

Abstract
Significance: All organs suffer wounds to some extent during an animal's lifetime and to compensate for cell loss, tissues often rely on cell division. However, many organs are made up of differentiated cells with only a limited capacity to divide. It is not well understood how cells are replaced in the absence of cell division.
Recent Advances: Recent studies in the model organism Drosophila melanogaster have proven that wound-induced polyploidy (WIP) is an essential mechanism to replace tissue mass and restore tissue integrity in the absence of cell division. In this repair mechanism, preexisting differentiated cells increase their DNA content and cell size by becoming polyploid.
Critical Issues: Cells within mammalian organs such as the liver, heart, and cornea have also been observed to increase their DNA ploidy in response to injury, suggesting that WIP may be an evolutionarily conserved mechanism to compensate for cell loss.
Future Directions: The Hippo signal transduction pathway is required for differentiated cells to initiate WIP in Drosophila. Continued studies in Drosophila will help to identify other signaling pathways required for WIP as well as the conserved mechanisms that polyploid cells may play during wound repair in all organisms.

Vicki P. Losick, PhD
Scope and Significance
A key step in wound repair is the replacement of lost cells. Cells that are lost or damaged from injury must be replaced to restore tissue integrity and homeostasis. Traditionally, research has focused on how cells divide to replace lost cells, but recent research in Drosophila melanogaster has demonstrated that cells can also enlarge their cell size and DNA content by wound-induced polyploidy (WIP). This critical review focuses on the recent observations in Drosophila that show polyploidy can restore tissue mass and integrity as an alternative to the replacement of cell number.1,2
Translational Relevance
Many cells in the human body increase their DNA content resulting in cells with more than the diploid chromosome number, which is defined as polyploidy. This genomic change has been observed in various cell types in human tissues following injury or as the consequence of aging. The morphological architecture of the tissue is often changed under these circumstances since there are fewer, but larger cells remaining. These changes are often considered pathological since they result in an altered tissue form. However, polyploidy can be beneficial to the immediate requirement to compensate for cell loss; without this change, it is possible that tissue integrity would be further compromised.
Clinical Relevance
Novel human wound healing therapies can be uncovered by using Drosophila as a model to address the importance of polyploidy in tissue repair. The breadth of genetic tools in the fruit fly will provide insight into the genetic factors required for WIP. Pharmacological drug libraries can also be used to identify targets that regulate polyploidy. These combinatorial screening strategies can be used to identify candidate genes or pharmacological agents that can promote the healing of human and mouse tissues, particularly in the liver, heart, and cornea where polyploidy has been observed to occur in response to injury.3–5
Background
Organs require an adequate level of cell proliferation during normal development. Too few or too many cells will compromise function resulting in atrophy or hyperplasia, respectively. This is also true when it comes to tissue repair. The cells that are damaged or lost due to injury will need to be replaced to restore tissue homeostasis. Stem cells are present in many tissues, including but not limited to the skin, the intestine, and the skeletal muscle; they serve as a reservoir for new cells when injury occurs.6–8 There is often a near perfect regeneration of the damaged tissue in these cases in terms of physiology and tissue architecture. However, not all tissues have stem cells and our understanding of how tissue repair occurs without stem cells is lacking.
D. melanogaster is emerging as a powerful model organism to study the cellular and molecular mechanisms of tissue repair and regeneration. Over the last decade, research has identified conserved mechanisms of wound healing in embryonic and larval animals.9,10 However, the adult remains one of the last systems to be characterized for tissue repair and regeneration in Drosophila and poses a great opportunity to examine how genetic factors impact these processes independent of the roles they serve in developmental contexts.
Tissues in the adult fruit fly repair by different cellular mechanisms. The Drosophila intestine was discovered to have stem cells, like the mammalian gut.11,12 The intestinal stem cells are required for normal gut homeostasis as well as to regenerate cells lost to damage.13 Stem cells have also been identified in the fly ovary and testes. Somatic and germline stem cells support the production of eggs and sperm throughout Drosophila's life, but research has mostly focused on how stem cells are regulated by normal homeostasis and aging.14 For the remaining adult fruit fly organs, the repair strategies are only beginning to be examined. Recent studies have shown that tissue damage is repaired solely by WIP in some Drosophila tissues lacking stem cells.1
Discussion
Cellular mechanisms generating polyploid cells
Cells have the capacity to divide and to grow. These terms are often used synonymously, but are actually distinct terms describing two different cellular processes: an increase in cell number versus an increase in cell size.15 Cells can increase their size by a variety of cellular mechanisms and these are not all equivalent. Some cell enlargement mechanisms rely on the cell cycle and some do not. An example of a cell cycle-independent mechanism that can lead to increases in cell size has been established in the skeletal muscle, within which the rate of transcription and translation exceeds the rate of protein turnover.16 This is known as cellular hypertrophy. This mechanism has also been observed in other muscle cell types such as the heart cardiomyocytes.17 Cellular hypertrophy is the leading cause of organ hypertrophy resulting in an enlarged organ that is no longer equally proportioned.
