

Abstract
Human sapovirus has been shown to be one of the most important etiologies in pediatric patients with acute diarrhea. However, very limited data are available about the causative roles and epidemiology of sapovirus in community settings. A nested matched case-control study within a birth cohort study of acute diarrhea in a peri-urban community in Peru from 2007 to 2010 was conducted to investigate the attributable fraction (AF) and genetic diversity of sapovirus. By quantitative reverse transcription–real-time PCR (qPCR) sapovirus was detected in 12.4% (37/299) of diarrheal and 5.7% (17/300) of nondiarrheal stools (P = 0.004). The sapovirus AF (7.1%) was higher in the second year (13.2%) than in the first year (1.4%) of life of children. Ten known genotypes and one novel cluster (n = 5) within four genogroups (GI, GII, GIV, and GV) were identified by phylogenetic analysis of a partial VP1 gene. Further sequence analysis of the full VP1 gene revealed a possible novel genotype, tentatively named GII.8. Notably, symptomatic reinfections with different genotypes within the same (n = 3) or different (n = 5) genogroups were observed in eight children. Sapovirus exhibited a high attributable burden for acute gastroenteritis, especially in the second year of life, of children in a Peruvian community. Further large-scale studies are needed to understand better the global burden, genetic diversity, and repeated infections of sapovirus.
INTRODUCTION
Acute diarrhea is one of the most important causes of morbidity and mortality in pediatric populations, especially in developing countries. Rotavirus, norovirus, and other viruses are common causative etiological agents, and rotavirus accounts for about 440,000 child deaths annually (1), while norovirus is a leading cause of epidemic and sporadic acute diarrhea (2). Currently a rotavirus vaccination program has been implemented in 80 countries as a part of national immunization programs (http://sites.path.org/rotavirusvaccine/rotavirus-vaccines/#global-intro). It successfully reduced the number of hospitalizations and deaths due to acute gastroenteritis (3, 4) and is cost-effective (5). Norovirus has now replaced rotavirus as the leading cause of medically attended acute diarrhea in pediatric populations (6, 7), and sapovirus, belonging to a separate genus of the Caliciviridae family, has been reported as the second most commonly detected virus after norovirus in children with acute diarrhea where rotavirus vaccination was implemented (8, 9). In addition, reports on sapovirus outbreaks across all age groups have increased in South Asia, Europe, and North America recently (10–14).
The genome of sapovirus consists of a positive-sense, single-stranded RNA with two open reading frames (ORFs) (12). ORF1 encodes the nonstructural proteins and a major capsid protein, VP1, and ORF2 encodes a protein whose function is still unknown (12). Like for norovirus, multiple genetic clusters of human sapovirus have been reported, including four genogroups (GI, GII, GIV, and GV) with 17 genotypes (GI.1 to GI.7, GII.1 to GII.7, GIV.1, and GV.1 and GV.2) (12). Mild acute diarrhea and severe and even fatal sapovirus infections have been documented (15, 16). Viral shedding lasts about 2 weeks after onset of symptoms (17, 18). Serological studies have identified that sapovirus infections are very common during early childhood; however, epidemiology and protective immunity in community settings have rarely been investigated. Sapovirus was identified as a significant viral etiological agent in children in limited etiological studies of acute diarrhea, which targeted multiple pathogens (19–21). To understand the etiological role and genetic diversity of sapovirus in acute diarrhea, we conducted a nested matched case-control study by analyzing selected samples from a birth cohort study of acute diarrhea in a Peruvian peri-urban community conducted between June 2007 and May 2010 (22).
MATERIALS AND METHODS
Stool sample selection.
Stool samples collected during or 1 day after diarrhea episodes (n = 862) were considered diarrheal samples in this study. We stratified the samples previously tested for norovirus (22) by eight age groups (0 to 2, 3 to 5, 6 to 8, 9 to 11, 12 to 14, 15 to 17, 18 to 20, and 21 to 23 months) and four seasons (December to February, March to May, June to August, and September to November). We assumed that sapovirus should be detected in at least 5% of diarrheal episodes in children under 2 years, based on detection rates of 5.4 to 16.6% by quantitative real-time PCR (qPCR) reported in two population-based studies (9, 21). To ensure that at least 1 positive sample was detected in each age group, at least 20 samples were tested per group. Finally, 300 diarrheal stools (26 to 44 stools per age group) representing 34.2% of 862 episodes were randomly selected by using Stata 13 (StataCorp, College Station, TX, USA). One nondiarrheal control specimen per diarrheal stool was selected at random, matched by age group and season using Stata 13. Nondiarrheal stools were defined as stools which were not collected within the diarrhea episode, within a week before the first day of diarrhea episodes, or within 2 weeks after the last day of diarrhea episodes, based on the sapovirus incubation period (12) and duration of virus shedding (17, 18). Both diarrheal and nondiarrheal samples were selected from the samples previously tested for norovirus and were included regardless of norovirus test results. Among 600 randomly selected samples, three samples (all negative for norovirus) were not available in the specimen bank, and two of them were replaced with other samples in the same age-season category; therefore, 599 samples were included in the analysis.
