

Abstract
The Tremarctinae are a subfamily of bears endemic to the New World, including two of the largest terrestrial mammalian carnivores that have ever lived: the giant, short-faced bears Arctodus simus from North America and Arctotherium angustidens from South America (greater than or equal to 1000 kg). Arctotherium angustidens became extinct during the Early Pleistocene, whereas Arctodus simus went extinct at the very end of the Pleistocene. The only living tremarctine is the spectacled bear (Tremarctos ornatus), a largely herbivorous bear that is today only found in South America. The relationships among the spectacled bears (Tremarctos), South American short-faced bears (Arctotherium) and North American short-faced bears (Arctodus) remain uncertain. In this study, we sequenced a mitochondrial genome from an Arctotherium femur preserved in a Chilean cave. Our molecular phylogenetic analyses revealed that the South American short-faced bears were more closely related to the extant South American spectacled bear than to the North American short-faced bears. This result suggests striking convergent evolution of giant forms in the two groups of short-faced bears (Arctodus and Arctotherium), potentially as an adaptation to dominate competition for megafaunal carcasses.
Keywords: Ursidae, great American biotic interchange, molecular dating, palaeontology
1. Introduction
The spectacled bear (Tremarctos ornatus) is the only living member of Tremarctinae, a previously diverse group of bears endemic to the Americas. The now-extinct Pleistocene diversity of Tremarctinae comprised the Florida spectacled bear (Tremarctos floridanus), South American short-faced bears (Arctotherium—five species; [1]) and North American short-faced bears (Arctodus—two species; [2]). These species ranged in size from the relatively small Arctotherium wingei (approx. 150 kg; [3]) to the giant short-faced bears Arctodus simus and Arctotherium angustidens, which may have attained body masses exceeding 1000 kg [4,5]. In addition, tremarctine bears displayed a diversity of foraging strategies, ranging from carnivorous/omnivorous (e.g. Arctodus simus, Arctotherium angustidens) to largely herbivorous (e.g. Arctotherium wingei, T. ornatus) [6–9]. The evolution and biogeography of this diverse group of bears is enigmatic, and currently lacks a robust phylogenetic framework.
Putative representatives of both Arctodus and Tremarctos are known from the Late Pliocene fossil record of North America [2,10,11], and genetic data suggest these two genera diverged during the Late Miocene or Early Pliocene [12]. In contrast, the earliest representative of Arctotherium appears in the Early Pleistocene fossil record of South America [1,4,10]. Concordantly, it is generally accepted that Arctotherium arose as a distinct clade following dispersal of North American ancestors to South America after the Panamanian Isthmus formed approximately 3 million years ago (Ma) [4,13–15], an event that precipitated a period of extensive faunal interchange between North and South America known as the great American biotic interchange (GABI) [16–18]. Conversely, Tremarctos does not appear in the South American fossil record until the Holocene, suggesting that the extant spectacled bear descends from an independent, later dispersal event.
Previous palaeontological studies have considered the short-faced bears (Arctotherium and Arctodus) as forming a monophyletic clade [19–21]. The proportionally larger and wider molars of both Arctodus and Arctotherium set them apart from other bears [19–21], and the cranium and mandible morphology of Arctodus simus and Arctotherium angustidens are strikingly similar [6]. However, the similarities between Arctodus and Arctotherium could alternatively result from convergent evolution driven by adaptation to the same ecological niche and/or feeding behaviour, as dental and mandible characters are frequently correlated with diet [8]. Similarly, the huge size of Arctodus simus and Arctotherium angustidens may have been convergent, and linked to their ecology by allowing predation on large herbivores and competition for carcasses. Consequently, resolving the phylogenetic relationships among these taxa would have important implications for our understanding of body size and foraging strategy evolution in tremarctines.
We sequenced a near-complete mitochondrial genome from a specimen of Arctotherium and compared these new data with previously published sequences from the extant spectacled bear T. ornatus, the North American giant short-faced bear Arctodus simus, and a number of additional outgroups.