Cells can also increase their size in a cell cycle-dependent manner by increasing their DNA content (C) resulting in cells with more than the diploid chromosome copy (Fig. 1). Cells with 4C or greater DNA content are considered to be polyploid. As a consequence, ploidy is determined by both the nuclear DNA content as well as the number of nuclei per cell. Cells can change their DNA content (and consequently, the cell size) by engaging in alternative cell cycles; two examples—endoreplication and endomitosis—are depicted in Fig. 1. During endoreplication (also referred to as the endocycle), a cell repetitively duplicates its genome in the DNA synthesis (S) phase and grows in the gap (G) phase skipping mitosis entirely, resulting in a mononucleated cell with increased ploidy. Successive endocycles will lead to higher ploidy values resulting in DNA content values of more than 2000C.18 Cells can also go through incomplete mitosis and fail cytokinesis, a process known as endomitosis, resulting in an enlarged binucleated cell (Fig. 1). Multinucleated polyploid cells can also be generated by cell-to-cell fusion events (Fig. 1). As a consequence, repetitive endocycles, endomitotic cycles, cell fusions, or a combination of these mechanisms will generate larger more polyploid cells. These alternative cell cycles are common in many plants and invertebrate cell types, but are also found in mammalian cells in the placenta, blood, heart, and liver tissues.4,18 In this study, WIP is defined as the enlargement of a cell size by the required increase in its DNA content.
Figure 1.

Cellular mechanisms of polyploid cell generation. Cells with more than diploid genomic content 4C or greater are considered polyploid. There are multiple mechanisms cells use to become polyploid. A diploid cell (brown) can enter an endocycle (A), where a cell duplicates its genome to become a mononucleated polyploid cell by bypassing mitosis. Cells can also go through endomitosis (B) where M phase is truncated (M*). In this case, a cell does not complete cytokinesis resulting in a binucleated polyploid cell. (C) Cells can also fuse together generating one enlarged multinucleated cell. In these cases, DNA ploidy is linked to cell size with increases in one value typically associated with the proportional increase in the other.18 Successive events or combinations of these processes will result in even larger cells with higher ploidy values.
WIP restores tissue mass and integrity
Most of the adult tissues in Drosophila are quiescent, including the epidermis, posterior region of the hindgut pylorus, brain, heart, muscle, and lower reproductive tract.1,19,* Quiescence means a state of dormancy with the potential to be reactivated. In the context of the cell cycle, this would describe cells that are not actively cycling, but have the potential to reenter the cell cycle. It is often the case that the regenerative potential of a tissue is not appreciated until damage occurs. In mice, quiescent muscle stem cells (known as satellite cells) only become activated to regenerate the skeletal muscle when there is severe trauma to the tissue.6 Injury to heart also stimulates quiescent cardiomyocytes to reenter the cell cycle even though their capacity to repair the heart is limited.4,20 Quiescent cells in Drosophila's posterior hindgut pylorus and epidermis respond to injury by reentering the cell cycle, but these cells grow through WIP to restore tissue integrity.1
The adult fly hindgut is functionally similar to the mammalian colon and acts in the absorption and excretion of water and ions.21 It lies posterior to the fly midgut and connects through a valve junction known as the pylorus. The most anterior strip of cells in the pylorus has a low level of mitotic activity, whereas the remaining posterior region consists of quiescent diploid cells.1,21 Internal organs are not readily accessible to some forms of injury and genetic approaches have become useful tools to induce damage to these tissues. Apoptotic genes can be expressed in a tissue-specific and conditional manner using the yeast-derived Gal4/UAS expression system.22 Quiescent pyloric cells endoreplicated in response to the apoptotic tissue damage and repair the tissue within 1 week by producing large polyploid cells.1
A similar wound-induced response is also evident in the adult Drosophila epidermis.1 The epidermis is made up of a single sheet of diploid epidermal cells that lies in direct contact with the cuticle exoskeleton (Fig. 2).1 It was first observed that puncture wounds heal quickly by forming a scab to close the cuticle thereby acting similarly to a mammalian scab to plug the wound as well as hold the edges of wound together to promote wound healing.23 The epithelium was observed to reseal under the scab,23 but the cellular mechanism of reepithelialization was only recently discovered.1 Epithelial cells surrounding the injury site reenter S phase (Fig. 3), but surprisingly, do not divide and instead heal a puncture wound by becoming polyploid. In this tissue, the polyploid cells are generated by two distinct mechanisms: endoreplication and cell fusion (Fig. 1). As a consequence, the epithelium repairs by generating a very large polyploid cell, which covers the area under the scab with smaller polyploid cells on the periphery (Fig. 2).1 The generated polyploid cells are critical for wound repair since when both endoreplication and cell fusion are perturbed simultaneously the wound fails to close (Fig. 2). Similar to the repair strategy of the hindgut pylorus, there was a close correlation with the number of quiescent cells that reentered S phase and the size of the wound.1 As a result, the number of polyploid cells observed in the epidermis and hindgut pylorus was sufficient to regenerate the number of cells that were lost.1 Therefore, WIP is an effective mechanism to retain the normal proportion and mass of the tissue helping to repair multiple types of wounds.
