

Abstract
Endurance performance depends on the integration of several phenotypic traits influenced by multiple environmental and genetic factors. Objectives of the study were: (1) to examine the genotypic frequencies of the ACE I/D, ACTN3 R577X polymorphisms and endurance performance-related phenotypes, (2) to evaluate the dynamics of endurance performance parameters during a 5-year period in relation to ACE I/D and ACTN3 R577X genotypes in Estonian young skiers. Determination of VO2peak was performed in 58 skiers aged 15-19 years (41 males, 17 females) during a 5-year period. The control group consisted of 322 healthy non-athletic subjects (145 males, 177 females). The study groups were genotyped for the ACE I/D and ACTN3 R577X variants. Frequencies of the ACE ID and ACTN3 RR genotypes were significantly higher (p = 0.047 and p = 0.003, respectively) and the RX genotype was lower (p = 0.008) in young male skiers compared with controls. A significant relationship was found between change (Δ) of training volume and ΔVO2peak (mL·kg-1·min-1) (r = 0.475, p = 0.002). No significant main effect was detected between VO2peak (mL·kg-1·min-1) dynamics (comparison with the previous age group data) and ACE I/D and ACTN3 R577X genotypes interactions (F = 0.571, p = 0.770 and F = 0.650 and p = 0.705, respectively) in all young skiers. Study results indicated a significantly higher frequency of the ACE ID and ACTN3 RR genotypes among Estonian young male skiers compared with the male control group. Significant genotype-related differences in dynamics of VO2peak during a 5-year period were not found. In the future, longitudinal research including different gene variants may contribute to a better understanding of the nature of endurance performance.
Key points.
Significantly higher prevalence of the ACE ID and the ACTN3 RR genotypes were found among Estonian young male skiers compared with the male control group, which may be an advantage for the explosive speed and power capacity in race skiing.
A more remarkable trend of increase in VO2peak (mL·kg-1·min-1) during the 5-year period was observed among male skiers with the ACTN3 XX genotype and among female skiers with the ACE ID genotype.
No significant genotype-related associations in the dynamics of VO2peak were found during the 5-year period.
Key words: Genes polymorphisms, endurance capacity, skiing, longitudinal study
Introduction
Genetic predisposition has a substantial role in the development of athletic physical performance and is characterized by a large number of genes polymorphisms while the influence of multiple environmental factors is essential on gene expression (MacArthur and North, 2005). Endurance performance depends mainly on the integration of cardiorespiratory fitness, metabolic processes in skeletal muscles and neurological factors, but biomechanical, psychological, physical, nutritional and other parameters are important as well (Joyner and Coyle, 2008; Rusko, 2003). The maximal oxygen uptake (VO2max) is recognized as one of the major quantitative traits in sports physiology and the most common parameter to demonstrate training effects. VO2max is influenced by different environmental and genetic factors (Rankinen et al., 2000). The maximal heritability for VO2max (adjusted for age, sex and body mass) in the sedentary cohort of the HERITAGE Family Study was observed to be up to 50% (Bouchard et al., 1998). On average, women’s VO2max values are 10% lower compared to men due to higher body fat mass and lower levels of haemoglobin (Pate et al., 1987; Durstine et al., 1987). During the growth and maturation period in 11-17-year-old humans, the increase of VO2peak is affected primarily by body lean mass (Armstrong and Welsman, 2001).
The angiotensin-converting enzyme gene insertion/deletion (ACE I/D) polymorphism (rs4340) and the α-actinin-3 gene (ACTN3) R577X polymorphism (rs1815739) are two of the most extensively studied genes in recent years in association with human physical performance. As part of the renin-angiotensin system, ACE (angiotensin-converting enzyme) has an essential role in the regulation of cardiovascular function and metabolic processes in muscles. ACE catalyses the conversion of angiotensin I to physiologically active angiotensin II, which causes vasoconstriction and through the release of aldosterone influences the regulation of electrolytes and water. In addition, ACE reduces the vasodilatory effect of bradykinin causing an increase in blood pressure (Thompson and Binder-Macleod, 2006). The functional polymorphism of human ACE I/D is related to the presence (insertion, I allele) or absence (deletion, D allele) of a 287-base-pair (bp) segment in intron 16. Alvarez et al. (2000) have reported that the activity of ACE enzyme in the DD homozygotes is 3-4 times higher than in the II homozygotes. The I allele has been associated with improved endurance performance and the D allele with higher ACE activity and enhanced strength and sprint performance (Myerson et al., 1999; Thompson and Binder-Macleod, 2006). On the contrary to the previous researches, there are studies not showing the correlation between the ACE I/D polymorphism and enhanced physical performance (Rankinen et al., 2000; Taylor et al., 1999).
