

Abstract
Bacterial resistance to conventional antibiotics has become a clinical and public health problem, making therapeutic decisions more challenging. Plant compounds and nanodrugs have been proposed as potential antimicrobial alternatives. Studies have shown that oregano (Origanum vulgare) essential oil (OEO) and silver nanoparticles have potent antibacterial activity, also against multidrug-resistant strains; however, the strong organoleptic characteristics of OEO and the development of resistance to these metal nanoparticles can limit their use. This study evaluated the antibacterial effect of a two-drug combination of biologically synthesized silver nanoparticles (bio-AgNP), produced by Fusarium oxysporum, and OEO against Gram-positive and Gram-negative bacteria, including multidrug-resistant strains. OEO and bio-AgNP showed bactericidal effects against all 17 strains tested, with minimal inhibitory concentrations (MIC) ranging from 0.298 to 1.193 mg/mL and 62.5 to 250 μM, respectively. Time-kill curves indicated that OEO acted rapidly (within 10 min), while the metallic nanoparticles took 4 h to kill Gram-negative bacteria and 24 h to kill Gram-positive bacteria. The combination of the two compounds resulted in a synergistic or additive effect, reducing their MIC values and reducing the time of action compared to bio-AgNP used alone, i.e., 20 min for Gram-negative bacteria and 7 h for Gram-positive bacteria. Scanning electron microscopy (SEM) revealed similar morphological alterations in Staphylococcus aureus (non-methicillin-resistant S. aureus, non-MRSA) cells exposed to three different treatments (OEO, bio-AgNP and combination of the two), which appeared cell surface blebbing. Individual and combined treatments showed reduction in cell density and decrease in exopolysaccharide matrix compared to untreated bacterial cells. It indicated that this composition have an antimicrobial activity against S. aureus by disrupting cells. Both compounds showed very low hemolytic activity, especially at MIC levels. This study describes for the first time the synergistic and additive interaction between OEO and bio-AgNP produced by F. oxysporum against multidrug-resistant bacteria, such as MRSA, and β-lactamase- and carbapenemase-producing Escherichia coli and Acinetobacter baumannii strains. These results indicated that this combination can be an alternative in the control of infections with few or no treatment options.
Keywords: oregano oil, biological silver nanoparticles, multidrug-resistant bacteria, synergism, antibacterial
Introduction
Bacterial antimicrobial resistance to most conventional antibiotics has become a clinical and public health problem. Infections due to multidrug-resistant microorganisms, such as bacteria that produce extended-spectrum β-lactamases (ESBL) and carbapenemases (Klebsiella pneumoniae carbapenemase, KPC) and MRSA can be challenging to control leading to high treatment costs, therapeutic failure and death (Silva and Lincopan, 2012; Cantas et al., 2013).
ESBL and KPC hydrolyze the β-lactam ring resulting in an inactive antimicrobial (Queenan and Bush, 2007; Drawz and Bonomo, 2010). ESBLs mediate resistance to most β-lactams, mainly in Gram-negative bacteria (Dhillon and Clark, 2012; Silva and Lincopan, 2012). In these cases, carbapenem antibiotics, such as imipenem, meropenem, and ertapenem, are drugs of choice for treatment. However, carbapenemases reduce treatment options because they inactivate penicillins, cephalosporins, monobactams, and carbapenems (Queenan and Bush, 2007).
Usually ESBLs and KPC are encoded by genes carried by mobile genetic elements which also carry resistance genes to other antimicrobial agents, contributing to the emergence of multidrug resistance and its rapid spread between different strains and species (Pitout, 2012; Silva and Lincopan, 2012; Patel and Bonomo, 2013; Shaikh et al., 2015). ESBL- and KPC-producing strains usually exhibit resistance to quinolones, tetracyclines, cotrimoxazol, trimethoprim, and aminoglycosides (Dhillon and Clark, 2012; Pitout, 2012; Patel and Bonomo, 2013).
In Gram-positive bacteria, the most common mechanism of resistance to β-lactam occurs due a mutant transpeptidase gene. Methicillin resistance in S. aureus occurs because of the mecA gene, which encodes transpeptidases that have low affinity for β-lactam antibiotics (Rice, 2012). MRSA strains can harbor resistance genes for other antimicrobials besides mecA, reducing treatment options (Turlej et al., 2011).
Multiresistant bacteria have been found in foods for human consumption, streams and effluents and thus it is a hospital, community, and environmental problem (Prado et al., 2007; Fontes et al., 2011; Silva and Lincopan, 2012; Rubin et al., 2014; Koga et al., 2015). As soon as new β-lactams are widely used, new β-lactamases are also discovered (Bush, 2010; Kong et al., 2010; Shaikh et al., 2015). Therefore, there is crucial need for research and development of new drugs with potential to combat resistant strains to minimize their selection.
Compounds from natural sources such as animals, plants, and microorganisms have been proposed as potential antimicrobial alternatives (Mandal and Mandal, 2011; Cardozo et al., 2013; Biasi-Garbin et al., 2015). Spice essential oils have been documented as being antimicrobial (Burt, 2004; Du et al., 2009; Betancourt et al., 2012; Nazzaro et al., 2013). Oil phenolic substances are mainly responsible for antibiotic properties (Rhayour et al., 2003; Burt, 2004; Nostro et al., 2007; Hyldgaard et al., 2012).
Studies on the antibacterial mechanisms of action of plant essential oils, including the OEO, suggest that hydrophobic bioactive compounds cause damage to cell membrane, increase cell permeability, affect ATP production, act on protein synthesis, cause cellular pH disturbance, induce cytoplasmic changes and interfere with quorum sensing (Helander et al., 1998; Ultee et al., 1999; Lambert et al., 2001; Rhayour et al., 2003; Souza et al., 2010, 2013; Szabó et al., 2010; Hyldgaard et al., 2012).
OEO, extracted mainly from the leaves of the herb O. vulgare by ecofriendly methods, has been reported as having broad antibacterial activity (Burt, 2004; Busatta et al., 2007; Betancourt et al., 2012). Other biological activities such as antifungal, antiviral, antioxidant and anticancer have been described for OEO (Cervato et al., 2000; Kalemba and Kunicka, 2003; Hyldgaard et al., 2012; Gautam et al., 2014; Gilling et al., 2014; Sobral et al., 2014).
Carvacrol and thymol are the main components of OEO (Nostro et al., 2004; Cleff et al., 2008; Hyldgaard et al., 2012; Stojković et al., 2013), and the antimicrobial activity of this oil varies according to their amounts. Synergistic and additive interactions between carvacrol and thymol have been reported (Bassolé and Juliani, 2012; Hyldgaard et al., 2012); furthermore, a mixture of compounds with antimicrobial activity could minimize the selection of resistant strains. Studies have shown that OEO has the potential to prevent food from being contaminated and control worrisome hospital infections (Nostro et al., 2004; Si et al., 2008; Barros et al., 2009; Amrouni et al., 2014; Honório et al., 2015). Despite the potential antimicrobial activity of OEO, its strong taste and smell seem to limit its use, so alternatives are needed to minimize or eliminate such undesirable characteristics (Burt, 2004; Alvarez et al., 2014).
Silver has been used for millennia to treat wounds and eye infections and to preserve food and water (Alexander, 2009). Nanotechnology has proved to be a useful tool for solving biomedical problems. Silver nanoparticles have been intensively studied as antimicrobial agents, including their use against multidrug-resistant bacteria (Li et al., 2010; Cardozo et al., 2013; Naqvi et al., 2013; Ansari et al., 2014; Bibbs et al., 2014; Palanisamy et al., 2014; Singh et al., 2014; Subashini et al., 2014; Theophel et al., 2014). Through nanotechnology, it has been possible to revive the use of silver to combat resistant bacteria. Nanoscale enhances the antibacterial activity of silver even at low concentrations; nanometer metallic particles show altered physical, chemical and biological properties compared to conventional silver, due to their high surface-to-volume ratio (Nowack et al., 2011; Rai et al., 2012; Herman and Herman, 2014). Furthermore, silver nanoparticles have been reported to be less toxic than silver ions to host (de Lima et al., 2012).
The bactericidal mechanism of silver nanoparticles is not clearly understood; however, it is believed that there is a multiple mechanism of action (Herman and Herman, 2014). Researchers have suggested that silver nanoparticles cause cell membrane disintegration and increase cell surface permeability, inactivate bacterial enzymes by interaction with thiol groups, deplete levels of intracellular ATP, cause DNA damage and induce free radical formation (Feng et al., 2000; Dibrov et al., 2002; Lok et al., 2006; Li et al., 2010; Kim et al., 2011; Theophel et al., 2014).
Although silver nanoparticles show excellent antimicrobial activity, silver-resistant bacteria have been described; and these microorganisms can quickly develop resistance to silver nanoparticles by genetic alterations (Losasso et al., 2014; Graves et al., 2015).
The combination of several antimicrobials seems to be the best strategy for controlling emergence of antibiotic-resistant microorganisms (Fischbach, 2011; Bass et al., 2015; Bollenbach, 2015). Many studies show additive or synergistic antibacterial effects of silver nanoparticles combined with alternative (eugenol, phenazine-1-carboxamide, and cinnamaldehyde) and conventional (ampicillin, kanamycin, erythromycin, chloramphenicol, amoxicillin, ciprofloxacin, and moxifloxacin; Li et al., 2005; Fayaz et al., 2010; Cardozo et al., 2013; Ghosh et al., 2013; Theophel et al., 2014; Biasi-Garbin et al., 2015) antimicrobial compounds. Synergistic and additive antimicrobial effects have been reported for OEO or its components combined with eugenol, linalool, menthol, cinnamaldehyde, Thymus vulgaris essential oil, Rosmarinus officinalis essential oil, gentamicin, polymyxin, kanamycin, levofloxacin or many others (Rosato et al., 2007; Si et al., 2008; de Souza et al., 2009; Bassolé and Juliani, 2012; Stojković et al., 2013; Honório et al., 2015).
Biological systems have been extensively used for rapid and eco-friendly biosynthesis of silver nanoparticles; instead of using chemical reagents, these processes involve fungi, bacteria, plants and other organisms or biomolecules (Sastry et al., 2003; Durán et al., 2005, 2011; Sankar et al., 2013; Singh et al., 2014). This study evaluated, for the first time, the antibacterial effect of a two-drug combination composed of bio-AgNP synthesized by F. oxysporum and OEO against standard and resistant Gram-positive and Gram-negative strains.
Materials and methods
Bacterial strains
All antimicrobial assays were performed against reference strains from American Type Culture Collection (ATCC) provided by the Laboratory of Basic and Applied Bacteriology of Londrina Stated University (Londrina, Paraná, Brazil), and clinical strains provided from Londrina University Hospital (Londrina, Paraná, Brazil). The standard bacterial strains used were as follows: methicillin-sensitive S. aureus (ATCC 25923), Streptococcus mutans (ATCC 25175), E. coli (ATCC 25922), K. pneumoniae (ATCC 10031), K. pneumoniae (ATCC 700603), Salmonella enterica serovar Enteritidis (ATCC 13076), S. enterica serovar Typhimurium UK-1 (ATCC 68169) and MRSA N315 provided by Dr. Elza Masae Mamizuka (São Paulo University, São Paulo – SP, Brazil). Several clinical isolates obtained from Londrina University Hospital were also tested (Table 1). Four isolates of ESBL-producing E. coli and two isolates of carbapenemase-producing E. coli from urinary tract infections (provided by Dr. Eliana Carolina Vespero, Londrina University Hospital, Londrina); one isolate of carbapenem-resistant A. baumannii from inguinal-rectal swabs (provided by Dr. Floristher Elaine Carrara Marroni, Londrina Clinics Hospital, Londrina) and two isolates of MRSA strains from secretions-general discharges (provided by Dr. Marcia Regina Eches Perugini, Londrina University Hospital, Londrina). The bacterial strains were stored in 25% glycerol (Merck) at −80°C.
Table 1.