Quantitative reverse transcription–real-time PCR (qPCR) for sapovirus.
Viral nucleic acid was extracted from 140 μl of a 10% stool suspension using QIAamp viral RNA minikits (Qiagen, Hilden, Germany). cDNA was synthesized by using Moloney murine leukemia virus (M-MLV) reverse transcriptase kits with random primers (Life Technologies, Carlsbad, CA, USA). qPCR targeting the polymerase-VP1 junction of sapovirus was performed on an ABI 7500 Fast real-time PCR system (Life Technologies), using previously described primers and probes (23). The detection limit of qPCR, which was determined by standard curves generated with 10-fold serial dilutions (107 to 10 copies/μl) of sapovirus GII.1 standard plasmid, was 100 copies/μl of cDNA, corresponding to a cycle threshold (CT) value of 38 at an amplification of total 45 cycles. A sample with a CT value of <38 was considered to be positive for sapovirus, and three positive controls and one negative control were included in each assay.
Genotyping and sequence analysis.
Sapovirus-positive samples were further amplified by hemi-nested PCR targeting a partial VP1 gene (720 nucleotides [nt]; corresponding to nt 5159 to 5878 of strain Manchester [accession number X86560]) with the external primers SV F13/F14 (24) and SaV 5857R-1/-2 and internal primers SaV F1245 (23) and SaV 5857R-1/-2 (see Table S1 in the supplemental material). Cycling conditions for the first and second rounds of hemi-nested PCR were 94°C for 5 min, 35 cycles of 94°C for 30 s, 50°C for 30 s, and 72°C for 1 min, and 1 cycle of 72°C for 10 min. For the possible new genotype of sapovirus, the full VP1 gene was amplified with primer pair SV F13/F14 and SaV GIIR under same cycling conditions as for hemi-nested PCR. PCR products were sequenced using BigDye Terminator chemistries V1.1 (Life Technologies) in a 3730 genetic analyzer (Life Technologies). The resulting sequences were BLAST verified. Genogroups and genotypes were characterized by phylogenetic analysis of a partial VP1 gene (606 nt; corresponding to nt 5236 to 5841 of strain Manchester [accession no. X86560]) (12) using the maximum-likelihood method (Kimura 2-parameter model) in MEGA5 (25). In addition, pairwise distances were calculated using the Kimura 2-parameter method in MEGA5, and the cutoff values to designate the same sapovirus genotypes and genogroups were ≤0.169 and ≤0.488, respectively (12, 26).
Statistical analysis.
The unadjusted attributable fraction (AF) for estimating the sapovirus burden of diarrhea was calculated by using the formula (27) AF = Pr(sapovirus/diarrhea)[1 − (1/OR)], where Pr(sapovirus/diarrhea) is the proportion of sapovirus in diarrhea stools and OR is the odds ratio of sapovirus for diarrhea and nondiarrhea. The median CT values for diarrheal and nondiarrheal stools were compared by using the Mann-Whitney U test with Stata 13 (StataCorp).
Ethics statement.
Mothers provided written informed consent, and the study was approved by the Ethics Committees of the Tohoku University Graduate School of Medicine, Asociación Benéfica PRISMA, Universidad Peruana Cayetano Heredia (UPCH), Johns Hopkins University, the Centers for Disease Control and Prevention (CDC), and the European Union.
Nucleotide sequence accession numbers.
All unique sequences determined in this study were deposited in GenBank (accession no. KT276516 to KT276558, KU886206 to KU886207, and KT306742).
RESULTS
Three hundred nondiarrheal and 299 diarrheal stools (one diarrheal stool in the age group 18 to 20 months was not available) were tested for sapovirus. Sapovirus was detected in 9.0% (54/599) of stools, with 12.4% (37/299) in diarrheal stools and 5.7% (17/300) in nondiarrheal stools (P = 0.004) (Fig. 1). In qPCR, the median CT values were significantly lower in diarrheal than in nondiarrheal samples (24 versus 29; P = 0.024). Among the 37 sapovirus-associated episodes, the median duration of diarrhea was 3 days (interquartile range [IQR], 2 to 5 days). The median maximum daily number of diarrheal stools was four (IQR, 3.5 to 5.5). Fifteen episodes (40.5%) had vomiting, and only four episodes (10.8%) had fever. Rates of detection of sapovirus in diarrheal stools were higher in children aged 12 to 23 months (18.6%; 27/145) than in children aged <12 months (6.5%; 10/154) (P = 0.002), and those in nondiarrheal stools were similar in the two age groups (5.2% versus 6.2%). The sapovirus AF (7.1%) was higher in the second (13.2%) than in the first (1.4%) year of life. Sapovirus was not detected in diarrheal stools of children aged 0 to 5 months (Fig. 1), although only 68 diarrheal samples from 194 episodes were tested for this age group. Sapovirus was detected together with norovirus in four diarrheal stools in the age group of 12 to 23 months.