Another widely investigated gene influencing athletic performance is the α-actinin-3 (ACTN3). The ACTN3 gene encodes an actinin-binding protein in the sarcomeric Z line of fast-twitch muscle fibres of skeletal muscle (North et al., 1999; Mills et al., 2001). A common genetic variation in the ACTN3 gene is the replacement of the arginine (R) with a stop codon (X) at amino acid 577 (C→T transition at position 1747 in exon 16) (North et al., 1999). The R577X polymorphism of the human ACTN3 gene results in two variants of alleles: a functional R allele and a non-functional X allele (North et al., 1999; Mills et al., 2001). Yang et al. (2003) detected a significant association between the ACTN3 R577X polymorphism and athletic performance. They found significantly higher frequencies of the R allele in elite sprint athletes than in controls. The non-functional X allele, which leads to the α-actinin-3 deficiency, supports better endurance ability. Eynon et al. (2012) reported that the ACTN3 XX genotype had a higher frequency in endurance athletes, and it was more prevalent in world-class endurance athletes compared to national level athletes. On the contrary, in the Genathlete study on Caucasian male elite endurance athletes conducted by Döring et al. (2010), no association between endurance performance and the ACTN3 R577X polymorphism was found. Furthermore, a study on Russian endurance athletes revealed that the α-actinin-3 deficiency may have a negative effect on performance in endurance athletes (Ahmetov et al., 2010). Ahmetov et al. (2010) disclosed that the XX genotype was significantly underrepresented among Russian endurance athletes compared with the control group. They pointed out that in addition to endurance capability, power and speed components are required as well to be successful in different endurance-oriented sports events.
Meta-analysis, which analyses relationships between the ACE I/D, the ACTN3 R577X genotypes and athletic performance conducted by Ma et al. (2013), revealed significant associations of the ACE II genotype with enhanced endurance performance and the ACTN3 R allele with better power performance.
Altogether, numerous previous studies have associated the ACE I/D and ACTN3 R577X polymorphisms with enhanced physical performance, although outcomes have been contradictory across investigations. Secondly, there are relatively few longitudinal studies observing the association of dynamics in training-induced changes in endurance performance with gene polymorphisms.
The primary aim of the present study was to examine the prevalence of the ACE I/D and ACTN3 R577X genotypes in Estonian young skiers compared to a control group. The secondary purpose was to evaluate the dynamics of endurance performance parameters during a 5-year period in relation to the ACE I/D and ACTN3 R577X genotypes in Estonian young skiers.
Methods
Participants
Participants in the present study were young cross-country and biathlon skiers (hereafter young skiers). The total number of subjects was 58 (41 males and 17 females). 52% of young skiers were current or former Estonian junior national team members. The database of the electronic records (eHL) of Tartu University Hospital from the period 2002-2013 was used. All study participants had five consecutive pre-participation health evaluations (PHE) between the ages 15 to 19 years. Study inclusion criteria for young skiers were regular specific cross-country skiing training during the observed age period, and the PHE had to include training characteristics and an exercise test on a treadmill determining the maximal oxygen uptake peak (VO2peak). The control group consisted of 322 healthy sedentary subjects without previous athletic training (145 males and 177 females). Male controls’ mean age was 33.9 ± 13.2 years (height 1.81 ± 0.07 m, weight 80.0 ± 13.0 kg) and female controls’ mean age was 37.1 ± 13.6 years (height 1.67 ± 0.06 m, weight 65.2 ± 11.0 kg). The ethnic origin of all participants was Caucasian.
Research Ethics Committee of the University of Tartu (protocol No. 196/M-30, and No. 207/M-9) approved the study protocol. Written informed consent was obtained from each participant.
Anthropometric measurements
Subjects’ height (m) was measured using a metal anthropometer (± 0.1 cm) and body mass (kg) was determined by using clinical scales (± 0.05 kg). The body mass index (BMI) was calculated.
Cardiopulmonary exercise test
All incremental continuous exercise tests were performed on the treadmill (Telineyhtymä, Kotka, Finland). A standardized stress test protocol for cross-country skiers increasing the treadmill inclination and speed was used. All tests were performed to volitional exhaustion to obtain the parameters of maximal aerobic endurance. It has been found that not all individuals achieve the maximum oxygen consumption plateau during maximal exertion (Bassett and Howley, 2000). In this case, the highest level of oxygen uptake (VO2peak) attained in an exercise test to exhaustion should be evaluated. VO2peak is considered to be an indicator in assessing the aerobic capacity of children and adolescents (Armstrong and Welsman, 2001). VO2peak (absolute, L · min-1 and relative, mL·kg-1·min-1) was determined by the direct breath-by-breath method using a gas analyzer (Oxycon Pro version 5.2, Hoechberg, Germany). The absolute value of VO2peak is influenced by age-related overall growth in body size and increase in body mass (Bongers et al., 2014). To exclude this main phenomenon, we used the relative value of VO2peak (mL·kg-1·min-1) to assess the dynamics of VO2peak related to the training capacity and investigated genes variants. In addition, the duration of the standardized exercise test (min) and maximal heart rate (HRmax, beats·min-1) were recorded, and exercise tolerance (W·kg-1) was calculated. Heart rate was recorded by using a sport-tester (Polar Electro, Finland). Gas analysis data were automatically calculated for every 30-second period. VO2peak was considered as the highest VO2 rate achieved within 30 seconds at the end of the exercise test. The criteria for maximal exercise test were the following: (1) respiratory exchange ratio (RER) equalled at least 1.1; (2) HRmax > 90% of the age-predicted maximum calculated according to the formula 220 – age (Davis, 2006).
