

Abstract
Insulin resistance has been characterized as attenuation of insulin sensitivity at target organs and tissues, such as muscle and fat tissues and the liver. The insulin signaling cascade is divided into major pathways such as the PI3K/Akt pathway and the MAPK/MEK pathway. In insulin resistance, however, these pathways are not equally impaired. For example, in the liver, inhibition of gluconeogenesis by the insulin receptor substrate (IRS) 2 pathway is impaired, while lipogenesis by the IRS1 pathway is preserved, thus causing hyperglycemia and hyperlipidemia. It has been recently suggested that selective impairment of insulin signaling cascades in insulin resistance also occurs in the kidney. In the renal proximal tubule, insulin signaling via IRS1 is inhibited, while insulin signaling via IRS2 is preserved. Insulin signaling via IRS2 continues to stimulate sodium reabsorption in the proximal tubule and causes sodium retention, edema, and hypertension. IRS1 signaling deficiency in the proximal tubule may impair IRS1-mediated inhibition of gluconeogenesis, which could induce hyperglycemia by preserving glucose production. In the glomerulus, the impairment of IRS1 signaling deteriorates the structure and function of podocyte and endothelial cells, possibly causing diabetic nephropathy. This paper mainly describes selective insulin resistance in the kidney, focusing on the proximal tubule.
1. Introduction
Insulin is one of the essential hormones in humans [1]. It is secreted from pancreatic β cells and regulates glucose homeostasis in various organs and tissues, such as the liver, muscle and fat tissues, and the kidney. However, the roles of insulin in these organs and tissues are pleiotropic and variable. In the liver, insulin inhibits gluconeogenesis, promotes glycogen synthesis, and activates de novo lipogenesis. In the muscle and adipocytes, insulin stimulates glucose uptake [2].
Insulin exerts its activity via signal transduction pathways that start from the binding of insulin to the insulin receptor (IR) [3]. Via insulin receptor substrate (IRS), the signal is transmitted to phosphoinositide 3-kinase (PI3K) and phosphoinositide-dependent kinase-1 (PDK1) and leads to the phosphorylation of Akt. The phosphorylated Akt mediates vital signals such as stimulating protein synthesis, cell survival, transcription, and glycogen synthesis [4]. There are four subtypes of IRS (IRS1 to IRS4), among which IRS1 and IRS2 are the primary mediators involved in insulin signaling [5]. There are other signal transduction pathways initiated by insulin, such as mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK) kinase (MEK) and ERK, which, partly under IRS-mediated signal and partly under Shc that directly mediates signals from the IR, serve to primarily regulate cell growth and proliferation [6, 7]. Figure 1 shows the simplified scheme of the insulin signal transduction network.
Figure 1.
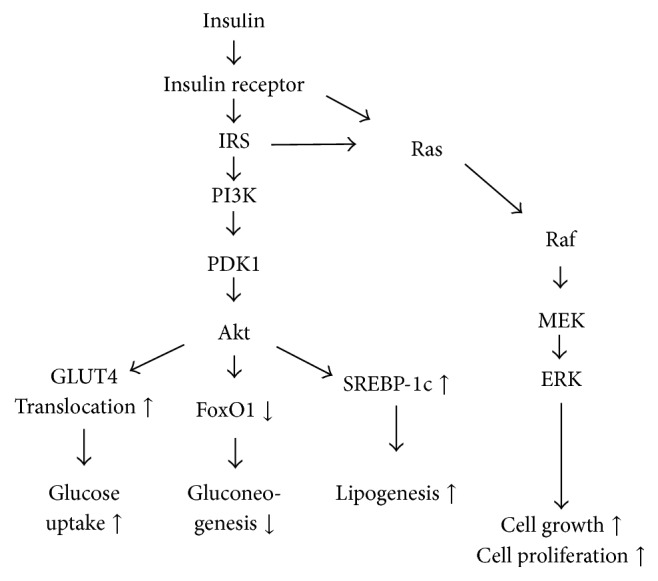
Typical insulin signaling cascade. Insulin binds to insulin receptor in the cell surface [7, 13–15]. The signal goes via IRS, PI3K, and PDK1 to Akt. Akt is a key regulator of this cascade and triggers various signals of physiological responses such as stimulation of glucose uptake, inhibition of gluconeogenesis, and stimulation of lipogenesis. Insulin also stimulates cell growth and proliferation via MEK and ERK cascade. IRS: insulin receptor substrate, PI3K: isoform of phosphatidylinositol 3′-kinase, PDK1: 3′-phosphoinositide-dependent protein kinase-1, GLUT4: glucose transporter type 4, FoxO1: Forkhead box protein O1, and SREBP-1c: sterol regulatory element-binding protein 1c.
However, in insulin resistance, the target organs and/or tissues do not fully respond to insulin. Insulin resistance is characterized by attenuation of the insulin effect. The problem is that the phenotype of insulin resistance is complex; for example, liver insulin receptor knockout mice (LIRKO) show hyperglycemia and hypolipidemia, due to total liver insulin signal deficiency [8]. In contrast, in humans and in animal models of metabolic syndrome-induced insulin resistance, there is hyperglycemia and hyperlipidemia caused by preserved lipogenesis [9]. Additionally, the mechanisms of insulin resistance is different among organs and tissues. For example, in adipose tissue, both IRS1-dependent and IRS2-dependent signals are impaired in insulin resistance. However, in the renal proximal tubule (PT), insulin signaling via IRS1 is impaired but insulin signaling via IRS2 is preserved [10–12].
2. Selective Insulin Resistance in the Liver and Vascular Endothelium
As shown in Figure 1, the insulin signaling cascade has various effects on glucose and lipid metabolism. The insulin signaling cascade mediates signals for gluconeogenesis inhibition by inhibiting the Forkhead box protein O1 (FoxO1) activity [22] and for activation of glucose uptake by inducing glucose transporter 4 (GLUT4) translocation to the plasma membrane [23]. The insulin signaling cascade also induces lipogenesis by activating sterol regulatory element-binding protein- (SREBP-) 1c [13, 24]. In insulin resistance, these effects are impaired to different extents.
In the liver, IRS1 mediates lipogenesis while IRS2 mediates glycogen synthesis. In insulin resistance, the signaling cascade via IRS2 seems to be impaired while the signaling cascade via IRS1 seems relatively intact [25, 26]. Furthermore, in hepatocytes with insulin resistance, SREBP-1c expression is increased, while the expressions of IRS2 and insulin-induced Akt phosphorylation are significantly decreased [26]. The IRS1-dependent pathway is essential for SREBP-1c expression triggered by insulin [25], while the PI3K signaling that is dependent on IRS2 has been shown to be essential for enhancement of glucose uptake by inhibiting glycogen synthase kinase (GSK) 3β [27]. These results suggest that both gluconeogenesis and lipogenesis are stimulated under hepatic insulin resistance, inducing hyperglycemia and hyperlipidemia. The difference between the role of IRS1 and IRS2 in insulin signaling cascade could account for the existence of selective insulin resistance in liver. In fact, the liver is the first organ in which selective insulin resistance was noticed [28–30].