FIG 1.
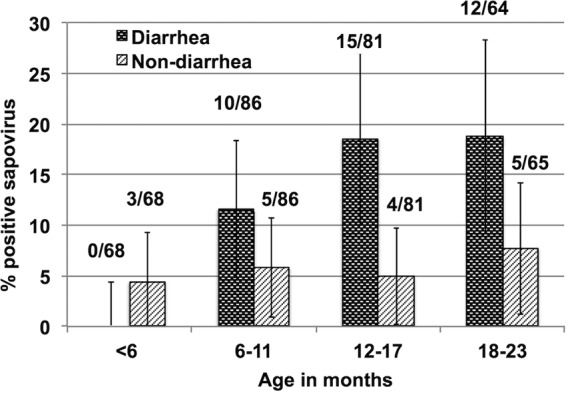
Prevalence of sapovirus in diarrheal and nondiarrheal stool samples by age in the case-control study in Peru. The y axis indicates the percentage of stools in which sapovirus was detected. The x axis shows age groups of infants. The black bars and gray bars indicate diarrheal and nondiarrheal stools, respectively. Error bars represent 95% confidence intervals. One diarrheal sample was not available in the age group of 18 to 23 months.
Of 54 qPCR-positive specimens, 45 specimens (83.3%) from 2007 to 2010 were successfully sequenced. Genogroup I (n = 21) and genogroup II (n = 16) were detected more frequently than genogroup IV (n = 3) and genogroup V (n = 5). Among these genogroups, 10 known genotypes (GI.1, GI.2, GI.6, GI.7, GII.1, GII.2 GII.4, GII.5, GIV, and GV.1) were identified (Fig. 2). Most genotypes were detected in both diarrheal and nondiarrheal stools except for two genotypes (GII.5 and GIV), which were found only in one and three diarrheal stools, respectively. Interestingly, five Peruvian sequences together with strains detected in the United States (HM59058) and Taiwan (KM092511) fell into a distinct cluster in GII with less than 78% similarity with other genotypes of GII based on the partial VP1 gene. We further sequenced the full VP1 gene of one diarrhea strain in this cluster and found that this Peruvian VP1 together with another full VP1 from Taiwan (KM092511) also formed a cluster distinct from other GII genotypes in the phylogenetic tree of nucleotide sequences (see Fig. S1 in the supplemental material) and amino acid sequences (Fig. 3). Moreover, the pairwise nucleotide distance of Peruvian VP1 from other GII.1 to GII.7 genotypes (0.298 to 0.468) was larger than 0.169. Therefore, this cluster could be tentatively classified as a novel genotype, and the tentative genotype GII.8 is proposed.
FIG 2.

Phylogenetic analysis of sapovirus based on a partial VP1 region (606 nt; corresponding to nt 5236 to 5841 of strain Manchester [accession no. X86560]) The phylogenetic tree was inferred using the maximum-likelihood method (Kimura 2-parameter model) with MEGA5. The tree is drawn to scale, with branch lengths in the units of the number of base substitutions per site. The percentage of trees (≥70%) in which the associated taxa clustered together is shown next to the branches. Peru strains are marked with triangles and are named as country-number tested-diarrheal (D) or nondiarrheal (ND).
FIG 3.

Phylogenetic analysis of full VP1 amino acid sequences of sapovirus. The phylogenetic tree was inferred using the maximum-likelihood method (general reverse transcriptase plus frequency model) with MEGA5. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. The VP1 obtained in this study is marked with a triangle.
Eight children had two sapovirus infections detected within short time (with the interval of two infections ranging from 20 days to 19 months) (Table 1). Five of these children had repeated infections with a virus belonging to a different sapovirus genogroup, and three children had repeated infections with a different genotype of the same genogroup. All first and second infections of the eight children were symptomatic, except in one child where the first infection with GII.1 was asymptomatic and the second with GII.2 was symptomatic. None of the repeated infections of sapovirus had concurrent norovirus infection. The median CT values of both the first (21; IQR, 17.5 to 25.5) and second (21; IQR, 15.8 to 27.6) infections were relatively low, suggesting that both infections had relatively large viral loads. In addition, one child was considered to have shed sapovirus at 25 days after the onset of diarrhea, since one diarrheal sample (Peru-406-D; CT = 24.2) and one nondiarrheal sample (Peru-426-ND; CT = 33.5) collected from the same child were found to contain the same GII.4 strain (Fig. 2).