The change in VO2peak (ΔVO2peak; absolute, L·min-1; relative mL·kg-1·min-1) was obtained by calculating the differences of VO2peak between 15 and 19 year-old age groups of female and male skiers. The change in exercise test duration (Δ duration of exercise test, min) and the training volume per week (Δ training hours, h · week1) were calculated similarly. Besides, the dynamics of VO2peak, duration of exercise test and training volume were observed in each age group separately compared with the previous age group.
Genotyping
Peripheral venous blood samples for DNA extraction were collected from all study subjects and controls during the period 2010-2013. The DNA was extracted from peripheral venous blood samples in 9 ml EDTA containing vacuettes by using the standard salting-out method (Miller et al., 1988). The EDTA tubes were stored at -202C until DNA extraction. Isolated DNA was dissolved in Tris-EDTA (TE) buffer. The purity and concentrations of the DNA were measured by a spectrophotometer (NanoDrop, ND-1000). The DNA samples were stored at -808C until usage. Genotyping of rs4343 (ACE, C_11942562_20) and rs1815739 (ACTN3, C_590093_1) was carried out by using the TaqMan® SNP Genotyping Assay (Applied Biosystems, Foster City, CA, USA), which is a multiplex endpoint assay that detects variants of a single nucleic acid sequence. PCR reactions were run on the ABI 7900 instrument (Applied Biosystems, Foster City, CA, USA) by using the following cycling parameters: after the first step at 95lC for 10 min, 40 cycles of denaturation at 92CC for 15 s and extension at 60CC for 1 min. Genomic DNA (20ng//l) was amplified in a total volume of 5ll containing 1x Amplification Master Mix (Applied Biosystems, Foster City, CA, USA) and 1x probe. Genotypes were analysed by using the allelic discrimination function of the system.
Statistical analysis
Statistical analysis was performed using the Statistical Package for Social Sciences (SPSS version 22.0 for WINDOWS). Descriptive data are presented as means (x), standard deviations (SD) and percentages (%). The chi-square (χ2-test) test was used to determine significant differences between the frequencies of the ACE I/D and the ACTN3 R577X genotypes. The dynamics of VO2peak (mL·kg-1·min-1) and the duration of the exercise test (min) as the change between five time points were analysed with repeated analysis of variance (dynamics as a factor of five levels). Two-factor ANOVA was used to determine the interaction between VO2peak (mL·kg-1·min-1) dynamics (five levels), the ACE I/D genotypes (three levels) and the ACTN3 R577X genotypes (three levels), respectively. The Pearson’s correlation coefficient (r) was applied to determine the relationships between variables. P values of ≤ 0.05 were set as statistically significant.
Results
Anthropometrical, physical performance and training characteristics of young skiers by age and sex are presented in Table 1. Among young male skiers, a remarkable increase of the training volume (h · week1) was observed at the age of 16 years (t = -3.059, p = 0.007). In female skiers, no significant changes in the dynamics in training hours per week were found up to 18 years, whereas a significant increase was observed at the age of 19 (t = -2.882, p = 0.024). A significant improvement in the duration of the standardized exercise test performance (min) was detected in the 16- and 17-year age groups (t = -2.317, p = 0.030; t = -2.508, p = 0.018, respectively) among young male skiers and in young female skiers of 18 years in comparison with the previous age group (t = -3.977, p = 0.002). A statistically significant improvement of the absolute VO2peak was found in males at the age of 16 (t = -3.849, p = 0.001), 17 (t = -3.232, p = 0.003) and 19 (t = -2.503, p = 0.020) compared with the previous age group. However, after correcting the differences for body size, the dynamics of relative VO2peak was not significant in comparison across all age groups in young male skiers. We found a significant positive trend of absolute VO2peak as well as relative VO2peak in young female skiers only in the 17-year-old age group compared with the previous age group (t = -3.535, p = 0.008 and t = -2.386, p = 0.044, respectively).
Table 1.
Characteristics of young skiers by age and sex groups. All data are presented as means (±SD).