Figure 2.

Wound-induced polyploidy is required to heal the adult fly epidermis. (A) An illustration of the adult Drosophila melanogaster abdomen and the area injured (boxed). (B) Immunofluorescent image of the ventral epidermis of the adult female fruit fly. The epidermis is organized into a monolayer of diploid cells organized into rows. For all images, epidermal nuclei are green and cell–cell FasIII septate junctions are in magenta. (C) Dramatic remodeling of the epidermis occurs after injury as a result of both the growth (via the endocycle) and fusion of the preexisting epidermal cells. As a consequence, a giant polyploid syncytium (outlined by the yellow dashed line) encompasses the area under the wound scab. Individual epidermal cells enlarge (arrowhead) and fuse into smaller syncytia (arrows) on the periphery. (D) Epidermal repair can be inhibited when polyploidy is blocked by perturbing both fusion and endoreplication simultaneously (reprinted by permission from Losick et al.1).
Figure 3.
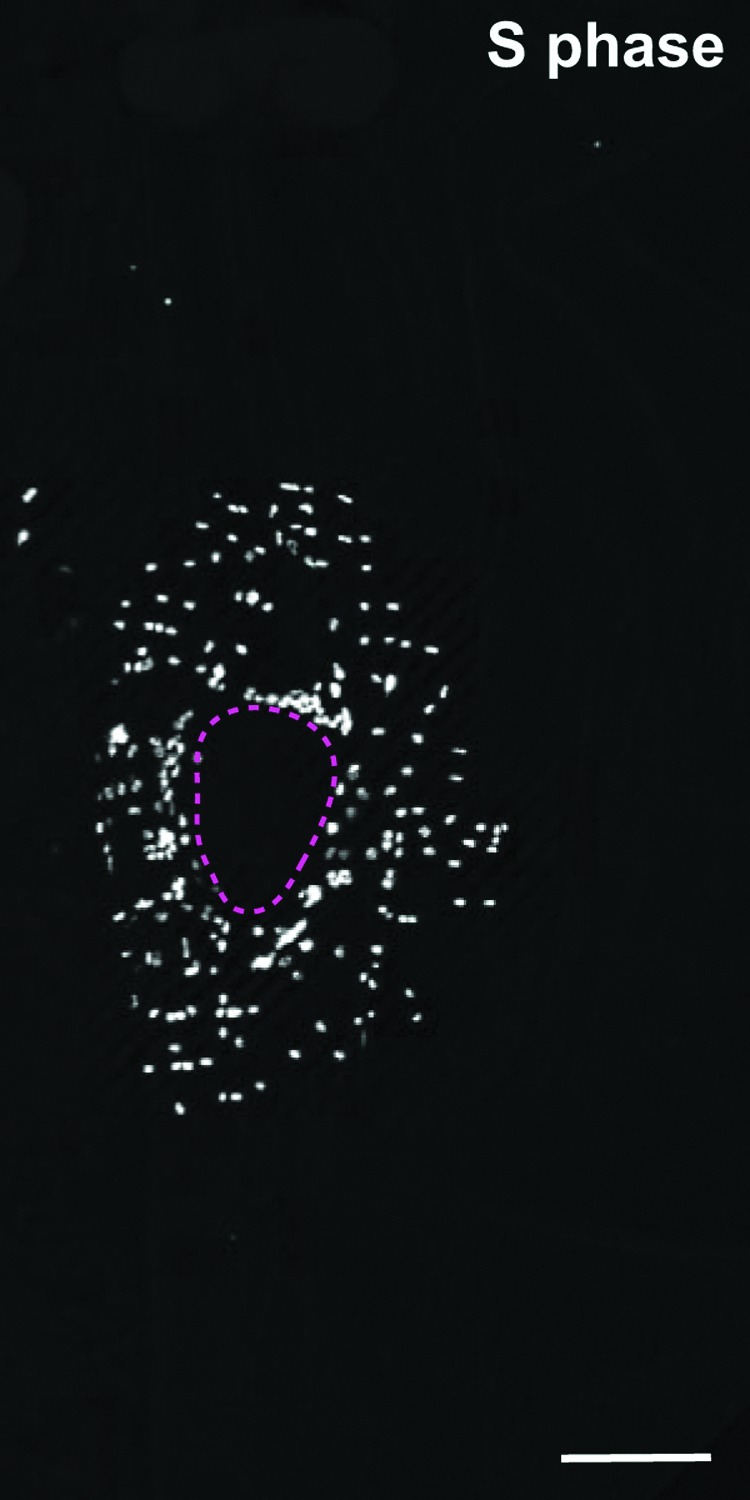
Quiescent epidermal cells in adult fly epidermis endoreplicate in response to injury. In response to injury, quiescent epidermal cells surrounding a puncture wound enter S phase and become ready labeled with the S phase marker EdU at 2 days postinjury. These EdU+cells grow instead of dividing by entering the endocycle. Shown is an immunofluorescent image of the wound zone with EdU+cells in white. The wound site is outlined by the magenta dashed line.