TABLE 1.
Characteristics of repeated infections of sapovirus observed in eight children in the birth cohort study in Peru
| Infant ID | Sampling date (day-mo-yr) | Age (mo) | CT value | Sapovirus genotype | Accession no. | Symptom(s) | Duration of symptoms (days) |
|---|---|---|---|---|---|---|---|
| PX046 | 13-July-08 | 13 | 16.0 | GI.6 | KT276525 | Diarrhea, vomiting | 2 |
| 16-Nov.-08 | 17 | 15.5 | GI.1 | KT672521 | Diarrhea | 3 | |
| PX152 | 19-Jan.-09 | 16 | 26.8 | GI.6 | KT276526 | Diarrhea | 5 |
| 10-Feb.-09 | 16 | 31.9 | GI.7 | KT276536 | Diarrhea, vomiting | 6 | |
| PX198 | 12-May-08 | 4 | 23.5 | GII.1 | KU886207 | None | 0 |
| 15-Dec.-09 | 23 | 31.4 | GII.2 | KT276555 | Diarrhea | 2 | |
| PX080 | 25-Aug.-08 | 12 | 21.5 | GIV | KT276558 | Diarrhea, vomiting | 2 |
| 25-May-09 | 21 | 21.1 | GII.2 | KT276554 | Diarrhea | 2 | |
| PX135 | 17-Mar.-08 | 6 | 34.4 | GIV | KU886206a | Diarrhea, fever | 12 |
| 19-June-09 | 21 | 15.9 | GV.1 | KT276538 | Diarrhea, vomiting | 2 | |
| PX159 | 19-June-08 | 9 | 16 | GII.4 | KT276549 | Diarrhea, vomiting | 5 |
| 27-Jan.-09 | 16 | 15.7 | GI.1 | KT276520 | Diarrhea | 4 | |
| PX212 | 20-Apr.-09 | 15 | 18.9 | GI.1 | KT276522 | Diarrhea, fever | 3 |
| 30-Oct.-09 | 21 | 23.9 | GII.8 | KT276546 | Diarrhea | 3 | |
| PX287 | 18-June-09 | 8 | 25.8 | GIV | KT276557 | Diarrhea, vomiting | 6 |
| 12-Oct.-09 | 11 | 19.4 | GI.1 | KT276519 | Diarrhea, vomiting | 3 |
This sequence has 450 bp and is not shown in Fig. 2.
DISCUSSION
We determined the positive rate and genotype distribution of sapovirus in a birth cohort study in Peru. Sapovirus had a considerable attributable burden of 7.1% for acute diarrhea in children under 2 years old in this community. In sapovirus-infected children less than 2 years old, vomiting was frequently reported, while fever was rarely reported. The median duration of sapovirus diarrhea observed in this study was similar to the previously reported duration of 3 to 5 days (8, 17). Compared to other etiological studies of sapovirus, the overall rate of detection of sapovirus in symptomatic children (12.4%) in this study was similar to those in the United Kingdom (11.1%) (19) and Nicaragua (16.6%) (9) but higher than those in the Netherlands (7.8%) (20) and the United States (5.4%) (21). The rate of detection of sapovirus in asymptomatic children observed in this study (5.7%) was similar to that in the United States (4.2%) (21) but higher than those in the Netherlands (1.8%) (20), the United Kingdom (2.4%) (19), and Nicaragua (1.9%) (9). Our results are comparable to recent reports from other continents, in which sapovirus was commonly detected in pediatric gastroenteritis (8, 28–31). Possible reasons of increased rates of detection of sapovirus include the emerging genotypes (10, 29, 31), the implementation of rotavirus vaccination (9), and application of more sensitive molecular diagnostic assays (32, 33). Limited data on sapovirus in South America are available (34), and more studies are needed to confirm clinical and public health significance of sapovirus in this region.
Our study found that sapovirus is more likely to be associated with acute diarrhea in the second year of life in children, as described for norovirus in the same cohort (22). This finding was supported by several recent reports, in which sapovirus was detected more commonly in symptomatic children aged 12 to 23 months than in those aged <1 year (9, 21, 35, 36). Lower detection rates of sapovirus in children aged <1 year possibly reflect a protective effect of breastfeeding and/or transferred maternal antibodies (22, 37), as supported by the fact that we did not detect sapovirus in symptomatic children under 6 months.