| Age group (yrs) | 15 years | 16 years | 17 years | 18 years | 19 years | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gender | Male | Female | Male | Female | Male | Female | Male | Female | Male | Female |
| Weight (kg) | 63.2 (8.0) | 58.4 (8.6) | 67.2 (8.4)c | 60.2 (6.8)b | 69.7 (7.8)c | 60.6 (5.2)a | 71.4 (7.6)c | 62.3 (4.4) | 72.5 (8.2)c | 61.8 (5.0) |
| Height (m) | 1.77 (.06) | 1.67 (.04) | 1.80 (.06)c | 1.70 (.04)c | 1.81 (.06)c | 1.69 (.04) | 1.82 (5.5)b | 1.71 (.04) | 1.82 (.06) | 1.69 (.04) |
| BMI (kg·m-2) | 20.2 (1.9) | 20.8 (2.3) | 20.8 (2.0) | 21.0 (1.9) | 21.3 (1.8) | 21.3 (1.6) | 21.5 (1.6) | 21.4 (1.5) | 21.9 (1.6) | 21.6 (1.3) |
| TrVol (h·week-1) | 8.5 (2.5) | 7.0 (1.4) | 9.9 (2.7)b | 7.6 (2.4) | 11.3 (2.9) | 9.3 (2.1) | 12.4 (2.9) | 10.9 (2.9) | 13.4 (2.9) | 13.2 (3.1)a |
| ET duration (min) | 24.6 (5.1) | 20.5 (3.4) | 27.0 (2.2)a | 21.1 (2.8) | 27.9 (2.0)a | 21.9 (2.4) | 28.3 (2.2) | 23.4 (1.7)b | 29.3 (1.6) | 24.6 (2.6) |
| VO2peak (L·min-1) | 4.0 (.6) | 3.0 (.3) | 4.4 (.5)c | 3.1 (.1) | 4.7 (.6)b | 3.4 (.3)b | 4.8 (.7) | 3.4 (.3) | 5.0 (.5)a | 3.5 (.4) |
|
VO2peak (mL·min-1·kg-1) |
64.1 (6.1) | 53.9 (5.9) | 66.1 (4.7) | 53.0 (5.2) | 67.0 (4.5) | 55.4 (5.5)a | 67.6 (5.7) | 54.9 (5.0) | 68.1 (5.0) | 57.1 (5.2) |
a p ≤ 0.05
b p ≤ 0.01
c p ≤ 0.001 compared to the previous age group; TrVol = training volume; ET = exercise test
Changes in performance parameters in young female and male skiers between the 15 and 19 year-old groups are shown in Table 2.
Table 2.
Changes in performance parameters (Δ) between 15 and 19 year-old age groups of female and male skiers. All data are presented as means (±SD).
| Young female skiers (n=17) | Young male skiers (n=41) | |
|---|---|---|
| Δ TrVol (h·week-1) | 6.0 (3.6) | 5.1 (3.5) |
| Δ ET duration (min) | 3.5 (4.2) | 3.1 (2.9) |
| Δ VO2peak (L·min-1) | .5 (.4) | .9 (.6) |
| Δ VO2peak (mL·kg-1·min-1) | 3.6 (4.6) | 4.0 (5.2) |
ET = exercise test; TrVol = training volume
There were no significant differences in the distribution of the ACE I/D genotypes between young female skiers and controls although the prevalence of the ACE II genotype compared with controls was higher, 39% and 23%, respectively (t = 2.177, p = 0.140) and the ACE ID genotype was lower, 33% and 50%, respectively (t = 1.757, p = 0.185). However, the frequency of the ACE ID genotype in young male skiers compared with controls was statistically higher (t = 3.940, p = 0.047). There was a non-significant lower trend of the ACE DD genotype among young male skiers compared to the male control group (t = 3.465, p = 0.063) (Figure 1).
Figure 1.
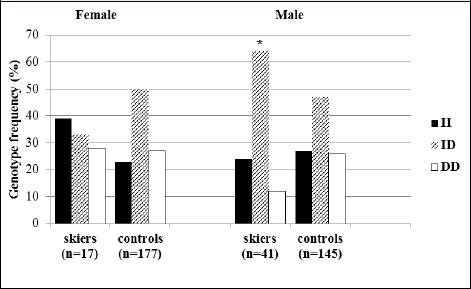
Genotypic frequencies of the ACE I/D polymorphism (rs4340) in young skiers and non-athletic controls. * p < 0.05 in comparison with controls (χ2-test).
In the distribution of the ACTN3 genotypes among young female skiers, there were no subjects carrying the ACTN3 XX genotype in comparison with the group of female controls (n = 13; 7.4%). Although there was a trend towards a higher frequency of the ACTN3 RR genotype in female skiers (33.3%) in comparison with female controls (15.3%), the difference remained statistically non-significant (t = 3.691, p = 0.055). When analysing the results of the distribution of the ACTN3 R577X genotypes frequencies in male skiers compared with male controls, the frequency of the RR genotype was statistically higher (t = 9.045, p = 0.003) and the frequency of the RX genotype was lower (t = 7.128, p = 0.008) (Figure 2).
Figure 2.
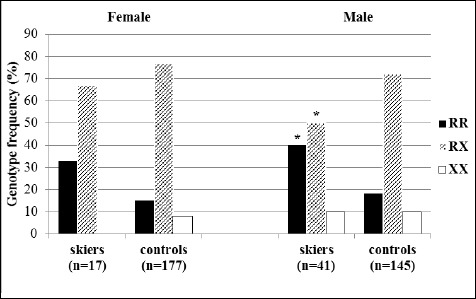
Genotypic frequencies of the ACTN3 R577X polymorphism (rs1815739) in young skiers and non-athletic controls. * p < 0.05 in comparison with controls (χ2-test).