Susceptibility to antibiotics of clinical strains.
| S. aureus | PEN | OXA | CIP | RIF | GEN | STR | TET | ERY | CLI | LNZ | SXT | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MRSA 101 | R | R | R | S | S | S | S | R | R | S | S | ||||||
| MRSA 107 | R | R | R | S | R | R | S | R | R | S | R | ||||||
| E. coli | PTZ | CEP | CFZ | CTX | CAZ | CPM | AZT | IPM | MRP | ERT | NOR | CIP | LVX | GEN | AMI | SXT | NIT |
| ESBL 167 | S | – | R | R | R | R | R | S | S | S | – | R | R | S | S | R | S |
| ESBL 169 | S | R | – | R | R | R | R | S | S | S | R | R | S | S | S | S | S |
| ESBL 176 | S | R | – | R | R | R | I | S | S | S | – | S | S | S | S | S | S |
| ESBL 192 | S | S | – | R | R | I | S | - | S | S | – | R | R | S | S | S | S |
| KPC 131 | R | – | – | R | S | S | R | R | R | R | – | R | – | – | S | – | – |
| KPC 133 | I | – | – | R | I | I | R | S | R | R | – | R | – | – | S | – | – |
| A. baumannii | PTZ | AMS | CTX | CAZ | CPM | AZT | IPM | MRP | ERT | CIP | LVX | GEN | AMI | TET | SXT | ||
| CR 01 | R | R | R | R | R | R | R | R | R | R | R | R | R | R | R | ||
AMI, amikacin; AMS, ampicillin + sulbactam; AZT, aztreonam; CAZ, ceftazidime; CEP, cephalothin; CFZ, cefazolin; CIP, ciprofloxacin; CLI, clindamycin; CPM, cefepime; CTX, cefotaxime; ERY, erythromycin; ETP, ertapenem; GEN, gentamicin; IPM, imipenem, LNZ, linezolid; LVX, levofloxacin; MRP, meropenem; NIT, nitrofurantoin; NOR, norfloxacin; OXA, oxacillin; PEN, penicillin; PTZ, piperacillin + tazobactam; RIF, rifampin; STR, streptomycin; SXT, trimethoprim + sulfamethoxazole; TET, tetracycline.
R, resistant; S, susceptible; I, intermediate; (-), not tested.
MRSA, methicillin-resistant S. aureus.
ESBL, extended-spectrum beta-lactamases; KPC, K. pneumoniae carbapenemase; RC, carbapenem-resistant.
101, 107, 167, 169, 176,192, 131, 133, and 01 are strain numbers at Laboratory of Basic and Applied Bacteriology-Universidade Estadual de Londrina.
Antimicrobial agents
Oregano essential oil
OEO was obtained from Ferquima Industry and Commerce of Essential Oils (São Paulo, Brazil). This oil (batch 224) was extracted by steam distillation and its density (0.954 g/mL) and composition (main components: 71% carvacrol, 3% thymol, 4.5% gamma terpinene, 3.5% para-cymene, and 4% beta-caryophyllene) were described in a technical report. A stock solution of 50% OEO was prepared in dimethylsulfoxide (DMSO, Sigma-Aldrich; v/v). DMSO maximum concentration in assays was 1%.
Silver nanoparticles
Bio-AgNP were prepared according to a previously established method (Durán et al., 2005, 2006). This method of production has been patented (Patent, 2006, PI 0605681-4A2; http://www.inpi.gov.br). Briefly, bio-AgNP were obtained after reduction of silver nitrate by F. oxysporum, strain 551, from the culture collection of the Molecular Genetics Laboratory of ESALQ-USP (Piracicaba, São Paulo, Brazil). F. oxysporum was cultivated on media containing 0.5% (w/v) yeast extract (Neogen), 2% (w/v) malt extract (Neogen), 2% (w/v) agar (Neogen) and distilled water at 28°C for 7 days. After growth, the fungal biomass was added to distilled water at 0.1 g/mL and incubated at 28°C for 72 h. Afterwards, the solution components were separated by filtration. AgNO3 (Nuclear) at 1 mM was added to fungal-free solution, and the system was incubated for several hours at 28°C in the absence of light. Periodically, aliquots of the solution system were removed and absorptions were measured using an ultraviolet-visible spectrophotometry (Varian Cary 50 Probe); the peak at 440 nm corresponded to the surface plasmon resonance of silver nanoparticles. After bio-AgNP purification, diameter was determined by photon correlation spectroscopy using ZetaSizer NanoZS (Malvern), and zeta potential measurement was performed using the same instrument.
Antibacterial activity of OEO and Bio-AgNP separately
MIC was determined by the broth dilution method according to Clinical and Laboratory Standards Institute guidelines (CLSI, 2012), with necessary modifications. Tested concentrations of OEO and bio-AgNP ranged from 0.075 to 9.540 mg/mL and 1.91 to 500 μM, respectively. Mueller-Hinton broth (MHB, Difco) alone, MHB plus OEO and MHB plus bio-AgNP were tested as sterility controls, and untreated bacteria inoculated on MHB alone and with 1% DMSO were tested as growth control. After 24 h incubation at 37°C, MIC was defined as the lowest concentration of antimicrobial agent that inhibited visible growth. Minimal bactericidal concentration (MBC) was determined by subculturing 10 μL from the broth dilution MIC, after 24 h of treatment, in Mueller-Hinton agar (MHA, Oxoid) with no antimicrobial agent. MBC was defined as the lowest concentration that kills ≥99.9% of bacteria after 24 h of antimicrobial treatment (NCCLS, 1999). All assays were carried out in triplicate, and at least on three different occasions.
Antibacterial combination assay
Interaction of OEO and bio-AgNP was determined by broth dilution in double-antimicrobial gradient as described by Traub and Kleber (1975), with modifications. Single colonies of bacterial cultures grown in MHA media were suspended in saline solution (0.9% sodium chloride, w/v, Merck) and adjusted to 0.5 McFarland suspension which corresponds to 1 × 108 colony-forming units/mL (CFU/mL). The inoculum at 1 × 108 CFU/mL was diluted 1:100 in MHB to yield 106 CFU/mL. A volume of 0.05 mL of bacterial inoculum at 106 CFU/mL was added to 0.05 mL of MHB complemented with combination of OEO and bio-AgNP whose concentrations ranged from 0.037 to 0.596 mg/mL and from 7.81 to 125 μM, respectively. Finally bacteria at 5 × 105 CFU/mL were grown in MHB with both antimicrobial agents in combination at 37°C for 24 h. Sterility and growth controls were performed as in the MIC determination assay described above. The interaction of both compounds was analyzed by fractional inhibitory concentrations index (FICI) according Chin et al. (1997), using the following equation: FICI = FICOEO + FICbio-AgNP, where FIC = MICcombination/MICindividual. FICI is interpreted as “synergistic” when ≤ 0.5, as “additive” when >0.5 and ≤ 1, as “indifferent” (no interaction) when >1 and < 4 and as “antagonist” when ≥4. All assays were carried out in triplicate, at least on three different occasions and against Gram-positive and Gram-negative bacteria, including multidrug-resistant strains.
Time-kill assay
Time-kill assay was carried out using the viable cells count method, according to NCCLS (1999), with modifications. Three conditions of treatments were tested, bacterial cultures treated with OEO, bio-AgNP and their combination; and a bacterial culture without antimicrobial agent served as growth control. At ten time points (0 h, 30 s, 10 min, 20 min, 30 min, 2 h, 4 h, 7 h, 10 h, and 24 h) of incubation at 37°C, 10 μL from serial dilutions of treated and untreated cultures were transferred to MHA and CFU/mL were determined. All assays were carried out in triplicate, and at least on two different occasions, with three bacterial strains (S. aureus ATCC 25923, E. coli ATCC 25922 and carbapenemase-producing E. coli 131).
Scanning electron microscopy
S. aureus (ATCC 25923, non-MRSA) at 109 CFU/mL were prepared in MHB and added to five tubes (5 mL of bacterial inoculum each). S. aureus was exposed to three different treatments; OEO at 0.594 mg/mL was added to the first tube, bio-AgNP at 250 μM were added to the second tube and the two compounds combined were added to the third tube (OEO at 0.297 mg/mL and bio-AgNP at 125 μM, 2x synergistic MIC values to account for the increased cell density used for SEM analyses). Incubation times at 37°C varied for each treatment, 30 min for OEO and 6 h for bio-AgNP and combination of antimicrobials. Two untreated controls were prepared, one with 30 min incubation and another with 6 h. After incubation, 20 μL of each sample were spotted onto poly-L-lysine (Sigma-Aldrich)-coated glass slides. Each slide containing treated or untreated bacteria was fixed (for 20 h) by immersion in 1 mL of 0.1 M sodium cacodylate buffer (pH 7.2) containing 2.5% glutaraldehyde and 2% paraformaldehyde. All samples were then post-fixed in 1% osmium tetroxide for 2 h. All reagents for both chemical fixations were provided from Electron Microscopy Sciences. Post-fixed cells were dehydrated in an ethanol gradient (Sigma-Aldrich) (70, 80, 90 and 100°GL), critical point-dried using CO2 (BALTEC CPD 030 Critical Point Dryer), coated with gold (BALTEC SDC 050 Sputter Coater) and observed under a scanning electron microscope (FEI Quanta 200).
Cytotoxicity assay with human red blood cells (RBC) and HEp-2cells
Hemolytic activity of OEO and bio-AgNP was determined according to Izumi et al. (2012), with necessary modifications. Blood was collected in heparinized tubes (Vacutainer) from a healthy human donor with voluntary consent, which was approved by the human ethics committee (CAAE 47661115.0.0000.5231, No. 1.268.019 – UEL). Erythrocytes were separated by centrifugation (5000 rpm, 4°C, 5 min) and were prepared in phosphate-buffered saline (0.1 M PBS, pH 7.2) at 6% (v/v). PBS was composed of 0.9% (w/v) sodium chloride (Merck), 0.2 M monobasic sodium phosphate (Chemco) and 0.2 M dibasic sodium phosphate (Nuclear). In 96-well plates, 100 μL of RBC at 6% were added to 100 μL of PBS with different concentrations of compounds individually and in combination. After 3 h of incubation at 37°C, supernatants were read at 550 nm to monitor release of hemoglobin. Triton X-100 (Sigma-Aldrich) at 1% was used as control for 100% hemolytic activity, and hemolysis percentage was calculated for each compound concentration.
Cytotoxicity to the human laryngeal epithelial carcinoma cell line HEp-2 was performed by the colorimetric dimethylthiazol diphenyl tetrazolium bromide (MTT, Sigma-Aldrich) assay in 96-well plates, according to the manufacturer's recommendations. HEp-2 cells were grown in RPMI medium 1640 (Gibco) at 37°C in 5% CO2 to form a monolayer. Non-adherent cells were removed using PBS and confluent cells were treated with different concentrations of compounds individually and in combination for 24 h at 37°C in 5% CO2. After incubation, the medium was removed and each well was washed with PBS. MTT solution (10 μL per well at 1.250 g/mL) was added to all wells, and plates were incubated at 37°C for 2 h. MTT solubilization solution (Sigma-Aldrich) containing 10% Triton X-100 in acidic isopropanol (0.1 N HCl) was added to each well (90 μL per well) to dissolve the dark blue crystals (formazan). After 15 min homogenization, the plate was read at 570 nm. Untreated HEp-2 cells were used as control for 100% viability, and viability percentage was calculated for each compound concentration.
In both cytotoxicity assays, the concentrations of OEO and bio-AgNP ranged 0.037–9.540 mg/mL and 1.95–250 μM, respectively. The 50% cytotoxic concentration (CC50) was defined as the antimicrobial concentration required to reduce cell viability by 50% compared to untreated control. CC50 of each compound was determined by regression analysis for both cell lines, RBC and HEp-2 tumor cells.
Statistical method
Results of MIC and time kill assay were analyzed using Wilcoxon or Kruskal-Wallis test followed by Dunn's test. Analyses were performed using R Statistical Software, version 3.1.0 (Foundation for Statistical Computing, Vienna, Austria). Values of p < 0.05 were considered significant.