Cell fusion, while not commonly observed during wound healing, was first observed during larval epidermal repair in Drosophila.24 Small syncytia containing less than ten nuclei were observed to form during repair of pinch or puncture wounds,24 similar to the much larger syncytia containing fifty or more nuclei observed during repair of the adult epidermis.1 The simple organization of fly epidermis has made cell-to-cell fusion events more readily visible, while cell fusion has not yet been observed as a repair mechanism in mammalian tissues. Improved methods for accurately detecting cell fusion will help to determine whether cell fusion is a conserved mechanism to compensate for cell loss during wound healing.
An open question in the field is why cells would opt to produce polyploid cells by one mechanism versus another. A plausible explanation in the case of cell fusion is the speed of tissue repair. Indeed, blocking cell fusion by expressing a dominant negative Rac GTPase in the adult Drosophila epidermis delayed wound closure.1 Interestingly, it was observed in the adult fly epidermis that epidermal cells spontaneously fuse as cells turnover during normal aging.25 This demonstrates that cell fusion may be used to compensate for different types of tissue insults to preserve integrity.
Polyploidy compensates for cell loss during organ development
This study marks for the first time that polyploidy has been demonstrated to be essential to heal a wound1; however, replacing cells by increasing polyploidy is a theme that has been observed in several different developmental contexts in Drosophila. In these cases, enlarged cells do not necessarily mean that organ development is morphologically distorted. The mitotic kinase Cdc2 was inactivated within a clone in Drosophila's larval wing imaginal disc making a mosaic tissue lacking an essential mitotic gene.26 The cdc2 mutant cells endoreplicate instead of undergoing cell division. The developing imaginal disc and adult wing do develop to a normal sized organ, despite the tissue exhibiting fewer but larger polyploid cells.26 Another interesting example occurs in somatic follicle cells in Drosophila's ovary.2 Follicle cells are developmentally programmed to switch from a mitotic cell cycle to an endocycle.27 When the follicle cell number was interfered with by gene mutation or by inducing cell death during egg development, the wild-type follicle cells undergo additional endocycles in a process named compensatory cellular hypertrophy (CCH).2 The resulting egg chambers consisted of larger cells making up for the reduced cell number. Similar to what was observed in epidermal and hindgut pyloric WIP, the follicle cells appear to sense the change in organ volume and compensate by stimulating the surviving wild-type cells to further increase their DNA content from 16C up to 64C.2 These examples argue that both during developmental and in adult quiescent tissues, an intrinsic organ size control mechanism exists to sense when cell loss occurs and to induce cells to grow through either CCH or WIP to restore tissue homeostasis.