In the liver, the evidence of specific zonation for metabolic pathways [31–34] has accumulated. Two zones, an afferent periportal area and an efferent perivenous region, are known so far. The periportal, upstream area is supplied with blood rich in oxygen and hormones such as insulin and is involved in oxidative energy metabolism and gluconeogenesis. On the other hand, the perivenous downstream area is supplied with blood which is poor in oxygen but rich in CO2 and metabolic products and is engaged in glucose uptake and lipogenesis [34–36]. Recent studies suggest that selective insulin resistance might be related to hepatic zonation; the periportal area becomes insulin resistant, while the perivenous area remains insulin sensitive and thus accompanied with enhanced gluconeogenesis and retained lipogenesis [37].
In vascular endothelial cells, IR/IRS1/PI3K/Akt pathways are thought to be attenuated in insulin resistance, while the ERK/MAPK pathways are not affected in obese Zucker rats [38], suggesting the existence of selective insulin resistance. Insulin increases endothelial nitric oxide (NO) production and endothelial NO synthase (eNOS) gene expression via PI3K and Akt, which is impaired in insulin resistance [39, 40]. The gene expression of eNOS is altered by IRS1 [41]. Mice lacking IRS1 present impaired endothelium-dependent vascular relaxation, suggesting that IRS1 plays a significant role in insulin signal transduction at the vascular endothelium [42]. IRS2−/− mice exhibit more intensive neointima formation compared to wild type and IRS1−/− mice [43]. This attenuation of vascular insulin signaling appears to mainly involve the IRS1-dependent pathway but IRS2-dependent pathway may also be partly involved. On the other hand, preserved ERK/MAPK pathway triggers the expression of endothelin-1, a strong vasoconstrictor that leads to blood pressure elevation [44]. Additionally, the lack of insulin receptor gene and resulting attenuation of insulin signaling were shown to increase the expression of vascular cell adhesion molecule 1 (VCAM-1) [38, 44, 45], a strong inducer of atherosclerosis, showing that loss of insulin signaling could accelerate atherosclerosis. Taken together, selective insulin resistance also exists in the vascular endothelium; that is, the attenuation of IRS/PI3K pathway could impair NO-derived vasodilatation and promote atherosclerosis, while the preserved ERK/MAPK pathway could induce vasoconstriction mediated by endothelin-1.
3. The Roles of IRS1 and IRS2 in Muscle and Adipose Tissue in Insulin Resistance
In muscle, insulin signaling via IRS and PI3K is attenuated, whereas the stimulation of the MAPK pathway by insulin is thought to be intact in hyperinsulinemia and type 2 diabetes mellitus (T2DM) [46–48]. Huang and colleagues showed that gene silencing of IRS1, but not IRS2, in L6 myotube cells reduced GLUT4 translocation and glucose uptake, suggesting that IRS1 is mainly involved in insulin-stimulated glucose uptake [49]. Other studies also suggest that muscle IRS1, but not IRS2, is reduced in insulin resistance [50]. In muscle, insulin resistance seems to attenuate predominantly IRS1-dependent GLUT4 translocation and glucose uptake, though some reports suggest that IRS2-dependent insulin signaling cascade could also be involved in glucose metabolism in myocytes [51, 52].
Adipose tissue is the major site responsible for systemic insulin resistance. Ueki and colleagues showed that suppressor of cytokine signaling 1 (SOCS-1) and SOCS-3 causes impaired insulin signaling through the phosphorylation of IRS1 and IRS2 by separate mechanisms [53]. In human adipocytes, exposure to glucose and high dose insulin for several hours reduced IRS1 expression [54] while, in rat adipocytes, high fat diet reduced both IRS1 and IRS2 content [55, 56]. The reason why IRS1 and IRS2 are not suppressed identically between human and rat adipocytes is not understood.
Insulin induces translocation of GLUT4, the main glucose transporter isoform expressed in adipocytes, from the cytosol to the plasma membrane [57, 58]. Overexpression of human IRS1 in rat adipocytes elevates surface GLUT4 level even without insulin [59]. In insulin resistance, the adipocyte itself is enlarged and the expression of IRS1 and GLUT4 is decreased. Inflammation factors such as TNFα and IL-1β are thought to induce reduction of IRS1 and GLUT4 expression [60, 61].
4. Chronic Kidney Disease (CKD) and Insulin Resistance
Substantial evidence indicates that insulin resistance is accompanied by chronic kidney disease (CKD) [62]. Insulin resistance is frequently seen in patients with advanced or end-stage renal disease and also in patients with mild renal dysfunction [63, 64]. The mechanisms involved in the occurrence and development of insulin resistance in CKD has been clarified to some extent [62]. Inflammation is a notorious contributor to the emergence of insulin resistance. Mediators of chronic inflammation, such as TNF-α, IL-6, and interferon-γ show increased levels in CKD patients [65–67]. The binding of insulin to IR is well preserved, but the signal transduction cascade after insulin binding to IR seems to be impaired [68, 69]. The main inhibitory step is the degradation of IRS1 by the ubiquitin complex, which reduces phosphorylation of Akt that is downstream of IRS1, resulting in abnormal glucose homeostasis and lipid metabolism [70, 71].
5. Recent Findings regarding Insulin Signaling in the Renal Proximal Tubule
In renal tubules, insulin stimulates sodium reabsorption in many tubular segments [12, 72, 73]: the PT [74–77], thick ascending loop of Henle (TAL) [78, 79], distal convoluted tubule (DCT) [80], and cortical collecting duct (CCD) [81]. As for the PT, signal transduction cascade initiated by insulin triggers Akt phosphorylation, mainly via IRS2/PI3K [11, 82]. In the DCT, the insulin signal pathway includes Akt and with-no-lysine kinase (WNK) [83].
We have recently showed [11] that in PTs of the Otsuka Long Evans Tokushima Fatty (OLETF) rat, an animal model of insulin resistance, the stimulatory effect of insulin on sodium bicarbonate cotransporter (NBCe1) activity via the PI3K/Akt pathway was also totally intact. This strongly suggests that, even in insulin resistance, the enhancement of sodium reabsorption via NBCe1 by insulin is preserved. We also showed that, in the kidney cortex of insulin resistant OLETF rats, the expression of IRS1 was decreased to some extent but the expression of IRS2 was totally retained. Why the expression of IRS2 in the kidney cortex is preserved in insulin resistance is not completely understood. However, after feeding and insulin administration, the protein expression of steroid regulatory element-binding protein 1 (SREBP1) was decreased in the liver but preserved in the kidney cortex, whereas the protein expression of Forkhead box protein O1 (FoxO1) was elevated in the liver but unchanged in the kidney cortex. This indicates that the liver and kidney cortex have different regulatory mechanisms for IRS2 expression [11]. Others have also showed that the kidney expression of IRS2 was preserved even in diabetic rats [10]. These facts support our findings of different IRS2 expression regulation between kidney and liver. We also confirmed that this stimulatory effect of insulin on NBCe1 in the PT is preserved in humans with insulin resistance as well. These results suggest a possibility that hyperinsulinemia accompanied with insulin resistance is an important factor for the onset and progression of hypertension in metabolic syndrome and is mediated through the IRS2/PI3K/Akt signaling pathway.
