTAT3y705), Alexa488-conjugated anti-pSTAT3-serine-727 (pSTAT3s727) and corresponding isotype-control mouse-IgG2a mAbs (BD Biosciences), as described (23). Cells were analyzed on Beckman-Coulter CyAN ADP cytometer (Brea, CA). Data-analyses were performed using Summit 4.3 software (Beckman-Coulter).
Quantification of cytokines
Serum cytokines were quantified using Millipore-multiplex mouse-cytokine kit, as recommended by the company. IFN-γ was measured in the cell-culture-conditioned media using mouse-IFN-γ Quantikine ELISA kit (R&D).
Statistics
Data were statistically evaluated using 10.0-SPSS (SPSS Inc., Chicago, IL) and 3.0.2.-R (www.R-project.org) softwares. Data are reported as means ± SD. Statistical significance was assessed using Student's t test. Additionally, cancer-appearance (tumor-development cumulative hazard) and survival-time was analyzed using Kaplan-Meier method. Exact-log-rank tests were used to compare cancer-appearance and survival-time curves of experimental and control groups. P ≤ 0.05 was considered significant.
Results
Soluble TNF sequestration prevents MCA-induced carcinogenesis
DN-TNF selectively sequesters sTNF without affecting tmTNF, and inhibits inflammations without impacting major immune mechanisms controlling intracellular pathogens and cancer (20,21). Differently, TNFR2-Fc neutralizes both sTNF and tmTNF, and inhibits both inflammations and major immune mechanisms (17-20). We initially examined the impact of XPro1595-DN-TNF and ENBREL-TNFR2-Fc on MCA-induced carcinogenesis in wild-type B6 mice (Fig. 1A). We found that 4-mm2 tumors appear in all mice regardless of treatment on week 4 after MCA injection. The incipient tumors were more frequent in PBS-treated than in ENBREL-treated and, especially, DN-TNF-treated mice. The differences in tumor frequencies between PBS-treated and ENBREL-treated, PBS-treated and DN-TNF-treated, or ENBREL-treated and DN-TNF-treated mice were significant (P = 0.009, P < 0.00005, and P = 0.027, respectively).
Figure 1. Sequestration of sTNF or deletion of TNF or TNFR1 genes protect mice from MCA-induced carcinogenesis.
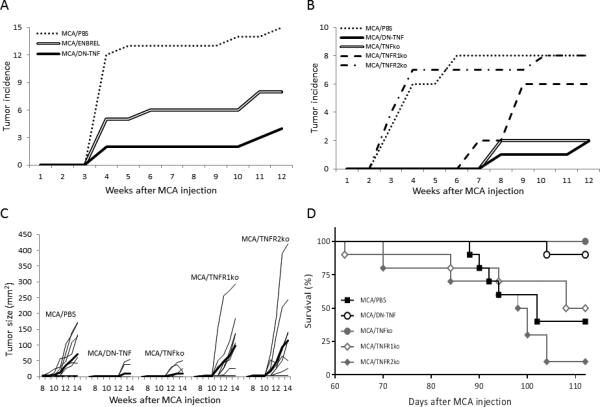
Panel A: Wild-type B6 mice were injected s.c. with MCA as described in Materials and Methods. The MCA-injected mice were randomized into 3 groups of 15 mice. The control-group mice were injected i.p. with PBS (0.5 mL). The two other groups’ mice were injected i.p. with either ENBREL or XPro1595 (DN-TNF) (200 μg/0.5 mL PBS/mouse), respectively. The injections of PBS, ENBREL and DN-TNF were performed twice a week, for 12 weeks, starting on the day of MCA injection. Panels B, C and D: Twenty wild-type, 10 TNF-deficient (TNFko), 10 TNFR1-deficient (TNFR1ko) and 10 TNFR2-deficient (TNFR2ko) B6 mice were injected with MCA, as described above. Ten of the MCA-injected wild-type mice were also injected i.p. with PBS, while the other 10 mice were injected with XPro1595 biologic. The MCA-injected gene-deficient mice were left untreated. MCA-injections and treatments were performed as described in Panel A. Panels A and B present the cumulative tumor incidence. Panel C presents the individual tumor sizes and their means (thick lines). Panel D presents the cumulative survival. The analyses were performed as described in Materials and Methods. In Panel A, the differences of MCA/PBS vs MCA/ENBREL or MCA/DN-TNF, and MCA/ENBREL vs MCA/DN-TNF are significant (P = 0.009, P < 0.00005, and P = 0.027 respectively). In Panel B, the differences of MCA/PBS vs MCA/DN-TNF, MCA/TNFko and MCA/TNFR1ko; and MCA/TNFR2ko vs MCA/DN-TNF, MCA/TNFko and MCA/TNFR1ko are significant (P = 0.0007, P = 0.0016 and P = 0.031; P = 0.0013, P = 0.0026, and P = 0.029; respectively). In Panel C, the differences of MCA/PBS vs MCA/DN-TNF and MCA/TNFko; MCA/TNFR1ko vs MCA/DN-TNF and MCA/TNFko; and MCA/TNFR2ko vs MCA/DN-TNF and MCA/TNFko are significant (P = 0.017, P = 0.027; P = 0.023, P = 0.031; P = 0.029 and P = 0.036, respectively). In Panel D, the differences of MCA/DN-TNF vs MCA/TNFR1ko, MCA/PBS and MCA/TNFR2ko; MCA/TNFko vs MCA/TNFR1ko, MCA/PBS and MCA/TNFR2ko; and MCA/TNFR2ko vs MCA/PBS and MCA/TNFR1ko are significant (P < 0.0001, P < 0.0003, P < 0.0001; P < 0.0001, P < 0.0001, P < 0.0001; P = 0.037, and P = 0.029, respectively).
To confirm the initial findings and evaluate in more details the roles of sTNF and tmTNF in carcinogenesis, we compared the effects of PBS- and DN-TNF-treatment (sTNF-sequestration) in wild-type mice with the effects of TNF, TNFR1, and TNFR2 deletion (exclusions: sTNF and tmTNF, sTNF-receptor and tmTNF-receptor, respectively) in the gene-deficient mice on MCA-induced carcinogenesis. Tumor incidence (Fig. 1B), tumor size (Fig. 1C) and survival (Fig. 1D) were monitored. The time and incidence of tumor occurrence varied between the experimental groups (Fig. 1B). In PBS-treated wild-type and TNFR2-deficient mice, 4-mm2 tumors appeared early and in the majority of animals (week 4: 6/10 and 7/10 mice, respectively). In these mouse-groups, tumor incidence reached maximum (8/10 mice) on weeks 6 and 10, respectively. In sharp contrast, DN-TNF-sequestration of sTNF, exclusion of sTNF activity by knocking-out TNFR1, or exclusion of both sTNF and tmTNF by knocking-out TNF led to a delayed (3 to 4 weeks) and infrequent (1/10, 2/10, and 2/10 mice, respectively) occurrence of tumors. While the low tumor incidence remained unchanged in TNF-deficient and DN-TNF-treated mice, it notably increased in TNFR1-deficient mice (from 2/10 to 6/10 mice) on week 9 after MCA injection. Tumor-frequency differences of PBS-treated vs DN-TNF-treated, TNF-deficient and TNFR1-deficient mice were significant (P = 0.0007, P = 0.0016, and P = 0.031, respectively). Similarly, the differences in tumor frequencies of TNFR2-deficient vs DN-TNF-treated, TNF-deficient and TNFR1-deficient mice were also significant (P = 0.0013, P = 0.0026 and P = 0.029, respectively).