The organ size control pathway Hippo regulates WIP
The Hippo signal transduction pathway has been characterized to control the organ size in fruit flies and mammals. The Hippo pathway was originally discovered in Drosophila by mutants in hippo (hpo), warts (wts), and salvador (sav) that caused a tumor-like phenotype (excessive proliferation) in mutant imaginal discs.28–30 These components make up a negative feedback kinase cascade. In its on state, the kinase cascade phosphorylates the cotranscriptional activator, Yorkie (Yki), also known as Yap in mammals. Tissue injury has been shown to inactivate the kinase cascade allowing Yki to translocate to the nucleus to induce cell survival and proliferative gene expression.31,32 This review will be limited to a discussion of the role of the Hippo pathway in Drosophila adults. For additional information on the Hippo pathway in other cellular contexts, please refer to the recent reviews by Schroeder and Schroeder31 and Zhao et al.32
Upon Hippo inactivation, Yki is known to turn on the expression of a suite of genes, including dIAP and expanded (encoding an antiapoptotic and a FERM-domain protein, respectively).33 These Yki regulated genes are induced after injury and Yki has been shown to be required for intestinal stem cells to divide and regenerate damaged tissue.34–36 In the adult fly, epidermis injury also triggers the upregulation of Yki's signature genes, however, in contrast to intestinal stem cells, Yki is required for epidermal cells to become polyploid.1 This demonstrates that the Hippo pathway regulates tissue repair through either cell division or endoreplication depending on the cell type (Fig. 4). It is unclear why inactivation of the Hippo pathway during tissue repair results in different cellular outcomes; there could be differences in the upstream activation cues or downstream effects. One plausible explanation could be due to an intrinsic cellular difference. It is appreciated from studies in Drosophila that endocycling cells downregulate mitotic cyclins enabling them to bypass M phase of the cell cycle.37,38 Yki is known to induce expression of the S phase gene cyclin E,30 and hence, the expression of cyclin E in the absence of mitotic cyclins may promote an endocycle instead of the mitotic cell cycle. Stem cells or undifferentiated cells may continue to express mitotic cyclins, whereas differentiated postmitotic cells may have lost the ability to express these genes (Fig. 4). This suggests that cells may have the capacity to switch back to a proliferative repair strategy if the necessary mitotic factors were supplied.
Figure 4.

The Hippo pathway regulates tissue repair by cell division or endoreplication. The Hippo signal transduction pathway regulates the cotranscriptional activator Yorkie (Yki) to control tissue repair by cell division or endoreplication depending on the cell's differentiated state. In stem cells, Yki induces expression of the S phase cyclin E stimulating cells to divide. In differentiated cell types, this same signaling pathway appears to induce endoreplication by stimulating cells to grow instead of dividing. This model suggests that there are intrinsic differences between an undifferentiated and differentiated cell that trigger different repair outcomes using the same core Hippo signaling components.
Observations of polyploid cells in injured mammalian tissues
Polyploid cells are often observed in mammalian tissues in response to injury, stress, and/or aging, but their role in wound healing remains unclear. One of the best examples of polyploidization occurs in the liver where hepatocytes use multiple cellular mechanisms to repair and regenerate lost cells after tissue damage. Hepatocytes can enter a mitotic cell cycle and divide or grow through entry into an endocycle or endomitosis.5 When hepatocytes are genetically prevented from dividing in cdk1, trf2, or separase conditional null mutants, the tissue can still function normally by generating only polyploid cells.39–41 Remarkably, these livers can regrow to normal mass after injury and appear to function normally despite being composed entirely of enlarged polyploid hepatocytes.40 This result strongly argues that polyploidy is equally effective at restoring tissue function during regenerative processes. Alternatively, conditional null mutant mice for either e2f7 and/or e2F8 do not develop polyploid hepatocytes yet also regenerate normally by hepatocyte mitotic division.42,43 Therefore, polyploidy and cell division function appear to be acting redundantly; when one process is impeded the other compensates. These redundant functions may explain why the liver is one of the most robust regenerative organs in the human body.
Enhanced polyploidy has also been observed in cardiomyocytes of the mammalian heart after myocardial infarctions.4 Human and mouse cardiomyocytes in the adult heart are polyploid with either a single polyploid nucleus or a binucleated cell, respectively. A careful study measuring the DNA synthesis with multi-isotope imaging mass spectrometry determined that cardiomyocyte ploidy is enhanced post-injury.20 Most of the adult cardiomyocytes reenter S phase, but do not complete cell division resulting in an even more polyploid cell. It remains to be investigated how these observed changes in cardiomyocyte DNA ploidy affect the repair and healing of the mammalian heart.
Last, the mammalian corneal endothelium appears to respond to injury and aging by inducing endoreplication and cell fusion,3,44,45 similar to how the adult Drosophila epidermis responds. The corneal endothelium is a single postmitotic sheet of diploid cells that forms the barrier between the cornea stroma and inner eye chamber transporting solutes that maintain transparency of the cornea and hydration of the stromal layers.46 The observed changes in the cell size have been called “pathological,”3,44,45 yet these changes may be critical to maintain the endothelial layer. It is possible that the generation of polyploid endothelial cells could compensate for cell loss and promote wound healing similar to that observed in flies. In all of these examples, it still remains to be determined how these polyploid cells function in tissue repair and what would happen to wound healing if the generation of polyploid cells was perturbed.
