

Abstract
Each year more than two billion songbirds cross the Sahara, but how they perform this formidable task is largely unknown. Using geolocation tracks from 27 pied flycatchers, a nocturnally migrating passerine, we show that most birds made diurnal flights in both autumn and spring. These diurnal flights were estimated to be part of non-stop flights of mostly 40–60 h. In spring, birds flew across the Sahara, while autumn migration probably circumpassed part of the desert, through a long oversea flight. Our data contradict claims that passerines cross the Sahara by intermittent flight and daytime resting. The frequent occurrence of long non-stop flights to cross the desert shows migrants' physiological abilities and poses the question why this would not be the general migration strategy to cross the Sahara.
Keywords: Ficedula hypoleuca, Afro-Palaearctic passerine, fuelling, migration strategy, Sahara desert
1. Introduction
Surprisingly, little is known of how the billions of small avian migrants cross the Sahara desert, while migrating from their temperate and boreal breeding grounds to the African wintering grounds [1,2]. Moreau [2] presumed that most passerines cross the Sahara in a non-stop 40–60 h flight, implying that nocturnal migrants also migrate during daytime. More recent studies argued that most nocturnally migrating passerines exhibit an intermitted rather than a non-stop migration to cross the Sahara. Radar observations in the western Sahara during spring estimated that only 17% of songbirds (exclusively Hirundinidae) prolonged migratory flights during the day, while in autumn, this appeared even less common [3]. Direct observations of large numbers of individuals sheltering during the day against the heat, without water or food, but in good condition [4], support this notion of intermittent flight.
In contrast to radar studies, geolocator studies yield species-specific tracking of individual movements and are thus less dependent on specific observation times and locations. Moreover, they provide insight into variation in migration strategies such as among seasons, species and individuals. Geolocation tracking revealed prolonged flights in small passerines of up to several days [5,6]: e.g. blackpoll warblers, Setophaga striata, of 12 g crossed the western Atlantic in a non-stop 2540 km flight of 60 h [6]. Given these remarkable flights, it would be rather surprising if migrants facing the Sahara do not use non-stop flights. Indeed, recent geolocation tracks showed prolonged night flights into the day over the Sahara in several passerines [7], but the authors of that study did not conclude that birds flew non-stop. However, an intermittent flight strategy probably increases the time individuals take to cross the Sahara, and hence requires larger reserves prior to migration. Here, we provide evidence from light-level geolocator loggers (hereafter, loggers) for an abundant, and presumed nocturnal long-distant migrant, the pied flycatcher Ficedula hypoleuca [8], that most individuals showed 1–2 day periods of diurnal flight. We argue that these diurnal flights are part of a non-stop flight strategy that reduces the costs of barrier crossing.
2. Material and methods
We studied migratory pied flycatchers from a nest-box population in Drenthe (NL, 52°49′ N, 6°22′ E [9]). Recent tracking work showed that our population wintered in the western Ivory Coast/eastern Guinea in sub-Saharan Africa (mean longitude 7.4°W ± 1.0° [10]) and migrated in spring in a mere two weeks to the breeding grounds, where they arrived between 5 April and 10 May. Most breeders migrate in early August in five weeks to their sub-Saharan African wintering grounds, with a major stop-over on the Iberian Peninsula [11]. For this study, we equipped 52 adult males and 28 females in 2013 with Intigeo-W50 geolocator loggers (Migrate Technology Ltd, Cambridge, UK) using leg-loop harnesses (avg total mass 0.52 g) just before chicks fledged (breeding adults), or during nest-box advertisement (unpaired males). We recaptured 27 birds with loggers in 2014–2015, and found no evidence for lower return rates or later spring arrival between birds with or without loggers [10]. From these loggers, 12 recorded maximum light levels every 10 min (measured 1 min intervals) and 15 recorded maximum light levels every 5 min and for each 4 h period the minimal and maximal temperature observed over this interval (measured 5 min intervals). For 15 birds, data included the onset of spring migration. Temperature was successfully recorded for 15 birds in autumn and seven in spring. Owing to unstable shading events, latitudes could not be calculated with large accuracy, but longitudes are reliable and therefore we report these [12].
Because flycatchers prefer wooded habitats, recorded daytime light levels by loggers are normally erratic (figure 1). Here we make use of changes in shading and temperature to show that pied flycatchers are not always nocturnal migrants. We found that twice a year a short period occurred with extremely smooth transitions without shading and high maximum daily light values in the light data. These periods ended with an abrupt occurrence of shading during the day. In woodland species, such ‘bright’ periods likely refer to windows of diurnal flight. Diurnal flight is associated with large changes in twilight times, mostly at sunrise, indicating fast movements in a southwesterly direction in autumn and northeasterly direction in spring (electronic supplementary material, figure S1 and table S1).