We did not detect any statistically significant relationships between the ACE I/D, the ACTN3 R577X genotypes and the ΔVO2peak (Table 3). However, a considerable increase of the ΔVO2peak was observed in female skiers with the ACE ID genotype (6.2 ± 5.7 mL·kg-1·min-1, n = 5) and in male skiers with the ACTN3 XX genotype (7.6 ± 4.8 mL·kg-1·min-1, n = 4), the increase was statistically nonsignificant.
Table 3.
VO2peak changes (Δ VO2peak, mL · kg-1 · min-1) according to the ACE I/D and the ACTN3 R577X genotypes between 15 and 19 year-old age groups of female (n =17) and male (n = 41) skiers. All data are presented as means (± SD).
| Young female skiers Δ VO2 peak |
Young male skiers Δ VO2 peak |
|
|---|---|---|
| ACE I/D genotypes | ||
| II | 2.0 (2.4) (n = 7) | 5.3 (5.5) (n = 10) |
| ID | 6.2 (5.7) (n = 5) | 3.2 (5.0) (n = 26) |
| DD | 3.3 (5.5) (n = 5) | 5.0 (6.2) (n = 5) |
| ACTN3 R577X genotypes | ||
| RR | 3.7 (2.6) (n = 5) | 3.8 (4.8) (n = 16) |
| RX | 3.7 (5.3) (n = 12) | 3.4 (5.5) (n = 21) |
| XX | - | 7.6 (4.8) (n = 4) |
The correlation analysis showed a statistically significant relationship between the Δ training volume and the ΔVO2peak (mL·kg-1·min-1) among young male skiers (r = 0.475, p = 0.002), but there was no significant relationship between these variables among female skiers (r = 0.331, p = 0.247).
According to the two-factor ANOVA analysis, a significant main effect of dynamics of the VO2peak (mL·kg-1·min-1) was detected in the total study group (F = 9.592, p < 0.001), as well as in males (F = 6.933, p < 0.001) and females (F = 5.741, p < 0.001). No significant effect was detected between the dynamics of the VO2peak (mL·kg-1·min-1) and the ACE I/D genotypes interaction (F = 0.571, p = 0.770) or between the dynamics of the VO2peak (mL·kg-1·min-1) and the ACTN3 R577X interaction (F = 0.650, p = 0.705) in all young skiers. There were no gender differences in all measured interactions.
Discussion
The results of our study revealed a significantly higher prevalence of the ACE ID and the ACTN3 RR genotypes and lower ACTN3 RX genotypes in male skiers in comparison with controls. A trend of a lower frequency of the ACE DD was observed among young male skiers compared with male controls.
Competitive cross-country skiing belongs to the group of one of the most demanding sports events. In the classification of sports activities by Mitchell et al. (2005), which is based on the type and intensity of exercise performed, cross-country skiing is characterized as a sport with high dynamic and up to moderate static (depending on the technique of skiing) capacities. Regardless of the very different length of race distances − from sprint distances of 1.5 km up to 50 km − all distances require high endurance performance (Rusko, 2003). It has been found in experimental studies that depending on the terrain profile, the proportion of aerobic energy production in longer distances can reach up to 90-99%, while during sprint ski-racing it may be 50% on average (Rusko, 2003).
Folland et al. (2000) associated a larger gain in muscle strength and hypertrophy after resistance training with the D allele of the ACE gene in their study. Muscle strength, especially upper body muscle strength plays an important role in addition to endurance capacity in competitive cross-country skiing (Alsobrook and Heil, 2009). Thus, the combination of the I and D allele of the ACE I/D polymorphism may be advantageous for good performance in skiing. In our study, it was not possible to distinguish between the sprint and distance skiers because young skiers participated in both distances during the observed age period. In addition, our results did not confirm reliable evidence for the association between the ACE II genotype and endurance performance.
Our findings showed a higher prevalence of the ACTN3 RR genotype among young male skiers, which does not concur with the evidence from meta-analysis by Alfred et al. (2011), where the RR genotype was found more common in sprint and power athletes compared with controls among Europeans (Alfred et al., 2011). However, no evidence was found in this meta-analysis that the X allele gives an advantage in endurance performance (Alfred et al., 2011).
Several studies (Alsobrook and Heil, 2009; Paavolainen et al., 1991) emphasize the importance of muscle mass and explosive power in the speed of skiing. A higher prevalence of the ACTN3 RR genotype among young male skiers compared with the male control group (p = 0.006) in our study may explain the previous argument. In a study among Russian endurance athletes, researchers found that the ACTN3 X allele was underrepresented in Russian endurance athletes and the α-actinin-3 deficiency was estimated as one possible limiting factor in improving power and strength in endurance events (Ahmetov et al., 2010). Data of the present study suggest that the distribution of the ACTN3 R577X genotypes among young male and female skiers was different. Although there was a statistically significant over-representation of the RR genotype in male skiers in comparison with male controls, no significant differences between young female skiers and female controls were found. One reason for this might be a relatively small sample of the female study group (n = 17).