The polyploid advantage
Researchers have been questioning for decades why some animal cells are polyploid. Polyploid cells have been discovered to be resistant to apoptosis in Drosophila tissues. When exposed to ionizing radiation, polyploid cells accumulate double-stranded DNA breaks, yet repress the DNA damage response checkpoint.47 In addition, genetically inducing an endocycle is sufficient to make a cell more resistant to apoptosis47 thereby giving polyploid cells a survival advantage.
Polyploidy has also long been correlated with an increased resistance to stress.18 Polyploid liver hepatocytes appear to adapt to metabolic stressors by having multiple copies of their genome. Polyploid liver hepatocytes, unlike most polyploid cells, readily divide sometimes resulting in aneuploid daughter cells.48 Hepatocyte aneuploidy is common in normal healthy livers, but is enhanced by injury resulting in the addition, loss, or breakage of chromosomes.48,49 As a result, karotype-specific hepatocytes emerged in the hereditary tyrosinemia mouse model, suggesting that polyploidy may give rise to genetic diversity and allow adaptive genomic selections under conditions of stress.49 In plants, researchers have also recently discovered advantages to polyploidy. Arabidopsis thaliana cells with higher ploidy are more resistant to stressful environmental conditions such as changes in salinity.50 It has been difficult for researchers to pinpoint the function of polyploidy in mammalian cells, but by further examining specific stresses or metabolic conditions, the polyploid advantage will be discovered.
Summary
The recent observations and results in D. melanogaster have proven that polyploidy can be beneficial to productive wound healing. There are many ways for cells to become polyploid and increase the cell size and depending on the cellular context different mechanisms appear to be used. Regardless, WIP allows tissues to compensate for lost or damaged cells when injury occurs without relying on cell division. The conserved organ size control pathway Hippo appears to function by sensing cell loss and by replacing tissue volume by either cell division or endoreplication depending on the cell type. What remains to be determined is whether there is an advantage to restoring the cell number by means of cell division or compensating by WIP during wound healing. Drosophila provides a simple model system to address this question and help to identify conserved mechanisms regulating the generation of polyploid cells during wound repair.
Take-Home Messages.
1. Polyploidy increases the cell size and DNA content
2. Tissue mass can be proportionally restored by polyploidization
3. WIP compensates for cell loss
4. The Hippo pathway regulates cell division or endoreplication depending on the cell's differentiated state
5. Drosophila can be used as a model to identify the molecular mechanisms of WIP
Abbreviations and Acronyms
- C
DNA content
- CCH
compensatory cellular hypertrophy
- Hpo
Hippo
- WIP
wound-induced polyploidy
- Wts
Warts
- Yki
Yorkie
Footnotes
Losick VP and Sprading AC: unpublished observations, Carnegie Institution for Science, 2009–2011.
Acknowledgments and Funding Sources
The author would like to apologize to her colleagues for the omission of their important contributions to the field. All the relevant work and their references could not be included in this short review due to space constraints. The author thanks Drs. Joyce Yang, Ming‐Chia Lee, and Allan Spradling for helpful discussions and critiques of this review article. This work was supported by the Howard Hughes Medical Institution and Carnegie Institution for Science in Baltimore.
Author Disclosure and Ghostwriting
No competing financial interests exist. This article was written by the author listed. No ghostwriters were used to write this article.
About the Authors
Vicki P. Losick, PhD, is a postdoctoral research associate with Dr. Allan C. Spradling in the Department of Embryology at Carnegie Institution for Science in Baltimore. She is a recipient of Jane Coffin Childs postdoctoral research fellowship for biomedical research. She received her PhD in Microbiology from Sackler School of Biomedical Sciences at Tufts University under the mentorship of Dr. Ralph R. Isberg. Her current research focuses on the molecular mechanisms of wound‐induced polyploidy and wound healing in Drosophila adults.