Results
Bio-AgNP characterization
Average bio-AgNP size and zeta potential were 77.68 nm and −34.6 mV, respectively (Supplementary Material).
MIC and antimicrobial interaction
OEO and bio-AgNP inhibited the growth of all bacterial strains tested, including multidrug-resistant strains. The mean MIC for OEO was 0.526 ± 0.130 mg/mL, and the mean MBC was 0.500 ± 0.158 mg/mL, MIC and MBC ranged from 0.298 to 1.193 mg/mL (Table 2). The mean MIC for bio-AgNP was 129.17 ± 55.25 μM ranging from 62.5 to 250μM, and the mean MBC was 154.17 ± 110.46 μM ranging from 62.5 μM to 500 μM (Table 2).
Table 2.
Mean of minimal inhibitory and bactericidal concentrations of oregano essential oil and biological silver nanoparticles.
| Bacteria | OEO (mg/mL) | bio-AgNP (μM) | ||
|---|---|---|---|---|
| MIC | MBC | MIC | MBC | |
| S. aureus (ATCC 25923) | 0.596 | 0.596 | 250.0 | 250.0 |
| S. mutans (ATCC 25175) | 0.596 | 0.596 | 125.0 | 250.0 |
| E. coli (ATCC 25922) | 0.596 | 0.596 | 62.50 | 62.50 |
| S. enterica Enteritidis (ATCC 13076) | 0.298 | 0.298 | 62.50 | 62.50 |
| K. pneumoniae (ATCC 10031) | 0.596 | 0.596 | 62.50 | 62.50 |
| K. pneumoniae (ATCC 700603) | 0.596 | 0.596 | 125.0 | 125.0 |
| S. enterica Typhimurium UK-1* | 0.298 | 0.298 | 125.0 | 125.0 |
| MRSA N315 | 0.596 | 0.596 | 250.0 | 250.0 |
| MRSA 101 | 1.193 | 1.193 | 250.0 | 500.0 |
| MRSA 107 | 1.193 | 1.193 | 250.0 | 500.0 |
| E. coli ESBL 167 | 0.596 | 0.596 | 125.0 | 125.0 |
| E. coli ESBL 169 | 0.596 | 0.596 | 125.0 | 125.0 |
| E. coli ESBL 176 | 0.596 | 0.596 | 125.0 | 125.0 |
| E. coli ESBL 192 | 0.596 | 0.596 | 125.0 | 125.0 |
| E. coli KPC 131 | 0.596 | 0.596 | 125.0 | 125.0 |
| E. coli KPC 133 | 0.596 | 0.596 | 125.0 | 125.0 |
| A. baumannii CR 01 | 0.298 | 0.298 | 125.0 | 125.0 |
OEO, oregano essential oil; bio-AgNP, biological silver nanoparticles.
MIC, minimal inhibitory concentration; MBC, minimal bactericidal concentration.
ATCC, American Type Culture Collection.
, ATCC 68169.
MRSA, methicillin resistant S. aureus.
ESBL, extended spectrum beta-lactamase; KPC, K. pneumoniae carbapenemase.
CR, Carbapenem-resistant;
101, 107, 167, 169, 176,192, 131, 133, and 01 are strain numbers at Laboratory of Basic and Applied Bacteriology-Universidade Estadual de Londrina.
There was no statistical difference between standard and multidrug-resistant strains with regard to MIC values of the two individual compounds (p > 0.05). No significant difference (p > 0.05) was observed between Gram-positive and Gram-negative sensitivity to OEO, where both bacterial groups were susceptible to this natural compound. Bio-AgNP showed better activity against Gram-negative bacteria (p < 0.05), where the mean MIC was 125 ± 56.53 μM in contrast to Gram-positive mean MIC, which was 225 ± 55.90 μM.
In combination, OEO and bio-AgNP showed significantly lower MIC values when compared with individual treatment (p < 0.05), where the two compounds together resulted in synergistic or additive antibacterial activity (Table 3).
Table 3.
Combinatory effect of oregano essential oil and biological silver nanoparticles and both minimal inhibitory concentrations in combination.
| MIC | FICI | ||
|---|---|---|---|
| Bacteria | OEO (mg/mL) | Bio-AgNP (μM) | |
| S. aureus (ATCC 25923) | 0.149 | 62.50 | 0.50 (S) |
| S. mutans (ATCC 25175) | 0.075 | 62.50 | 0.62 (A) |
| E. coli (ATCC 25922) | 0.298 | 15.62 | 0.75 (A) |
| S. enterica Enteritidis (ATCC 13076) | 0.037 | 31.25 | 0.62 (A) |
| K. pneumoniae (ATCC 10031) | 0.075 | 15.62 | 0.37 (S) |
| K. pneumoniae (ATCC 700603) | 0.075 | 62.50 | 0.62 (A) |
| S. enterica Typhimurium UK-1* | 0.149 | 62.50 | 1.00 (A) |
| MRSA N315 | 0.075 | 125.0 | 0.62 (A) |
| MRSA 101 | 0.596 | 62.50 | 0.75 (A) |
| MRSA 107 | 0.596 | 62.50 | 0.75 (A) |
| E. coli ESBL 167 | 0.149 | 15.62 | 0.37 (S) |
| E. coli ESBL 169 | 0.149 | 15.62 | 0.37 (S) |
| E. coli ESBL 176 | 0.149 | 31.25 | 0.50 (S) |
| E. coli ESBL 192 | 0.149 | 31.25 | 0.50 (S) |
| E. coli KPC 131 | 0.075 | 31.25 | 0.37 (S) |
| E. coli KPC 133 | 0.075 | 62.50 | 0.62 (A) |
| A. baumannii CR 01 | 0.149 | 15.62 | 0.62 (A) |
OEO, oregano essential oil; bio-AgNP, biological silver nanoparticles.
ATCC, American Type Culture Collection.
, S. enterica Typhimurium ATCC 68169.
MRSA, methicillin resistant S. aureus.
ESBL, extended spectrum beta-lactamase; KPC, K. pneumoniae carbapenemase.
CR, Carbapenem-resistant.
101, 107, 167, 169, 176,192, 131, 133 and 01 are strain numbers at Laboratory of Basic and Applied Bacteriology-Universidade Estadual de Londrina.
FICI, fractional inhibitory concentration index.
FIC index were interpreted as follows: ≤ 0.5, synergy; > 0.5 to 1.0, addition; > 1.0 to < 4.0, indifference; and ≥ 4, antagonism.
(S), Synergistic interaction; (A), Additive interaction.
Time-kill curve
OEO reduced the number of CFU/mL rapidly. For all three strains tested (S. aureus ATCC 25923, E. coli ATCC 25922, KPC-producing E. coli), after 10 min of treatment with OEO at MIC (0.596 mg/mL), there were no viable cells (p < 0.05; Figures 1–3). The bacterial inhibition by OEO was immediate, where after 30 s of treatment, there was a 0.57 log (p < 0.05), 1.54 log (p < 0.05), and 0.27 log (p < 0.05) reduction of S. aureus ATCC 25923 (Figure 1), E. coli ATCC 25922 (Figure 2), and carbapenemase-producing E. coli (Figure 3) cell populations, respectively.
Figure 1.
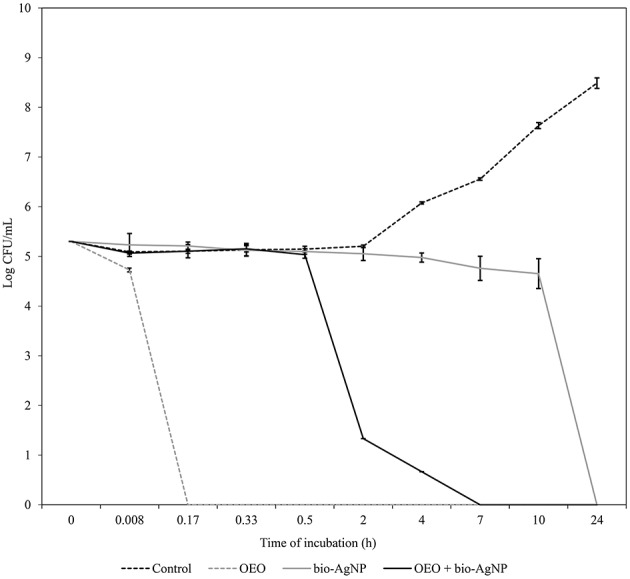
Time-kill curves of S. aureus 25923 (ATCC) exposed to OEO and bio-AgNP individually and in combination at MIC values. Bacteria at 5 × 105 CFU/mL were exposed to three different treatments; OEO alone (0.596 mg/mL), bio-AgNP alone (250μM), and OEO + bio-AgNP (0.298 mg/mL + 125 μM). Control indicates bacterial growth without antimicrobial compounds. Values of CFU/mL are the mean ± standard deviation.
Figure 2.
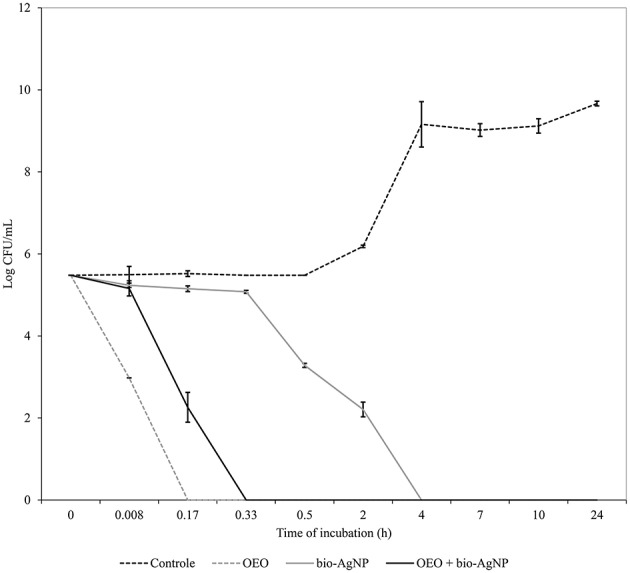
Time-kill curves of E. coli 25922 (ATCC) exposed to OEO and bio-AgNP individually and in combination at MIC values. Bacteria at 5 × 105 CFU/mL were exposed to three different treatments; OEO alone (0.596 mg/mL), bio-AgNP alone (62.5 μM) and OEO + bio-AgNP (0.298 mg/mL + 15.62 μM). Control indicates bacterial growth without antimicrobial compounds. Values of CFU/mL are the mean ± standard deviation.
Figure 3.
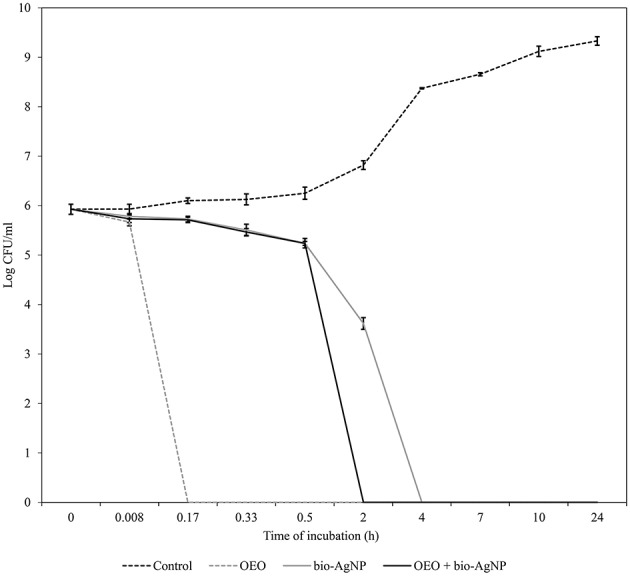
Time-kill curves of carbapenemase-producing E. coli exposed to OEO and bio-AgNP individually and in combination at MIC values. Bacteria at 5 × 105 CFU/mL were exposed to three different treatments; OEO alone (0.596 mg/mL), bio-AgNP alone (125 μM), and OEO + bio-AgNP (0.075 mg/mL + 31.25 μM). Control indicates bacterial growth without antimicrobial compounds. Values of CFU/mL are the mean ± standard deviation.