Tumors grew slowly or remained dormant in all groups of mice until week 10 after MCA-injection. Afterwards, in the test groups with frequent tumors (PBS-treated wild-type and TNFR1- and TNFR2-deficient mice), the tumors grew with a variable velocity and differed widely in size within each group, at all-time points of their measurements. In sharp contrast, the infrequent tumors in DN-TNF-treated and TNF-deficient mice remained small (4-9 mm2) and dormant until week 12 after MCA injection. Thus, between weeks 10 and 14 after MCA injection, tumors were significantly larger in PBS-treated, TNFR1-deficient and TNFR2-deficient mice than in DN-TNF-treated and TNF-deficient mice (vs DN-TNF: P = 0.017, P = 0.023 and P = 0.029; vs TNF-knockout: P = 0.027, P = 0.031 and P = 0.036, respectively) (Fig. 1C).
The lower tumor frequency and slower tumor growth in DN-TNF-treated and TNF-deficient mice than in PBS-treated, TNFR1-deficient and TNFR2-deficient mice resulted in significant prolongation of mouse-survival. At experiment end-point, day-112 (week-16) after MCA injection, 9/10 DN-TNF-treated and 10/10 TNF-deficient mice were alive. In sharp contrast, 5/10 TNFR1-deficient (vs DN-TNF, TNFko: P = 0.001, P = 0.003, respectively), 4/10 PBS-treated (vs DN-TNF and TNFko: P = 0.0026, P = 0.0037, respectively) and 1/10 TNFR2-deficient (vs DN-TNF and TNFko: P = 0.00033, P = 0.00012, respectively) mice were alive (Fig. 1D).
These findings demonstrate that sTNF is critical, while tmTNF is dispensable, for MCA-induced carcinogenesis. They also suggest that tmTNF, in contrast to sTNF, could have a protective role in carcinogenesis.
Sequestration of sTNF modulates immunoregulatory cytokines
MCA-induced cancers are immunogenic (24). Therefore, sTNF exclusion-induced increased resistance to MCA-induced carcinogenesis could be caused by a modified and/or enhanced anti-tumor immune response. Type and quantity of secreted cytokines define type and effectiveness of anti-tumor immune responses. We next examined cytokines in the sera of healthy/untreated (Control) and MCA-injected/PBS-, ENBREL- or DN-TNF-treated mice, two weeks after MCA injection (Fig. 2). In MCA-injected/PBS-treated mice, the central proinflammatory cytokines IL1β, IL12p40/p70 and IL17 were either unchanged (IL1β, IL17; Figs. 2A,2C) or slightly decreased (IL12p40/p70, Fig. 2B) relative to control mice. In contrast, the levels of these cytokines increased significantly in MCA-injected/DN-TNF-treated mice (IL1β: P = 0.026; IL12p40/70: P = 0.05; IL17; P = 0.039). The cytokines did not change in MCA-injected/ENBREL-treated mice. In contrast, IL1α levels were high in control mice, notably increased in MCA-injected/PBS-treated mice, but strongly decreased below control levels by ENBREL (P = 0.012) or DN-TNF (P = 0.023) treatment. These findings indicate that sTNF suppresses and sTNF-sequestration enables inflammasome (IL1β), Th1 (IL12) and Th17 (IL17) responses in MCA-injected mice. Because these responses didn't differ in control and MCA-injected/ENBREL-treated mice (neutralized sTNF and tmTNF), the increased responses after sTNF-sequestration could be mediated by tmTNF. In addition, the strong suppression of IL1α by ENBREL or DN-TNF indicates that sTNF up-regulates IL1α. Intriguingly, at this early stage of carcinogenesis, sTNF was not detectable in the sera of MCA-injected mice. However, sTNF could be initially present in biologically significant quantities in tumor tissues (1).
Figure 2. Sequestration of sTNF by DN-TNF and neutralization of both sTNF and tmTNF by TNFR2-Fc affect secretion of immunoregulatory cytokines in MCA-injected mice.
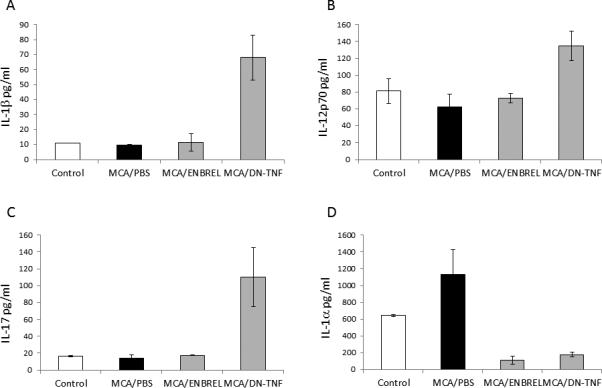
Healthy/untreated (Control), MCA-injected/PBS-treated (MCA/PBS), MCA-injected/ENBREL-treated (MCA/ENBREL) and MCA-injected/XPro1595-treated (MCA/DN-TNF) wild-type mice (3 mice per each group) were sacrificed and their sera obtained 14 days after MCA-injection. Sera were examined for the presence of IL1β (A), IL12p40/70 (B), IL17 (C), and IL1α (D) using the LUMINEX assay. The data represent mean values of triplicates ± SE. In (A), (B), and (C) the increases of cytokine levels in the sera of MCA/DN-TNF-treated mice were significant relative to that of Control and MCA/PBS-treated mice (P = 0.026, P = 0.05 and P = 0.039, respectively). In (D), the decreases of IL1α levels in the sera of MCA/ENBREL- and DN-TNF-treated mice vs Control and MCA/PBS-treated mice are significant (P = 0.012 and P = 0.023, respectively).