2. Methods
Analyses were performed on an Arctotherium femur collected from Cueva del Puma, Chile (complete right femur, no. 32104, Centro de Estudios del Hombre Austral, Instituto de la Patagonia, Universidad de Magallanes). DNA extraction and genomic library preparation were performed in the ancient DNA laboratory at the Australian Centre for Ancient DNA, University of Adelaide. The Arctotherium genomic library was enriched for mitochondrial DNA, and sequenced on an Illumina MiSeq; the resulting reads were assembled into a mitochondrial genome. We aligned the Arctotherium mitochondrial genome sequence to 13 previously published carnivoran sequences, used PartitionFinder v. 1.1.1 [22] to determine optimal partitioning and substitution models, and performed phylogenetic analyses with RAxML v. 8.2.0 [23] and beast v. 1.8.0 [24]. See the electronic supplementary material for detailed methods.
3. Results
Our analyses recovered relationships among outgroup taxa that were consistent with previous genetic studies ([12,25], but see [26]). Within Tremarctinae, we recovered strong support for a clade comprising Tremarctos ornatus and Arctotherium to the exclusion of Arctodus simus (Bayesian posterior probability = 1.0, maximum-likelihood bootstrap percentage = 98%; figure 1). Our mean estimate for the time of divergence between Tremarctos and Arctotherium was 4.1 Ma (95% highest posterior density, HPD = 3.0–5.3 Ma), and 4.8 Ma for the most recent common ancestor of Tremarctos, Arctotherium and Arctodus (95% HPD = 3.6–6.2 Ma). In general, node age estimates from our molecular dating analyses were slightly younger than those of a previous study of ursid mitochondrial DNA [12], although our 95% highest posterior densities (HPDs) overlapped with theirs substantially for equivalent nodes. These differences likely arise from our less restrictive calibration on the root of the tree, which allowed for the possibility that the Eocene Parictis is not a true member of the bear lineage. Conversely, a study of nuclear DNA obtained dates for the radiation of Ursinae that were much younger than ours [26], likely as a result of our conservative constraint on the crown-age of Ursidae, which permitted this node to substantially predate its first unequivocal fossil representative (electronic supplementary material).
Figure 1.
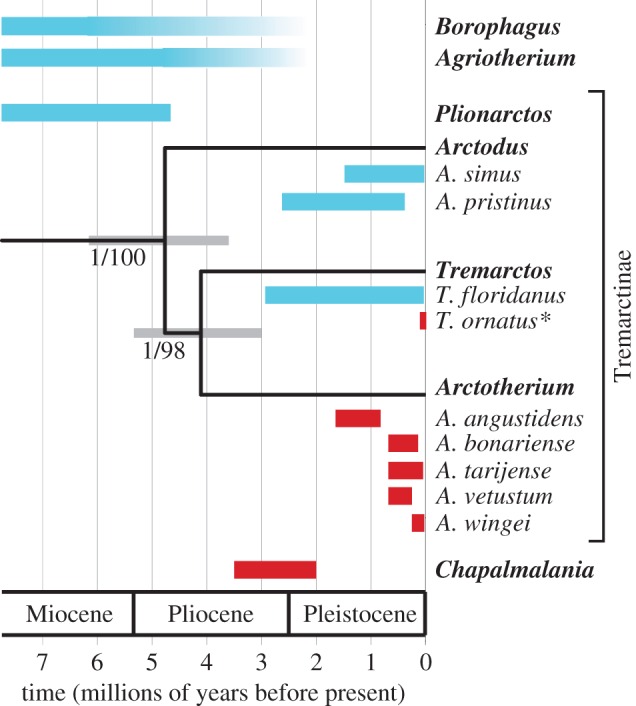
Relationships among tremarctine genera resulting from phylogenetic analysis of our mitochondrial genome dataset. Nodes reflect mean age estimates, whereas grey bars reflect 95% highest posterior densities (HPDs). Branch support values (BEAST posterior probability/RAxML bootstrap %) are given for each clade. The approximate temporal range of taxa of interest (see main text) is plotted based on the fossil record, and coloured according to distribution (North America, blue; South America, red). The extant taxon is marked with an asterisk.