We have also subsequently reported that, in OLETF rats with overt diabetic nephropathy accompanied with massive proteinuria, the stimulatory effect of insulin on renal proximal sodium reabsorption was preserved [16]. The expression of IRS2 and the insulin-induced phosphorylation of Akt in the kidney cortex were also preserved in these rats. Moreover, in the PT of human subjects with type 2 diabetic nephropathy, insulin significantly stimulated NBCe1 activity. These results indicate that the stimulatory effect of insulin on PT sodium reabsorption is preserved even in overt diabetic nephropathy with massive proteinuria. This could at least partially explain why intensive glycemic control for patients with T2DM and CKD is often complicated with massive weight gain, prolonged hyperinsulinemia, and hypoglycemia due to decreased renal function and poor prognosis due to increased cardiovascular risk [84].
In contrast, OLETF rats have a significantly decreased expression of IRS1 in the kidney cortex. This may be relevant to the fact that gluconeogenesis, which is restricted to the PT in the kidney, is enhanced in DM, and this is supported by control experimental animals having suppressed PT gluconeogenesis due to an intact IRS1 pathway [85–88]. In insulin resistance, diabetes, and overt diabetic nephropathy, IRS2-dependent stimulation of sodium transport by insulin in the PT is preserved and this may induce sodium retention, whereas IRS1-dependent suppression of gluconeogenesis is attenuated and might induce hyperglycemia. These findings suggest that insulin signaling is selectively impaired in the PT, under conditions of systemic insulin resistance, diabetes, and even overt diabetic nephropathy.
6. Recent Findings regarding Insulin Signaling in Glomeruli
The glomerulus is composed of three cell types: podocytes, endothelial glomerular cells, and mesangial cells. All of these cells have been shown to respond to insulin stimulation. In the glomerular endothelial cells, insulin can increase nitric oxide (NO) production by stimulating eNOS activity [39]. This effect seems to be impaired in animal models of insulin resistance and diabetes [89, 90]. In primary cultures, podocytes have the highest IR and IRS1 expression levels compared with endothelial cells and mesangial cells [10]. Insulin was suggested to play a role in the regulation of podocyte contractility, which may contribute to glomerular permeability [91, 92]. Podocyte-specific IR knockout mice develop albuminuria, the effacement of podocyte foot process, and podocyte apoptosis. These mice also have increased glomerular matrix level, glomerulosclerosis, and glomerular basement membrane (GBM) thickening, which recapitulates some features of diabetic nephropathy. This suggests that podocyte-specific insulin signaling is crucial for glomerular function [93]. Insulin is also reported to modulate glomerular permeability by controlling podocyte contractility [17]. Insulin exerts its effect on mesangial cells; insulin has been shown to inhibit mesangial cell apoptosis, by activating the PI3K pathway and enhancing mesangial cell proliferation [94–96].
Similarly in glomeruli, many reports suggest that insulin signaling is altered in insulin resistance and diabetes. Using animal models of insulin resistance and T2DM, Mima and colleagues showed an attenuation of glomerular IRS1 expression, IRS1 phosphorylation, and glomerular endothelial signaling. In contrast, IRS2 expression was preserved in these glomerular endothelial cells [10]. The glomerular insulin signaling cascade via Akt2 is thought to be crucial for the maintenance of glomerular function and structure [93, 97]. The impairment of IRS2 signaling in the podocyte in the onset of diabetic nephropathy has been very recently suggested; Santamaria and colleagues demonstrated that phosphatase and tensin homolog (PTEN) and IRS2 were essential for insulin signaling in podocytes [18].
These findings could help to elucidate the mechanisms of the emergence and the progression of diabetic nephropathy. The existence of insulin resistance in type 1 diabetes (T1D) is a remarkable risk factor for the progression to overt nephropathy [98–100]. Additionally, in various animal models of renal injury, thiazolidinediones (TZDs), used clinically for improving insulin sensitivity, are reported to have renoprotective effects on the glomeruli [101–103], suggesting that glomerular insulin resistance could be an important factor for the impairment of renal function.
Taken together, the attenuation of glomerular insulin signaling cascade accompanied with insulin resistance and DM could be related to the emergence and progression of diabetic nephropathy. Conversely, selective insulin resistance in the PT could result in the preservation of the stimulatory effect of insulin on sodium transport as well as the attenuation of inhibitory effect of insulin on gluconeogenesis. This may explain the pathogenetic mechanisms of hypertension accompanied with hyperinsulinemia, edema, and fluid retention as a complication of metabolic syndrome and intensive insulin treatment. Unsuppressed renal gluconeogenesis could also at least partially contribute to hyperglycemia in DM. Thus, the selective insulin resistance in the kidney seems to be a common mechanism linking all negative effects on the emergence and progression of diabetic nephropathy and other complications, in both glomeruli and renal tubules, making the prevention and therapy of diabetic nephropathy even more challenging. In the glomeruli, the decrease of IRS1-dependent signaling impairs the functions of glomerular cells, endothelial cells, podocytes, and mesangial cells. In the PT, IRS2-dependent stimulation of sodium reabsorption could cause hypertension and edema, while impaired IRS1-dependent signaling could induce unsuppressed gluconeogenesis, possibly contributing to hyperglycemia. TZDs are suggested to have protective effects on the glomeruli by ameliorating insulin sensitivity. However, in the PT, TZDs enhance sodium retention [104] and can abolish the beneficial effect of TZDs on glomeruli. Figure 2 summarizes selective insulin resistance in the kidney.
Figure 2.