Figure 1.

Examples of three pied flycatchers (male: F272/F291; female: F384) that used diurnal flight twice a year when facing ecological barriers, like the Sahara. Light and temperature profiles (a) show 2 days before, 2 days during and 2 days after diurnal flight in autumn and spring, and changes in longitude positions (b) across 2013–2014. Grey lines indicate diurnal flight (periods). Temperatures (in a) are approximate changes in ambient temperature rather than being absolute measures and were saved at the end of each 4 h period.
3. Results and discussion
Of the 15 flycatchers with loggers recording data in spring, 14 showed 1–2 day cycles with higher and less erratic light levels, which directly documented non-stop diurnal migration (figure 1). These periods coincided with the start of spring migration in a northeasterly direction (range: March 23–April 29 [10], figure 2; electronic supplementary material, table S1). Days without shading are unlikely to have been caused by resting in the desert, because the six loggers recording temperature showed lower and more stable temperatures during these days than before and after diurnal flight (figure 1). The observed reduction in temperature amplitudes during daytime are in agreement with radar studies showing that birds cross the Sahara in spring while flying at high altitudes in cool anti-trade winds [13]. Some individuals ended their diurnal flight during the day, as visible in the reappearance of shading events and a sharp increase in temperature (figure 1, individual F272, 1 April). In three birds, short temperature peaks at night occurred during spring diurnal flight periods (e.g. figure 1, individual F384), which may indicate lower altitude flights or short nightly resting stops. For the one individual with ambiguous diurnal flight during spring, temperature profiles were inconclusive to support intermitted or non-stop flight (see the electronic supplementary material, figure S2).
Figure 2.
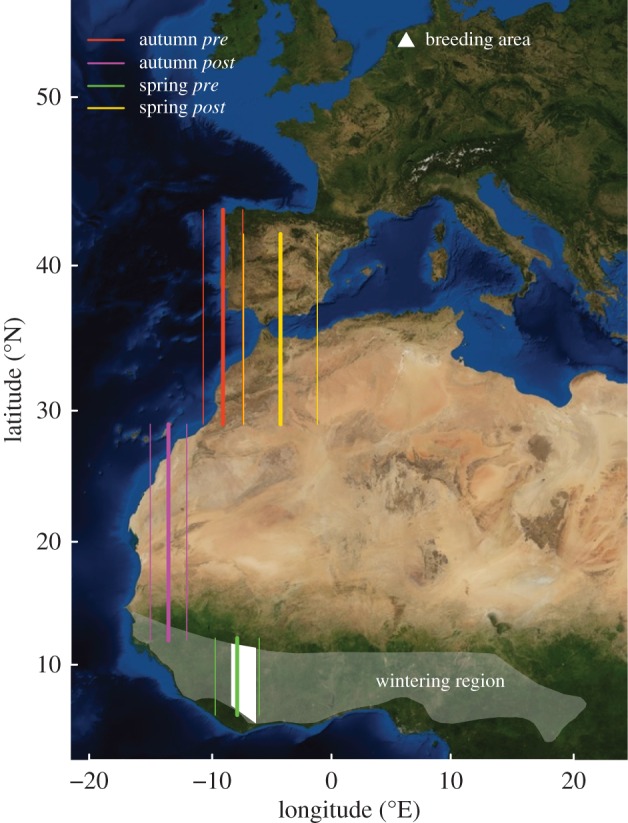
Longitudes (average, bold line; s.d.: thin line) of Dutch pied flycatchers before and after diurnal flight (see the electronic supplementary material, table S1) roughly indicate onset locations and destinations of non-stop flight. Latitudes prior to diurnal flight were chosen to include fuelling sites in autumn (Iberia, less so North Africa) or spring (wintering range). Latitudes after diurnal flight indicate possible destinations, assuming 15.3 m s−1 flight speed and the maximal flight duration (electronic supplementary material, table S1). Wintering longitude (average±s.d.) is depicted in white.
The estimated maximum duration of the prolonged flight in spring was 45.3 ± 5.89 h (n = 12) and possibly even longer (electronic supplementary material, table S1). Maximum flight duration ranged between 40 and 61 h, and with an average ground speed of 15.3 m s−1 [3], flycatchers could cover 2200–3350 km and likely reached at least the northern edge of the Sahara (figure 2).