In our study, we did not find a significant association between the ACE I/D, the ACTN3 R577X genotypes and increase of VO2peak (ΔVO2peak, mL·kg-1·min-1) in either the male or female study group. However, a larger increase of the VO2peak (mL·kg-1·min-1) during the 5-year training process can be observed among female skiers with the ACE ID genotype, but it remained statistically nonsignificant (t = 0.760, p = 0.472). On the contrary, the ΔVO2peak (mL·kg-1·min-1) in male skiers during the 5-year observation period was not associated with the ACE II, ID or DD genotype. We detected a recognizable, although statistically nonsignificant improvement in the ΔVO2peak (mL·kg-1·min-1) among male skiers with the ACTN3 XX genotype, which is in agreement with the studies, where a higher XX genotype prevalence in endurance sport has been found (Eynon et al., 2012; Yang et al., 2003). However, a limiting factor of the present study is the small sub-group of the ACTN3 XX genotype − four subjects of 41 male skiers. At the same time, Thompson et al. (2006) did not find significant associations between the dynamics of the VO2max and ACE I/D genotypes after a 6-week training program. In our study, the ΔVO2peak (L·min-1), the ΔVO2peak (mL·kg-1·min-1) and the Δ duration of the standardized exercise test did not show significant associations with the ACE I/D and ACTN3 R577X genotypes. We can assume that the ACE II and the ACTN3 XX genotypes might not provide an advantage in endurance performance phenotypes in our study group. The latter supposition conforms to several previous studies, as conducted by Alfred et al. (2011) and Lucia et al. (2006). We observed the significant correlation between the Δ training volume and the ΔVO2peak (mL·kg-1·min-1), i.e. increase in training hours related positively to the dynamics of the VO2peak (mL·kg-1·min-1). Rusko (1992) has described in his review article that longitudinal studies among skiers have referred to the VO2max increase with age and training during the 15-20-year-old period, and the annual increase may amount up to 1-3 mL·kg-1·min-1 per year. In our study, a significant main effect of the VO2peak (mL·kg-1·min-1) was detected in the total study group, but no significant effect was found between the VO2peak (mL·kg-1·min-1) and the ACE I/D and ACTN3 R577X genotypes interactions.
Conclusion
The main result of this study was a significantly higher prevalence of the ACE ID and the ACTN3 RR genotypes among Estonian young male skiers compared with the male control group, which may be an advantage for the explosive speed and power capacity in race skiing. Our study showed a more remarkable trend of increase in the VO2peak (mL·kg-1·min-1) during the 5-year period among male skiers with the ACTN3 XX genotype and among female skiers with the ACE ID genotype. No significant genotype-related associations in the dynamics of the VO2peak during the 5-year period were found.
From a practical viewpoint, the genetic profile combined with physiological and biochemical characteristics could form a strong predictive package, which could be used by athletes and their teams for personalized management, monitoring and better optimization of training processes in the future. As multiple combinations of gene variants influence physical performance phenotypes, future longitudinal research, including different gene variants, may contribute to a better understanding of the nature of physical performance.
Acknowledgements
This study was supported by Estonian Institutional Research Funding No. IUT 02-7 and IUT 20-46. The authors wish to thank Helen Kaptein for excellent technical assistance. The authors declare that they have no conflicts of interest regarding the publication of this paper.
Biographies

Agnes MÄGI
Employment
Department of Sports Medicine and Rehabilitation, Faculty of Medicine, University of Tartu, Estonia. Sports Medicine and Rehabilitation Clinic, Tartu University Hospital, Estonia.
Degree
MD
Research interests
Genetics and physical performance; preparticipation evaluation of young athletes, prevention of health disorders in athletes.
E-mail: agnes.magi@kliinikum.ee

Eve UNT
Employment
Associate Professor of Sports Medicine and Rehabilitation, Department of Sports Medicine and Rehabilitation, Faculty of Medicine, University of Tartu. Estonia. Sports Medicine and Rehabilitation Clinic, Tartu University Hospital, Estonia. Department of Cardiology, Faculty of Medicine, University of Tartu, Estonia
Degree
PhD, MD
Research interests
Genetics and physical performance; preparticipation evaluation of young athletes, cardiovascular risk factors in athletes
E-mail: eve.unt@ut.ee
Ele PRANS
Employment
Research Fellow of Pathophysiology, Department of Pathophysiology, Institute of Biomedicine and Translational Medicine, Faculty of Medicine, University of Tartu, Estonia
Degree
MSc
Research interests
Health, biomedicine
E-mail: ele.prans@ut.ee
Liina RAUS
Employment
Department of Sports Medicine and Rehabilitation, Faculty of Medicine, University of Tartu, Estonia.