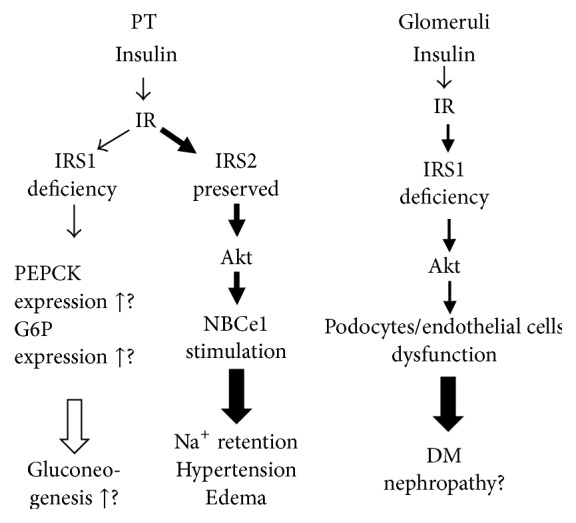
Insulin signaling in insulin resistance in the kidney [10, 11, 16–21]. In the PT, the expression of IRS1 is suppressed while that of IRS2 is preserved. IRS1 signaling deficiency may cause the upregulation of G6P and PEPCK. In this condition, the inhibition of gluconeogenesis does not work sufficiently, causing enhanced gluconeogenesis and hyperglycemia. On the other hand, the expression of IRS2 is preserved, causing stimulation of sodium reabsorption via NBCe1, leading to sodium retention, hypertension, and edema. In the glomeruli, IRS1 signaling is also impaired similar to the PT. This causes podocyte dysfunction, possibly leading to DM nephropathy. Both the PT and glomeruli have selective insulin resistance, with differences between signaling via IRS1 and signaling via IRS2. IR: insulin receptor, IRS: insulin receptor substrate, FoxO1: Forkhead box protein O1, NBCe1: sodium bicarbonate cotransporter type 1, G6P: glucose-6-phosphatase, and PEPCK: phosphoenolpyruvate carboxykinase.
7. Conclusion
This paper provides an overview of the recent findings regarding selective insulin resistance in the kidney. This condition of “selective insulin resistance” has been previously recognized, but it is only recently that the existence of renal “selective insulin resistance” and some of its detailed mechanism have started to be elucidated. The characteristics of insulin resistance differ among organs and tissues; in liver IRS2-mediated pathway is impaired, causing hyperglycemia while IRS1-mediated pathway is preserved to some extent, inducing hyperlipidemia. In the glomerulus, deficiency of IRS1-mediated pathway causes glomerular dysfunction and possibly contributes to diabetic nephropathy. In the PT, IRS2-mediated pathway is preserved and the stimulation of sodium reabsorption by insulin causes sodium retention and possibly subsequent hypertension, whereas the potential impairment of the IRS1-mediated pathway could lead to unsuppressed gluconeogenesis. These facts support the existence of selective insulin resistance in the kidney. TZDs, drugs for improvement of insulin sensitivity, enhance sodium retention in the renal tubules. “Selective insulin resistance” in diabetic nephropathy could explain the challenges of treatment for hypertension and congestive heart failure accompanied with diabetes. Future investigation targeted at the improvement of selective insulin resistance will be of significance for the treatment of diabetes and its complications.
Competing Interests
The authors declare that they have no competing interests.
References
- 1.Banting F. G., Best C. H., Collip J. B., Campbell W. R., Fletcher A. A. Pancreatic extracts in the treatment of diabetes mellitus. Canadian Medical Association Journal. 1922;12(3):141–146. [PMC free article] [PubMed] [Google Scholar]
- 2.Samuel V. T., Shulman G. I. Mechanisms for insulin resistance: common threads and missing links. Cell. 2012;148(5):852–871. doi: 10.1016/j.cell.2012.02.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hale L. J., Coward R. J. The insulin receptor and the kidney. Current Opinion in Nephrology and Hypertension. 2013;22:100–106. doi: 10.1097/MNH.0b013e32835abb52. [DOI] [PubMed] [Google Scholar]
- 4.Hale L. J., Coward R. J. M. Insulin signalling to the kidney in health and disease. Clinical Science. 2013;124(6):351–370. doi: 10.1042/CS20120378. [DOI] [PubMed] [Google Scholar]
- 5.White M. F. IRS proteins and the common path to diabetes. American Journal of Physiology—Endocrinology and Metabolism. 2002;283(3):E413–E422. doi: 10.1152/ajpendo.00514.2001. [DOI] [PubMed] [Google Scholar]
- 6.Gehart H., Kumpf S., Ittner A., Ricci R. MAPK signalling in cellular metabolism: stress or wellness? EMBO Reports. 2010;11(11):834–840. doi: 10.1038/embor.2010.160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Könner A. C., Brüning J. C. Selective insulin and leptin resistance in metabolic disorders. Cell Metabolism. 2012;16(2):144–152. doi: 10.1016/j.cmet.2012.07.004. [DOI] [PubMed] [Google Scholar]
- 8.Michael M. D., Kulkarni R. N., Postic C., et al. Loss of insulin signaling in hepatocytes leads to severe insulin resistance and progressive hepatic dysfunction. Molecular Cell. 2000;6(1):87–97. doi: 10.1016/S1097-2765(05)00015-8. [DOI] [PubMed] [Google Scholar]
- 9.Biddinger S. B., Kahn C. R. From mice to men: insights into the insulin resistance syndromes. Annual Review of Physiology. 2006;68:123–158. doi: 10.1146/annurev.physiol.68.040104.124723. [DOI] [PubMed] [Google Scholar]
- 10.Mima A., Ohshiro Y., Kitada M., et al. Glomerular-specific protein kinase C-β-induced insulin receptor substrate-1 dysfunction and insulin resistance in rat models of diabetes and obesity. Kidney International. 2011;79(8):883–896. doi: 10.1038/ki.2010.526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Nakamura M., Yamazaki O., Shirai A., et al. Preserved Na/HCO3 cotransporter sensitivity to insulin may promote hypertension in metabolic syndrome. Kidney International. 2015;87(3):535–542. doi: 10.1038/ki.2014.351. [DOI] [PubMed] [Google Scholar]
- 12.Nakamura R., Emmanouel D. S., Katz A. I. Insulin binding sites in various segments of the rabbit nephron. The Journal of Clinical Investigation. 1983;72(1):388–392. doi: 10.1172/jci110979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Wu X., Chen K., Williams K. J. The role of pathway-selective insulin resistance and responsiveness in diabetic dyslipoproteinemia. Current Opinion in Lipidology. 2012;23(4):334–344. doi: 10.1097/MOL.0b013e3283544424. [DOI] [PubMed] [Google Scholar]
- 14.Bakan I., Laplante M. Connecting mTORC1 signaling to SREBP-1 activation. Current Opinion in Lipidology. 2012;23(3):226–234. doi: 10.1097/MOL.0b013e328352dd03. [DOI] [PubMed] [Google Scholar]
- 15.Sin T. K., Yung B. Y., Siu P. M. Modulation of SIRT1-foxo1 signaling axis by resveratrol: implications in skeletal muscle aging and insulin resistance. Cellular Physiology and Biochemistry. 2015;35(2):541–552. doi: 10.1159/000369718. [DOI] [PubMed] [Google Scholar]