In autumn, all individuals showed similar patterns with little daytime shading, suggesting diurnal flight during southward migration as well. In contrast to spring, less pronounced (but notable) changes in temperature profiles were observed (figure 1), because either flycatchers profit from favourable trade winds that bring warm and dry winds, or they fly over sea [13]. That these light curves indeed are the result of diurnal flight rather than resting in the desert is strongly suggested by the sudden appearance of shading, mostly happening half-way during the day (e.g. F384 on 25 August, figure 1). The maximum duration of autumn non-stop flights was variable: the shortest flights lasted probably 23–25 h (n = 4), but most took 37–48 h (n = 23, average 41.2 + 2.8 h), and possibly even longer (up to 60 h; see the electronic supplementary material, table S1).
It has been suggested that in autumn, migrants suffer more from dehydration when crossing the Sahara, and hence have to stop during the hot day [3,4]. Alternatively, we suggest that most flycatchers cross the ocean and thereby circumpass the desert (figure 2). A direct flight across the ocean is supported by pied flycatchers being the most abundant prey of breeding Eleonora's falcons, Falco eleonorae, in September on the Canary Islands [14]. Our data from longitudes before the onset of diurnal flight agree with major stop-over areas in the west of the Iberian Peninsula [8,11], and after this flight they have consistently shifted to more western longitudes in West Africa (figure 2; electronic supplementary material, table S1). Based on 15.3 m s−1 ground speed [3], mean departure weights of 16.8 g from southwest Iberia [11], and a flight duration of 37–48(−60) h (n = 24), most flycatchers can cover ca 2000–2600(−3300) km and thus reach the south edge of the desert in this prolonged flight. Along the coast south of the Sahara, suitable scattered vegetation seems to support further autumn passage, and pied flycatchers were mostly in good condition at these stop-over sites [15]. More southwards, flycatchers likely changed direction eastwards towards their final wintering sites [12,16].
Our data are indirect but highly suggestive evidence that pied flycatchers from our breeding population commonly flew non-stop when migrating across the Sahara. This pattern contradicts claims that the general pattern of Sahara crossings in passerines is by intermittent flight. However, these claims are mainly based on the relative rarity of daytime passage on radar observations at few spots and the assumption that most migrants depart for spring migration from the southern Sahara edge. By contrast, our flycatchers departed directly from their more southern wintering grounds [10], and if they departed shortly after sunset, they likely passed these radar observation sites after ca 24–28 h flight, during the night. High departure fat loads [11,17] and relatively few observations of pied flycatchers resting in or near oases during autumn and spring migration [15,18] support a non-stop migration strategy. Observations with geolocators of diurnal flights in various small nocturnal migrants [6,7], suggest that the non-stop strategy is not limited to the relatively fast migrating pied flycatcher [10]. We argue that for many small migrants, like pied flycatchers, a non-stop flight strategy must be superior to an intermittent strategy both in autumn and in spring, probably because it reduces time, energy and risk of dehydration while passing the Sahara.
Our finding of non-stop Sahara crossing by a 12 g passerine challenges the view on the severity of such a barrier. Our observation that flycatchers in spring take off on a non-stop flight from at least 500 km south of the start of the desert suggests that physiology does not constrain a broader crossing. Furthermore, flycatchers seem to perceive barriers differently between seasons, as only in spring do they seem to fly directly over the desert. The next challenge is to compare species' migration strategies, to unravel how variable these patterns are and their associated costs and benefits [19]. This is especially important as long-distance migrants face large changes at the wintering grounds [1] that may hamper their preparation for successful barrier crossings.
Supplementary Material
Acknowledgements
We thank J. Fox for data extraction, R. G. Bijlsma, R. H. G. Klaassen and two anonymous referees for valuable comments on the manuscript.
Ethics
Experiments were approved by the ethical committee on animal experiments of the University of Groningen, The Netherlands (permit: 5588).
Data accessibility
Data supporting this article are included as the electronic supplementary material.
Authors' contributions
J.O. analysed data. J.O./C.B. performed the experiment, wrote, approved and are accountable for the final content of the manuscript.
Competing interests
We declare we have no competing interests.
Funding
Financial support was provided by The Netherlands Organisation for Scientific Research (NWO-ALW-814.01.010 to J.O./C.B., VIDI-NWO-864.06.004 to C.B.) and KNAW Schure-Beijerink Popping foundation (to C.B.).