Sequestration of sTNF averts MDSC accumulation
Both carcinogens and cancer induce strong immunosuppression, which enable cancer development and growth (25-27). MDSCs are a heterogeneous population of immature myeloid cells that strongly suppress immune functions (28). While MDSCs are rare in healthy organisms, their frequency and activity highly increase in cancer-host bone marrow, peripheral lymphoid tissue and tumor. It is believed that MDSCs play significant roles in tumor-immune-escape and failure of immunotherapy. However, MDSC presence and function in chemically-induced carcinogenesis are underexplored (28). We examined changes in frequency of MDSCs (Suppl. Fig. 1, Figs. 3A-3D) and macrophages (Suppl. Fig. 2) following MCA injection and/or exclusion of sTNF or tmTNF. MCA-induced increases of spleen monocytic (CD11b+Gr1lo/−Ly6C+/hi, Figs. 3A, 3B) and granulocytic (CD11b+Gr1hiLy6C+, Figs. 3C, 3D) MDSC frequency in wild-type mice. Increased frequencies of MDSCs were notable at the cancer-initiation (day 14: 1.4- and 1.3-fold, respectively), and highly augmented in the tumor-established stage (day 84: 6.7- and 6.8-fold, respectively) of carcinogenesis. In contrast, MCA-induced carcinogenesis didn't change splenic F4/80+ macrophage frequency (Suppl. Figs. 2A-B). However, ENBREL-neutralization of both TNF forms decreased frequencies of monocytic and granulocytic MDSCs, and macrophages in both tumor-free and tumor-bearing MCA-injected mice. Sequestration of sTNF by XPro1595-DN-TNF invariably and dramatically decreased frequencies of monocytic MDSCs, granulocytic MDSCs and macrophages to or below their control levels, in MCA-injected both tumor-free and tumor-bearing mice. Similar decreases of MDSC and macrophage frequencies were observed in MCA-injected tumor-bearing TNF- and TNFR1-deficient mice. In contrast to MCA-injected/DN-TNF- or ENBREL-treated wild-type and TNF- or TNFR1-deficient mice, and similar to MCA-injected/PBS-treated wild-type mice, MCA-injected/TNFR2-deficient mice lacking preferential tmTNF-receptor possessed highly increased frequencies of both monocytic and granulocytic MDSCs, and unchanged frequencies of macrophages, relative to control mice. FoxP3+CD4+CD25+ Treg frequencies were found unchanged in all MCA-injected groups of mice (Suppl. Fig. 3). These data show that MCA induces increased MDSC accumulation during carcinogenesis. They also provide evidence that sTNF, but not tmTNF, is required for the MCA-induced accumulation of MDSCs and maintenance of the macrophage population, and that treatment with the selective sTNF inhibitor XPro1595-DN-TNF efficiently prevents MDSC accumulation.
Figure 3. sTNF exclusion averts MCA-induced MDSC accumulation.
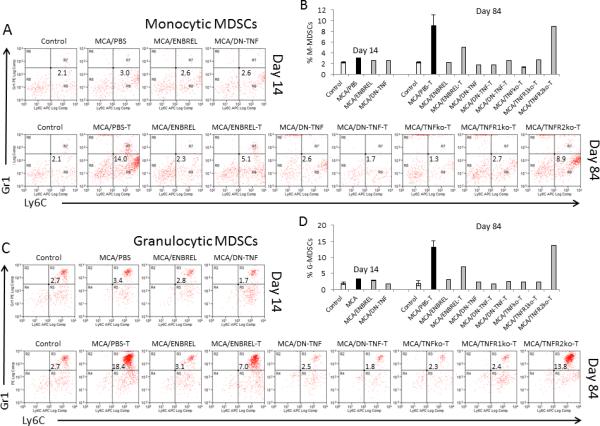
Similar experimental groups were set up as described in Fig. 1 legend. Both tumor-free and tumor-bearing (T) mice were examined: healthy/untreated control (Control); MCA-injected/PBS-treated (tumor-free:tumor-bearing: MCA/PBST), ENBREL-treated (tumor-free: MCA/ENBREL; tumor-bearing : MCA/ENBREL-T), XPro1595-treated (tumor-free: MCA/DN-TNF; tumor-bearing: MCA/DN-TNF-T); and/or tumor-bearing MCA-injected/TNFko (MCA/TNFko-T), TNFR1ko (MCA/TNFR1ko-T) and TNFR2ko-T (MCA/TNFR2ko-T) mice. Spleens were obtained from the listed groups of mice on days 14 (A, B) and/or 84 (C, D) following MCA injection, and stained with fluorochrome-conjugated antibodies to CD11b, Gr1, and Ly6C. Stained splenocytes were analyzed by three-color flow cytometry. The gating/analysis strategy is presented in Suppl. Fig. 1. CD11b+Gr1lo/−Ly6C+/hi monocytic MDSCs (A, B), and CD11b+Gr1hiLy6C+ granulocytic MDSCs (C, D) were scored. Dot-plot data present non-granular Gr1−Ly6C+CD11b+ (A), and granular Gr1+Ly6C+CD11b+ (C) cells. Histograms show individual and mean (2-3 replicates ± SD: Control, MCA/PBS-T and DN-TNF-T) percentages of monocytic (B) and granulocytic (D) MDSCs,.
Soluble TNF-dependent STAT3 activation in myeloid cells
The transcription factor STAT3 plays an essential role in regulation of MDSC development and function (28,29). Its active, phosphorylated form, pSTAT3, is considered a hallmark of MDSCs. To validate the carcinogen-induced sTNF-dependent expansion of MDSCs, we investigated whether MCA induces sTNF-dependent pSTAT3 in myeloid cells. Splenocytes of healthy/untreated-control and MCA-injected/PBS-, ENBREL- or DN-TNF-treated wild-type mice were examined by flow cytometry for the presence of pSTAT3y705 and pSTAT3s727 residues day-84 after MCA injection (Fig. 4). Monocytes and granulocytes were defined by forward-scatter (size) and side-scatter (granularity), as non-granular/large- and granular/medium-size cells, respectively (Suppl. Fig. 4). The identities of cell populations were confirmed using their labeling with fluorochrome-conjugated antibodies to Gr1, CD11b, Ly6C and F4/80, and back-gating (Suppl. Figs. 1, 2, not-shown data). In controls, the frequencies of splenic monocytes and granulocytes expressing pSTAT3 were low (Fig. 4). In contrast, frequencies of STAT3y705+ (Figs, 4A, 4B) and pSTAT3s727+ (Figs, 4C, 4D) monocytes (Figs, 4A, 4C) and granulocytes (Figs, 4B, 4D) were highly increased in MCA-injected/PBS-treated mice. The MCA-induced increases in pSTAT3+ cell-frequencies were prevented in both tumor-free and tumor-bearing mice by ENBREL or DN-TNF treatment. These data show that MCA and/or MCA-induced tumors activate the essential MDSC transcription factor STAT3 in myeloid cells, which parallels and correlates with MDSC accumulation. They also show that sTNF is not only important for MCA-induced accumulation of MDSCs but also for MCA-induced activation of STAT3 in myeloid cells.
Figure 4. sTNF-sequestration prevents MCA-induced activation of STAT3 in myeloid cells.
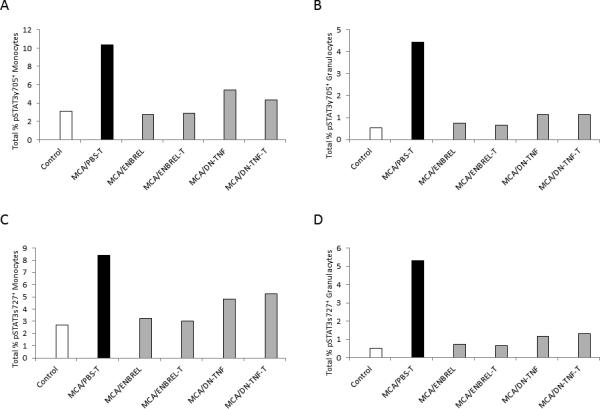
Splenocytes of healthy/untreated control (Control) and MCA-injected/PBS-treated (tumor-bearing: MCA-T), ENBREL-treated (tumor-free: MCA/ENBREL; and tumor-bearing: MCA/ENBREL-T), and XPro1595-treated (tumor-free: MCA/DN-TNF; and tumor-bearing MCA/DN-TNF-T) wild-type mice were obtained on day 84 after MCA injection, and examined by flow cytometry for intracellular presence of the phosphorylated STAT3y705 (A, B) and STAT3s727 (C, D). The analyses of gated large non-granular mononuclear cells (Monocytes) (A, C) and middle-size granular (Granulocytes) (B, D) were performed as shown in Suppl. Fig. 3. Data represent total percentages of stained monocytes and granulocytes.