- 16.Nakamura M., Satoh N., Suzuki M., et al. Stimulatory effect of insulin on renal proximal tubule sodium transport is preserved in type 2 diabetes with nephropathy. Biochemical and Biophysical Research Communications. 2015;461(1):154–158. doi: 10.1016/j.bbrc.2015.04.005. [DOI] [PubMed] [Google Scholar]
- 17.Lay A., Coward R. J. Recent advances in our understanding of insulin signalling to the podocyte. Nephrology Dialysis Transplantation. 2014;29(6):1127–1133. doi: 10.1093/ndt/gft471. [DOI] [PubMed] [Google Scholar]
- 18.Santamaria B., Marquez E., Lay A., et al. IRS2 and PTEN are key molecules in controlling insulin sensitivity in podocytes. Biochimica et Biophysica Acta—Molecular Cell Research. 2015;1853(12):3224–3234. doi: 10.1016/j.bbamcr.2015.09.020. [DOI] [PubMed] [Google Scholar]
- 19.Sun Y., Liu S., Ferguson S., et al. Phosphoenolpyruvate carboxykinase overexpression selectively attenuates insulin signaling and hepatic insulin sensitivity in transgenic mice. The Journal of Biological Chemistry. 2002;277(26):23301–23307. doi: 10.1074/jbc.m200964200. [DOI] [PubMed] [Google Scholar]
- 20.Taniguchi C. M., Ueki K., Kahn C. R. Complementary roles of IRS-1 and IRS-2 in the hepatic regulation of metabolism. The Journal of Clinical Investigation. 2005;115(3):718–727. doi: 10.1172/jci200523187. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 21.Zhang J., Nakatsu Y., Shinjo T., et al. Par14 protein associates with insulin receptor substrate 1 (IRS-1), thereby enhancing insulin-induced IRS-1. The Journal of Biological Chemistry. 2013;288(28):20692–20701. doi: 10.1074/jbc.m113.485730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Matsumoto M., Han S., Kitamura T., Accili D. Dual role of transcription factor FoxO1 in controlling hepatic insulin sensitivity and lipid metabolism. The Journal of Clinical Investigation. 2006;116(9):2464–2472. doi: 10.1172/jci27047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gonzalez E., Flier E., Molle D., Accili D., McGraw T. E. Hyperinsulinemia leads to uncoupled insulin regulation of the GLUT4 glucose transporter and the FoxO1 transcription factor. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(25):10162–10167. doi: 10.1073/pnas.1019268108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Manning B. D., Cantley L. C. AKT/PKB signaling: navigating downstream. Cell. 2007;129(7):1261–1274. doi: 10.1016/j.cell.2007.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Matsumoto M., Ogawa W., Teshigawara K., et al. Role of the insulin receptor substrate 1 and phosphatidylinositol 3-kinase signaling pathway in insulin-induced expression of sterol regulatory element binding protein 1c and glucokinase genes in rat hepatocytes. Diabetes. 2002;51(6):1672–1680. doi: 10.2337/diabetes.51.6.1672. [DOI] [PubMed] [Google Scholar]
- 26.Shimomura I., Matsuda M., Hammer R. E., Bashmakov Y., Brown M. S., Goldstein J. L. Decreased IRS-2 and increased SREBP-1c lead to mixed insulin resistance and sensitivity in livers of lipodystrophic and ob/ob mice. Molecular Cell. 2000;6(1):77–86. doi: 10.1016/S1097-2765(05)00010-9. [DOI] [PubMed] [Google Scholar]
- 27.Landis J., Shaw L. M. Insulin receptor substrate 2-mediated phosphatidylinositol 3-kinase signaling selectively inhibits glycogen synthase kinase 3β to regulate aerobic glycolysis. The Journal of Biological Chemistry. 2014;289(26):18603–18613. doi: 10.1074/jbc.m114.564070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Biddinger S. B., Hernandez-Ono A., Rask-Madsen C., et al. Hepatic insulin resistance is sufficient to produce dyslipidemia and susceptibility to atherosclerosis. Cell Metabolism. 2008;7(2):125–134. doi: 10.1016/j.cmet.2007.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Brown M. S., Goldstein J. L. Selective versus total insulin resistance: a pathogenic paradox. Cell Metabolism. 2008;7(2):95–96. doi: 10.1016/j.cmet.2007.12.009. [DOI] [PubMed] [Google Scholar]
- 30.Laplante M., Sabatini D. M. mTORC1 activates SREBP-1c and uncouples lipogenesis from gluconeogenesis. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(8):3281–3282. doi: 10.1073/pnas.1000323107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gumucio J. J. Hepatocyte heterogeneity: the coming of age from the description of a biological curiosity to a partial understanding of its physiological meaning and regulation. Hepatology. 1989;9(1):154–160. doi: 10.1002/hep.1840090124. [DOI] [PubMed] [Google Scholar]
- 32.Guzman M., Castro J. Zonation of fatty acid metabolism in rat liver. Biochemical Journal. 1989;264(1):107–113. doi: 10.1042/bj2640107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Jungermann K., Kietzmann T. Zonation of parenchymal and nonparenchymal metabolism in liver. Annual Review of Nutrition. 1996;16:179–203. doi: 10.1146/annurev.nu.16.070196.001143. [DOI] [PubMed] [Google Scholar]
- 34.Postic C., Girard J. Contribution of de novo fatty acid synthesis to hepatic steatosis and insulin resistance: lessons from genetically engineered mice. The Journal of Clinical Investigation. 2008;118(3):829–838. doi: 10.1172/jci34275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Braeuning A., Ittrich C., Köhle C., et al. Differential gene expression in periportal and perivenous mouse hepatocytes. FEBS Journal. 2006;273(22):5051–5061. doi: 10.1111/j.1742-4658.2006.05503.x. [DOI] [PubMed] [Google Scholar]
- 36.Katz N. R. Metabolic heterogeneity of hepatocytes across the liver acinus. The Journal of Nutrition. 1992;122(3):843–849. doi: 10.1093/jn/122.suppl_3.843. [DOI] [PubMed] [Google Scholar]
- 37.Hijmans B. S., Grefhorst A., Oosterveer M. H., Groen A. K. Zonation of glucose and fatty acid metabolism in the liver: mechanism and metabolic consequences. Biochimie. 2014;96(1):121–129. doi: 10.1016/j.biochi.2013.06.007. [DOI] [PubMed] [Google Scholar]
- 38.Jiang Z. Y., Lin Y.-W., Clemont A., et al. Characterization of selective resistance to insulin signaling in the vasculature of obese Zucker (fa/fa) rats. The Journal of Clinical Investigation. 1999;104(4):447–457. doi: 10.1172/jci5971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kuboki K., Jiang Z. Y., Takahara N., et al. Regulation of endothelial constitutive nitric oxide synthase gene expression in endothelial cells and in vivo: a specific vascular action of insulin. Circulation. 2000;101(6):676–681. doi: 10.1161/01.cir.101.6.676. [DOI] [PubMed] [Google Scholar]