References
- 1.Zwarts L, Bijlsma RG, van der Kamp J, Wymenga E. 2009. Living on the edge: wetlands and birds in a changing Sahel. Zeist, The Netherlands: KNNV Uitgeverij. [Google Scholar]
- 2.Moreau RE. 1972. The Palearctic-African bird migration systems. New York, NY: Academic Press. [Google Scholar]
- 3.Schmaljohann H, Liechti F, Bruderer B. 2007. Songbird migration across the Sahara: the non-stop hypothesis rejected! Proc. R. Soc. B 274, 735–739. ( 10.1098/rspb.2006.0011) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Newton I. 2008. The migration ecology of birds. London, UK: Academic Press. [Google Scholar]
- 5.Bairlein F, Norris DR, Nagel R, Bulte M, Voigt CC, Fox JW, Hussell DJT, Schmaljohann H. 2012. Cross-hemisphere migration of a 25 g songbird. Biol. Lett. 8, 505–507. ( 10.1098/rsbl.2011.1223) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.DeLuca WV, Woodworth BK, Rimmer CC, Marra PP, Taylor PD, McFarland KP, Mackenzie SA, Norris DR. 2015. Transoceanic migration by a 12 g songbird. Biol. Lett. 11, 20141045 ( 10.1098/rsbl.2014.1045) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Adamík P, et al. 2016. Barrier crossing in small avian migrants: individual tracking reveals prolonged nocturnal flights into the day as a common migratory strategy. Sci. Rep. 6, 21560 ( 10.1038/srep21560) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lundberg A, Alatalo RV. 1992. The pied flycatcher. London, UK: T&AD Poyser. [Google Scholar]
- 9.Both C, Bijlsma RG, Ouwehand J. 2016. Repeatability in spring arrival dates in pied flycatchers varies among years and sexes. Ardea 104, 3–20. [Google Scholar]
- 10.Ouwehand J, Both C. Submitted. African departure rather than migration duration determines variation in spring arrival in pied flycatchers. [DOI] [PubMed] [Google Scholar]
- 11.Bibby CJ, Green RE. 1980. Foraging behaviour of migrant pied flycatchers, Ficedula hypoleuca, on temporary territories. J. Anim. Ecol. 49, 507–521. ( 10.2307/4260) [DOI] [Google Scholar]
- 12.Ouwehand J, et al. 2016. Light-level geolocators reveal migratory connectivity in European populations of pied flycatchers Ficedula hypoleuca. J. Avian Biol. 47, 69–83. ( 10.1111/jav.00721) [DOI] [Google Scholar]
- 13.Schmaljohann H, Liechti F, Bruderer B. 2009. Trans-Sahara migrants select flight altitudes to minimize energy costs rather than water loss. Behav. Ecol. Sociobiol. 63, 1609–1619. ( 10.1007/s00265-009-0758-x) [DOI] [Google Scholar]
- 14.de Leon L, Rodrigues B, Martin A, Nogales M, Alonso J, Izquierdo C. 2007. Status, distribution, and diet of Eleonora's falcon (Falco eleonorae) in the Canary Islands. J. Raptor Res. 41, 331–336. ( 10.3356/0892-1016(2007)41[331:SDADOE]2.0.CO;2) [DOI] [Google Scholar]
- 15.Salewski V, Schaub M. 2007. Stopover duration of Palearctic passerine migrants in the western Sahara—independent of fat stores? Ibis 149, 223–236. ( 10.1111/j.1474-919X.2006.00608.x) [DOI] [Google Scholar]
- 16.Liechti F, Komenda-Zehnder S, Bruderer B. 2012. Orientation of passerine trans-Sahara migrants: the directional shift (‘Zugknick’) reconsidered for free-flying birds. Anim. Behav. 83, 63–68. ( 10.1016/j.anbehav.2011.10.005) [DOI] [Google Scholar]
- 17.Smith VW. 1966. Autumn and spring weights of some palaearctic migrants in central Nigeria. Ibis 108, 492–512. ( 10.1111/j.1474-919X.1966.tb07206.x) [DOI] [Google Scholar]
- 18.Jenni-Eiermann S, Almasi B, Maggini I, Salewski V, Bruderer B, Liechti L, Jenni L. 2011. Numbers, foraging and refuelling of passerine migrants at a stopover site in the western Sahara: diverse strategies to cross a desert. J. Ornithol. 151, 113–128. ( 10.1007/s10336-010-0572-2) [DOI] [Google Scholar]
- 19.Deppe JL, et al. 2015. Fat, weather, and date affect migratory songbirds’ departure decisions, routes, and time it takes to cross the Gulf of Mexico. Proc. Natl Acad. Sci. USA 112, E6331–E6338. ( 10.1073/pnas.1503381112) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data supporting this article are included as the electronic supplementary material.