4. Discussion
Our results suggest that the North and South American short-faced bears (Arctodus and Arctotherium, respectively) do not form a monophyletic clade (figure 1), contrary to the suggestions of previous palaeontological studies [19]. In addition, our molecular dating analyses indicate that Arctotherium, Arctodus and Tremarctos all diverged from one another during the Late Miocene or Pliocene. This inferred timeframe suggests that the Miocene/Pliocene genus Plionarctos is ancestral to the Quaternary tremarctine genera. Our observations are consistent with the idea that giant representatives of Arctodus and Arctotherium evolved independently in both North and South America during the Pleistocene [4], as all known Plionarctos specimens are relatively small-bodied (as was the earliest occurring species of Arctodus, Arctodus pristinus). The largest tremarctine bears (Arctodus simus and Arctotherium angustidens) appear to have been among the most inclined towards carnivory, although plant matter would likely still have made up a substantial proportion of their diets [6–8]. Consequently, the convergent morphological evolution of giant short-faced bears may have occurred as an adaptation for securing and scavenging large carcasses, as none of the tremarctine bears was specialized for active predation [27,28].
Many carnivoran species inhabited North America during the Pliocene and Pleistocene, including wolves (Canis lupus/Canis dirus), lions (Panthera leo) and the sabre-toothed cats (e.g. Smilodon, Xenosmilus, Homotherium). During the Pliocene, carcasses resulting from carnivore kills were probably scavenged by the ‘bone-crushing dogs’ (Borophagus spp.; [29]) or the bear Agriotherium [27], because the only known North American hyaenid (Chasmaporthetes ossifragus) does not appear to have been as well equipped for bone-crushing as extant Hyena and Crocuta species [30]. However, both Agriotherium and Borophagus appear to have become extinct by the end of the Pliocene, which may have vacated an ecological niche that Arctodus subsequently exploited. This explanation is compatible with the observation that the giant Arctodus simus is only known from the Pleistocene, whereas the temporal range of the smaller-bodied Arctodus pristinus overlaps with Borophagus and Agriotherium. It is also possible that changes in the large herbivore community during the Pleistocene—for example, the immigration of bison into North America—may have played a central role in the evolution of Arctodus simus.
The oldest known Arctotherium specimens are giant-sized [3,8,10], suggesting either that their size evolution occurred very rapidly or that fossils from the early stages of Arctotherium evolution have not yet been recorded. Our molecular dating estimates are compatible with both possibilities. A substantial proportion of the 95% HPD for our estimated time of divergence between Arctotherium and Tremarctos is distributed in the Early Pliocene, suggesting that Arctotherium may have existed in North and/or Central America for several million years without being detected in the fossil record (but see [31]) or perhaps even island-hopped to mainland South America prior to formation of the Isthmus of Panama approximately 3 Ma. However, our date estimates also allow for a Late Pliocene origin of Arctotherium, approximately coincident with the establishment of direct land connection between North and South America. This latter hypothesis is more consistent with the fossil record, as the earliest unequivocal records of Arctotherium are from the Early Pleistocene of South America (Ensenadan age).
The first recorded Arctotherium specimens in South America occur alongside the earliest known South American records of several other carnivorans: the sabre-toothed cats Smilodon and Homotherium, the puma (Puma concolor), the jaguar (Panthera onca), some large 25–35 kg canids, and several smaller less than 15 kg mustelids, canids, felids and mephitids [14,15,32,33]. This Ensenadan carnivore guild was dramatically more diverse than the communities recorded in immediately preceding layers, which included only a handful of carnivorous mammal species and none larger than approximately 30 kg [13,14,33]. Thus, species migrating southwards into South America during the GABI would initially have encountered an abundance of large mammalian herbivores and a paucity of carnivores [14,17,33]. Further, there would have been no South American mammals specialized for scavenging, because the large (more than 30 kg) procyonid Chapalmalania, which appears to have filled this niche [34], became extinct in the Late Pliocene/Early Pleistocene. Thus, a lack of competition for carcass scavenging during this period of faunal turnover may have driven the evolution of the giant Arctotherium angustidens [4].