MCA induces and sTNF inhibition averts cell-mediated suppression of NK cell/DC crosstalk
NK-cell/DC crosstalk is a central immunoregulatory mechanism that defines type and extent of innate and adaptive immune responses (30-32). We have previously determined that the crosstalk leading to a high Th1 response, which efficiently controls cancer growth, is mainly mediated via cell-to-cell contact and tmTNF (17-19,33). Carcinogens and tumors suppress NK cells and/or DCs (26,27,34) that may disable NK-cell/DC crosstalk and allow carcinogenesis and tumor growth. Because MCA concomitantly induced sTNF-dependent carcinogenesis, suppression of central immunoregulatory cytokines that are up-regulated in NK-cell/DC crosstalk, and accumulation of MDSCs, we examined whether MDSC-mediated suppression of NK cell/DC crosstalk is an MCA-induced immunosuppression mechanism (Fig. 5). We initially assessed the endogenous NK-cell/DC crosstalk among splenocytes of healthy/untreated-control and MCA-injected/PBS-, ENBREL- and DN-TNF-treated wild-type mice (Figs. 5A, 5B). Splenocytes were stimulated with IL2 and LPS, to activate NK cells and DCs, respectively, and promote their crosstalk and IFNγ secretion (17,19). IL2/LPS-stimulation induced enhanced NK-cell/DC crosstalk and IFNγ secretion in healthy untreated mouse splenocytes (Control 2) relative to their unstimulated counterpart (Control 1). Both 14 and 84 days after carcinogen injection, the IL2/LPS-stimulated splenocytes of all MCA-injected mice regardless of treatment, except of DN-TNF-treated tumor-free mice 84 days after MCA injection, showed significant decreases of Th1 response (Figs. 5A, 5B). The decreases of activity were particularly pronounced in splenocytes of ENBREL-treated mice. These findings show that NK cell/DC crosstalk is suppressed throughout MCA-induced carcinogenesis. The enhanced suppression of NK-cell/DC-crosstalk in all ENBREL-treated mice, and lack of suppression in DN-TNF-treated tumor-free mice day 84 after MCA, confirm that endogenous NK-cell/DC crosstalk is primarily mediated by tmTNF (17-19).
Figure 5. sTNF-sequestration prevents MCA induction of cell-mediated suppression of NK-cell/DC crosstalk.
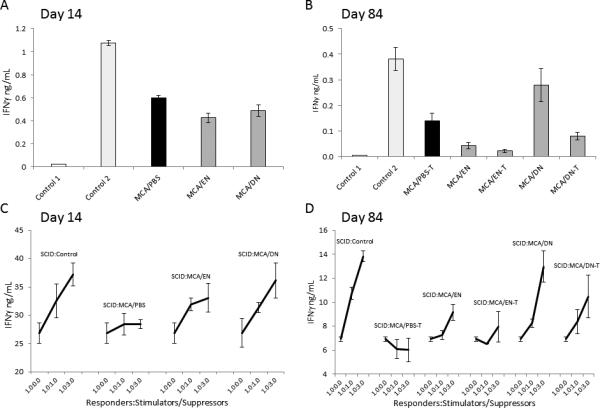
As described in Fig. 3 legend, splenocytes of healthy/untreated (Control), and MCA-injected/PBS-treated (tumor-free: MCA/PBS; and tumor-bearing: MCA/PBS-T), ENBREL-treated (tumor-free: MCA/EN; and tumor-bearing: MCA/EN-T) and XPro1595-treated (tumor-free: MCA/DN; and tumor-bearing: MCA/DN-T) wild-type mice were obtained on days 14 (A, C) and/or 84 (B, D) after MCA injection. These splenocytes, either alone (A, B) or mixed with SCID splenocytes (SCID) at 1:1 and 3:1 ratios (C, D), were stimulated with LPS (1 μg/mL) and IL2 (6,000 IU/mL), for 24 h. After this stimulation, the cell culture-conditioned media were collected and examined for the presence of IFNγ using ELISA. In A and B, Control 1 is unstimulated, and Control 2 is IL2/LPS-stimulated splenocytes of healthy/untreated mice. Data are means of triplicates ± SD of IFNγ ng/mL. In A, MCA/PBS, MCA/EN and MCA/DN are significantly lower than Control 2 (P = 0.00041, P = 0.00057 and P = 0.0016, respectively). In B, MCA/PBS and MCA/EN are significantly lower than Control 2 (P = 0.0047, P = 0.0016, respectively). In C, SCID:Control (1.0:0.0) and SCID:MCA/PBS (1.0:1.0-1.0:3.0) are significantly lower than SCID:Control (1.0:1.0-1.0:3.0) (P = 0.026, P = 0.001, respectively); and SCID:MCA/EN (1.0:1.0-1.0:3.0) and SCID:MCA/DN (1.0:1.0-1.0:3.0) are significantly higher than SCID:MCA/PBS (1.0:1.0-1.0:3.0) (P = 0.019, and P = 0.001, respectively). In D, SCID:Control (1.0:0.0) and SCID:MCA/PBS-T (1.0:1.0-1.0:3.0) are significantly lower than SCID:Control (1.0:1.0-1.0:3.0) (P = 0.0034, P = 0.0033, respectively); and SCID:MCA/EN (1.0:3.0), SCID:MCA/DN (1.0:1.0-1.0:3.0) and SCID:MCA/DN-T (1.0:1.0-1.0:3.0) are significantly higher than SCID:MCA/PBS-T (1.0:1.0-1.0:3.0) (P = 0.027, P = 0.028, P = 0.046, respectively).