- 40.Montagnani M., Chen H., Barr V. A., Quon M. J. Insulin-stimulated Activation of eNOS Is Independent of Ca2+ but Requires Phosphorylation by Akt at Ser1179. The Journal of Biological Chemistry. 2001;276(32):30392–30398. doi: 10.1074/jbc.m103702200. [DOI] [PubMed] [Google Scholar]
- 41.Federici M., Pandolfi A., De Filippis E. A., et al. G972R IRS-1 variant impairs insulin regulation of endothelial nitric oxide synthase in cultured human endothelial cells. Circulation. 2004;109(3):399–405. doi: 10.1161/01.cir.0000109498.77895.6f. [DOI] [PubMed] [Google Scholar]
- 42.Abe H., Yamada N., Kamata K., et al. Hypertension, hypertriglyceridemia, and impaired endothelium-dependent vascular relaxation in mice lacking insulin receptor substrate-1. The Journal of Clinical Investigation. 1998;101(8):1784–1788. doi: 10.1172/jci1594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kubota T., Kubota N., Moroi M., et al. Lack of insulin receptor substrate-2 causes progressive neointima formation in response to vessel injury. Circulation. 2003;107(24):3073–3080. doi: 10.1161/01.CIR.0000070937.52035.25. [DOI] [PubMed] [Google Scholar]
- 44.Rask-Madsen C., King G. L. Mechanisms of disease: endothelial dysfunction in insulin resistance and diabetes. Nature Clinical Practice Endocrinology and Metabolism. 2007;3(1):46–56. doi: 10.1038/ncpendmet0366. [DOI] [PubMed] [Google Scholar]
- 45.Rask-Madsen C., Li Q., Freund B., et al. Loss of insulin signaling in vascular endothelial cells accelerates atherosclerosis in apolipoprotein E null mice. Cell Metabolism. 2010;11(5):379–389. doi: 10.1016/j.cmet.2010.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Cusi K., Maezono K., Osman A., et al. Insulin resistance differentially affects the PI 3-kinase- and MAP kinase-mediated signaling in human muscle. The Journal of Clinical Investigation. 2000;105(3):311–320. doi: 10.1172/jci7535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Goodyear L. J., Giorgino F., Sherman L. A., Carey J., Smith R. J., Dohm G. L. Insulin receptor phosphorylation, insulin receptor substrate-1 phosphorylation, and phosphatidylinositol 3-kinase activity are decreased in intact skeletal muscle strips from obese subjects. The Journal of Clinical Investigation. 1995;95(5):2195–2204. doi: 10.1172/jci117909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kim Y.-B., Nikoulina S. E., Ciaraldi T. P., Henry R. R., Kahn B. B. Normal insulin-dependent activation of Akt/protein kinase B, with diminished activation of phosphoinositide 3-kinase, in muscle in type 2 diabetes. The Journal of Clinical Investigation. 1999;104(6):733–741. doi: 10.1172/jci6928. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Huang C., Thirone A. C. P., Huang X., Klip A. Differential contribution of insulin receptor substrates 1 versus 2 to insulin signaling and glucose uptake in L6 myotubes. The Journal of Biological Chemistry. 2005;280(19):19426–19435. doi: 10.1074/jbc.m412317200. [DOI] [PubMed] [Google Scholar]
- 50.Thirone A. C. P., Huang C., Klip A. Tissue-specific roles of IRS proteins in insulin signaling and glucose transport. Trends in Endocrinology and Metabolism. 2006;17(2):72–78. doi: 10.1016/j.tem.2006.01.005. [DOI] [PubMed] [Google Scholar]
- 51.Agarwal P., Srivastava R., Srivastava A. K., Ali S., Datta M. MiR-135a targets IRS2 and regulates insulin signaling and glucose uptake in the diabetic gastrocnemius skeletal muscle. Biochimica et Biophysica Acta—Molecular Basis of Disease. 2013;1832(8):1294–1303. doi: 10.1016/j.bbadis.2013.03.021. [DOI] [PubMed] [Google Scholar]
- 52.Pirola L., Bonnafous S., Johnston A. M., Chaussade C., Portis F., Van Obberghen E. Phosphoinositide 3-kinase-mediated reduction of insulin receptor substrate-1/2 protein expression via different mechanisms contributes to the insulin-induced desensitization of its signaling pathways in L6 muscle cells. The Journal of Biological Chemistry. 2003;278(18):15641–15651. doi: 10.1074/jbc.m208984200. [DOI] [PubMed] [Google Scholar]
- 53.Ueki K., Kondo T., Kahn C. R. Suppressor of cytokine signaling 1 (SOCS-1) and SOCS-3 cause insulin resistance through inhibition of tyrosine phosphorylation of insulin receptor substrate proteins by discrete mechanisms. Molecular and Cellular Biology. 2004;24(12):5434–5446. doi: 10.1128/MCB.24.12.5434-5446.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Renström F., Burén J., Svensson M., Eriksson J. W. Insulin resistance induced by high glucose and high insulin precedes insulin receptor substrate 1 protein depletion in human adipocytes. Metabolism: Clinical and Experimental. 2007;56(2):190–198. doi: 10.1016/j.metabol.2006.09.012. [DOI] [PubMed] [Google Scholar]
- 55.Anai M., Ono H., Funaki M., et al. Different subcellular distribution and regulation of expression of insulin receptor substrate (IRS)-3 from those of IRS-1 and IRS-2. The Journal of Biological Chemistry. 1998;273(45):29686–29692. doi: 10.1074/jbc.273.45.29686. [DOI] [PubMed] [Google Scholar]
- 56.Renström F., Burén J., Eriksson J. W. Insulin receptor substrates-1 and -2 are both depleted but via different mechanisms after down-regulation of glucose transport in rat adipocytes. Endocrinology. 2005;146(7):3044–3051. doi: 10.1210/en.2004-1675. [DOI] [PubMed] [Google Scholar]
- 57.Marette A., Richardson J. M., Ramlal T., et al. Abundance, localization, and insulin-induced translocation of glucose transporters in red and white muscle. American Journal of Physiology—Cell Physiology. 1992;263(2):C443–C452. doi: 10.1152/ajpcell.1992.263.2.C443. [DOI] [PubMed] [Google Scholar]
- 58.Satoh S., Nishimura H., Clark A. E., et al. Use of bismannose photolabel to elucidate insulin-regulated GLUT4 subcellular trafficking kinetics in rat adipose cells. Evidence that exocytosis is a critical site of hormone action. The Journal of Biological Chemistry. 1993;268(24):17820–17829. [PubMed] [Google Scholar]