Supplementary Material
Acknowledgements
We thank the Centro de Estudios del Hombre Austral (F. Morello) for providing access to the Arctotherium specimen, J. Weinstock for assistance with aDNA replication, and A. Camens and T. Worthy for discussions regarding early drafts of this manuscript. Grid computing facilities were provided by eResearch SA and CIPRES (Cyberinfrastructure for Phylogenetic Research).
Data accessibility
Data from this study are available on GenBank (KU886001) and the Dryad Data Repository (http://dx.doi.org/10.5061/dryad.v7f30).
Authors' contributions
A.C., J.J.A., F.P., A.P., L.S. and F.M. conceived of the study. S.C.B., K.J.M. and P.B. performed the laboratory work. S.C.B., K.J.M., L.S., B.W.S. and F.P. significantly contributed to the design of the analyses. K.J.M wrote the manuscript. All authors helped with interpretation of results and critically revised the manuscript. All authors approve the final version of this manuscript and agree to be held accountable for all aspects of the work performed.
Competing interests
The authors declare no competing interests.
Funding
The authors were supported by the Australian Research Council, Marie Curie International Outgoing Fellowship (MEDITADNA, PIOF-GA-2011-300854, FP7-PEOPLE), Consejo Nacional de Investigaciones Científicas y Técnicas (PIP 2011-164) and Agencia Nacional de Promoción Científica y Tecnológica (PICT 2011-309).
References
- 1.Soibelzon LH. 2004. Revisión sistemática de los Tremarctinae (Carnivora, Ursidae) fósiles de América del Sur [Systematic review of fossil Tremarctinae (Carnivora, Ursidae) from South America]. Rev. Mus. Argent. Cienc. Nat. 6, 107–133. [Google Scholar]
- 2.Kurtén B. 1967. Pleistocene bears of North America, II: genus Arctodus, short-faced bears. Acta Zool. Fenn. 117, 1–160. [Google Scholar]
- 3.Soibelzon LH, Tarantini VB. 2009. Estimación de la masa corporal de las especies de osos fósiles y actuales (Ursidae, Tremarctinae) de América del Sur [Body mass estimation of extinct and extant South American bears (Ursidae, Tremarctinae)]. Rev. Mus. Argent. Cienc. Nat. 11, 243–254. [Google Scholar]
- 4.Soibelzon LH, Schubert BW. 2011. The largest known bear, Arctotherium angustidens, from the Early Pleistocene pampean region of Argentina: with a discussion of size and diet trends in bears. J. Paleontol. 85, 69–75. ( 10.1666/10-037.1) [DOI] [Google Scholar]
- 5.Christiansen P. 1999. What size were Arctodus simus and Ursus spelaeus (Carnivora: Ursidae)? Ann. Zool. Fennici 36, 93–102. [Google Scholar]
- 6.Figueirido B, Soibelzon LH. 2009. Inferring palaeoecology in extinct tremarctine bears (Carnivora, Ursidae) using geometric morphometrics: palaeoecology in extinct tremarctines. Lethaia 43, 209–222. ( 10.1111/j.1502-3931.2009.00184.x) [DOI] [Google Scholar]
- 7.Figueirido B, Palmqvist P, Pérez-Claros JA. 2009. Ecomorphological correlates of craniodental variation in bears and paleobiological implications for extinct taxa: an approach based on geometric morphometrics. J. Zool. 277, 70–80. ( 10.1111/j.1469-7998.2008.00511.x) [DOI] [Google Scholar]