Next, we directly assessed whether MCA-induced MDSCs could suppress NK cell/DC crosstalk. In these experiments, the responder cells (Responders) were bulk SCID-mouse splenocytes, composed of 50% NK cells, 30% DCs and 20% monocytes/macrophages. The stimulator/suppressor cells (Stimulators/Suppressors) were bulk splenocytes of healthy/untreated (Control), MCA-injected/PBS-treated (tumor-free: MCA/PBS; tumor-bearing: MCA/PBS-T), MCA-injected/ENBREL-treated (tumor-free: MCA/EN; tumor-bearing: MCA/EN-T) and MCA-injected/DN-TNF-treated (tumor-free: MCA/DN; tumor-bearing: MCA/DN-T) wild-type mice, obtained on days 14 and 84 after MCA injection. Responders and stimulators/suppressors, either alone or mixed in 1:1 and 1:3 ratios, were incubated in cell-to-cell contact in the presence of IL2/LPS, for 24 h. Very similar results were obtained with splenocytes of mice harvested on days 14 (Fig. 5C) and 84 (Fig. 5D) after MCA injection. At both time-points, SCID NK cells and DCs interacted strongly and secreted large amounts of IFNγ (SCID:Control, 1.0:0.0). The reaction was highly increased in a dose-dependent manner by adding to SCID-mouse splenocytes healthy/untreated wild-type-mouse splenocytes, which had low MDSC frequency (Figs. 3). Importantly, the observed increases in IFNγ secretion were 6 to 10 fold greater than the responses of the corresponding stimulators/suppressors alone (Control 2, Figs. 5A, 5B), indicating a possible synergistic cooperation of SCID and healthy/untreated wild-type mouse splenocytes. In contrast, MCA/PBS-mouse splenocytes containing a large population of MDSCs (Fig. 3) strikingly decreased the SCID-mouse NK cell/DC crosstalk to baseline, on day 14, or below baseline, on day 84. In sharp contrast, MCA/ENBREL-mouse splenocytes, and more prominently MCA/DN-TNF-mouse splenocytes stimulated SCID NK cell/DC crosstalk measured by IFNγ secretion. Both tumor-free and tumor-bearing MCA/DN-TNF-mouse splenocytes lacking MDSCs (Fig. 3) stimulated SCID-mouse NK-cell/DC crosstalk as strongly as wild-type control-mouse splenocytes. In contrast, MCA/ENBREL-treated mouse splenocytes having low to moderate frequency of MDSCs (Fig. 3) slightly (day 14) and moderately (day 84) inhibited SCID-NK cell/DC crosstalk relative to that of control- or MCA/DN-TNF-treated-mouse splenocytes. These findings indicate that MCA and MCA-induced tumor strongly stimulate not only the expansion but also the immunosuppressive activity of MDSCs, which in turn inhibits NK-cell/DC crosstalk. They also suggest that the expansion and immunosuppressive activity of MDSCs are both dependent on sTNF and can be efficiently prevented by XPro1595-DN-TNF treatment.
Discussion
Previous studies showed that mice lacking either TNF or TNFR1 genes are resistant to chemically induced carcinogenesis in the skin (9,10) or colon (11), respectively. Similarly, wild-type mice that undergo chemically induced carcinogenesis and treatment with ENBREL show reduced tumorigenesis (10). These studies, however, didn't consider possible distinct roles in carcinogenesis of functionally different two TNF forms. Here we showed that selective exclusion of sTNF by XPro1595-DN-TNF treatment or TNFR1 deletion, or elimination of both sTNF and tmTNF by ENBREL treatment or TNF deletion prevented carcinogenesis, decreased tumor growth and prolonged survival of MCA-injected mice. In contrast, selective exclusion of tmTNF by TNFR2 deletion enhanced MCA-induced carcinogenesis. We therefore have demonstrated that sTNF is essential in MCA-induced carcinogenesis. The data also suggest that tmTNF, opposite to sTNF, could have a protective role in carcinogenesis.
Notably, while DN-TNF treatment or TNF deletion, excluding sTNF or both TNF forms, almost completely prevented carcinogenesis, ENBREL treatment, neutralizing the two TNF forms and also LTα and possibly LTα2β1 (35), partially prevented carcinogenesis. These findings indicate that besides tmTNF, LTα, or LTα2β1 could be protective in MCA-induced carcinogenesis.
A perplexing finding in our study was that exclusion of sTNF by TNFR1 deletion only temporarily prevented carcinogenesis, and led to the late development of fast growing tumors in the majority of MCA-injected mice. This may indicate that in addition to sTNF playing a major role, some other, less potent, factor(s) could also promote carcinogenesis by inducing late generation of TNFR1-expressing cancer cells. These cancer cells could be susceptible to and controlled by TNF-mediated cytotoxicity in wild-type mice (27,36,37). In contrast, in TNFR1-deficient mice, the MCA-induced cancer cells lack TNFR1 and may be resistant to TNF-mediated killing, and vigorously grow in the absence of the immune control.
Our findings support the following scenario for the initiation of carcinogenesis. Carcinogens induce injury and cell necrosis in target tissues. Necrotic cells release damage associated molecular pattern (DAMPs) molecules, including heat-shock proteins, high-mobility group box 1 (HMGB1), DNA, RNA, S100 molecules, and purine metabolites (38). DAMPs induce activation of innate immunity effector cells such as macrophages, DCs, and NK cells, which consequently release proinflammatory cytokines such as sTNF. The proinflammatory cytokines induce pro-carcinogenic inflammation. In parallel, tissue protective/healing anti-inflammatory and immunosuppressive feedback mechanisms develop. In our study, IL1β, IL12, and IL17, which are immunoregulatory cytokines produced by innate-immunity activated effectors and mediate the effective Th1 and Th17 anticancer immune mechanisms, are unchanged during the initiation of MCA-induced carcinogenesis, but enhanced after sTNF exclusion by DN-TNF. In the context of carcinogenesis, these findings indicate that, during carcinogenesis, the activation of anticancer immune mechanisms might be stimulated, but down-regulated by sTNF-induced immunosuppression. In support of this possibility, potentially immunosuppressive soluble IL1α (39) is notably secreted in healthy mice, significantly increased during the initiation of MCA-induced carcinogenesis, and strikingly decreased below its baseline levels by sTNF sequestration with DN-TNF.
IL1α and IL1β bind to the same receptors (39). Both cytokines are produced as 31-kD precursors, which are processed by the proteases calpain and IL1β converting enzyme (ICE), respectively, generating 17-kD soluble forms that are released outside of cells. Soluble IL1β is immunologically active. In contrast, soluble IL1α is seemingly immunologically inactive in spite of its ability to bind to IL1 receptors. However, soluble IL1α becomes immunologically active after binding to a cell-membrane mannose-like receptor. The calpain-mediated cleavage of IL1α precursor and release of 17-kD IL1α also leads to the generation of the functional 14-kD IL1α N-terminal propiece. The IL1α propiece functions in cells as a transcription factor, activating oncogenes and inhibiting tumor-suppressor genes, and is considered as an “endokine”. Complementarily, secreted 17-kD IL1α can function as a growth factor for malignant cells. Moreover, immunologically inactive soluble IL1α, capable of binding IL1 receptors, could block and prevent IL1 receptor interaction with the immunologically active forms of IL1, and thus function as a potent negative regulator of immune responses.
As IL1α is overexpressed in cells exposed to chemical carcinogens (39), our findings that soluble IL1α level is elevated in MCA-injected and strikingly decreased in DN-TNF-treated mice indicate that sTNF can enhance generation of propiece and soluble IL1α. As a result, the two upregulated forms of IL1α could promote malignant transformation of target cells, mediate growth of newly generated malignant cells and inhibit IL1-induced immune responses.