- 59.Quon M. J., Butte A. J., Zarnowski M. J., Sesti G., Cushman S. W., Taylor S. I. Insulin receptor substrate 1 mediates the stimulatory effect of insulin on GLUT4 translocation in transfected rat adipose cells. The Journal of Biological Chemistry. 1994;269(45):27920–27924. [PubMed] [Google Scholar]
- 60.Gustafson B., Gogg S., Hedjazifar S., Jenndahl L., Hammarstedt A., Smith U. Inflammation and impaired adipogenesis in hypertrophic obesity in man. American Journal of Physiology—ndocrinology and Metabolism. 2009;297(5):E999–E1003. doi: 10.1152/ajpendo.00377.2009. [DOI] [PubMed] [Google Scholar]
- 61.Gustafson B., Hammarstedt A., Andersson C. X., Smith U. Inflamed adipose tissue: a culprit underlying the metabolic syndrome and atherosclerosis. Arteriosclerosis, Thrombosis, and Vascular Biology. 2007;27(11):2276–2283. doi: 10.1161/atvbaha.107.147835. [DOI] [PubMed] [Google Scholar]
- 62.Thomas S. S., Zhang L., Mitch W. E. Molecular mechanisms of insulin resistance in chronic kidney disease. Kidney International. 2015;88(6):1233–1239. doi: 10.1038/ki.2015.305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.DeFronzo R. A., Alvestrand A., Smith D., Hendler R., Hendler E., Wahren J. Insulin resistance in uremia. The Journal of Clinical Investigation. 1981;67(2):563–568. doi: 10.1172/jci110067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fliser D., Pacini G., Engelleiter R., et al. Insulin resistance and hyperinsulinemia are already present in patients with incipient renal disease. Kidney International. 1998;53(5):1343–1347. doi: 10.1046/j.1523-1755.1998.00898.x. [DOI] [PubMed] [Google Scholar]
- 65.Almquist T., Jacobson S. H., Mobarrez F., Näsman P., Hjemdahl P. Lipid-lowering treatment and inflammatory mediators in diabetes and chronic kidney disease. European Journal of Clinical Investigation. 2014;44(3):276–284. doi: 10.1111/eci.12230. [DOI] [PubMed] [Google Scholar]
- 66.Meuwese C. L., Snaedal S., Halbesma N., et al. Trimestral variations of C-reactive protein, interleukin-6 and tumour necrosis factor-α are similarly associated with survival in haemodialysis patients. Nephrology Dialysis Transplantation. 2011;26(4):1313–1318. doi: 10.1093/ndt/gfq557. [DOI] [PubMed] [Google Scholar]
- 67.Pereira B. J. G., Sundaram S., Snodgrass B., Hogan P., King A. J. Plasma lipopolysaccharide binding protein and bactericidal/permeability increasing factor in CRF and HD patients. Journal of the American Society of Nephrology. 1996;7(3):479–487. doi: 10.1681/ASN.V73479. [DOI] [PubMed] [Google Scholar]
- 68.Friedman J. E., Dohm G. L., Elton C. W., et al. Muscle insulin resistance in uremic humans: glucose transport, glucose transporters, and insulin receptors. American Journal of Physiology—Endocrinology and Metabolism. 1991;261(1):E87–E94. doi: 10.1152/ajpendo.1991.261.1.E87. [DOI] [PubMed] [Google Scholar]
- 69.Smith D., DeFronzo R. A. Insulin resistance in uremia mediated by postbinding defects. Kidney International. 1982;22(1):54–62. doi: 10.1038/ki.1982.132. [DOI] [PubMed] [Google Scholar]
- 70.Lecker S. H., Goldberg A. L., Mitch W. E. Protein degradation by the ubiquitin-proteasome pathway in normal and disease states. Journal of the American Society of Nephrology. 2006;17(7):1807–1819. doi: 10.1681/ASN.2006010083. [DOI] [PubMed] [Google Scholar]
- 71.Mitch W. E., Goldberg A. L. Mechanisms of muscle wasting. The role of the ubiquitin-proteasome pathway. The New England Journal of Medicine. 1996;335(25):1897–1905. doi: 10.1056/nejm199612193352507. [DOI] [PubMed] [Google Scholar]
- 72.Natali A., Quinones Galvan A., Santoro D., et al. Relationship between insulin release, antinatriuresis and hypokalaemia after glucose ingestion in normal and hypertensive man. Clinical Science. 1993;85(3):327–335. doi: 10.1042/cs0850327. [DOI] [PubMed] [Google Scholar]
- 73.Quiñones-Galvan A., Ferrannini E. Renal effects of insulin in man. Journal of Nephrology. 1997;10(4):188–191. [PubMed] [Google Scholar]
- 74.Baum M. Insulin stimulates volume absorption in the rabbit proximal convoluted tubule. The Journal of Clinical Investigation. 1987;79(4):1104–1109. doi: 10.1172/jci112925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Feraille E., Carranza M. L., Rousselot M., Favre H. Insulin enhances sodium sensitivity of Na-K-ATPase in isolated rat proximal convoluted tubule. American Journal of Physiology. 1994;267(1, part 2):F55–F62. doi: 10.1152/ajprenal.1994.267.1.F55. [DOI] [PubMed] [Google Scholar]
- 76.Gesek F. A., Schoolwerth A. C. Insulin increases Na+-H+ exchange activity in proximal tubules from normotensive and hypertensive rats. American Journal of Physiology—Renal Fluid and Electrolyte Physiology. 1991;260(5):F695–F703. doi: 10.1152/ajprenal.1991.260.5.F695. [DOI] [PubMed] [Google Scholar]
- 77.Ruiz O. S., Qiu Y.-Y., Cardoso L. R., Arruda J. A. L. Regulation of the renal Na-HCO3 cotransporter: IX. Modulation by insulin, epidermal growth factor and carbachol. Regulatory Peptides. 1998;77(1–3):155–161. doi: 10.1016/s0167-0115(98)00115-3. [DOI] [PubMed] [Google Scholar]
- 78.Ito O., Kondo Y., Takahashi N., et al. Insulin stimulates NaCl transport in isolated perfused MTAL of Henle's loop of rabbit kidney. American Journal of Physiology—Renal Fluid and Electrolyte Physiology. 1994;267(2):F265–F270. doi: 10.1152/ajprenal.1994.267.2.F265. [DOI] [PubMed] [Google Scholar]
- 79.Mandon B., Siga E., Chabardes D., Firsov D., Roinel N., De Rouffignac C. Insulin stimulates Na+, Cl-, Ca2+, and Mg2+ transports in TAL of mouse nephron: cross-potentiation with AVP. American Journal of Physiology—Renal Physiology. 1993;265(3, part 2):F361–F369. doi: 10.1152/ajprenal.1993.265.3.F361. [DOI] [PubMed] [Google Scholar]
- 80.Sohara E., Rai T., Yang S.-S., et al. Acute insulin stimulation induces phosphorylation of the Na-Cl cotransporter in cultured distal mpkDCT cells and mouse kidney. PLoS ONE. 2011;6(8) doi: 10.1371/journal.pone.0024277.e24277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Feraille E., Rousselot M., Rajerison R., Favre H. Effect of insulin on Na+,K(+)-ATPase in rat collecting duct. The Journal of Physiology. 1995;488(1):171–180. doi: 10.1113/jphysiol.1995.sp020955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zheng Y., Yamada H., Sakamoto K., et al. Roles of insulin receptor substrates in insulin-induced stimulation of renal proximal bicarbonate absorption. Journal of the American Society of Nephrology. 2005;16(8):2288–2295. doi: 10.1681/ASN.2005020193. [DOI] [PubMed] [Google Scholar]