Degree
MSc
Research interests
Genetics and physical performance
E-mail: liinaraus@gmail.com
Jaan EHA
Employment
Professor of Cardiology, Department of Cardiology, Faculty of Medicine, University of Tartu, Estonia. Heart Clinic, Tartu University Hospital, Estonia
Degree
PhD, MD
Research interests
Invasive diagnostics of ischemic heart disease, reperfusion in acute myocardial infarction
E-mail: jaan.eha@ut.ee

Alar VERAKSITŠ
Employment
Senior Assistant in Human Physiology. Department of Physiology, Institute of Biomedicine and Translational Medicine, Faculty of Medicine, University of Tartu, Estonia.
Degree
PhD, MD
Research interests
Genetics, cardiology, muscle physiology
E-mail: alar.veraksits@ut.ee
Külli KINGO
Employment
Professor in Dermatology and Venereology, Department of Dermatology, Faculty of Medicine, University of Tartu, Estonia. Dermatology Clinic, Tartu University Hospital, Estonia
Degree
PhD, MD
Research interests
To identify network of genes and biomolecules which are involved in and malfunction of which leads to development chronic dermatoses.
E-mail: kylli.kingo@ut.ee
Sulev KÕKS
Employment
Professor in Pathophysiology, Lead Research Fellow of Physiological Genomics, Department of Pathophysiology, Institute of Biomedicine and Translational Medicine, Faculty of Medicine, University of Tartu, Estonia.
Degree
PhD, MD
Research interests
Genomics, pathophysiology
E-mail: sulev.koks@ut.ee
References
- Ahmetov I.I., Druzhevskaya A.M., Astratenkova I.V., Popov D.V., Vinogradova O.L., Rogozkin V.A. (2010) The ACTN3 R577X polymorphism in Russian endurance athletes. British Journal of Sports Medicine 44, 649-652. [DOI] [PubMed] [Google Scholar]
- Alfred T., Ben-Shlomo Y., Cooper R., Hardy R., Cooper C., Deary I.J., Gunnell D., Harris S.E., Kumari M., Martin R.M., Moran C.N., Pitsiladis Y.P., Ring S.M., Sayer A.A., Smith G.D., Starr J.M., Kuh D., Day I.N.HALCyon study team. (2011) ACTN3 genotype, athletic status, and life course physical capability: meta-analysis of the published literature and findings from nine studies. Human Mutation 32, 1008-1018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alsobrook N.G., Heil D.P. (2009) Upper body power as a determinant of classical cross-country ski performance. European Journal of Applied Physiology 105, 633-641. [DOI] [PubMed] [Google Scholar]
- Alvarez R., Terrados N., Ortolano R., Iglesias-Cubero G., Reguero J.R., Batalla A., Cortina A., Fernández-García B., Rodríguez C., Braga S., Alvarez V., Coto E. (2000) Genetic variation in the renin-angiotensin system and athletic performance. European Journal of Applied Physiology 82, 117-120. [DOI] [PubMed] [Google Scholar]
- Armstrong N., Welsman J.R. (2001) Peak oxygen uptake in relation to growth and maturation in 11- to 17-year-old humans. European Journal of Applied Physiology 85, 546-551. [DOI] [PubMed] [Google Scholar]
- Bassett D.R., Jr., Howley E.T. (2000) Limiting factors for maximum oxygen uptake and determinants of endurance performance. Medicine and Science in Sports and Exercise 32, 70-84. [DOI] [PubMed] [Google Scholar]
- Bongers B.C., Brussel M., Hulzebos E.H.J., Takken T. (2014) Pediatric norms for the cardiopulmonary exercise test. In: Pediatric norms for cardiopulmonary exercise testing. In relation to sex and age. 2 nd edition Uitgeverij BOXPress, ‘s-Hertogenbosch: 48-57. [Google Scholar]
- Bouchard C., Daw E.W., Rice T., Pérusse l., Gagnon J., Privince M.A., Leon A.S., Rao D.C., Skinner J.S., Wilmore J.H. (1998) Familial resemblance for VO2max in the sedentary state: the HERITAGE family study. Medicine and Science in Sports and Exercise 30, 252-258. [DOI] [PubMed] [Google Scholar]
- Davis J.A. (2006) Direct determination of aerobic power. In: Physiological assessment of human fitness. Eds: Maud P.J., Foster C. 2 nd edition United States: Human Kinetics; 9-18. [Google Scholar]
- Döring F.E., Onur S., Geisen U., Boulay M.R., Pérusse L., Rankinen T., Rauramaa R., Wolfahrt B., Bouchard C. (2010) ACTN3 R577X and other polymorphisms are not associated with elite endurance athlete status in the Genathlete study. Journal of Sports Sciences 28, 1355-1359. [DOI] [PubMed] [Google Scholar]
- Durstine J.L., Pate R.R., Sparling P.B., Wilson G.E., Senn M.D., Bartoli W.P. (1987) Lipid, lipoprotein and iron status of elite women distance runners. International Journal of Sports and Medicine 8, 119-123. [DOI] [PubMed] [Google Scholar]