References
- 1.Losick VP, Fox DT, Spradling AC. Polyploidization and cell fusion contribute to wound healing in the adult Drosophila epithelium. Curr Biol 2013;23:2224–2232 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Tamori Y, Deng WM. Tissue repair through cell competition and compensatory cellular hypertrophy in postmitotic epithelia. Dev Cell 2013;25:350–363 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ikebe H, Takamatsu T, Itoi M, Fujita S. Changes in nuclear DNA content and cell size of injured human corneal endothelium. Exp Eye Res 1988;47:205–215 [DOI] [PubMed] [Google Scholar]
- 4.Laflamme MA, Murry CE. Heart regeneration. Nature 2011;473:326–335 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Miyaoka Y, Ebato K, Kato H, Arakawa S, Shimizu S, Miyajima A. Hypertrophy and unconventional cell division of hepatocytes underlie liver regeneration. Curr Biol 2012;22:1166–1175 [DOI] [PubMed] [Google Scholar]
- 6.Brack AS, Rando TA. Tissue-specific stem cells: lessons from the skeletal muscle satellite cell. Cell Stem Cell 2012;10:504–514 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Clevers H. The intestinal crypt, a prototype stem cell compartment. Cell 2013;154:274–284 [DOI] [PubMed] [Google Scholar]
- 8.Plikus MV, Gay DL, Treffeisen E, Wang A, Supapannachart RJ, Cotsarelis G. Epithelial stem cells and implications for wound repair. Semin Cell Dev Biol 2012;23:946–953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Razzell W, Wood W, Martin P. Swatting flies: modelling wound healing and inflammation in Drosophila. Dis Model Mech 2011;4:569–574 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Worley MI, Setiawan L, Hariharan IK. Regeneration and transdetermination in Drosophila imaginal discs. Annu Rev Genet 2012;46:289–310 [DOI] [PubMed] [Google Scholar]
- 11.Ohlstein B, Spradling A. The adult Drosophila posterior midgut is maintained by pluripotent stem cells. Nature 2006;439:470–474 [DOI] [PubMed] [Google Scholar]
- 12.Micchelli CA, Perrimon N. Evidence that stem cells reside in the adult Drosophila midgut epithelium. Nature 2006;439:475–479 [DOI] [PubMed] [Google Scholar]
- 13.Lucchetta EM, Ohlstein B. The Drosophila midgut: a model for stem cell driven tissue regeneration. Wiley Interdiscip Rev Dev Biol 2012;1:781–788 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Losick VP, Morris LX, Fox DT, Spradling A. Drosophila stem cell niches: a decade of discovery suggests a unified view of stem cell regulation. Dev Cell 2011;21:159–171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Su TT, O'Farrell PH. Size control: cell proliferation does not equal growth. Curr Biol 1998;8:R687–689 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Miyazaki M, Esser KA. Cellular mechanisms regulating protein synthesis and skeletal muscle hypertrophy in animals. J Appl Physiol 2009;106:1367–1373 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Aoyagi T, Matsui T. Phosphoinositide-3 kinase signaling in cardiac hypertrophy and heart failure. Curr Pharm Des 2011;17:1818–1824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fox DT, Duronio RJ. Endoreplication and polyploidy: insights into development and disease. Development 2013;140:3–12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kato K, Awasaki T, Ito K. Neuronal programmed cell death induces glial cell division in the adult Drosophila brain. Development 2009;136:51–59 [DOI] [PubMed] [Google Scholar]
- 20.Senyo SE, Steinhauser ML, Pizzimenti CL, et al. Mammalian heart renewal by pre-existing cardiomyocytes. Nature 2013;493:433–436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fox DT, Spradling AC. The Drosophila hindgut lacks constitutively active adult stem cells but proliferates in response to tissue damage. Cell Stem Cell 2009;5:290–297 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Smith-Bolton RK, Worley MI, Kanda H, Hariharan IK. Regenerative growth in Drosophila imaginal discs is regulated by Wingless and Myc. Dev Cell 2009;16:797–809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ramet M, Lanot R, Zachary D, Manfruelli P. JNK signaling pathway is required for efficient wound healing in Drosophila. Dev Biol 2002;241:145–156 [DOI] [PubMed] [Google Scholar]
- 24.Galko MJ, Krasnow MA. Cellular and genetic analysis of wound healing in Drosophila larvae. PLoS Biol 2004;2:E239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Scherfer C, Han VC, Wang Y, Anderson AE, Galko MJ. Autophagy drives epidermal deterioration in Drosophila model of tissue aging. Aging (Albany NY) 2013;5:276–287 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Weigmann K, Cohen SM, Lehner CF. Cell cycle progression, growth and patterning in imaginal discs despite inhibition of cell division after inactivation of Drosophila Cdc2 kinase. Development 1997;124:3555–3563 [DOI] [PubMed] [Google Scholar]