Bio-AgNP at MIC (250 μM) showed bactericidal effect at 24 h to S. aureus ATCC 25923 (p < 0.05; Figure 1). After 10 h of treatment, the S. aureus cell population decreased by 0.57 log (p < 0.05); and between 10 and 24 h of treatment, there was a 4.73 log reduction (p < 0.05). Gram-negative bacteria lost total viability after 4 h of incubation with bio-AgNP at 62.5 and 125 μM for E. coli ATCC 25922 (Figure 2) and carbapenemase-producing E. coli (Figure 3), respectively (p < 0.05). In both Gram-negative bacteria, after 2 h of exposure to bio-AgNP, there was a significant decrease in CFU/mL, 3.3 and 2.31 log reduction in standard E. coli (Figure 2) and KPC-producing E. coli (Figure 3), respectively (p < 0.05).
The combination of OEO and bio-AgNP led to faster reduction of CFU/mL than in individual treatment with bio-AgNP. Against S. aureus ATCC 25923, the combination of the two compounds at synergistic MIC (0.149 mg/mL and 62.5 μM for OEO and bio-AgNP, respectively) showed bacteriostatic activity (data not shown). But the combination with additive MIC values (0.298 mg/mL OEO and 125 μM bio-AgNP) against S. aureus ATCC 25923 caused a 3.48 log decrease in cell population in 2 h (p < 0.05) and resulted in no viable bacterial cells at 7 h (p < 0.05; Figure 1).
Against standard E. coli (ATCC 25922), OEO and bio-AgNP at additive MIC (0.298 mg/mL and 15.62 μM, respectively) caused a 2.3 log decrease (p < 0.05) within 10 min of treatment and there were no viable cells after 20 min (p < 0.05; Figure 2). At 30 min of incubation with synergistic MIC values (0.075 mg/mL and 31.25 μM for OEO and bio-AgNP, respectively), the population of carbapenemase-producing E. coli decreased 2.7 log (p < 0.05) and after 2 h there was a total reduction of bacterial population (p < 0.05; Figure 3).
Scanning electron microscopy
To investigate antimicrobial activity against S. aureus ATCC 25923 (non-MRSA) by OEO, bio-AgNP and combination of the two treatment, the morphology changes were investigated with SEM. Untreated cells showed their unique shapes (spherical shaped) 30 min (Figure 4A) and 6 h (Figure 4B) after incubation. Inset images showed, seen clearly in the higher magnification, intact surface of S. aureus ATCC 25923 (Figures 4A,B - inset). Although cells treated after 30 min with OEO alone was still present, most of them were damaged and extensively disappeared (Figure 4C). Inset showed details, in higher magnification, of morphological changes on the cells surface (Figure 4C - inset). Treatment with bio-AgNP alone after 6 h showed a decrease in cell density and caused morphological changes (Figure 4D). Inset show, in the higher magnification, cells walls covered with substance resulting from serious disruptions in the surface of S. aureus ATCC 25923 (Figure 4D - inset). Cells treated with combination of OEO plus bio-AgNP (Figure 4E) showed very deformed cells, with cell debris and damages appeared as cell surface blebbing after 6 h treatment (Figure 4E - inset). Treatments with OEO plus bio-AgNP showed a decrease in cell density, exopolysaccharide, morphology changes, and cell destruction, compared to the non-treated cells. It indicated that this composition have an antimicrobial activity against S. aureus by disrupting cells.
Figure 4.
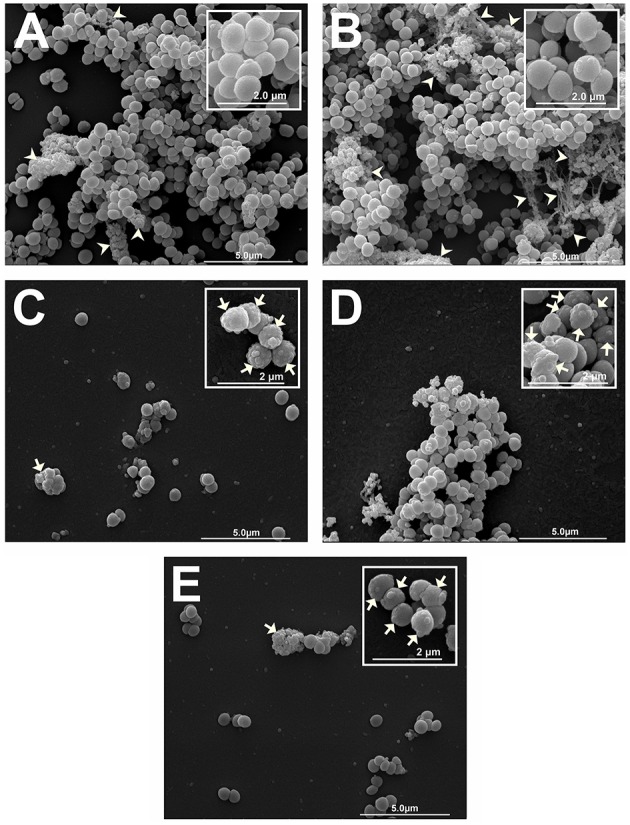
Scanning electron micrograph of antibacterial effect of OEO and bio-AgNP alone and in combination against S. aureus ATCC 25923 (non-MRSA). (A) Untreated control after 30 min incubation. (B) Untreated control after 6 h incubation. (C) Treated cells with OEO at 0.596 mg/mL for 30 min. (D) Treated cells with bio-AgNP at 250 μM for 6 h. (E) Treated cells with combination of OEO and bio-AgNP at 0.298 mg/mL and 125 μM, respectively, for 6 h. Micrographs (A–E) show cell density and exopolysaccharide matrix (15,000 x). Inset images show detail of morphological alterations of treated cells and typical cell morphology of untreated controls (30,000 x). Arrows: morphological changes (surface protrusions) and cellular debris. Arrowheads: exopolysaccharide.
Cytotoxicity assay in human RBC and HEp-2 cells
Results of MTT assay showed that OEO was toxic to tumor cells (HEp-2 cells), reducing cell metabolism by 50% or more at concentrations higher than 0.075 mg/mL (Figure 5A). But this essential oil was less toxic to RBC, and the CC50 was 7.519 mg/mL (Figure 6A). CC50 of bio-AgNP in HEp-2 cells was 97.22 μM (Figure 5B), and very low hemolytic activity was observed even at the highest concentration tested (250 μM; Figure 6B).
Figure 5.
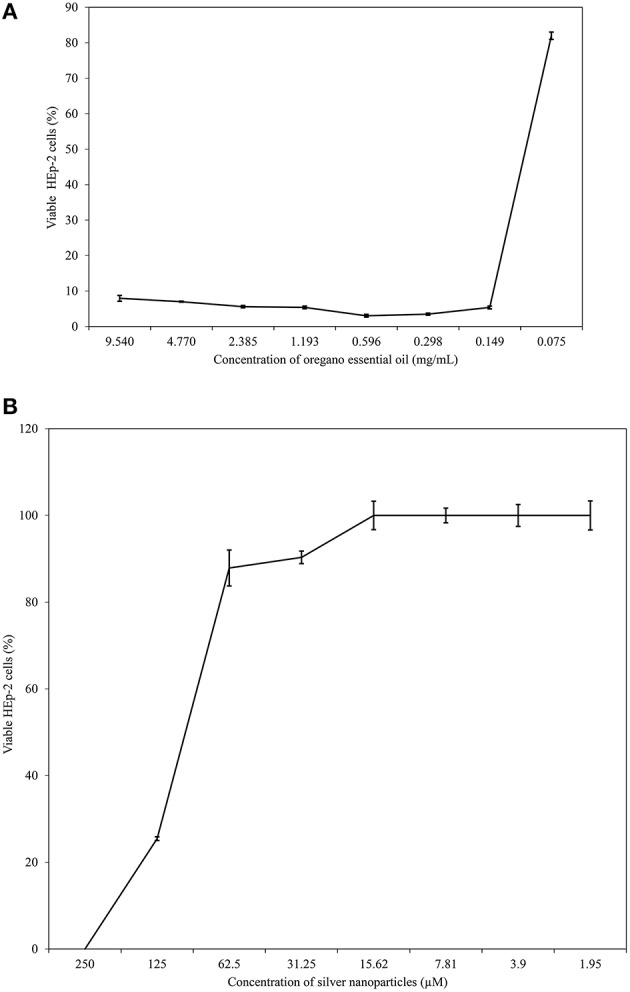
HEp-2 cell viability at different concentrations of OEO and bio-AgNP individually. (A) HEp-2 cell exposed to OEO alone ranging from 0.075 to 9.540 mg/mL. (B) HEp-2 cell exposed to bio-AgNP alone ranging from 1.95 to 250 μM. Percentage values of cell viability are the mean ± standard deviation.
Figure 6.
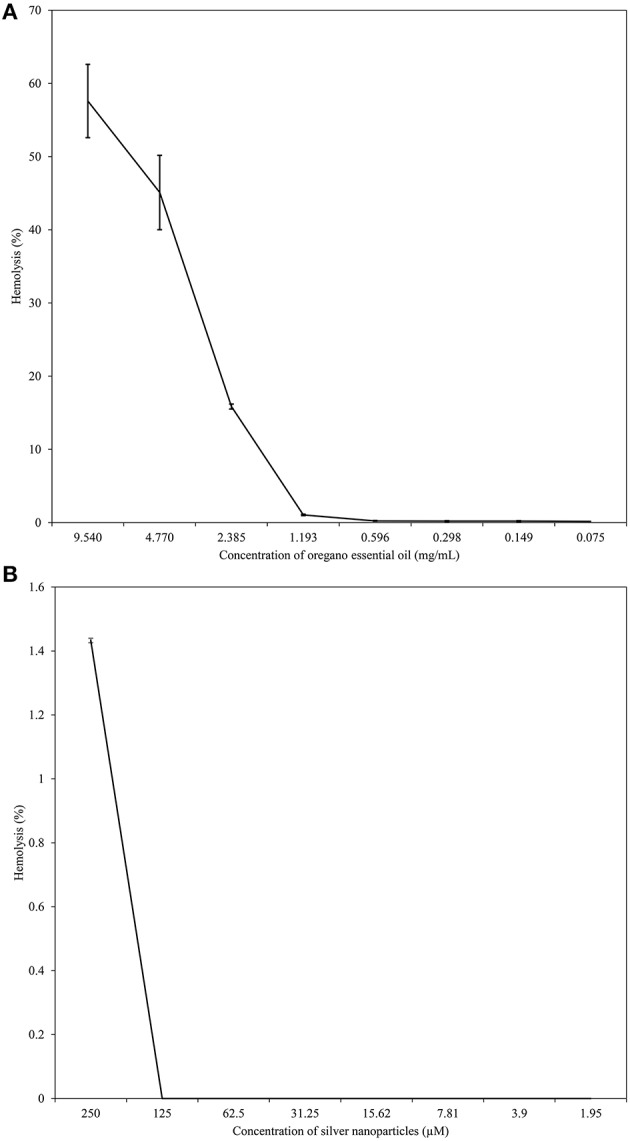
Hemolytic activity of different concentrations of OEO and bio-AgNP individually. (A) Human erythrocytes exposed to OEO alone ranging from 0.075 to 9.540 mg/mL. (B) Human erythrocytes exposed to bio-AgNP ranging from 1.95 to 250 μM. Values of hemolysis percentage are the mean ± standard deviation.
The compounds in combination were non-toxic to HEp-2 cells at concentrations of 0.075 mg/mL and 31.25 μM to OEO and bio-AgNP respectively, or at lower values concentrations. The combination of 0.075 mg/mL of OEO and 125 μM or 62.5 μM of bio-AgNP showed toxicity to HEp-2 cells (Table 4). However, the combination of OEO and bio-AgNP was non-toxic to RBC at all concentrations tested (Table 5).
Table 4.