In this study, 10 known genotypes and one novel cluster were detected, and none of them appeared to be predominant, which differed from norovirus GII.4, having been predominant in the same cohort (22) or worldwide (38) the for past 2 decades. The emerging genotypes GI.2 and GIV, which caused outbreaks in various countries recently (10, 11, 14, 15), were detected, but not as dominant genotypes, in this Peruvian community. Some uncommon genotypes of sapovirus, such as GI.6, GI.7, GII.4, and GII.5, were observed. Five strains detected from 2008 to 2009 formed a distinct cluster and appear to belong to a novel genotype. BLAST searches from GenBank showed similar sequences deposited from South Africa (2010 and 2013), the United States (2010), China (2011), and Taiwan (2014). It is noteworthy that the strains in the United States and Taiwan were associated with outbreaks of diarrhea, suggesting their wide distribution and pathogenicity. Currently, two complete VP1 sequences for this possible novel genotype are available from two countries (Peru and Taiwan), and it is tentatively proposed as GII.8, following both phylogenetic analysis of amino acid sequence (39) and pairwise distance classification proposed by Oka et al. (12).
The cohort study design allowed us to identify repeated sapovirus infections in same child. Five children had symptomatic reinfections with different genogroups, and a similar observation was reported in two patients in Japan (40). Also, the occurrence of natural reinfections in pigs by different porcine sapovirus genogroups has been recently reported (41). In addition, we found that three children experienced a second diarrheal episode caused by a different sapovirus genotype in a period of less than 1 month to 19 months. In the same cohort, children were found to commonly experience multiple diarrheal episodes with different norovirus GII genotypes, and therefore it was speculated that children can develop genotype-specific immunity of norovirus with only a modest level of cross-protection even within the same genogroup (22). In contrast, very little information is available about protective immunity against sapovirus infection. Sera collected from naturally infected humans or immunized animals with recombinant VP1 protein of sapovirus were shown to be moderately cross-reactive against the heterologous genotypes and weakly cross-reactive with heterologous genogroups of sapovirus (42–44). Large-scale studies are needed to investigate whether protective immunity for sapovirus is genotype specific or not, since the number of children with reinfections in this study was small due to a limited sample size.
One of the limitations of this study was that samples were not tested for enteric pathogens other than norovirus; therefore, coinfections of sapovirus with other pathogens was not analyzed. The attribution of sapovirus in acute diarrhea should be validated in future studies including other major enteric pathogens. Also, a small proportion of symptomatic cases may shed sapovirus longer than 2 weeks after onset of illness, as seen in the current study and other studies (17, 18); therefore, the prevalence of sapovirus in asymptomatic children in this study could be slightly overestimated, and the viral shedding pattern needs to be furthered explored.
In conclusion, recent epidemiological reports have highlighted the importance of sapovirus in acute diarrhea, especially after rotavirus vaccine implementation, but the etiological role and epidemiology of sapovirus remain to be clarified. We found that sapovirus exhibited a high attributable burden of acute diarrhea, especially in the second year of life, in a peri-urban community in Peru. Cocirculation of multiple genotypes with a possible novel genotype and symptomatic reinfection with different genotypes within the same or different genogroups were observed. Further large-scale studies are needed to define the global burden, incidence, and protective immunity of sapovirus infection.
Supplementary Material
ACKNOWLEDGMENTS
We thank the community of San Juan de Miraflores for their long-term collaboration and the field staff for their hard work. We acknowledge K. Tohma and I. Khandaker for their technical assistance.
The findings and conclusions in this report are those of the authors and do not necessarily represent the views of the Centers for Disease Control and Prevention.
All authors report no potential conflict of interest.
Footnotes
Supplemental material for this article may be found at http://dx.doi.org/10.1128/JCM.03133-15.