- Eynon N., Ruiz J.R., Femia P., Pushkarev V.P., Cieszczyk P., Maciejewska-Karlowska A., Sawczuk M., Dyatlov D.A., Lekontsev E.V., Kulikov L.M., Birk R., Bishop D.J., Lucia A. (2012) The ACTN3 R577X polymorphism across three groups of elite male European athletes. PLoS One 7, e43132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folland J., Leach B., Little T., Hawker K., Myerson S., Montgomery H., Jones D. (2000) Angiotensin-converting enzyme genotype affects the response of human skeletal muscle to functional overload. Experimental Physiology 85, 575-579. [PubMed] [Google Scholar]
- Joyner M.J., Coyle E.F. (2008) Endurance exercise performance: the physiology of champions. The Journal of Physiology 586, 35-44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lucia A., Gómez-Gallego F., Santiago C., Bandrés F., Earnest C., Rabadán M., Alonso J.M., Hoyos J., Córdova A., Villa G., Foster C. (2006) ACTN3 genotype in professional endurance cyclists. International Journal of Sports Medicine 27, 880-884. [DOI] [PubMed] [Google Scholar]
- Ma F., Yang Y., Li X., Zhou F., Gao C., Li M., Gao L. (2013) The association of sport performance with ACE and ACTN3 genetic polymorphisms: a systematic review and meta-analysis. PLoS One 8, e54685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacArthur D.G., North K.N. (2005) Genes and human elite athletic performance. Human Genetics 116, 331-339. [DOI] [PubMed] [Google Scholar]
- Miller S.A., Dykes D.D., Polesky H.F. (1988) A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Research 16 1215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mills M., Yang N., Weinberger R., Vander Woude D.L., Beggs A.H., Easteal S., North K. (2001) Differential expression of the actin-binding proteins, alpha-actinin-2 and -3, in different species: implications for the evolution of functional redundancy. Human Molecular Genetics 10, 1335-1346. [DOI] [PubMed] [Google Scholar]
- Mitchell J.H., Haskell W., Snell P., van Camp S.P. (2005) Task Force 8: classification of sports. Journal of the American College of Cardiology 45, 1364-1367. [DOI] [PubMed] [Google Scholar]
- Myerson S., Hemingway H., Budget R., Martin J., Humphries S., Montgomery H. (1999) Human angiotensin I-converting enzyme gene and endurance performance. Journal of Applied Physiology 87, 1313-1316. [DOI] [PubMed] [Google Scholar]
- North K.N., Yang N., Wattanasirichaigoon D., Mills M., Easteal S., Beggs A.H. (1999) A common nonsense mutation results in alpha-actinin-3 deficiency in the general population. Nature Genetics 21, 353-354. [DOI] [PubMed] [Google Scholar]
- Paavolainen L., Häkkinen K., Rusko H. (1991) Effects of explosive type strength training on physical performance characteristics in cross-country skiers. European Journal of Applied Physiology and Occupational Physiology 62, 251-255. [DOI] [PubMed] [Google Scholar]
- Pate R.R., Sparling P.B., Wilson G.E., Cureton K.J., Miller B.J. (1987) Cardiorespiratory and metabolic responses to submaximal and maximal exercise in elite woman distance runners. International Journal of Sports Medicine 8, 91-95. [DOI] [PubMed] [Google Scholar]
- Rankinen T., Wolfarth B., Simoneau J.A., Maier-Lenz D., Rauramaa R., Rivera M.A., Boulay M.R., Chagnon Y.C., Pérusse L., Keul J., Bouchard C. (2000) No association between the angiotensin-converting enzyme ID polymorphism and elite endurance athlete status. Journal of Applied Physiology 88, 1571-1575. [DOI] [PubMed] [Google Scholar]
- Rusko H. (2003) Physiology of cross-country skiing. In: Cross country skiing. Ed: Rusko H. United Kingdom: Blackwell Science; 1-31. [Google Scholar]
- Rusko H.K. (1992) Development of aerobic power in relation to age and training in cross-country skiers. Medicine and Science in Sports and Exercise 24, 1040-1047. [PubMed] [Google Scholar]
- Taylor R.R., Mamotte C.D., Fallon K., van Bockxmeer F.M. (1999) Elite athletes and the gene for angiotensin-converting enzyme. Journal of Applied Physiology 87, 1035-1037. [DOI] [PubMed] [Google Scholar]
- Thompson P.D., Tsongalis G.J., Ordovas J.M., Seip R.L., Bilbie C., Miles M., Zoeller R., Visich P., Gordon P., Angelopoulos T.J., Pescatello L., Moyna N. (2006) Angiotensin-converting enzyme genotype and adherence to aerobic exercise training. Preventive Cardiology 9, 21-24. [DOI] [PubMed] [Google Scholar]
- Thompson W.R., Binder-Macleod S.A. (2006) Association of genetic factors with selected measures of physical performance. Physical Therapy 86, 585-591. [PMC free article] [PubMed] [Google Scholar]
- Yang N., MacArthur D.G., Gulbin J.P., Hahn A.G., Beggs A.H., Easteal S., North K. (2003) ACTN3 genotype is associated with human elite athletic performance. American Journal of Human Genetics 73, 627-631. [DOI] [PMC free article] [PubMed] [Google Scholar]