Soluble TNF and tmTNF preferentially trigger TNFR1 and TNFR2 signaling pathways, respectively. However, both TNF receptors are associated with TRAF2 adapter molecule, share TRAF1, have similar inflammatory/survival signaling pathways, and can crosstalk and cross-transduce signals one to the other (15,16). Two complementary mechanisms of sTNF-dependent MDSC accumulation at the periphery have been defined (40,41). The first one is dominant in chronic-inflammation, and is mediated by sTNF-induced S100A8/A9-RAGE signaling in MDSC progenitors leading to MDSC generation and differentiation arrest (40). As MDSC progenitors might lack TNFR2 and express only TNFR1, sTNF mediates this mechanism solely via TNFR1. The second mechanism is dominant in mice rapidly developing transplanted tumors and likely acute inflammation (41). This mechanism leads to protection from apoptosis and prolongation of survival of MDSCs by inducing NF-κB activation, cFLIP upregulation and caspase-8 inhibition. Seemingly paradoxical, the latter sTNF-mediated mechanism requires TNFR2. In this mechanism, sTNF could selectively trigger TNFR1 cell-survival signal that could be either executed solely through TNFR1 signaling pathway, if the expression of TNFR1 is prevalent; or the TNFR1-originated survival signal could be cross-transduced, amplified and executed through TNFR2 signaling pathway, if the expression of TNFR2 is prevalent. In agreement with these findings and considerations, we showed that MCA-induced chronic inflammation-based carcinogenesis caused in both tumor-free and tumor-bearing mice STAT3 phosphorylation in myeloid cells and accumulation of MDSCs, both of which were prevented by the exclusion of sTNF or TNFR1, but not TNFR2. These findings demonstrate that sTNF mediates MDSC accumulation during carcinogenesis, and suggest that this mainly occurs by selective triggering TNFR1 on MDSC progenitors leading to MDSC generation and differentiation arrest.
During carcinogenesis initiation, low increases of MDSC frequency occur. In contrast, during tumorigenesis, high increases of MDSC frequency occur. This difference could be caused by the involvement of different MDSC regulatory cytokines at different stages of carcinogenesis. sTNF appears to be involved throughout the carcinogenic process. Secreted IL1α, which can also mediate growth and homing of myeloid cells (39), could be involved in the MDSC growth and regulation at least during carcinogenesis initiation. On the other hand, VEGF and GM-CSF, which are implicated in MDSC growth in tumor-bearing hosts, could be involved when tumors are established (28). Our data suggest that sTNF has a central regulatory role in MDSC accumulation during carcinogenesis, which could comprise their generation and differentiation arrest (40), survival extension (41), and growth stimulated by IL1α, VEGF and GM-CSF (28,39).
Similar to MDSC accumulation, two TNF-dependent mechanisms are implicated in regulation of MDSC immunosuppressive functions. Thus, while sTNF mediates the acquisition and maintenance of MDSC immunosuppressive functions (40), tmTNF highly expressed on noncleavable-TNF-transduced tumor cells, amplifies these functions (42). As activated immune cells of both innate and adaptive immunity, but not tumor cells, highly express tmTNF, the latter mechanism might be induced by activated immune cells interacting with MDSCs. In parallel with the sTNF dependent accumulation of MDSCs in the spleen of MCA-injected mice, and in agreement with the published results and considerations, we observed potent sTNF-dependent MDSC-mediated immunosuppression of the central immunoregulatory NK cell/DC crosstalk. The ultimate MDSC immunosuppressive function could be a resultant of sTNF-dependent baseline suppressive activity and its tmTNF-mediated amplification induced by activated NK cells and DCs (17).
Similar to MDSC expansion, immunosuppressive activity was completely eliminated by sTNF exclusion with DN-TNF treatment in both tumor-free and tumor-bearing mice. As the frequency of other potential immunosuppressive cells, including CD4+CD25+Foxp3+ Treg and F4/80+ macrophages, did not change during MCA-induced tumorigenesis, it is conceivable that MDSCs are the main mediators of the splenocyte immunosuppression that evolves during chemical carcinogenesis. MDSCs strongly suppress T-cell responses by producing a number of immunosuppressive molecules, including prostaglandin E2 (PGE2), transforming growth factor β (TGFβ), IL10, arginase, nitric oxide (NO) and reactive oxygen species (28,43,44). MDSC-derived immunosuppressive molecules, especially PGE2, TGFβ and IL10, are also known as potent suppressants and/or modulators of NK cells and DCs (45-47). Therefore, our findings could indicate that MDSCs potently suppress NK cells and/or DCs, which leads to abrogation of their crosstalk in MCA-induced carcinogenesis. As NK cell/DC crosstalk is the central immunoregulatory mechanism that defines the quality and extent of effective anticancer immune responses, MDSC-mediated immunosuppression could be a critically important mechanism of cancer immune escape in MCA-induced carcinogenesis.
In conclusion, the manuscript reports on the novel findings that specific sTNF sequestration prevents chemically-induced carcinogenesis, expansion of MDSCs and suppression of Th1-type innate immunity in mice. The study reveals a pivotal role of sTNF in carcinogenesis, and defines a sTNF-TNFR1 negative immunoregulatory axis of innate immunity. The newly defined negative immunoregulatory axis balances the positive tmTNF-TNFR2 immunoregulatory axis of innate immunity (17-9). It also complements the well-established immune checkpoints of adaptive immunity, which targeting with antibodies leads to durable clinical responses in a large proportion of patients with advanced cancer (48-50). In this context, the sTNF-TNFR1 axis of innate immunity could be viewed as additional immune checkpoint, which targeting might be beneficial in both immunoprevention and immunotherapy of cancer.
Supplementary Material
Acknowledgements
We thank Dr. David Szymkowski (Xencor, Monrovia, CA) for providing us with the dominant-negative-TNF XPro1595 biologic.
Grant Support
This study was supported by NIH award P30CA047904. The study used the UPCI shared resources Flow Cytometry Facility and LUMINEX Laboratory supported in part by NIH P30CA047904.
Footnotes
Authors’ Contributions
Conception and design: N.L. Vujanovic and L. Vujanovic
Development of methodology: L. Vujanovic, A. DeLeo, A. Sobo-Vujanovic, F. Concha-Benavente and R.L. Ferris, N.L. Vujanovic
Acquisition of data (provided animals, acquired and managed patients, provided facilities, etc.): L. Vujanovic, A. Sobo-Vujanovic, F. Concha-Benavente, N.L. Vujanovic
Analysis and interpretation of data (e.g., statistical analysis, biostatistics, computation analysis): Y. Lin, L. Vujanovic, A. DeLeo, A. Sobo-Vujanovic, F. Concha-Benavente, N.L. Vujanovic
Writing, review, and/or revision of the manuscript: L. Vujanovic (writing), A. DeLeo (writing), F. Concha-Benavente (review), N.L. Vujanovic (writing)
Administrative, technical, or material support (i.e., reporting or organizing data, constructing databases): L. Vujanovic, A. DeLeo, R.L. Ferris, N.L. Vujanovic
Study supervision: N.L. Vujanovic
Disclosure of Potential Conflicts of Interest: RLF discloses service as a consultant for or on the advisory boards of ONO Pharmaceutical, Merck, Celgene, AstraZeneca, and Bristol Meyers-Squibb.