REFERENCES
- 1.Parashar UD, Hummelman EG, Bresee JS, Miller MA, Glass RI. 2003. Global illness and deaths caused by rotavirus disease in children. Emerg Infect Dis 9:565–572. doi: 10.3201/eid0905.020562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Patel MM, Hall AJ, Vinje J, Parashar UD. 2009. Noroviruses: a comprehensive review. J Clin Virol 44:1–8. doi: 10.1016/j.jcv.2008.10.009. [DOI] [PubMed] [Google Scholar]
- 3.Payne DC, Boom JA, Staat MA, Edwards KM, Szilagyi PG, Klein EJ, Selvarangan R, Azimi PH, Harrison C, Moffatt M, Johnston SH, Sahni LC, Baker CJ, Rench MA, Donauer S, McNeal M, Chappell J, Weinberg GA, Tasslimi A, Tate JE, Wikswo M, Curns AT, Sulemana I, Mijatovic-Rustempasic S, Esona MD, Bowen MD, Gentsch JR, Parashar UD. 2013. Effectiveness of pentavalent and monovalent rotavirus vaccines in concurrent use among US children <5 years of age, 2009-2011. Clin Infect Dis 57:13–20. doi: 10.1093/cid/cit164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.do Carmo GM, Yen C, Cortes J, Siqueira AA, de Oliveira WK, Cortez-Escalante JJ, Lopman B, Flannery B, de Oliveira LH, Carmo EH, Patel M. 2011. Decline in diarrhea mortality and admissions after routine childhood rotavirus immunization in Brazil: a time-series analysis. PLoS Med 8:e1001024. doi: 10.1371/journal.pmed.1001024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Paternina-Caicedo A, De la Hoz-Restrepo F, Alvis-Guzman N. 2015. Epidemiological and economic impact of monovalent and pentavalent rotavirus vaccines in low and middle income countries: a cost-effectiveness modeling analysis. Pediatr Infect Dis J 34:e176–184. doi: 10.1097/INF.0000000000000727. [DOI] [PubMed] [Google Scholar]
- 6.Payne DC, Vinje J, Szilagyi PG, Edwards KM, Staat MA, Weinberg GA, Hall CB, Chappell J, Bernstein DI, Curns AT, Wikswo M, Shirley SH, Hall AJ, Lopman B, Parashar UD. 2013. Norovirus and medically attended gastroenteritis in U.S. children. N Engl J Med 368:1121–1130. doi: 10.1056/NEJMsa1206589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hemming M, Rasanen S, Huhti L, Paloniemi M, Salminen M, Vesikari T. 2013. Major reduction of rotavirus, but not norovirus, gastroenteritis in children seen in hospital after the introduction of RotaTeq vaccine into the National Immunization Programme in Finland. Eur J Pediatr doi: 10.1007/s00431-013-1945-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Pang XL, Zeng SQ, Honma S, Nakata S, Vesikari T. 2001. Effect of rotavirus vaccine on Sapporo virus gastroenteritis in Finnish infants. Pediatr Infect Dis J 20:295–300. doi: 10.1097/00006454-200103000-00015. [DOI] [PubMed] [Google Scholar]
- 9.Becker-Dreps S, Bucardo F, Vilchez S, Zambrana LE, Liu L, Weber DJ, Pena R, Barclay L, Vinje J, Hudgens MG, Nordgren J, Svensson L, Morgan DR, Espinoza F, Paniagua M. 2014. Etiology of childhood diarrhea after rotavirus vaccine introduction: a prospective, population-based study in Nicaragua. Pediatr Infect Dis J 33:1156–1163. doi: 10.1097/INF.0000000000000427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Svraka S, Vennema H, van der Veer B, Hedlund KO, Thorhagen M, Siebenga J, Duizer E, Koopmans M. 2010. Epidemiology and genotype analysis of emerging sapovirus-associated infections across Europe. J Clin Microbiol 48:2191–2198. doi: 10.1128/JCM.02427-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Pang XL, Lee BE, Tyrrell GJ, Preiksaitis JK. 2009. Epidemiology and genotype analysis of sapovirus associated with gastroenteritis outbreaks in Alberta, Canada: 2004-2007. J Infect Dis 199:547–551. doi: 10.1086/596210. [DOI] [PubMed] [Google Scholar]
- 12.Oka T, Wang Q, Katayama K, Saif LJ. 2015. Comprehensive review of human sapoviruses. Clin Microbiol Rev 28:32–53. doi: 10.1128/CMR.00011-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Iritani N, Kaida A, Abe N, Kubo H, Sekiguchi J, Yamamoto SP, Goto K, Tanaka T, Noda M. 2014. Detection and genetic characterization of human enteric viruses in oyster-associated gastroenteritis outbreaks between 2001 and 2012 in Osaka City, Japan. J Med Virol 86:2019–2025. doi: 10.1002/jmv.23883. [DOI] [PubMed] [Google Scholar]