HEp-2 cell viability at different concentrations of oregano essential oil (OEO) and biological silver nanoparticles (bio-AgNP) in combination.
| Combination | Tested concentrations | Viable cells (%) | |
|---|---|---|---|
| OEO (mg/mL) | bio-AgNP (μM) | ||
| A1 | 0.596 | 125.0 | 0.95 ± 0.69 |
| A2 | 0.596 | 62.50 | 1.36 ± 0.59 |
| A3 | 0.596 | 31.25 | 0.99 ± 0.15 |
| A4 | 0.596 | 15.62 | 1.27 ± 0.40 |
| B1 | 0.298 | 125.0 | 1.18 ± 0.20 |
| B2 | 0.298 | 62.50 | 1.72 ± 0.57 |
| B3 | 0.298 | 31.25 | 1.04 ± 0.20 |
| B4 | 0.298 | 15.62 | 1.99 ± 4.64 |
| C1 | 0.149 | 125.0 | 7.80 ± 2.00 |
| C2 | 0.149 | 62.50 | 21.81 ± 12.19 |
| C3 | 0.149 | 31.25 | 22.94 ± 14.00 |
| C4 | 0.149 | 15.62 | 72.01 ± 17.60NC |
| D1 | 0.075 | 125.0 | 38.68 ± 8.49 |
| D2 | 0.075 | 62.50 | 51.61 ± 3.25 |
| D3 | 0.075 | 31.25 | 94.24 ± 3.67 NC |
| D4 | 0.075 | 15.62 | 100 ± 0.00NC |
| E1 | 0.037 | 125.0 | 61.54 ± 6.92NC |
| E2 | 0.037 | 62.50 | 82.27 ± 10.55NC |
| E3 | 0.037 | 31.25 | 100 ± 0.00NC |
| E4 | 0.037 | 15.62 | 100 ± 0.00NC |
Concentrations of OEO (mg/mL) in combination: A, 0.596; B, 0.298; C, 0.149; D, 0.075; E, 0.037.
Concentrations of bio-AgNP (μM) in combination: (1) 125; (2) 62.5; (3) 31.25; (4) 15.62.
, non-cytotoxic, ±, standard deviation.
Table 5.
Hemolytic activity of oregano essential oil (OEO) and biological silver nanoparticles (bio-AgNP) in combination.
| Combination | Tested concentrations | Hemolysis (%) | |
|---|---|---|---|
| OEO (mg/mL) | bio-AgNP (μM) | ||
| A1 | 0.596 | 125.0 | 6.49 ± 2.29NC |
| A2 | 0.596 | 62.50 | 2.86 ± 0.11NC |
| A3 | 0.596 | 31.25 | 1.95 ± 0.11NC |
| A4 | 0.596 | 15.62 | 0.49 ± 0.13NC |
| B1 | 0.298 | 125.0 | 7.56 ± 1.16NC |
| B2 | 0.298 | 62.50 | 2.60 ± 1.33NC |
| B3 | 0.298 | 31.25 | 0.77 ± 0.37NC |
| B4 | 0.298 | 15.62 | 0.19 ± 0.32NC |
| C1 | 0.149 | 125.0 | 8.44 ± 0.98NC |
| C2 | 0.149 | 62.50 | 4.58 ± 0.23NC |
| C3 | 0.149 | 31.25 | 0.19 ± 0.17NC |
| C4 | 0.149 | 15.62 | 1.26 ± 0.19NC |
| D1 | 0.075 | 125.0 | 7.75 ± 0.93NC |
| D2 | 0.075 | 62.50 | 2.67 ± 0.73NC |
| D3 | 0.075 | 31.25 | 0.42 ± 0.13NC |
| D4 | 0.075 | 15.62 | 0.00 ± 0.00NC |
| E1 | 0.037 | 125.0 | 5.81 ± 2.27NC |
| E2 | 0.037 | 62.50 | 1.57 ± 0.27NC |
| E3 | 0.037 | 31.25 | 0.42 ± 0.53NC |
| E4 | 0.037 | 15.62 | 0.00 ± 0.00NC |
Concentrations of OEO (mg/mL) in combination: A, 0.596; B, 0.298; C, 0.149; D, 0.075; E, 0.037.
Concentrations of bio-AgNP (μM) in combination: (1) 125; (2) 62.5; (3) 31.25; (4) 15.62.
, non-cytotoxic, ±, standard deviation.
Discussion
This study showed that OEO has potent bactericidal activity at low concentration with fast action, in agreement with previous studies (Burt and Reinders, 2003; Souza et al., 2010; Betancourt et al., 2012; Alvarez et al., 2014). Busatta et al. (2007) reported for OEO MIC values of 0.23 and 0.46 mg/mL against methicillin-sensitive S. aureus and E. coli ATCC 25922 respectively. Betancourt et al. (2012) found for OEO a MIC of 0.780 mg/mL against S. enterica and 3.125 mg/mL against E. coli. OEO MIC values obtained in our study ranged from 0.298 to 1.193 mg/mL (Table 2), which were in line with literature results. Our analysis showed that OEO at a low concentration was bactericidal against all multidrug-resistant bacteria tested, with MBC values of 0.298 mg/mL against A. baumannii, 0.596 mg/mL against ESBL and KPC-producing E. coli isolates and 1.193 mg/mL against both clinical MRSA isolates.
Time-kill assays showed that OEO reduced cell populations nearly 5 log (after 10 min of treatment) in all three bacterial strains tested (p < 0.05), so our results indicated that OEO acted within a few minutes against S. aureus ATCC 25923 (Figure 1), E. coli ATCC 25922 (Figure 2), and KPC-producing E. coli (Figure 3). Burt and Reinders (2003) showed that when E. coli O157:H7 at 5 × 105 CFU/mL were exposed to OEO (0.06%, v/v), there were no viable cells after 1 min of treatment. In a study with S. aureus isolated from food, Barros et al. (2009) found that OEO at MIC (0.06%, v/v) caused 3 log reduction of initial inoculum (108 CFU/mL) after 2 h of treatment. In our study using an equal initial inoculum (S. aureus at 108 CFU/mL) and same OEO concentration (0.596 mg/mL; corresponding to 0.06 %, v/v), there were no viable cells after 30 min (p < 0.05), due to a 8 log reduction of S. aureus ATCC 25923 population (data not shown).
Some studies have found that terpenoid compounds from OEO are more active against Gram-positive bacteria (Smith-Palmer et al., 1998; Lambert et al., 2001; Stojković et al., 2013). Our results indicated that OEO has broad-spectrum action in agreement with previous studies (Busatta et al., 2007; Rosato et al., 2010; Stojković et al., 2013; Alvarez et al., 2014), where it was equally effective according to MIC (Table 2) and time of action (Figures 1–3) against Gram-positive and Gram-negative bacteria (p > 0.05); these results are in line with those obtained by other researchers (Dorman and Deans, 2000; Rosato et al., 2010). This variable susceptibility of bacteria to essential oils can be due to the chemical composition of essential oils, which varies according to seasonal and geographical factors (Medini et al., 2009).
Many studies suggest that essential oils affect the bacterial cell membrane resulting in growth inhibition, since this structure supports the most essential functions in prokaryotes. Bennis et al. (2004) reported cell surface changes such as curling and cracks in Saccharomyces cerevisiae treated with thymol. Lambert et al. (2001) suggested that OEO inhibits bacterial growth due to damage to membrane integrity, affecting pH homeostasis and equilibrium of inorganic ions. Suzuki et al. (2015) found physical damage and morphological alterations in Staphylococcus epidermidis treated with OEO products. Souza et al. (2010) showed loss of 260 nm-absorbing material and release of potassium ions of S. aureus cells treated with OEO, results suggesting increased membrane permeability. In our study, SEM observations confirmed physical damage and considerable morphological alteration as cell surface blebbing in S. aureus ATCC 25923 (non-MRSA) cells exposed to OEO for 30 min (Figure 4C), compared to control (untreated bacteria) showing no morphological changes after 30 min (Figure 4A). This analysis by electron microscopy revealed a decrease in cell density of OEO-treated bacteria sample when compared to control sample, according with time kill results at 30 min of incubation (data not shown).
In our RBC toxicity assay, OEO CC50 was 7.519 mg/mL (Figure 6A). OEO did not appear to be toxic to RBC, since MIC values ranged from 0.037 mg/mL to 1.193 mg/mL (Tables 2, 3). The lack of cytotoxicity of OEO to erythrocytes has also been found by other investigators. Mancini et al. (2014) reported no hemolytic activity of OEO against bovine erythrocytes, and Cacciatore et al. (2015) showed that carvacrol derivatives did not produce human blood hemolysis at their MIC values. OEO was toxic to HEp-2 cells (Figure 5A), and there are previous studies using tumor cells, including HEp-2 cells, that reported the antitumor activity of OEO or its components (Mehdi et al., 2011; Gautam et al., 2014; Sobral et al., 2014).
Due to its strong antimicrobial property, OEO has potential to replace antibiotics in the food industry and can be incorporated in cosmetic products, among other applications (Alvarez et al., 2014; Suzuki et al., 2015). However, the high volatile character and undesirable organoleptic features can limit its use; therefore, alternatives to solve these limitations are required (Burt, 2004; Alvarez et al., 2014; Hernández-Hernández et al., 2014).
In this study, bio-AgNP exhibited broad-spectrum antibacterial action, inhibiting growth of both Gram-positive and -negative bacteria, in agreement with the literature (Busatta et al., 2007; Jain et al., 2009; Kim et al., 2011). The mean MIC value obtained in our study (129 μM) was according with previous studies using silver nanoparticles produced by F. oxysporum that reported MIC value of 125 μM (Cardozo et al., 2013; Biasi-Garbin et al., 2015). However, we found that Gram-positive bacteria were more tolerant to silver nanoparticles because bio-AgNP MIC values were higher (Table 2) and time of action was slower, compared to Gram-negative bacteria (Figures 1–3), as reported in other studies (Jain et al., 2009; Durán et al., 2010; Agnihotri et al., 2014). Kim et al. (2011) showed that Gram-negative bacteria were more susceptible to silver nanoparticles compared to Gram-positive strains, where there was increased bactericidal activity in time kill assay, loss of protein through the membrane and inactivation of lactate dehydrogenase (LDH), besides marked morphological changes.
The size of the nanoparticles is a key point in their antibacterial activity, where many studies show that antibacterial activity is particle size dependent; antimicrobial activity of nanoparticles increases with smaller size (Panacek et al., 2006; Ayala-Núñez et al., 2009; Agnihotri et al., 2014), and this fact accounts for the variation in MIC data between different studies, limiting comparison.
Our analysis by SEM showed surface protrusions on bio-AgNP treated S. aureus cells (non-MRSA) (Figure 4D). Kim et al. (2011) reported that bacterial cells treated with silver nanoparticles had irregular fragments on the surface, a large protein leakage and decrease in bacterial growth; suggesting that fragments can be lost cytoplasmic material resulting in damage to the membrane with a consequent increase in permeability. Studies also report that silver ions have an affinity for thiol groups and may damage respiratory enzymes of cytoplasmic membrane (Holt and Bard, 2005; Li et al., 2010).
In our RBC toxicity assay, bio-AgNP were not cytotoxic to human blood cells (Figure 6B). Our metal compound did not exhibit cytotoxic effect on RBC at the concentrations studied, 250 μM (37.5 μg/mL) to 1.95 μM (0.29 μg/mL), so it was not possible to calculate CC50 for human erythrocytes. Spectrophotometric analysis of supernatants indicated that bio-AgNP did not increase absorbance, demonstrating that little hemoglobin was released from the RBC. Choi et al. (2011) showed that the silver nanoparticles at 700 μg/mL lysed 50% of RBC. According to ASTM E2524-08 (2013)1 standards, hemolysis > 5% indicates damage to RBC; in our study, the highest bio-AgNP concentration tested (250 μM) showed only 1.4% hemolysis. Our MTT assay results showed that bio-AgNP was toxic to HEp-2 cells only at high concentrations (CC50 = 97.22 μM), often above that needed to inhibit bacterial growth. Some authors have reported antitumor activity of silver nanoparticles, supporting our finding of bio-AgNP being more toxic to HEp-2 cells than to RBC (Devi and Bhimba, 2012; Antony et al., 2013).