- 27.Klusza S, Deng WM. At the crossroads of differentiation and proliferation: precise control of cell-cycle changes by multiple signaling pathways in Drosophila follicle cells. Bioessays 2011;33:124–134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Harvey KF, Pfleger CM, Hariharan IK. The Drosophila Mst ortholog, hippo, restricts growth and cell proliferation and promotes apoptosis. Cell 2003;114:457–467 [DOI] [PubMed] [Google Scholar]
- 29.Tapon N, Harvey KF, Bell DW, et al. salvador promotes both cell cycle exit and apoptosis in Drosophila and is mutated in human cancer cell lines. Cell 2002;110:467–478 [DOI] [PubMed] [Google Scholar]
- 30.Huang J, Wu S, Barrera J, Matthews K, Pan D. The Hippo signaling pathway coordinately regulates cell proliferation and apoptosis by inactivating Yorkie, the Drosophila Homolog of YAP. Cell 2005;122:421–434 [DOI] [PubMed] [Google Scholar]
- 31.Schroeder MC, Halder G. Regulation of the Hippo pathway by cell architecture and mechanical signals. Semin Cell Dev Biol 2012;23:803–811 [DOI] [PubMed] [Google Scholar]
- 32.Zhao B, Tumaneng K, Guan KL. The Hippo pathway in organ size control, tissue regeneration and stem cell self-renewal. Nat Cell Biol 2011;13:877–883 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wu S, Huang J, Dong J, Pan D. hippo encodes a Ste-20 family protein kinase that restricts cell proliferation and promotes apoptosis in conjunction with salvador and warts. Cell 2003;114:445–456 [DOI] [PubMed] [Google Scholar]
- 34.Karpowicz P, Perez J, Perrimon N. The Hippo tumor suppressor pathway regulates intestinal stem cell regeneration. Development 2010;137:4135–4145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ren F, Wang B, Yue T, Yun EY, Ip YT, Jiang J. Hippo signaling regulates Drosophila intestine stem cell proliferation through multiple pathways. Proc Natl Acad Sci U S A 2010;107:21064–21069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Shaw RL, Kohlmaier A, Polesello C, Veelken C, Edgar BA, Tapon N. The Hippo pathway regulates intestinal stem cell proliferation during Drosophila adult midgut regeneration. Development 2010;137:4147–4158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Maqbool SB, Mehrotra S, Kolpakas A, et al. Dampened activity of E2F1-DP and Myb-MuvB transcription factors in Drosophila endocycling cells. J Cell Sci 2010;123:4095–4106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Zielke N, Kim KJ, Tran V, et al. Control of Drosophila endocycles by E2F and CRL4(CDT2). Nature 2011;480:123–127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Diril MK, Ratnacaram CK, Padmakumar VC, et al. Cyclin-dependent kinase 1 (Cdk1) is essential for cell division and suppression of DNA re-replication but not for liver regeneration. Proc Natl Acad Sci U S A 2012;109:3826–3831 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lazzerini Denchi E, Celli G, de Lange T. Hepatocytes with extensive telomere deprotection and fusion remain viable and regenerate liver mass through endoreduplication. Genes Dev 2006;20:2648–2653 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Wirth KG, Wutz G, Kudo NR, et al. Separase: a universal trigger for sister chromatid disjunction but not chromosome cycle progression. J Cell Biol 2006;172:847–860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chen HZ, Ouseph MM, Li J, et al. Canonical and atypical E2Fs regulate the mammalian endocycle. Nat Cell Biol 2012;14:1192–1202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Pandit SK, Westendorp B, Nantasanti S, et al. E2F8 is essential for polyploidization in mammalian cells. Nat Cell Biol 2012;14:1181–1191 [DOI] [PubMed] [Google Scholar]
- 44.Ikebe H, Takamatsu T, Itoi M, Fujita S. Age-dependent changes in nuclear DNA content and cell size of presumably normal human corneal endothelium. Exp Eye Res 1986;43:251–258 [DOI] [PubMed] [Google Scholar]
- 45.Roh DS, Du Y, Gabriele ML, Robinson AR, Niedernhofer LJ, Funderburgh JL. Age-related dystrophic changes in corneal endothelium from DNA repair-deficient mice. Aging Cell 2013;12:1122–1131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Joyce NC. Proliferative capacity of the corneal endothelium. Prog Retin Eye Res 2003;22:359–389 [DOI] [PubMed] [Google Scholar]
- 47.Hassel C, Zhang B, Dixon M, Calvi BR. Induction of endocycles represses apoptosis independently of differentiation and predisposes cells to genome instability. Development 2014;141:112–123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Duncan AW, Taylor MH, Hickey RD, et al. The ploidy conveyor of mature hepatocytes as a source of genetic variation. Nature 2010;467:707–710 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Duncan AW, Hanlon Newell AE, Bi W, et al. Aneuploidy as a mechanism of stress-induced liver adaption. J Clin Invest 2012;122:3307–3315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Chao DY, Dilkes B, Luo H, et al. Polyploids exhibit higher potassium uptake and salinity tolerance in Arabidopsis. Science 2013;341:658–659 [DOI] [PMC free article] [PubMed] [Google Scholar]