References
- 1.Balkwill F, Mantovani A. Inflammation and cancer: back to Virchow? Lancet. 2001;357:539–45. doi: 10.1016/S0140-6736(00)04046-0. [DOI] [PubMed] [Google Scholar]
- 2.Demaria S, Pikarsky E, Karin M, Coussens LM, Chen YC, El-Omar EM, Trinichieri G, et al. Cancer and inflammation: promise for biological therapy. J Immunother. 2010;33:335–51. doi: 10.1097/CJI.0b013e3181d32e74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bidwell J, Keen L, Gallagher G, Kimberly R, Huizinga T, McDermott M F, et al. Cytokine gene polymorphism in human disease: on-line databases. Genes Immun. 1999;1:3–19. doi: 10.1038/sj.gene.6363645. [DOI] [PubMed] [Google Scholar]
- 4.Howell WM, Calder PC, Grimble RF. Gene polymorphisms, inflammatory diseases and cancer. Proc Nutr Soc. 2002;61:447–56. doi: 10.1079/pns2002186. [DOI] [PubMed] [Google Scholar]
- 5.Karin M, Greten FR. NF-κB linking inflammation and immunity to cancer development and progression. Nature Rev Immunol. 2005;5:749–59. doi: 10.1038/nri1703. [DOI] [PubMed] [Google Scholar]
- 6.Pikarsky E, Porat RM, Stein I, Abramovitch R, Amit S, Kasem S, et al. NF-κB functions as a tumor promoter in inflammation-associated cancer. Nature. 2004;431:461–6. doi: 10.1038/nature02924. [DOI] [PubMed] [Google Scholar]
- 7.Greten F R, Eckmann L, Greten TF, Park JM, Li ZW, Egan LJ, et al. IKKβ links inflammation and tumorigenesis in a mouse model of colitis-associated cancer. Cell. 2004;118:285–96. doi: 10.1016/j.cell.2004.07.013. [DOI] [PubMed] [Google Scholar]
- 8.Tu S, Bhagat G, Cui G, Takaishi S, Kurt-Jones EA, Rickman B, Betz KS, et al. Overexpression of interleukin-1β induces gastric inflammation and cancer and mobilizes myeloid-derived suppressor cells in mice. Cancer Cell. 2008;14:408–19. doi: 10.1016/j.ccr.2008.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Moore RJ, Owens DM, Stamp G, Arnott C, Burke F, East N, et al. Mice deficient in tumor necrosis factor-α are resistant to skin carcinogenesis. Nat Med. 1999;5:828–31. doi: 10.1038/10552. [DOI] [PubMed] [Google Scholar]
- 10.Suganuma M, Okabe S, Marino MW, Sakai A, Sueoka E, Fujiki H. Esential role of tumor necrosis factor α (TNF-α) in tumor promotion as revealed by TNF-α-deficient mice. Cancer Res. 1999;59:4516–8. [PubMed] [Google Scholar]
- 11.Popivanova BK, Kitamura K, Wu Y, Kondo T, Kagaya T, Kaneko S, et al. Blocking TNF-α in mice reduces colorectal carcinogenesis associated with chronic colitis. J Clin Invest. 2008;118:560–70. doi: 10.1172/JCI32453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Jaiswal M, LaRusso NF, Burgart LJ, Gores GJ. Inflammatory cytokines induce DNA damage and inhibit DNA repair in cholangiocarcinoma cells by nitric oxide-dependent mechanism. Cancer Res. 2000;60:184–90. [PubMed] [Google Scholar]
- 13.Komori A, Yatsunami J, Suganuma M, Okabe S, Abe S, Sakai A, et al. Tumor necrosis factor acts as a tumor promoter in BALB/3T3 cell transformation. Cancer Res. 1993;53:1982–5. [PubMed] [Google Scholar]
- 14.Philip M, Rowley DA, Schreiber H. Inflammation as a tumor promoter in cancer induction. Semin Cancer Biol. 2004;14:433–9. doi: 10.1016/j.semcancer.2004.06.006. [DOI] [PubMed] [Google Scholar]
- 15.Brenner D, Blaser H, Mak TW. Regulation of tumour necrosis factor signaling: live or let die. Nat Rev Immunol. 2015;15:362–74. doi: 10.1038/nri3834. [DOI] [PubMed] [Google Scholar]
- 16.Naude PJ, den Boer JA, Luiten PG, Eisel UL. Tumor necrosis factor receptor cross-talk. FEBS J. 2011;278:888–98. doi: 10.1111/j.1742-4658.2011.08017.x. [DOI] [PubMed] [Google Scholar]
- 17.Xu J, Chakrabarti AK, Tan JL, Ge L, Gambotto A, Vujanovic NL. Essential role of the TNF-TNFR2 cognate interaction in mouse dendritic cell-natural killer cell cross-talk. Blood. 2007;109:3333–41. doi: 10.1182/blood-2006-06-026385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vujanovic L, Szymkowski DE, Alber S, Watkins SC, Vujanovic NL, Butterfield LH. Virally-infected and matured human dendritic cells activate natural killer cells via cooperative activity of plasma membrane-bound TNF and IL-15. Blood. 2010;116:575–83. doi: 10.1182/blood-2009-08-240325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Vujanovic NL. Role of TNF superfamily ligands in innate immunity. Immunol Res. 2011;50:159–74. doi: 10.1007/s12026-011-8228-8. [DOI] [PubMed] [Google Scholar]
- 20.Steed PM, Tansey MG, Zalevsky J, Zhukovsky EA, Desjarlais JR, Szymkowski DE, et al. Inactivation of TNF signaling by rationally designed dominant-negative TNF variants. Science. 2003;301:1895–8. doi: 10.1126/science.1081297. [DOI] [PubMed] [Google Scholar]
- 21.Van Hauwermeiren F, Vandenbroucke RE, Libert C. Treatment of TNF mediated diseases by selective inhibition of soluble TNF or TNFR1. Cytokine Growth Factor Rev. 2011;22:311–9. doi: 10.1016/j.cytogfr.2011.09.004. [DOI] [PubMed] [Google Scholar]
- 22.Sobo-Vujanovic A, Munich S, Vujanovic NL. Dendritic-cell exosomes cross-present Toll-like receptor-ligands and activate bystander dendritic cells. Cell Immunol. 2014;289:119–27. doi: 10.1016/j.cellimm.2014.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Krutzik PO, Nolan GP. Intracellular phospho-protein staining techniques for flow cytometry: monitoring single cell signaling events. Cytometry A. 2003;55:61–70. doi: 10.1002/cyto.a.10072. [DOI] [PubMed] [Google Scholar]
- 24.Cicinnati VR, Dworacki G, Albers A, Beckebaum S, Tuting T, Kaczmarek E, et al. Impact of p53-based immunization on primary chemically-induced tumors. Int J Cancer. 2005;113:961–70. doi: 10.1002/ijc.20686. [DOI] [PubMed] [Google Scholar]
- 25.Wojdani A, Alfred LJ. Alternations in cell-mediated immune functions induced in mouse splenic lymphocytes by polycyclic aromatic hydrocarbons. Cancer Res. 1984;44:942–5. [PubMed] [Google Scholar]