Despite the effective antimicrobial action of bio-AgNP with broad spectrum activity and low cytotoxicity, bacteria can easily develop resistance to these nanoparticles by simple genetic changes (Graves et al., 2015). Thus, alternative studies to work around this problem are needed.
Combined drug treatment is recommended as a strategy to control antimicrobial resistance (Fischbach, 2011; Bollenbach, 2015). Li et al. (2005) observed a synergistic effect of silver nanoparticles combined with amoxicillin against E. coli, and other studies have shown a synergistic interaction with other natural alternative compounds such as cinnamaldehyde (Ghosh et al., 2013), eugenol (Biasi-Garbin et al., 2015), and phenazine-1-carboxamide (Cardozo et al., 2013).
Other studies have shown that the OEO acts in synergism with some conventional antimicrobials such as gentamicin against S. aureus 29213, E. coli 25922, and A. baumannii 19606 (Rosato et al., 2010), or has an additive effect in combination with amoxicillin, polymyxin, and lincomycin against ESBL-producing E. coli (Si et al., 2008). OEO shows synergistic effects with natural substances such as essential oil of Thymus vulgare and Rosmarinus officinalis (Stojković et al., 2013; Honório et al., 2015).
In the literature, OEO (Nostro et al., 2004; Si et al., 2008; Amrouni et al., 2014) and silver nanoparticles (Cardozo et al., 2013; Singh et al., 2014; Subashini et al., 2014) were shown to be active against multidrug-resistant bacteria. But this study showed, for the first time, synergistic and additive effects of a combination of OEO and bio-AgNP against bacterial strains, including MRSA and beta-lactamase- or carbapenemase-producing bacteria. Synergistic and additive drug interactions may be potential strategies for controlling resistance evolution, since administration of multiple drugs may disrupt several bacterial functions and thus minimize selection of resistant strains (Yeh et al., 2009; Bollenbach, 2015). In combination, both compounds were effective at concentrations that individually do not affect human erythrocyte (Figure 5), and their synergistic or additive effects neither impact such cells (Table 5). However, OEO combined with bio-AgNP was cytotoxic to HEp-2 (Table 4); as observed for these compounds individually. The combination also showed antibacterial activity in short time (Figures 1–3). Furthermore, the undesirable organoleptic characteristics of OEO are decreased, since the synergistic or additive interaction reduces the concentration of both compounds.
Our results highlight the powerful action of OEO combined with bio-AgNP against Gram-negative bacteria, including those producing ESBL and carbapenemases (Table 3, Figures 2, 3). The combination of the two substances also showed activity against Gram-positive bacteria, since the FICI indicated synergistic and additive interactions (Table 3), and SEM confirmed this antimicrobial action through changes in the S. aureus (non-MRSA) cell surface and decrease in cell density (Figure 4E) compared to control (Figure 4B).
In conclusion, the combination of bio-AgNP with OEO resulted in synergistic and additive antimicrobial activities against the multidrug-resistant bacterial strains of E. coli, A. baumannii and MRSA. Therefore bio-AgNP combined with OEO has potential to be applied in industry (cosmetics, food and pharmaceutical, for example) and clinical and hospital settings (i.e., for treating wounds and burns infections, or for disinfecting hospital to combat resistant strains as A. baumannii).
Author contributions
SS, conception and drafting of the work, analysis and interpretation of results, writing of this manuscript. LD, conception of the work, analysis of the results (especially with essential oil assays and time-kill curves), review critically of this manuscript. CC, conception and analysis of MTT assay. SY, conception and analysis of MTT assay. CN, design of SEM methodology, image acquisition, analysis and interpretation of SEM results. AD, design of SEM methodology, image acquisition, analysis and interpretation of SEM results. CA, design of SEM methodology, image acquisition, analysis, and interpretation of SEM results. ND, biosynthesis and characterization of silver nanoparticles. GN, analysis and interpretation of results, review critically of this manuscript, final approval of the manuscript version to be published. RK, design of the work, drafting the work, analysis and interpretation of results, guidance in all stages of the work, review critically, and correction of this manuscript, final approval of the manuscript version to be published.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This study was supported by CNPq BIOTEC 402728/2013-0 and Postgraduate Program in Microbiology of Universidade Estadual de Londrina. SS was funded by a student scholarship from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). The authors thank Dr. Carlos Nozawa for providing the HEp-2 cells. Dr. A. Leyva helped with English editing of the manuscript. Laboratory of Electron Microscopy and Microanalysis of Universidade Estadual de Londrina for performing the electron micrographs.
Glossary
Abbreviations
- OEO
oregano essential. oil
- bio-AgNP
biological synthesized silver nanoparticle
- RBC
red blood cells.
Footnotes
1ASTM E2524-08 (2013). Standard Test Method for Analysis of Hemolytic Properties of Nanoparticles. Available online at: http://www.astm.org/Standards/E2524.htm [Accessed October 17, 2015].
Supplementary material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.00760
References
- Agnihotri S., Mukherji S., Mukherji S. (2014). Size-controlled silver nanoparticles synthesized over the range 5–100 nm using the same protocol and their antibacterial efficacy. RSC Adv. 4, 3974–3983. 10.1039/C3RA44507K [DOI] [Google Scholar]
- Alexander J. W. (2009). History of the medical use of silver. Surg. Infect. 10, 289–292. 10.1089/sur.2008.9941 [DOI] [PubMed] [Google Scholar]
- Alvarez M. V., Ortega-Ramirez L. A., Gutierrez-Pacheco M. M., Bernal-Mercado A. T., Rodriguez-Garcia I., Gonzalez-Aguilar G. A., et al. (2014). Oregano essential oil-pectin edible films as anti-quorum sensing and food antimicrobial agents. Front. Microbiol. 5:699. 10.3389/fmicb.2014.00699 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amrouni S., Touati M., Hadef Y., Djahoudi A. (2014). Effet de l'huile essentielle d'Origanum vulgare et de Thymus ciliatus sur Pseudomonas aeruginosa VIM-2 carbapénèmase. Phytothérapie 12, 309–313. 10.1007/s10298-014-0842-x [DOI] [Google Scholar]
- Ansari M. A., Khan H. M., Khan A. A., Cameotra S. S., Saquib Q., Musarrat J. (2014). Gum arabic capped-silver nanoparticles inhibit biofilm formation by multi-drug resistant strains of Pseudomonas aeruginosa. J. Basic Microbiol. 54, 688–699. 10.1002/jobm.201300748 [DOI] [PubMed] [Google Scholar]
- Antony J. J., Sithika M. A. A., Joseph T. A., Suriyakalaa U., Sankarganesh A., Siva D., et al. (2013). In vivo antitumor activity of biosynthesized silver nanoparticles using Ficus religiosa as a nanofactory in DAL induced mice model. Colloids Sur. B Biointerfaces 108, 185–190. 10.1016/j.colsurfb.2013.02.041 [DOI] [PubMed] [Google Scholar]
- Ayala-Núñez N. V., Villegas H. H. L., Turrent L. C. I., Padilla C. R. (2009). Silver nanoparticles toxicity and bactericidal effect against methicillin-resistant Staphylococcus aureus: nanoscale does matter. Nanobiotechnology 5, 2–9. 10.1007/s12030-009-9029-1 [DOI] [Google Scholar]
- Barros J. C., Conceição M. L., Gomes Neto N. J., Costa A. C. V., Siqueira Júnior J. P., Basílio Júnior I. D. (2009). Interference of Origanum vulgare L. essential oil on the growth and some physiological characteristics of Staphylococcus aureus strains isolated from foods. LWT Food Sci. Tech. 42, 1139–1143. 10.1016/j.lwt.2009.01.010 [DOI] [Google Scholar]
- Bass S. N., Bauer S. R., Neuner E. A., Lam S. W. (2015). Mortality risk factors for critically III patients with carbapenem-resistant bacteremia: impact of combination antimicrobial therapy. Antimicrob. Agents Chemother. 59, 3748–3753. 10.1128/AAC.00091-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassolé I. H. N., Juliani H. R. (2012). Essential oils in combination and their antimicrobial properties. Molecules 17, 3989–4006. 10.3390/molecules17043989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennis S., Chami F., Chami N., Bouchikhi T., Remmal A. (2004). Surface alteration of Saccharomyces cerevisiae induced by thymol and eugenol. Lett. Appl. Microbiol. 38, 454–458. 10.1111/j.1472-765X.2004.01511.x [DOI] [PubMed] [Google Scholar]
- Betancourt L., Phandanauvong V., Patiño R. (2012). Composition and bactericidal activity against beneficial and pathogenic bacteria of oregano essential oils from four chemotypes of Origanum and Lippia genus. Arq. Bras. de Med. Veterinária e Zootecnia 59, 21–31. [Google Scholar]
- Biasi-Garbin R. P., Otaguiri E. S., Morey A. T., da Silva M. F., Morguette A. E. B., Lancheros C. A. C., et al. (2015). Effect of eugenol against Streptococcus agalactiae and synergistic interaction with biologically produced silver nanoparticles. Evid. Based Complement. Altern. Med. 2015:861497. 10.1155/2015/861497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bibbs R. K., Harris R. D., Peoples V. A., Barnett C., Singh S. R., Dennis V. A., et al. (2014). Silver polyvinyl pyrrolidone nanoparticles exhibit a capsular polysaccharide influenced bactericidal effect against Streptococcus pneumoniae. Front. Microbiol. 5:665. 10.3389/fmicb.2014.00665 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bollenbach T. (2015). Antimicrobial interactions: mechanisms and implications for drug discovery and resistance evolution. Curr. Opin. Microbiol. 27, 1–9. 10.1016/j.mib.2015.05.008 [DOI] [PubMed] [Google Scholar]
- Burt S. (2004). Essential oils: their antibacterial properties and potential applications in foods - a review. Int. J. Food Microbiol. 94, 223–253. 10.1016/j.ijfoodmicro.2004.03.022 [DOI] [PubMed] [Google Scholar]
- Burt S. A., Reinders R. D. (2003). Antibacterial activity of selected plant essential oils against Escherichia coli O157:H7. Lett. Appl. Microbiol. 36, 162–167. 10.1046/j.1472-765X.2003.01285.x [DOI] [PubMed] [Google Scholar]
- Busatta C., Mossi A. J., Rodrigues M. R. A., Cansian R. L., Oliveira J. V. (2007). Evaluation of Origanum vulgare essential oil as antimicrobial agent in sausage. Braz. J. Microbiol. 38, 610–616. 10.1590/S1517-83822007000400006 [DOI] [Google Scholar]
- Bush K. (2010). Bench-to-bedside review: the role of beta-lactamases in antibiotic-resistant Gram-negative infections. Critical Care. 14:224. 10.1186/cc8892 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cacciatore I., Di Giulio M., Fornasari E., Di Stefano A., Cerasa L. S., Marinelli L., et al. (2015). Carvacrol Codrugs: a new approach in the antimicrobial plan. PLoS One 10:e0120937. 10.1371/journal.pone.0120937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cantas L., Shah S. Q. A., Cavaco L. M., Manaia C. M., Walsh F., Popowska M., et al. (2013). A brief multi-disciplinary review on antimicrobial resistance in medicine and its linkage to the global environmental microbiota. Front. Microbiol. 4:96. 10.3389/fmicb.2013.00096 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardozo V. F., Oliveira A. G., Nishio E. K., Perugini M. R. E., Andrade C. G. T. J., Silveira W. D., et al. (2013). Antibacterial activity of extracellular compounds produced by a Pseudomonas strain against methicillin-resistant Staphylococcus aureus (MRSA) strains. Ann. Clin. Microbiol. Antimicrob. 12:12. 10.1186/1476-0711-12-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cervato G., Carabelli M., Gervasio S., Cittera A., Cazzola R., Cestaro B. (2000). Antioxidant properties of (Oregano vulgare) leaf extracts. J. Food Biochem. 24, 453–465. 10.1111/j.1745-4514.2000.tb00715.x [DOI] [Google Scholar]
- Chin N. X., Weitzman I., Della-Latta P. (1997). In vitro activity of fluvastatin, a cholesterol-lowering agent, and synergy with flucanazole and itraconazole against candida species and Cryptococcus neoformans. Antimicrob. agents chemother. 41, 850–852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi J., Reipa V., Hitchins V. M., Goering P. L., Malinauskas R. A. (2011). Physicochemical characterization and in vitro hemolysis evaluation of silver nanoparticles. Toxicol. Sci. 123, 133–143. 10.1093/toxsci/kfr149 [DOI] [PubMed] [Google Scholar]
- Cleff M. B., Meinerz A. R., Sallis E. S., Antunes T. A., Mattei A., Rodrigues M. R., et al. (2008). Toxicidade pré-clínica em doses repetidas do óleo essencial do Origanum vulgare L. (Orégano) em ratas Wistar. Latin Am. J. Pharm. 27, 704–709. [Google Scholar]
- CLSI (2012). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically. Available online at: http://antimicrobianos.com.ar/ATB/wp-content/uploads/2012/11/03-CLSI-M07-A9-2012.pdf [Accessed October 17, 2015].