- 8.Soibelzon LH, Grinspan GA, Bocherens H, Acosta WG, Jones W, Blanco ER, Prevosti F. 2014. South American giant short-faced bear (Arctotherium angustidens) diet: evidence from pathology, morphology, stable isotopes, and biomechanics. J. Paleontol. 88, 1240–1250. ( 10.1666/13-143) [DOI] [Google Scholar]
- 9.Prevosti FJ, Martin FM. 2013. Paleoecology of the mammalian predator guild of Southern Patagonia during the latest Pleistocene: ecomorphology, stable isotopes, and taphonomy. Quat. Int. 305, 74–84. ( 10.1016/j.quaint.2012.12.039) [DOI] [Google Scholar]
- 10.Soibelzon LH, Tonni EP, Bond M. 2005. The fossil record of South American short-faced bears (Ursidae, Tremarctinae). J. South Am. Earth Sci. 20, 105–113. ( 10.1016/j.jsames.2005.07.005) [DOI] [Google Scholar]
- 11.Kurtén B. 1966. Pleistocene bears of North America, I: genus Tremarctos, spectacled bears. Acta Zool. Fenn. 115, 1–120. [Google Scholar]
- 12.Krause J, et al. 2008. Mitochondrial genomes reveal an explosive radiation of extinct and extant bears near the Miocene–Pliocene boundary. BMC Evol. Biol. 8, 220 ( 10.1186/1471-2148-8-220) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Soibelzon LH, Prevosti F. 2007. Los carnívoros (Carnivora, Mammalia) terrestres del Cuaternario de América del Sur [The Quaternary terrestrial carnivores (mammalia, carnivora) of South America]. In Geomorfologia Litoral i Quaternari Homenatge a Joan Cuerda Barceló (eds Pons GX, Vicens D). Palma, Majorca: Monografies de la Societat d'História Natural de Balears. [Google Scholar]
- 14.Prevosti F, Soibelzon LH. 2012. Evolution of the South American carnivores (Mammalia, Carnivora): a paleontological perspective. In Bones, clones, and biomes: the history and geography of Recent Neotropical mammals (eds Patterson BD, Costa LP), pp. 102–122. Chicago, IL: University of Chicago Press. [Google Scholar]
- 15.Soibelzon LH, Prevosti F. 2012. Fossils of South American land carnivores (Carnivora, Mammalia). In Molecular population genetics, evolutionary biology and biological conservation of neotropical carnivores (eds Ruiz M, Shostell J), pp. 509–530. New York, NY: Nova Science Publisher. [Google Scholar]
- 16.Marshall LG, Webb SD, Sepkoski JJ Jr, Raup DM. 1982. Mammalian evolution and the great American interchange. Science 215, 1351–1357. ( 10.1126/science.215.4538.1351) [DOI] [PubMed] [Google Scholar]
- 17.Cione A, Gasparini G, Soibelzon E, Soibelzon L, Tonni E. 2015. The GABI in southern South America. In The great American biotic interchange, pp. 71–96. Amsterdam, The Netherlands: Springer. [Google Scholar]
- 18.Woodburne MO. 2010. The great American biotic interchange: dispersals, tectonics, climate, sea level and holding pens. J. Mamm. Evol. 17, 245–264. ( 10.1007/s10914-010-9144-8) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Trajano E, Ferrarezzi H. 1995. A fossil bear from northeastern Brazil, with a phylogenetic analysis of the South American extinct Tremarctinae (Ursidae). J. Vertebr. Paleontol. 14, 552–561. ( 10.1080/02724634.1995.10011577) [DOI] [Google Scholar]
- 20.Soibelzon LH. 2002. Los ursidae (Carnivora: Fissipedia) fósiles de la República Argentina. Aspectos sistemáticos y paleoecológicos [The Ursidae (Carnivora: Fissipedia) fossils of Argentina. Systematic and palaeoecological aspects]. PhD thesis, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata.