- 83.Nishida H., Sohara E., Nomura N., et al. Phosphatidylinositol 3-kinase/akt signaling pathway activates the WNK-OSR1/SPAK-NCC phosphorylation cascade in hyperinsulinemic db/db mice. Hypertension. 2012;60(4):981–990. doi: 10.1161/HYPERTENSIONAHA.112.201509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Papademetriou V., Lovato L., Doumas M., et al. Chronic kidney disease and intensive glycemic control increase cardiovascular risk in patients with type 2 diabetes. Kidney International. 2015;87(3):649–659. doi: 10.1038/ki.2014.296. [DOI] [PubMed] [Google Scholar]
- 85.Gatica R., Bertinat R., Silva P., et al. Altered expression and localization of insulin receptor in proximal tubule cells from human and rat diabetic kidney. Journal of Cellular Biochemistry. 2013;114(3):639–649. doi: 10.1002/jcb.24406. [DOI] [PubMed] [Google Scholar]
- 86.Gerich J. E., Woerle H. J., Meyer C., Stumvoll M. Renal gluconeogenesis: its importance in human glucose homeostasis. Diabetes Care. 2001;24(2):382–391. doi: 10.2337/diacare.24.2.382. [DOI] [PubMed] [Google Scholar]
- 87.Meyer C., Stumvoll M., Nadkarni V., Dostou J., Mitrakou A., Gerich J. Abnormal renal and hepatic glucose metabolism in type 2 diabetes mellitus. The Journal of Clinical Investigation. 1998;102(3):619–624. doi: 10.1172/jci2415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Meyer C., Woerle H. J., Dostou J. M., Welle S. L., Gerich J. E. Abnormal renal, hepatic, and muscle glucose metabolism following glucose ingestion in type 2 diabetes. American Journal of Physiology—Endocrinology and Metabolism. 2004;287(6):E1049–E1056. doi: 10.1152/ajpendo.00041.2004. [DOI] [PubMed] [Google Scholar]
- 89.Bohlen H. G. Protein kinase βII in Zucker obese rats compromises oxygen and flow-mediated regulation of nitric oxide formation. American Journal of Physiology—Heart and Circulatory Physiology. 2004;286(2):H492–H497. doi: 10.1152/ajpheart.00818.2003. [DOI] [PubMed] [Google Scholar]
- 90.Naruse K., Rask-Madsen C., Takahara N., et al. Activation of vascular protein kinase C-β inhibits Akt-dependent endothelial nitric oxide synthase function in obesity-associated insulin resistance. Diabetes. 2006;55(3):691–698. doi: 10.2337/diabetes.55.03.06.db05-0771. [DOI] [PubMed] [Google Scholar]
- 91.Kim E. Y., Anderson M., Dryer S. E. Insulin increases surface expression of TRPC6 channels in podocytes: role of NADPH oxidases and reactive oxygen species. American Journal of Physiology—Renal Physiology. 2012;302(3):F298–F307. doi: 10.1152/ajprenal.00423.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kim E. Y., Dryer S. E. Effects of insulin and high glucose on mobilization of slo1 BKCa channels in podocytes. Journal of Cellular Physiology. 2011;226(9):2307–2315. doi: 10.1002/jcp.22567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Welsh G. I., Hale L. J., Eremina V., et al. Insulin signaling to the glomerular podocyte is critical for normal kidney function. Cell Metabolism. 2010;12(4):329–340. doi: 10.1016/j.cmet.2010.08.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Alessi D. R., Downes C. P. The role of PI 3-kinase in insulin action. Biochimica et Biophysica Acta—Lipids and Lipid Metabolism. 1998;1436(1-2):151–164. doi: 10.1016/s0005-2760(98)00133-7. [DOI] [PubMed] [Google Scholar]
- 95.Khwaja A. Apoptosis: Akt is more than just a bad kinase. Nature. 1999;401(6748):33–34. doi: 10.1038/43354. [DOI] [PubMed] [Google Scholar]
- 96.Knight S. F., Imig J. D. Obesity, insulin resistance, and renal function. Microcirculation. 2007;14(4-5):349–362. doi: 10.1080/10739680701283018. [DOI] [PubMed] [Google Scholar]
- 97.Canaud G., Bienaimé F., Viau A., et al. AKT2 is essential to maintain podocyte viability and function during chronic kidney disease. Nature Medicine. 2013;19(10):1288–1296. doi: 10.1038/nm.3313. [DOI] [PubMed] [Google Scholar]
- 98.Bjornstad P., Snell-Bergeon J. K., Rewers M., et al. Early diabetic nephropathy: a complication of reduced insulin sensitivity in type 1 diabetes. Diabetes Care. 2013;36(11):3678–3683. doi: 10.2337/dc13-0631. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Orchard T. J., Chang Y.-F., Ferrell R. E., Petro N., Ellis D. E. Nephropathy in type 1 diabetes: a manifestation of insulin resistance and multiple genetic susceptibilities? Further evidence from the Pittsburgh Epidemiology of Diabetes Complication Study. Kidney International. 2002;62(3):963–970. doi: 10.1046/j.1523-1755.2002.00507.x. [DOI] [PubMed] [Google Scholar]
- 100.Pop A., Clenciu D., Anghel M., et al. Insulin resistance is associated with all chronic complications in type 1 diabetes. Journal of Diabetes. 2016;8:220–228. doi: 10.1111/1753-0407.12283. [DOI] [PubMed] [Google Scholar]
- 101.Lee S., Kim W., Kang K. P., et al. Agonist of peroxisome proliferator-activated receptor-γ, rosiglitazone, reduces renal injury and dysfunction in a murine sepsis model. Nephrology Dialysis Transplantation. 2005;20:1057–1065. doi: 10.1093/ndt/gfh705. [DOI] [PubMed] [Google Scholar]
- 102.Sivarajah A., Chatterjee P. K., Patel N. S. A., et al. Agonists of peroxisome-proliferator activated receptor-gamma reduce renal ischemia/reperfusion injury. American Journal of Nephrology. 2003;23(4):267–276. doi: 10.1159/000072088. [DOI] [PubMed] [Google Scholar]
- 103.Yang H.-C., Deleuze S., Zuo Y., Potthoff S. A., Ma L.-J., Fogo A. B. The PPARγ agonist pioglitazone ameliorates aging-related progressive renal injury. Journal of the American Society of Nephrology. 2009;20(11):2380–2388. doi: 10.1681/asn.2008111138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Endo Y., Suzuki M., Yamada H., et al. Thiazolidinediones enhance sodium-coupled bicarbonate absorption from renal proximal tubules via PPARγ-dependent nongenomic signaling. Cell Metabolism. 2011;13(5):550–561. doi: 10.1016/j.cmet.2011.02.015. [DOI] [PubMed] [Google Scholar]