- de Lima R., Seabra A. B., Durán N. (2012). Silver nanoparticles: a brief review of cytotoxicity and genotoxicity of chemically and biogenically synthesized nanoparticles. J. Appl. Toxicol. 32, 867–879. 10.1002/jat.2780 [DOI] [PubMed] [Google Scholar]
- de Souza E. L., Barros J. C., Conceição M. L., Gomes Neto N. J., Costa A. C. V. (2009). Combined application of Origanum vulgare L. essential oil and acetic acid for controlling the growth of Staphylococcus aureus in foods. Braz. J. Microbiol. 40, 387–393. 10.1590/S1517-83822009000200032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devi J. S., Bhimba B. V. (2012). Anticancer activity of silver nanoparticles synthesized by the seaweed Ulva lactuca in vitro. J. Nanomedine. Biotherapeutic Discov. 01:242 10.4172/scientificreports.242 [DOI] [Google Scholar]
- Dhillon R. H. P., Clark J. (2012). ESBLs: a clear and present danger? Crit. Care Res. Pract. 2012:625170. 10.1155/2012/625170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dibrov P., Dzioba J., Gosink K. K., Häse C. C. (2002). Chemiosmotic mechanism of antimicrobial activity of Ag+ in Vibrio cholerae. Antimicrob. Agents Chemother. 46, 8–11. 10.1128/aac.46.8.2668-2670.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dorman H. J., Deans S. G. (2000). Antimicrobial agents from plants: antibacterial activity of plant volatile oils. J. Appl. Microbiol. 88, 308–316. 10.1046/j.1365-2672.2000.00969.x [DOI] [PubMed] [Google Scholar]
- Drawz S. M., Bonomo R. A. (2010). Three decades of β-lactamase inhibitors. Clin. Microbiol. Rev. 23, 160–201. 10.1128/CMR.00037-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du W. X., Olsen C. W., Avena-Bustillos R. J., McHugh T. H., Levin C. E., Mandrell R., et al. (2009). Antibacterial effects of allspice, garlic, and oregano essential oils in tomato films determined by overlay and vapor-phase methods. J. Food Sci. 74, 390–397. 10.1111/j.1750-3841.2009.01289.x [DOI] [PubMed] [Google Scholar]
- Durán N., Alves O. L., Esposito E., de Souza G. I., Marcato P. D. (2006). Processo de produção de nanopartículas de prata estabilizadas por proteínas na produção de produtos têxteis antibacterianos e o tratamento dos efluentes produzidos. Braz. patent PI 0605681-4 A. Campinas: BR. Instituto Nacional da Propriedade Industrial (INPI). [Google Scholar]
- Durán N., Marcato P. D., Alves O. L., Souza G. I. H., Esposito E. (2005). Mechanistic aspects of biosynthesis of silver nanoparticles by several Fusarium oxysporum strains. J. Nanobiotechnology. 3:8. 10.1186/1477-3155-3-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durán N., Marcato P. D., De Conti R., Alves O. L., Costa F. T. M., Brocchi M. (2010). Potential use of silver nanoparticles on pathogenic bacteria, their toxicity and possible mechanisms of action. J. Braz. Chem. Soc. 21, 949–959. 10.1590/S0103-50532010000600002 [DOI] [Google Scholar]
- Durán N., Marcato P. D., Durán M., Yadav A., Gade A., Rai M. (2011). Mechanistic aspects in the biogenic synthesis of extracellular metal nanoparticles by peptides, bacteria, fungi, and plants. Appl. Microbiol. Biotechnol. 90, 1609–1624. 10.1007/s00253-011-3249-8 [DOI] [PubMed] [Google Scholar]
- Fayaz A. M., Balaji K., Girilal M., Yadav R., Kalaichelvan P. T., Venketesan R. (2010). Biogenic synthesis of silver nanoparticles and their synergistic effect with antibiotics: a study against Gram-positive and Gram-negative bacteria. Nanomed. Nanotechnol. Biol. Med. 6, 103–109. 10.1016/j.nano.2009.04.006 [DOI] [PubMed] [Google Scholar]
- Feng Q., Wu J., Chen G. Q., Cui F. Z., Kim T. N., Kim J. O. (2000). A mechanistic study of the antibacterial effect of silver ions on Escherichia coli and Staphylococcus aureus. J. Biomed. Mater. Res. 52, 662–668. [DOI] [PubMed] [Google Scholar]
- Fischbach M. A. (2011). Combination therapies for combating antimicrobial resistance. Curr. Opin. Microbiol. 14, 519–523. 10.1016/j.mib.2011.08.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fontes L. C., Neves P. R., Oliveira S., Silva K. C., Hachich E. M., Sato M. I. Z., et al. (2011). Isolation of Pseudomonas aeruginosa coproducing metallo-β-lactamase SPM-1 and 16S rRNA methylase RmtD1 in an urban river. Antimicrob. Agents Chemother. 55, 3063–3064. 10.1128/AAC.00138-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gautam N., Mantha A. K., Mittal S. (2014). Essential oils and their constituents as anticancer agents: a mechanistic view. Biomed. Res. Int. 2014:154106. 10.1155/2014/154106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghosh I. N., Patil S. D., Sharma T. K., Srivastava S. K., Pathania R., Navani N. K. (2013). Synergistic action of cinnamaldehyde with silver nanoparticles against spore-forming bacteria: a case for judicious use of silver nanoparticles for antibacterial applications. Int. J. Nanomed. 8, 4721–4731. 10.2147/IJN.S49649 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilling D. H., Kitajima M., Torrey J. R., Bright K. R. (2014). Antiviral efficacy and mechanisms of action of oregano essential oil and its primary component carvacrol against murine norovirus. J. Appl. Microbiol. 116, 1149–1163. 10.1111/jam.12453 [DOI] [PubMed] [Google Scholar]
- Graves J. L., Jr., Tajkarimi M., Cunningham Q., Campbell A., Nonga H., Harrison S. H., et al. (2015). Rapid evolution of silver nanoparticle resistance in Escherichia coli. Front. Genet. 6:42. 10.3389/fgene.2015.00042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helander I. M., Alakomi H.-L., Latva-Kala K., Mattila-Sandholm T., Pol I., Smid E. J., et al. (1998). Characterization of the action of selected essential oil components on Gram-negative bacteria. J. Agric. Food Chem. 46, 3590–3595. 10.1021/jf980154m [DOI] [Google Scholar]
- Herman A., Herman A. P. (2014). Nanoparticles as antimicrobial agents: their toxicity and mechanisms of action. J. Nanosci. Nanotechnol. 14, 946–957. 10.1166/jnn.2014.9054 [DOI] [PubMed] [Google Scholar]
- Hernández-Hernández E., Regalado-González C., Vázquez-Landaverde P., Guerrero-Legarreta I., García-Almendárez B. E. (2014). Microencapsulation, chemical characterization, and antimicrobial activity of mexican (Lippia graveolens H. B. K.) and european (Origanum vulgare L.) oregano essential oils. Sci. World J. 2014:641814. 10.1155/2014/641814 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holt K., Bard A. (2005). The interaction of silver (I) ions with the respiratory chain of Escherichia coli: an electrochemical and scanning electrochemical microscopy study of the antimicrobial mechanism of micromolar Ag+. Biochemistry 44, 13214–13223. 10.1021/bi0508542 [DOI] [PubMed] [Google Scholar]
- Honório V. G., Bezerra J., Souza G. T., Carvalho R. J. (2015). Inhibition of Staphylococcus aureus cocktail using the synergies of oregano and rosemary essential oils or carvacrol and 1,8-cineole. Front. Microbiol. 6:1223. 10.3389/fmicb.2015.01223 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyldgaard M., Mygind T., Meyer R. L. (2012). Essential oils in food preservation: mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 3:12. 10.3389/fmicb.2012.00012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Izumi E., Ueda-Nakamura T., Veiga V. F., Jr., Pinto A. C., Nakamura C. V. (2012). Terpenes from copaifera demonstrated in vitro antiparasitic and synergic activity. J. Med. Chem. 55, 2994–3001. 10.1021/jm201451h [DOI] [PubMed] [Google Scholar]
- Jain J., Arora S., Rajwade J. M., Omray P., Khandelwal S., Paknikar K. M. (2009). Silver nanoparticles in therapeutics: development of an antimicrobial gel formulation for topical use. Mol. Pharm. 6, 1388–1401. 10.1021/mp900056g [DOI] [PubMed] [Google Scholar]
- Kalemba D., Kunicka A. (2003). Antibacterial and antifungal properties of essential oils. Curr. Med. Chem. 10, 813–829. 10.2174/0929867033457719 [DOI] [PubMed] [Google Scholar]
- Kim S. H., Lee H. S., Ryu D. S., Choi S. J., Lee D. S. (2011). Antibacterial activity of silver-nanoparticles against Staphylococcus aureus and Escherichia coli. J. Microbiol. Biotechnol. 39, 77–85. [Google Scholar]
- Koga V. L., Rodrigues G. R., Scandorieiro S., Vespero E. C., Oba A., De Brito B. G. (2015). Evaluation of the antibiotic resistance and virulence of Escherichia coli strains isolated from chicken carcasses in 2007 and 2013 from Paraná, Brazil. Foodborne Pathog. Dis. 12, 479–485. 10.1089/fpd.2014.1888 [DOI] [PubMed] [Google Scholar]
- Kong K. F., Schneper L., Mathee K. (2010). Beta-lactam antibiotics: from antibiosis to resistance and bacteriology. APMIS 118, 1–36. 10.1111/j.1600-0463.2009.02563.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambert R. J. W., Skandamis P. N., Coote P. J., Nychas G. J. E. (2001). A study of the minimum inhibitory concentration and mode of action of oregano essential oil, thymol and carvacrol. J. Appl. Microbiol. 91, 453–462. 10.1046/j.1365-2672.2001.01428.x [DOI] [PubMed] [Google Scholar]
- Li P., Li J., Wu C., Wu Q., Li J. (2005). Synergistic antibacterial effects of β-lactam antibiotic combined with silver nanoparticles. Nanotechnology 16, 1912–1917. 10.1088/0957-4484/16/9/082 [DOI] [Google Scholar]
- Li W. R., Xie X. B., Shi Q. S., Zeng H. Y., Ou-Yang Y. S., Chen Y. B. (2010). Antibacterial activity and mechanism of silver nanoparticles on Escherichia coli. Appl. Microbiol. Biotechnol. 85, 1115–1122. 10.1007/s00253-009-2159-5 [DOI] [PubMed] [Google Scholar]
- Lok C. N., Ho C. M., Chen R., He Q. Y., Yu W. Y., Sun H., et al. (2006). Proteomic analysis of the mode of antibacterial action of silver nanoparticles. J. Proteome Res. 5, 916–924. 10.1021/pr0504079 [DOI] [PubMed] [Google Scholar]
- Losasso C., Belluco S., Cibin V., Zavagnin P., Mičetić I., Gallocchio F., et al. (2014). Antibacterial activity of silver nanoparticles: sensitivity of different Salmonella serovars. Front. Microbiol. 5:227. 10.3389/fmicb.2014.00227 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mancini E., Camele I., Elshafie H. S., De Martino L., Pellegrino C., Grulova D., et al. (2014). Chemical composition and biological activity of the essential oil of Origanum vulgare ssp. hirtum from different areas in the Southern Apennines (Italy). Chem. Biodivers. 11, 639–651. 10.1002/cbdv.201300326 [DOI] [PubMed] [Google Scholar]
- Mandal M. D., Mandal S. (2011). Honey: its medicinal property and antibacterial activity. Asian Pac. J. Trop. Biomed. 1, 154–160. 10.1016/S2221-1691(11)60016-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Medini H., Elaissi A., Farhat F., Khouja M. L., Chemli R., Harzallah-Skhiri F. (2009). Seasonal and geographical influences on the chemical composition of Juniperus phoenicea L. essential oil leaves from the northern Tunisia. Chem. Biodivers. 6, 1378–1387. 10.1002/cbdv.200800202 [DOI] [PubMed] [Google Scholar]
- Mehdi S. J., Ahmad A., Irshad M., Manzoor N., Rizvi M. M. A. (2011). Cytotoxic effect of carvacrol on human cervical cancer cells. Biol. Med. 3, 307–312. [Google Scholar]
- Naqvi S. Z. H., Kiran U., Ali M. I., Jamal A., Hameed A., Ahmed S., et al. (2013). Combined efficacy of biologically synthesized silver nanoparticles and different antibiotics against multidrug-resistant bacteria. Int. J. Nanomed. 8, 3187–3195. 10.2147/IJN.S49284 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nazzaro F., Fratianni F., De Martino L., Coppola R., De Feo V. (2013). Effect of essential oils on pathogenic bacteria. Pharmaceuticals 6, 1451–1474. 10.3390/ph6121451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- NCCLS (1999). Methods for Determining Bactericidal Activity of Antimicrobial Agents. Available online at: http://shop.clsi.org/site/Sample_pdf/M26A_sample.pdf [Accessed October 17, 2015].