- 14.Wu FT, Oka T, Takeda N, Katayama K, Hansman GS, Muo CH, Liang SY, Hung CH, Dah-Shyong Jiang D, Hsin Chang J, Yang JY, Wu HS, Yang CF. 2008. Acute gastroenteritis caused by GI/2 sapovirus, Taiwan, 2007. Emerg Infect Dis 14:1169–1171. doi: 10.3201/eid1407.071531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lee LE, Cebelinski EA, Fuller C, Keene WE, Smith K, Vinje J, Besser JM. 2012. Sapovirus outbreaks in long-term care facilities, Oregon and Minnesota, USA, 2002-2009. Emerg Infect Dis 18:873–876. doi: 10.3201/eid1805.111843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liu X, Yamamoto D, Saito M, Imagawa T, Ablola A, Tandoc AO III, Segubre-Mercado E, Lupisan SP, Okamoto M, Furuse Y, Saito M, Oshitani H. 2015. Molecular detection and characterization of sapovirus in hospitalized children with acute gastroenteritis in the Philippines. J Clin Virol 68:83–88. doi: 10.1016/j.jcv.2015.05.001. [DOI] [PubMed] [Google Scholar]
- 17.Rockx B, De Wit M, Vennema H, Vinje J, De Bruin E, Van Duynhoven Y, Koopmans M. 2002. Natural history of human calicivirus infection: a prospective cohort study. Clin Infect Dis 35:246–253. doi: 10.1086/341408. [DOI] [PubMed] [Google Scholar]
- 18.Iwakiri A, Ganmyo H, Yamamoto S, Otao K, Mikasa M, Kizoe S, Katayama K, Wakita T, Takeda N, Oka T. 2009. Quantitative analysis of fecal sapovirus shedding: identification of nucleotide substitutions in the capsid protein during prolonged excretion. Arch Virol 154:689–693. doi: 10.1007/s00705-009-0358-0. [DOI] [PubMed] [Google Scholar]
- 19.Amar CF, East CL, Gray J, Iturriza-Gomara M, Maclure EA, McLauchlin J. 2007. Detection by PCR of eight groups of enteric pathogens in 4,627 faecal samples: re-examination of the English case-control Infectious Intestinal Disease Study (1993-1996). Eur J Clin Microbiol Infect Dis 26:311–323. doi: 10.1007/s10096-007-0290-8. [DOI] [PubMed] [Google Scholar]
- 20.de Wit MA, Koopmans MP, Kortbeek LM, Wannet WJ, Vinje J, van Leusden F, Bartelds AI, van Duynhoven YT. 2001. Sensor, a population-based cohort study on gastroenteritis in the Netherlands: incidence and etiology. Am J Epidemiol 154:666–674. doi: 10.1093/aje/154.7.666. [DOI] [PubMed] [Google Scholar]
- 21.Chhabra P, Payne DC, Szilagyi PG, Edwards KM, Staat MA, Shirley SH, Wikswo M, Nix WA, Lu X, Parashar UD, Vinje J. 2013. Etiology of viral gastroenteritis in children <5 years of age in the United States, 2008-2009. J Infect Dis 208:790–800. doi: 10.1093/infdis/jit254. [DOI] [PubMed] [Google Scholar]
- 22.Saito M, Goel-Apaza S, Espetia S, Velasquez D, Cabrera L, Loli S, Crabtree JE, Black RE, Kosek M, Checkley W, Zimic M, Bern C, Cama V, Gilman RH, Norovirus Working Group in Peru. 2014. Multiple norovirus infections in a birth cohort in a Peruvian periurban community. Clin Infect Dis 58:483–491. doi: 10.1093/cid/cit763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Oka T, Katayama K, Hansman GS, Kageyama T, Ogawa S, Wu FT, White PA, Takeda N. 2006. Detection of human sapovirus by real-time reverse transcription-polymerase chain reaction. J Med Virol 78:1347–1353. doi: 10.1002/jmv.20699. [DOI] [PubMed] [Google Scholar]
- 24.Okada M, Yamashita Y, Oseto M, Shinozaki K. 2006. The detection of human sapoviruses with universal and genogroup-specific primers. Arch Virol 151:2503–2509. doi: 10.1007/s00705-006-0820-1. [DOI] [PubMed] [Google Scholar]
- 25.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28:2731–2739. doi: 10.1093/molbev/msr121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Oka T, Mori K, Iritani N, Harada S, Ueki Y, Iizuka S, Mise K, Murakami K, Wakita T, Katayama K. 2012. Human sapovirus classification based on complete capsid nucleotide sequences. Arch Virol 157:349–352. doi: 10.1007/s00705-011-1161-2. [DOI] [PubMed] [Google Scholar]
- 27.Blackwelder WC, Biswas K, Wu Y, Kotloff KL, Farag TH, Nasrin D, Graubard BI, Sommerfelt H, Levine MM. 2012. Statistical methods in the Global Enteric Multicenter Study (GEMS). Clin Infect Dis 55(Suppl 4):S246–S253. doi: 10.1093/cid/cis788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Johnsen CK, Midgley S, Bottiger B. 2009. Genetic diversity of sapovirus infections in Danish children 2005-2007. J Clin Virol 46:265–269. doi: 10.1016/j.jcv.2009.07.008. [DOI] [PubMed] [Google Scholar]
- 29.Harada S, Okada M, Yahiro S, Nishimura K, Matsuo S, Miyasaka J, Nakashima R, Shimada Y, Ueno T, Ikezawa S, Shinozaki K, Katayama K, Wakita T, Takeda N, Oka T. 2009. Surveillance of pathogens in outpatients with gastroenteritis and characterization of sapovirus strains between 2002 and 2007 in Kumamoto Prefecture, Japan. J Med Virol 81:1117–1127. doi: 10.1002/jmv.21454. [DOI] [PubMed] [Google Scholar]