- 21.Soibelzon LH, Schubert BW, Posadas P. 2010. A new phylogenetic analysis of tremarctine bears. In Paleontologia em Destaque. Edição Especial VII Simpósio Brasileiro de Paleontologia de Vertebrados (eds Costa da Silva R, Avilla L), pp. 115. Rio de Janeiro, Brazil: Sociedade Brasileira de Paleontologia. [Google Scholar]
- 22.Lanfear R, Calcott B, Ho SYW, Guindon S. 2012. PartitionFinder: combined selection of partitioning schemes and substitution. Mol. Biol. Evol. 29, 1695–1701. ( 10.1093/molbev/mss020) [DOI] [PubMed] [Google Scholar]
- 23.Stamatakis A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22, 2688–2690. ( 10.1093/bioinformatics/btl446) [DOI] [PubMed] [Google Scholar]
- 24.Drummond AJ, Rambaut A. 2007. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 7, 214 ( 10.1186/1471-2148-7-214) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pagès M, Calvignac S, Klein C, Paris M, Hughes S, Hanni C. 2008. Combined analysis of fourteen nuclear genes refines the Ursidae phylogeny. Mol. Phylogenet. Evol. 47, 73–83. ( 10.1016/j.ympev.2007.10.019) [DOI] [PubMed] [Google Scholar]
- 26.Kutschera VE, Bidon T, Hailer F, Rodi JL, Fain SR, Janke A. 2014. Bears in a forest of gene trees: phylogenetic inference is complicated by incomplete lineage sorting and gene flow. Mol. Biol. Evol. 31, 2004–2017. ( 10.1093/molbev/msu186) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Sorkin B. 2006. Ecomorphology of the giant short-faced bears Agriotherium and Arctodus. Hist. Biol. 18, 1–20. ( 10.1080/08912960500476366) [DOI] [Google Scholar]
- 28.Figueirido B, Perez-Claros JA, Torregrosa V, Martin-Serra A, Palmqvist P. 2010. Demythologizing Arctodus simus, the ‘short-faced’ long-legged and predaceous bear that never was. J. Vertebr. Paleontol. 30, 262–275. ( 10.1080/02724630903416027) [DOI] [Google Scholar]
- 29.Wang X, Tedford R, Taylor B. 1999. Phylogenetic systematics of the Borophaginae. Bull. Am. Mus. Nat. Hist. 243, 1–391. [Google Scholar]
- 30.Hartstone-Rose A. 2011. Reconstructing the diets of extinct South African carnivorans from premolar ‘intercuspid notch’ morphology. J. Zool. 285, 119–127. ( 10.1111/j.1469-7998.2011.00821.x) [DOI] [Google Scholar]
- 31.Soibelzon LH, Romero MR, Huziel Aguilar D, Tarantini VB. 2008. A Blancan (Pliocene) short-faced bear from El Salvador and its implications for tremarctines in South America. Neues Jb. Geol. Paläontol. 250, 1–8. ( 10.1127/0077-7749/2008/0250-0001) [DOI] [Google Scholar]
- 32.Prevosti FJ, Vizcaíno SF. 2006. Paleoecology of the large carnivore guild from the late Pleistocene of Argentina. Acta Palaeontol. Pol. 51, 407–422. [Google Scholar]
- 33.Prevosti FJ, Forasiepi A, Zimicz N. 2013. The evolution of the Cenozoic terrestrial mammalian predator guild in South America: competition or replacement? J. Mamm. Evol. 20, 3–21. ( 10.1007/s10914-011-9175-9) [DOI] [Google Scholar]
- 34.de los Reyes M, Poiré D, Soibelzon LH, Zurita AE, Arrouy MJ. 2013. First evidence of scavenging of a glyptodont (Mammalia, Glyptodontidae) from the Pliocene of the pampean region (Argentina): taphonomic and paleoecological remarks. Palaeontol. Electron. 16, 1–13. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data from this study are available on GenBank (KU886001) and the Dryad Data Repository (http://dx.doi.org/10.5061/dryad.v7f30).