- Nostro A., Blanco A. R., Cannatelli M. A., Enea V., Flamini G., Morelli I., et al. (2004). Susceptibility of methicillin-resistant Staphylococci to oregano essential oil, carvacrol and thymol. FEMS Microbiol. Lett. 230, 191–195. 10.1016/S0378-1097(03)00890-5 [DOI] [PubMed] [Google Scholar]
- Nostro A., Roccaro A. S., Bisignano G., Marino A., Cannatelli M. A., Pizzimenti F. C., et al. (2007). Effects of oregano, carvacrol and thymol on Staphylococcus aureus and Staphylococcus epidermidis biofilms. J. Med. Microbiol. 56, 519–523. 10.1099/jmm.0.46804-0 [DOI] [PubMed] [Google Scholar]
- Nowack B., Krug H. F., Height M. (2011). 120 years of nanosilver history: implications for policy makers. Environ. Sci. Technol. 45, 1177–1183. 10.1021/es103316q [DOI] [PubMed] [Google Scholar]
- Palanisamy N. K., Ferina N., Amirulhusni A. N., Mohd-Zain Z., Hussaini J., Ping L. J., et al. (2014). Antibiofilm properties of chemically synthesized silver nanoparticles found against Pseudomonas aeruginosa. J. Nanobiotechnol. 12:2. 10.1186/1477-3155-12-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panacek A., Kvítek L., Prucek R., Kolar M., Vecerova R., Pizurova N., et al. (2006). Silver colloid nanoparticles : synthesis, characterization and their antibacterial activity. J. Phys. Chem. B. 110, 16248–16253. 10.1021/jp063826h [DOI] [PubMed] [Google Scholar]
- Patel G., Bonomo R. A. (2013). “Stormy waters ahead”: global emergence of carbapenemases. Front. Microbiol. 4:48. 10.3389/fmicb.2013.00048 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitout J. D. D. (2012). Extraintestinal pathogenic Escherichia coli: a combination of virulence with antibiotic resistance. Front. Microbiol. 3:9. 10.3389/fmicb.2012.00009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prado T., Pereira W. C., Silva D. M., Seki L. M., Carvalho A. P. D. A., Asensi M. D. (2007). Detection of extended-spectrum β-lactamase-producing Klebsiella pneumoniae in effluents and sludge of a hospital sewage treatment plant. Lett. Appl. Microbiol. 46, 136–141. 10.1111/j.1472-765x.2007.02275.x [DOI] [PubMed] [Google Scholar]
- Queenan A. M., Bush K. (2007). Carbapenemases: the versatile β-lactamases. Clin. Microbiol. Rev. 20, 440–458. 10.1128/CMR.00001-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rai M. K., Deshmukh S. D., Ingle A. P., Gade A. K. (2012). Silver nanoparticles: the powerful nanoweapon against multidrug-resistant bacteria. J. Appl. Microbiol. 112, 841–852. 10.1111/j.1365-2672.2012.05253.x [DOI] [PubMed] [Google Scholar]
- Rhayour K., Bouchikhi T., Tantaoui-Elaraki A., Sendide K., Remmal A. (2003). The mechanism of bactericidal action of oregano and clove essential oils and of their phenolic major components on Escherichia coli and Bacillus subtilis. J. Essent. Oil Res. 15, 286–292. 10.1080/10412905.2003.9712144 [DOI] [Google Scholar]
- Rice L. B. (2012). Mechanisms of resistance and clinical relevance of resistance to β-lactams, glycopeptides, and fluoroquinolones. Mayo Clin. Proc. 87, 198–208. 10.1016/j.mayocp.2011.12.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosato A., Piarulli M., Corbo F., Muraglia M., Carone A., Vitali M. E., et al. (2010). In vitro synergistic antibacterial action of certain combinations of gentamicin and essential oils. Curr. Med. Chem. 17, 3289–3295. 10.2174/092986710792231996 [DOI] [PubMed] [Google Scholar]
- Rosato A., Vitali C., De Laurentis N., Armenise D., Milillo M. A. (2007). Antibacterial effect of some essential oils administered alone or in combination with norfloxacin. Phytomedicine 14, 727–732. 10.1016/j.phymed.2007.01.005 [DOI] [PubMed] [Google Scholar]
- Rubin J. E., Ekanayake S., Fernando C. (2014). Carbapenemase-producing Organism in Food, 2014. Emerg. Infect. Dis. 20, 1264–1265. 10.3201/eid2007.140534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sankar R., Karthik A., Prabu A., Karthik S., Shivashangari K. S., Ravikumar V. (2013). imediated biosynthesis of silver nanoparticles for its antibacterial and anticancer activity. Colloids Surf. B Biointerfaces 108, 80–84. 10.1016/j.colsurfb.2013.02.033 [DOI] [PubMed] [Google Scholar]
- Sastry M., Ahmad A., Islam Khan M., Kumar R. (2003). Biosynthesis of metal nanoparticles using fungi and actinomycete. Curr. Sci. 85, 162–170. [Google Scholar]
- Shaikh S., Fatima J., Shakil S., Rizvi S. M. D., Kamal M. A. (2015). Antibiotic resistance and extended spectrum beta-lactamases: types, epidemiology and treatment. Saudi J. Biol. Sci. 22, 90–101. 10.1016/j.sjbs.2014.08.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Si H., Hu J., Liu Z., Zeng Z. L. (2008). Antibacterial effect of oregano essential oil alone and in combination with antibiotics against extended-spectrum β-lactamase-producing Escherichia coli. FEMS Immunol. Med. Microbiol. 53, 190–194. 10.1111/j.1574-695X.2008.00414.x [DOI] [PubMed] [Google Scholar]
- Silva K. C., Lincopan N. (2012). Epidemiologia das betalactamases de espectro estendido no Brasil : impacto clínico e implicações para o agronegócio. J. Bras. Patol. Med. Lab. 48, 91–99. 10.1590/S1676-24442012000200004 [DOI] [Google Scholar]
- Singh K., Panghal M., Kadyan S., Chaudhary U., Yadav J. (2014). Green silver nanoparticles of Phyllanthus amarus: as an antibacterial agent against multi drug resistant clinical isolates of Pseudomonas aeruginosa. J. Nanobiotechnol. 12:40. 10.1186/s12951-014-0040-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith-Palmer A., Stewart J., Fyfe L. (1998). Antimicrobial properties of plant essential oils and essences against five important food-borne pathogens. Lett. Appl. Microbiol. 26, 118–122. 10.1046/j.1472-765X.1998.00303.x [DOI] [PubMed] [Google Scholar]
- Sobral M. V., Xavier A. L., Lima T. C., de Sousa D. P. (2014). Antitumor activity of monoterpenes found in essential oils. Sci. World J. 2014:953451. 10.1155/2014/953451 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Souza E. L., Barros J. C., Oliveira C. E. V., Conceição M. L. (2010). Influence of Origanum vulgare L. essential oil on enterotoxin production, membrane permeability and surface characteristics of Staphylococcus aureus. Int. J. Food Microbiol. 137, 308–311. 10.1016/j.ijfoodmicro.2009.11.025 [DOI] [PubMed] [Google Scholar]
- Souza E. L., Oliveira C. E. V., Stamford T. L. M., Conceição M. L., Gomes Neto N. J. (2013). Influence of carvacrol and thymol on the physiological attributes, enterotoxin production and surface characteristics of Staphylococcus aureus strains isolated from foods. Braz. J. Microbiol. 44, 29–35. 10.1590/S1517-83822013005000001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stojković D., Glamočlija J., Ćirić A., Nikolić M., Ristić M., Šiljegović J. (2013). Investigation on antibacterial synergism of Origanum vulgare and Thymus vulgaris essential oils. Arch. Biol. Sci. 65, 639–644. 10.2298/ABS1302639S [DOI] [Google Scholar]
- Subashini J., Khanna V. G., Kannabiran K. (2014). Anti-ESBL activity of silver nanoparticles biosynthesized using soil Streptomyces species. Bioprocess Biosyst. Eng. 37, 999–1006. 10.1007/s00449-013-1070-8 [DOI] [PubMed] [Google Scholar]
- Suzuki É. Y., Soldati P. P., Chaves M. G. A. M., Nádia R. B. (2015). Essential oil from Origanum vulgare Linnaeus: an alternative against microorganisms responsible for bad perspiration odor. J. Young Pharm. 7, 12–20. 10.5530/jyp.2015.1.4 [DOI] [Google Scholar]
- Szabó M. Á., Varga G. Z., Hohmann J., Schelz Z., Szegedi E., Amaral L., et al. (2010). Inhibition of quorum-sensing signals by essential oils. Phytother. Res. 24, 782–786. 10.1002/ptr.3010 [DOI] [PubMed] [Google Scholar]
- Theophel K., Schacht V. J., Schlüter M., Schnell S., Stingu C. S., Schaumann R., et al. (2014). The importance of growth kinetic analysis in determining bacterial susceptibility against antibiotics and silver nanoparticles. Front. Microbiol. 5:544. 10.3389/fmicb.2014.00544 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Traub W. H., Kleber I. (1975). In vitro additive effect of polymyxin B and rifampin against Serratia marcescens. Antimicrob. Agents Chemother. 7, 874–876. 10.1128/AAC.7.6.874 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turlej A., Hryniewicz W., Empel J. (2011). Staphylococcal Cassette Chromosome mec (SCCmec) classification and typing methods: an overview. Pol. J. Microbiol. 60, 95–103. [PubMed] [Google Scholar]
- Ultee A., Kets E. P. W., Smid E. J. (1999). Mechanisms of action of carvacrol on the food-borne pathogen. Appl. Environ. Microbiol. 65, 4606–4610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeh P. J., Hegreness M. J., Aiden A. P., Kishony R. (2009). Drug interactions and the evolution of antibiotic resistance. Nat. Rev. Microbiol. 7, 460–466. 10.1038/nrmicro2133 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.