- 30.Matussek A, Dienus O, Djeneba O, Simpore J, Nitiema L, Nordgren J. 2015. Molecular characterization and genetic susceptibility of sapovirus in children with diarrhea in Burkina Faso. Infect Genet Evol 32:396–400. doi: 10.1016/j.meegid.2015.03.039. [DOI] [PubMed] [Google Scholar]
- 31.Iritani N, Yamamoto SP, Abe N, Kubo H, Oka T, Kaida A. 2015. Epidemics of GI.2 sapovirus in gastroenteritis outbreaks during 2012-2013 in Osaka City, Japan. J Med Virol doi: 10.1002/jmv.24451. [DOI] [PubMed] [Google Scholar]
- 32.Tam CC, O'Brien SJ, Tompkins DS, Bolton FJ, Berry L, Dodds J, Choudhury D, Halstead F, Iturriza-Gomara M, Mather K, Rait G, Ridge A, Rodrigues LC, Wain J, Wood B, Gray JJ. 2012. Changes in causes of acute gastroenteritis in the United Kingdom over 15 years: microbiologic findings from 2 prospective, population-based studies of infectious intestinal disease. Clin Infect Dis 54:1275–1286. doi: 10.1093/cid/cis028. [DOI] [PubMed] [Google Scholar]
- 33.Chan MC, Sung JJ, Lam RK, Chan PK, Lai RW, Leung WK. 2006. Sapovirus detection by quantitative real-time RT-PCR in clinical stool specimens. J Virol Methods 134:146–153. doi: 10.1016/j.jviromet.2005.12.013. [DOI] [PubMed] [Google Scholar]
- 34.Marques Mendanha de Oliveira D, Souza M, Souza Fiaccadori F, Cesar Pereira Santos H, das Dores de Paula Cardoso D. 2014. Monitoring of calicivirus among day-care children: evidence of asymptomatic viral excretion and first report of GI.7 norovirus and GI.3 sapovirus in Brazil. J Med Virol 86:1569–1575. doi: 10.1002/jmv.23791. [DOI] [PubMed] [Google Scholar]
- 35.Pang XL, Preiksaitis JK, Lee BE. 2013. Enhanced enteric virus detection in sporadic gastroenteritis using a multi-target real-time PCR panel: a one-year study. J Med Virol doi: 10.1002/jmv.23851. [DOI] [PubMed] [Google Scholar]
- 36.Thongprachum A, Takanashi S, Kalesaran AFC, Okitsu S, Mizuguchi M, Hayakawa S, Ushijima H. 2015. Four-year study of viruses that cause diarrhea in Japanese pediatric outpatients. J Med Virol 87:1141–1148. doi: 10.1002/jmv.24155. [DOI] [PubMed] [Google Scholar]
- 37.Makita K, Hayakawa Y, Okame M, Homma K, Phan TG, Okitsu S, Ushijima H. 2007. First detection of IgA against norovirus in breast milk. Clin Lab 53:125–128. [PubMed] [Google Scholar]
- 38.Vinje J. 2015. Advances in laboratory methods for detection and typing of norovirus. J Clin Microbiol 53:373–381. doi: 10.1128/JCM.01535-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kroneman A, Vega E, Vennema H, Vinje J, White PA, Hansman G, Green K, Martella V, Katayama K, Koopmans M. 2013. Proposal for a unified norovirus nomenclature and genotyping. Arch Virol 158:2059–2068. doi: 10.1007/s00705-013-1708-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Harada S, Oka T, Tokuoka E, Kiyota N, Nishimura K, Shimada Y, Ueno T, Ikezawa S, Wakita T, Wang Q, Saif LJ, Katayama K. 2012. A confirmation of sapovirus re-infection gastroenteritis cases with different genogroups and genetic shifts in the evolving sapovirus genotypes, 2002-2011. Arch Virol 157:1999–2003. doi: 10.1007/s00705-012-1387-7. [DOI] [PubMed] [Google Scholar]
- 41.Lauritsen KT, Hansen MS, Johnsen CK, Jungersen G, Bottiger B. 2015. Repeated examination of natural sapovirus infections in pig litters raised under experimental conditions. Acta Vet Scand 57:60. doi: 10.1186/s13028-015-0146-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hansman GS, Oka T, Sakon N, Takeda N. 2007. Antigenic diversity of human sapoviruses. Emerg Infect Dis 13:1519–1525. doi: 10.3201/eid1310.070402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Farkas T, Deng X, Ruiz-Palacios G, Morrow A, Jiang X. 2006. Development of an enzyme immunoassay for detection of sapovirus-specific antibodies and its application in a study of seroprevalence in children. J Clin Microbiol 44:3674–3679. doi: 10.1128/JCM.01087-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Jiang X, Cubitt WD, Berke T, Zhong W, Dai X, Nakata S, Pickering LK, Matson DO. 1997. Sapporo-like human caliciviruses are genetically and antigenically diverse. Arch Virol 142:1813–1827. doi: 10.1007/s007050050199. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.