- 26.Horiguchi S, Petersson M, Nakazawa T, Kanda M, Zea AH, Ochoa AC, Kissling R. Primary chemically induced tumors induce profound immunosuppression concomitant with apoptosis and alteration in signal transduction in T cells and NK cells. Cancer Res. 1999;59:2950–6. [PubMed] [Google Scholar]
- 27.Baskic D, Vujanovic L, Arsenijevic N, Whiteside TL, Myers EN, Vujanovic NL. Natural killer cell and dendritic cell apoptotic tumoricidal activity is suppressed in head and neck cancer patients. Head Neck. 2013;35:388–98. doi: 10.1002/hed.22968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gabrilovich DI, Nagaraj S. Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol. 2009;9:162–74. doi: 10.1038/nri2506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Vasquez-Dunddel D, Pan F, Gorbounov M, Albesiano E, Fu J, Blosser RL, et al. STAT3 regulates arginase-I in myeloid-derived suppressor cells from cancer patients. J Clin Invest. 2013;123:1580–9. doi: 10.1172/JCI60083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Moretta A. Natural killer cells and dendritic cells: rendezvous in abused tissues. Nat Rev Immunol. 2002;2:957–64. doi: 10.1038/nri956. [DOI] [PubMed] [Google Scholar]
- 31.Cooper MA, Fehniger TA, Fuchs A, Colonna M, Caligiuri MA. NK cell and DC interactions. Trends Immunol. 2004;25:47–52. doi: 10.1016/j.it.2003.10.012. [DOI] [PubMed] [Google Scholar]
- 32.Walzer T, Dalod M, Robbins SH, Zitvogel L, Vivier E. Natural killer cells and dendritic cells: “l'union fait la force”. Blood. 2005;106:2252–8. doi: 10.1182/blood-2005-03-1154. [DOI] [PubMed] [Google Scholar]
- 33.Makarenkova V, Chakrabarti AK, Liberatore JA, Popovic P, Lu G, Watkins S, et al. Dendritic cells and natural killer cells interact via multiple TNF family molecules. J Leukoc Biol. 2005;77:408–13. doi: 10.1189/jlb.1104675. [DOI] [PubMed] [Google Scholar]
- 34.Gabrilovich DI, Ciernik IF, Carbone DP. Dendritic cells in antitumor immune responses. I. Defective antigen presentation in tumor-bearing hosts. Cell Immunol. 1996;170:101–10. doi: 10.1006/cimm.1996.0139. [DOI] [PubMed] [Google Scholar]
- 35.Tracey D, Klareskog L, Sasso EH, Salfeld JG, Tak PP. Tumor necrosis factor antagonist mechanisms of action: a comprehensive review. Pharmacol Ther. 2008;117:244–79. doi: 10.1016/j.pharmthera.2007.10.001. [DOI] [PubMed] [Google Scholar]
- 36.Vujanovic NL, Nagashima S, Herberman RB, Whiteside TL. Non-secretory apoptotic killing by human NK cells. J Immunol. 1996;157:1117–26. [PubMed] [Google Scholar]
- 37.Lu G, Janjic BM, Janjic J, Whiteside TL, Storkus WJ, Vujanovic NL. Innate direct anticancer effector function of human immature dendritic cells. II. Role of TNF, lymphotoxin-alpha(1)beta(2), Fas ligand, and TNF-related apoptosis-inducing ligand. J Immunol. 2002;168:1831–9. doi: 10.4049/jimmunol.168.4.1831. [DOI] [PubMed] [Google Scholar]
- 38.Tang D, Kang R, Coyne CB, Zeh HJ, Lotze MT. PAMPs and DAMPs: signal 0s that spur autophagy and immunity. Immunol Rev. 2012;249:158–75. doi: 10.1111/j.1600-065X.2012.01146.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Apte RN, Dotan S, Elkabets M, White MR, Reich E, Carmi Y, et al. The involvement of IL-1 in tumorigenesis, tumor invasiveness, metastasis and tumor-host interaction. Cancer Metastasis Rev. 2006;25:387–408. doi: 10.1007/s10555-006-9004-4. [DOI] [PubMed] [Google Scholar]
- 40.Sade-Feldman M, Kanterman J, Ish-Shalom E, Elnekava M, Horwitz E, Baniyash M. Tumor necrosis factor-α blocks differentiation and enhances suppressive activity of immature myeloid cells during chronic inflammation. Immunity. 2013;38:541–54. doi: 10.1016/j.immuni.2013.02.007. [DOI] [PubMed] [Google Scholar]
- 41.Zhao X, Rong L, Zhao X, Li X, Liu X, Deng J, et al. TNF signaling drives myeloid-derived suppressor cell accumulation. J Clin Invest. 2012;122:4094–104. doi: 10.1172/JCI64115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hu X, Li B, Li X, Zhao X, Wan L, Lin G, et al. Transmembrane TNF-α promotes suppressive activities of myeloid-derived suppressor cells via TNFR2. J Immunol. 2014;192:1320–31. doi: 10.4049/jimmunol.1203195. [DOI] [PubMed] [Google Scholar]
- 43.Serafini P. Editorial: PGE2-producing MDSC: a role in tumor progression. J Leukoc Biol. 2010;88:827–9. doi: 10.1189/jlb.0510303. [DOI] [PubMed] [Google Scholar]
- 44.Yang L, Moses HL. Transforming growth factor β: tumor suppressor or promoter? Are host immune cells the answer? Cancer Res. 2008;68:9107–11. doi: 10.1158/0008-5472.CAN-08-2556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Harizi H. Reciprocal crosstalk between dendritic cells and natural killer cells under the effects of PGE2 in immunity and immunopathology. Cell Mol Immunol. 2013;10:213–21. doi: 10.1038/cmi.2013.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Li MO, Wan YY, Sanjabi S, Robertson AK, Flavell RA. Transforming growth factor-β regulation of immune responses. Annu Rev Immunol. 2006;24:99–146. doi: 10.1146/annurev.immunol.24.021605.090737. [DOI] [PubMed] [Google Scholar]
- 47.Moore KW, de Waal Malefyt R, Coffman RL, O'Garra A. Interleukin-10 and the interleukin-10 receptor. Annu Rev Immunol. 2001;19:683–765. doi: 10.1146/annurev.immunol.19.1.683. [DOI] [PubMed] [Google Scholar]
- 48.Postow MA, Harding J, Wolchok JD. Targeting immune checkpoints releasing the restrains on anti-tumor immunity for patients with melanoma. Cancer J. 2012;18:153–9. [PMC free article] [PubMed] [Google Scholar]
- 49.Topalian SL, Drake CG, Pardoll DM. Targeting the PD-1/B7-H1(PD-L1) pathway to activate anti-tumor immunity. Cur Opin Immunol. 2012;24:207–12. doi: 10.1016/j.coi.2011.12.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Ott PA, Hodi FS, Robert C. CTLA-4 and PD-1/PD-L1 blockade: New immunotherapeutic modalities with durable clinical benefit in melanoma patients. Clin Cancer Res. 2013;19:5300–9. doi: 10.1158/1078-0432.CCR-13-0143. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.